
Improvements in Maturity and Stability of 3D iPSC-Derived Hepatocyte-like Cell Cultures
 , , ,
, , ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Induced Pluripotent Stem Cell (iPSC) Reprogramming and Cell Culture
2.2. Differentiation of iPSCs into Hepatocyte-like Cells (HLCs)
2.3. Isolation and Culture of Human Adipose Tissue-Derived Stem/Stromal Cells
2.4. Isolation and Culture of Human Umbilical Vein Endothelial Cells (HUVECs)
2.5. Establishing Microvascular Network
2.6. iPSC-HLC Spheroid Formation and Culturing
2.7. Culturing Cells on a Microfluidic Chip
2.8. Culturing the 3D iPSC-HLC Spheroids in Inserts with Vasculature
2.9. iPSC-HLC + HUVEC Spheroids
2.10. iPSC-HLCs with HUVECs on Microfluidic Chips
2.11. Multilineage Cell Cultures
2.11.1. Medium Tests for Multilineage Spheroids
2.11.2. Further Multilineage Spheroid Experiments
2.11.3. Multilineage Chip Cultures
2.12. Assessing Biomaterial Integrity on the Chip Cultures
2.13. Assessing Cell Viability
2.13.1. Cell Viability in the Spheroid Cultures
2.13.2. Cell Viability in the Microfluidic Chip Cultures
2.14. RNA Extraction and Real-Time Quantitative PCR (qPCR)
2.14.1. Spheroids
2.14.2. Microfluidic Chips
2.15. Staining
2.15.1. Immunocytochemical (ICC) Analysis of 2D iPSC-HLC Cultures
2.15.2. Immunohistochemical (IHC) Analysis of Spheroid Sections
2.15.3. Hematoxylin-Eosin Staining of Spheroid Sections
2.15.4. Immunocytochemical Analysis of Cells Cultured on Microfluidic Chips
2.16. Media Analyses
2.16.1. Albumin
2.16.2. Urea
2.17. Statistical Methods
2.18. The Selective Plane Illumination Microscopy (SPIM) Imaging
2.19. The Optical Projection Tomography (OPT) and Electrical Impedance Tomography (EIT) Imaging
3. Results
3.1. The 3D iPSC-HLC Monoculture Spheroid and Microfluidic Chip Cultures
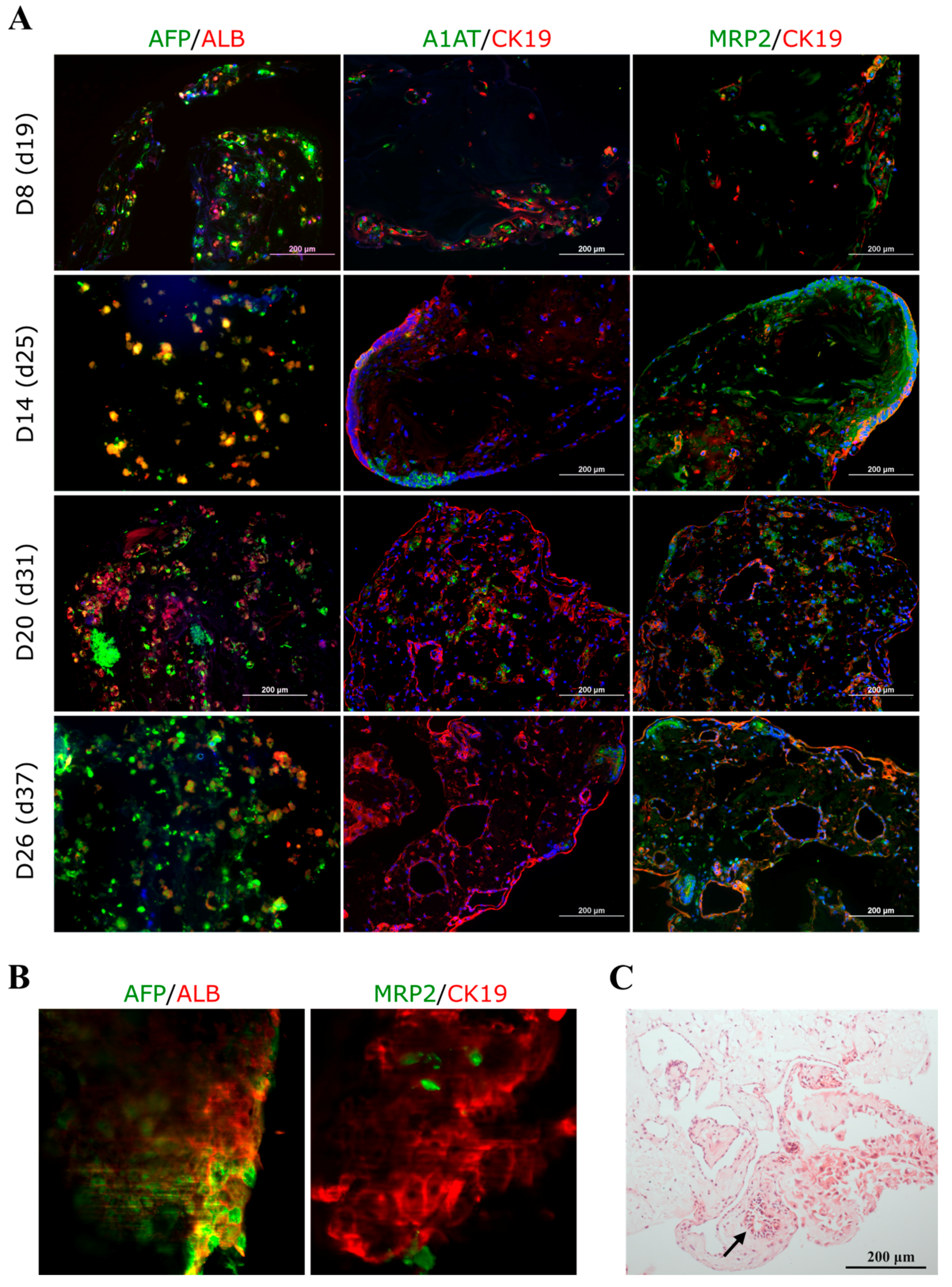
3.1.1. Mature Liver Marker Gene and Protein Expression Was Increased in the 3D iPSC-HLC Monoculture Spheroids during Culture
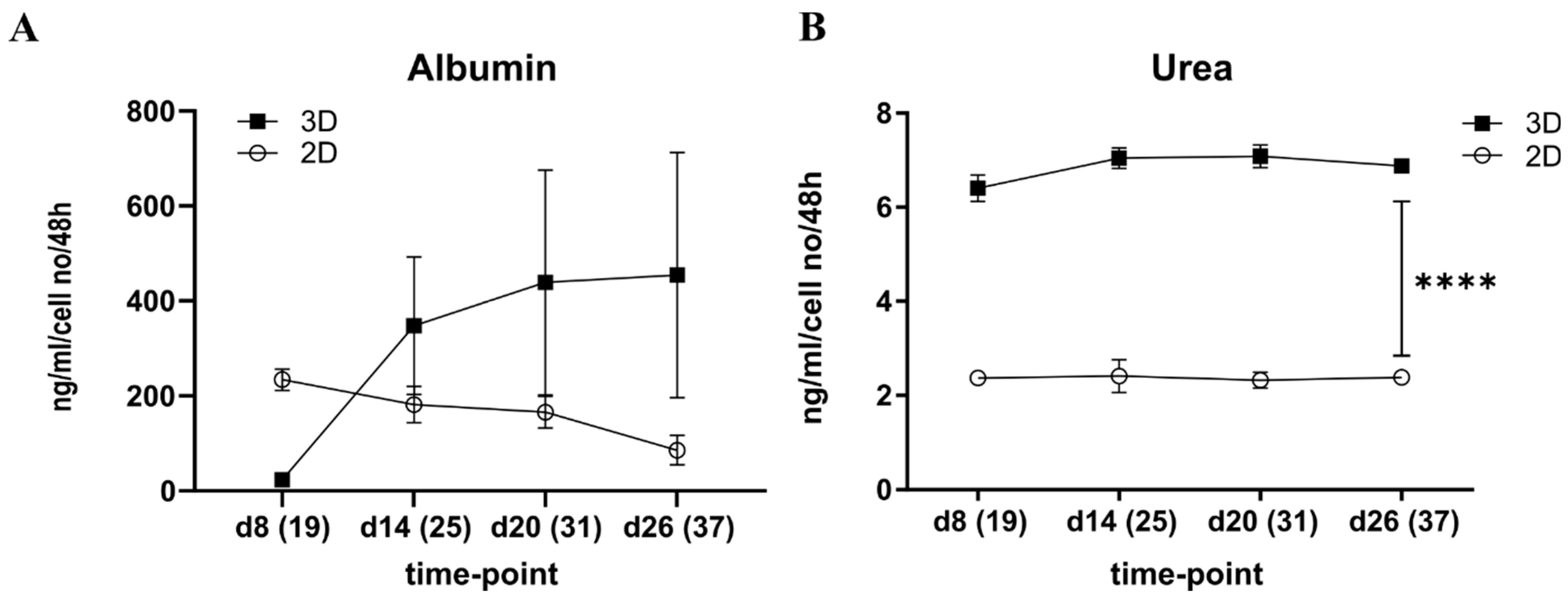
3.1.2. Albumin Production was Increased in the 3D Spheroids over Time Whereas Urea Stayed Constant
3.1.3. Biomaterial Testing for 3D on-Chip Hepatocyte Monocultures
3.2. The 3D iPSC-HLC Spheroids in Insert Culture with Vasculature
3.2.1. Gene Expression
3.2.2. Albumin and Urea Production
3.3. The 3D Co-Culture of iPSC-HLCs and HUVECs
3.3.1. Gene Expression in Co-Culture Spheroids
3.3.2. The OPT, EIT, and SPIM Imaging and Immunocytochemical Staining Reveal Decreasing Spheroid Size, Increasing Conductivity, and Persisting AFP Expression
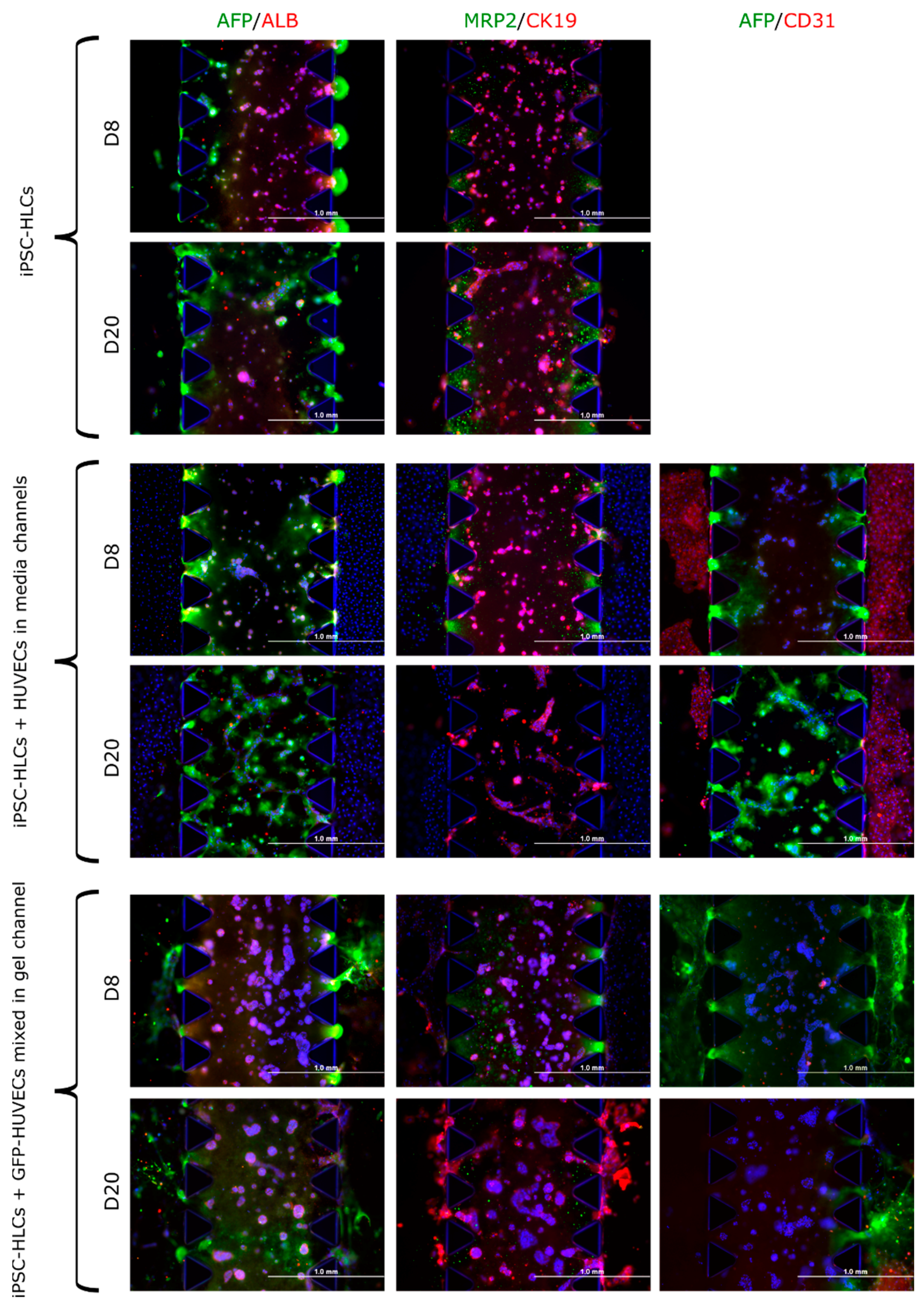
3.3.3. iPSC-HLCs in 3D Co-Culture with HUVECs on Microfluidic Chips
3.4. Multilineage Cultures of iPSC-HLCs, HUVECs and hASCs
3.4.1. Gene Expression in Multilineage Spheroids
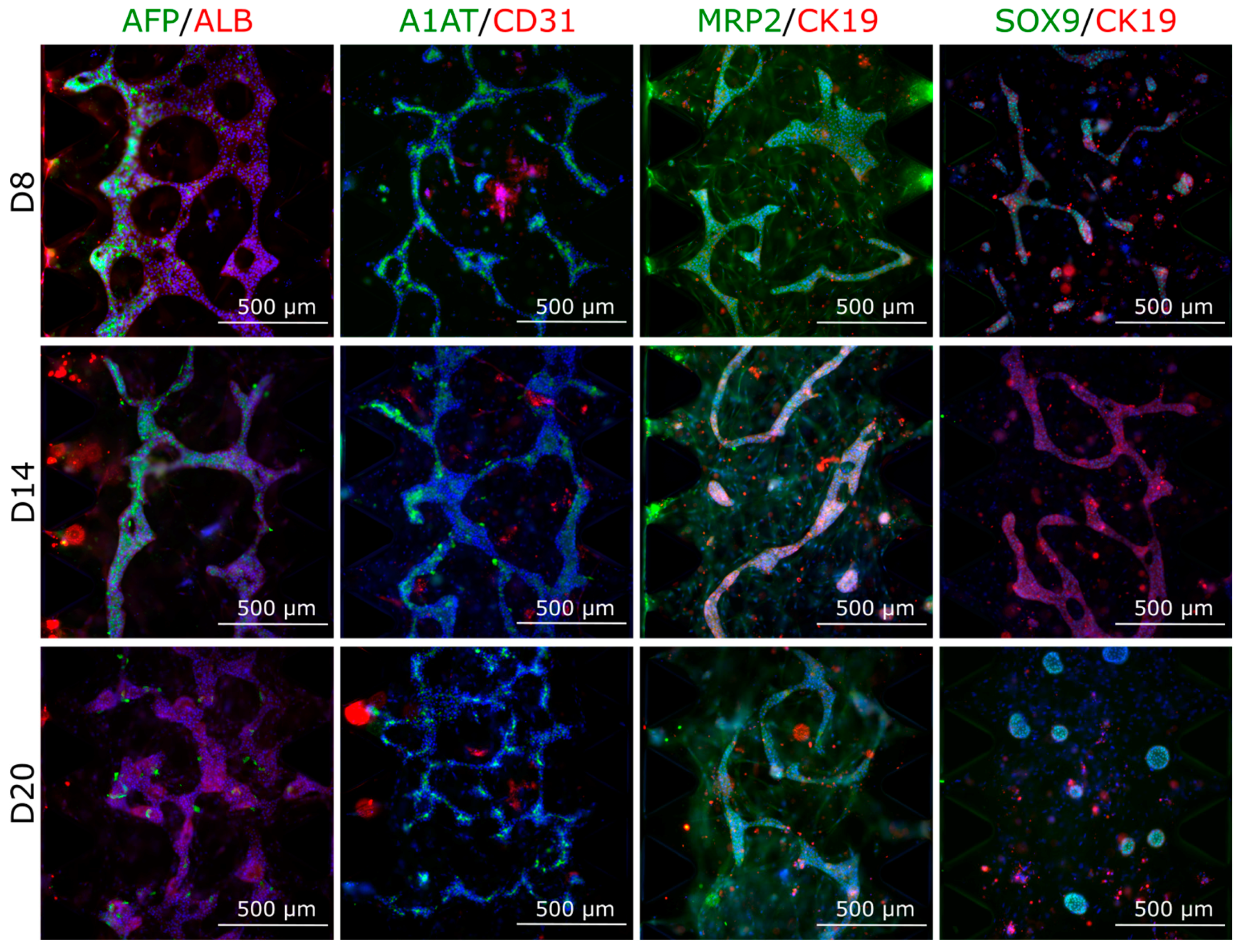
3.4.2. Protein Expression in Multilineage Spheroids
3.4.3. Multilineage Chip Cultures
3.4.4. Albumin Production Stayed Constant in the Multilineage Chip Cultures
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Heydari, Z.; Najimi, M.; Mirzaei, H.; Shpichka, A.; Ruoss, M.; Farzaneh, Z.; Montazeri, L.; Piryaei, A.; Timashev, P.; Gramignoli, R.; et al. Tissue Engineering in Liver Regenerative Medicine: Insights into Novel Translational Technologies. Cells 2020, 9, 304. [Google Scholar] [CrossRef] [PubMed]
- Godoy, P.; Hewitt, N.J.; Albrecht, U.; Andersen, M.E.; Ansari, N.; Bhattacharya, S.; Bode, J.G.; Bolleyn, J.; Borner, C.; Böttger, J.; et al. Recent Advances in 2D and 3D in Vitro Systems Using Primary Hepatocytes, Alternative Hepatocyte Sources and Non-Parenchymal Liver Cells and Their Use in Investigating Mechanisms of Hepatotoxicity, Cell Signaling and ADME. Arch. Toxicol. 2013, 87, 1315–1530. [Google Scholar] [CrossRef] [PubMed]
- Kang, H.K.; Sarsenova, M.; Kim, D.-H.; Kim, M.S.; Lee, J.Y.; Sung, E.-A.; Kook, M.G.; Kim, N.G.; Choi, S.W.; Ogay, V.; et al. Establishing a 3D In Vitro Hepatic Model Mimicking Physiologically Relevant to In Vivo State. Cells 2021, 10, 1268. [Google Scholar] [CrossRef] [PubMed]
- Lee, G.; Kim, H.; Park, J.Y.; Kim, G.; Han, J.; Chung, S.; Yang, J.H.; Jeon, J.S.; Woo, D.-H.; Han, C.; et al. Generation of Uniform Liver Spheroids from Human Pluripotent Stem Cells for Imaging-Based Drug Toxicity Analysis. Biomaterials 2021, 269, 120529. [Google Scholar] [CrossRef] [PubMed]
- Danoy, M.; Tauran, Y.; Poulain, S.; Jellali, R.; Bruce, J.; Leduc, M.; Le Gall, M.; Koui, Y.; Arakawa, H.; Gilard, F.; et al. Investigation of the Hepatic Development in the Coculture of HiPSCs-Derived LSECs and HLCs in a Fluidic Microenvironment. APL Bioeng. 2021, 5, 26104. [Google Scholar] [CrossRef]
- Heslop, J.A.; Rowe, C.; Walsh, J.; Sison-Young, R.; Jenkins, R.; Kamalian, L.; Kia, R.; Hay, D.; Jones, R.P.; Malik, H.Z.; et al. Mechanistic Evaluation of Primary Human Hepatocyte Culture Using Global Proteomic Analysis Reveals a Selective Dedifferentiation Profile. Arch. Toxicol. 2017, 91, 439–452. [Google Scholar] [CrossRef]
- Kiamehr, M.; Heiskanen, L.; Laufer, T.; Düsterloh, A.; Kahraman, M.; Käkelä, R.; Laaksonen, R.; Aalto-Setälä, K. Dedifferentiation of Primary Hepatocytes Is Accompanied with Reorganization of Lipid Metabolism Indicated by Altered Molecular Lipid and MiRNA Profiles. Int. J. Mol. Sci. 2019, 20, 2910. [Google Scholar] [CrossRef]
- Bonanini, F.; Kurek, D.; Previdi, S.; Nicolas, A.; Hendriks, D.; de Ruiter, S.; Meyer, M.; Clapés Cabrer, M.; Dinkelberg, R.; García, S.B.; et al. In Vitro Grafting of Hepatic Spheroids and Organoids on a Microfluidic Vascular Bed. Angiogenesis 2022, 25, 455. [Google Scholar] [CrossRef]
- Serras, A.S.; Rodrigues, J.S.; Cipriano, M.; Rodrigues, A.V.; Oliveira, N.G.; Miranda, J.P. A Critical Perspective on 3D Liver Models for Drug Metabolism and Toxicology Studies. Front. Cell Dev. Biol. 2021, 9, 203. [Google Scholar] [CrossRef]
- Matsuda, K.; Falkenberg, K.J.; Woods, A.A.; Choi, Y.S.; Morrison, W.A.; Dilley, R.J. Adipose-Derived Stem Cells Promote Angiogenesis and Tissue Formation for In Vivo Tissue Engineering. Tissue Eng. Part A 2013, 19, 1327. [Google Scholar] [CrossRef]
- Wang, G.; Zheng, Y.; Wang, Y.; Cai, Z.; Liao, N.; Liu, J.; Zhang, W. Co-Culture System of Hepatocytes and Endothelial Cells: Two in Vitro Approaches for Enhancing Liver-Specific Functions of Hepatocytes. Cytotechnology 2018, 70, 1279–1290. [Google Scholar] [CrossRef] [PubMed]
- Mykuliak, A.; Yrjänäinen, A.; Mäki, A.-J.; Gebraad, A.; Lampela, E.; Kääriäinen, M.; Pakarinen, T.-K.; Kallio, P.; Miettinen, S.; Vuorenpää, H. Vasculogenic Potency of Bone Marrow- and Adipose Tissue-Derived Mesenchymal Stem/Stromal Cells Results in Differing Vascular Network Phenotypes in a Microfluidic Chip. Front. Bioeng. Biotechnol. 2022, 10, 764237. [Google Scholar] [CrossRef]
- Freyer, N.; Greuel, S.; Knöspel, F.; Strahl, N.; Amini, L.; Jacobs, F.; Monshouwer, M.; Zeilinger, K. Effects of Co-Culture Media on Hepatic Differentiation of HiPSC with or without HUVEC Co-Culture. Int. J. Mol. Sci. 2017, 18, 1724. [Google Scholar] [CrossRef] [PubMed]
- Manzini, S.; Viiri, L.E.; Marttila, S.; Aalto-Setälä, K. A Comparative View on Easy to Deploy Non-Integrating Methods for Patient-Specific IPSC Production. Stem Cell Rev. 2015, 11, 900–908. [Google Scholar] [CrossRef] [PubMed]
- Kiamehr, M.; Viiri, L.E.; Vihervaara, T.; Koistinen, K.M.; Hilvo, M.; Ekroos, K.; Käkelä, R.; Aalto-Setälä, K. Lipidomic Profiling of Patient-Specific IPSC-Derived Hepatocyte-like Cells. Dis. Model. Mech. 2017, 10, 1141–1153. [Google Scholar] [CrossRef] [PubMed]
- Kajiwara, M.; Aoi, T.; Okita, K.; Takahashi, R.; Inoue, H.; Takayama, N.; Endo, H.; Eto, K.; Toguchida, J.; Uemoto, S.; et al. Donor-Dependent Variations in Hepatic Differentiation from Human-Induced Pluripotent Stem Cells. Proc. Natl. Acad. Sci. USA 2012, 109, 12538–12543. [Google Scholar] [CrossRef]
- Kiamehr, M.; Alexanova, A.; Viiri, L.E.; Heiskanen, L.; Vihervaara, T.; Kauhanen, D.; Ekroos, K.; Laaksonen, R.; Käkelä, R.; Aalto-Setälä, K. HiPSC-Derived Hepatocytes Closely Mimic the Lipid Profile of Primary Hepatocytes: A Future Personalised Cell Model for Studying the Lipid Metabolism of the Liver. J. Cell. Physiol. 2019, 234, 3744–3761. [Google Scholar] [CrossRef]
- Kyllönen, L.; Haimi, S.; Mannerström, B.; Huhtala, H.; Rajala, K.M.; Skottman, H.; Sándor, G.K.; Miettinen, S. Effects of Different Serum Conditions on Osteogenic Differentiation of Human Adipose Stem Cells in Vitro. Stem Cell Res. Ther. 2013, 4, 17. [Google Scholar] [CrossRef]
- Vuornos, K.; Ojansivu, M.; Koivisto, J.T.; Häkkänen, H.; Belay, B.; Montonen, T.; Huhtala, H.; Kääriäinen, M.; Hupa, L.; Kellomäki, M.; et al. Bioactive Glass Ions Induce Efficient Osteogenic Differentiation of Human Adipose Stem Cells Encapsulated in Gellan Gum and Collagen Type I Hydrogels. Mater. Sci. Eng. C 2019, 99, 905–918. [Google Scholar] [CrossRef]
- Jaffe, E.A.; Nachman, R.L.; Becker, C.G.; Minick, C.R. Culture of Human Endothelial Cells Derived from Umbilical Veins. Identification by morphologic and immunologic criteria. J. Clin. Investig. 1973, 52, 2745. [Google Scholar] [CrossRef]
- Sarkanen, J.-R.; Vuorenpää, H.; Huttala, O.; Mannerström, B.; Kuokkanen, H.; Miettinen, S.; Heinonen, T.; Ylikomi, T. Adipose Stromal Cell Tubule Network Model Provides a Versatile Tool for Vascular Research and Tissue Engineering. Cells Tissues Organs 2012, 196, 385–397. [Google Scholar] [CrossRef]
- Kiamehr, M.; Verfaillie, C. Spheroidal Self-Assembled Peptide Hydrogels Comprising Cells. WO 2022079272-A1, 21 April 2022. [Google Scholar]
- Livak, K.J.; Schmittgen, T.D. Analysis of Relative Gene Expression Data Using Real-Time Quantitative PCR and the 2−ΔΔCT Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]
- Gualda, E.J.; Vale, T.; Almada, P.; Feijó, J.A.; Martins, G.G.; Moreno, N. OpenSpinMicroscopy: An Open-Source Integrated Microscopy Platform. Nat. Methods 2013, 10, 599–600. [Google Scholar] [CrossRef]
- Lehti-Polojärvi, M.; Räsänen, M.J.; Viiri, L.E.; Vuorenpää, H.; Miettinen, S.; Seppänen, A.; Hyttinen, J. Retrieval of the Conductivity Spectrum of Tissues in Vitro with Novel Multimodal Tomography. Phys. Med. Biol. 2021, 66, 205016. [Google Scholar] [CrossRef] [PubMed]
- Gupta, R.; Schrooders, Y.; Hauser, D.; van Herwijnen, M.; Albrecht, W.; ter Braak, B.; Brecklinghaus, T.; Castell, J.V.; Elenschneider, L.; Escher, S.; et al. Comparing in Vitro Human Liver Models to in Vivo Human Liver Using RNA-Seq. Arch. Toxicol. 2021, 95, 573–589. [Google Scholar] [CrossRef] [PubMed]
- Lu, C.; Li, A.P.; Lu, C.; Li, A.P. Species Comparison in P450 Induction: Effects of Dexamethasone, Omeprazole, and Rifampin on P450 Isoforms 1A and 3A in Primary Cultured Hepatocytes from Man, Sprague-Dawley Rat, Minipig, and Beagle Dog. Chem. Biol. Interact. 2001, 134, 271–281. [Google Scholar] [CrossRef] [PubMed]
- Poorna, M.R.; Jayakumar, R.; Chen, J.P.; Mony, U. Hydrogels: A Potential Platform for Induced Pluripotent Stem Cell Culture and Differentiation. Colloids Surf. B Biointerfaces 2021, 207, 111991. [Google Scholar] [CrossRef] [PubMed]
- Bhattacharya, M.; Malinen, M.M.; Lauren, P.; Lou, Y.R.; Kuisma, S.W.; Kanninen, L.; Lille, M.; Corlu, A.; Guguen-Guillouzo, C.; Ikkala, O.; et al. Nanofibrillar Cellulose Hydrogel Promotes Three-Dimensional Liver Cell Culture. J. Control. Release 2012, 164, 291–298. [Google Scholar] [CrossRef]
- Roth, A.D.; Lama, P.; Dunn, S.; Hong, S.; Lee, M.Y. Polymer Coating on a Micropillar Chip for Robust Attachment of PuraMatrix Peptide Hydrogel for 3D Hepatic Cell Culture. Mater. Sci. Eng. C 2018, 90, 634–644. [Google Scholar] [CrossRef]
- Ardalani, H.; Sengupta, S.; Harms, V.; Vickerman, V.; Thomson, J.A.; Murphy, W.L. 3-D Culture and Endothelial Cells Improve Maturity of Human Pluripotent Stem Cell-Derived Hepatocytes. Acta Biomater. 2019, 95, 371–381. [Google Scholar] [CrossRef]
- Gieseck III, R.L.; Hannan, N.R.F.; Bort, R.; Hanley, N.A.; Drake, R.A.L.; Cameron, G.W.W.; Wynn, T.A.; Vallier, L.; Elaut, G.; Henkens, T.; et al. Maturation of Induced Pluripotent Stem Cell Derived Hepatocytes by 3D-Culture. PLoS ONE 2014, 9, e86372. [Google Scholar] [CrossRef]
- Luo, Y.; Lou, C.; Zhang, S.; Zhu, Z.; Xing, Q.; Wang, P.; Liu, T.; Liu, H.; Li, C.; Shi, W.; et al. Three-Dimensional Hydrogel Culture Conditions Promote the Differentiation of Human Induced Pluripotent Stem Cells into Hepatocytes. Cytotherapy 2018, 20, 95–107. [Google Scholar] [CrossRef] [PubMed]
- Asai, A.; Aihara, E.; Watson, C.; Mourya, R.; Mizuochi, T.; Shivakumar, P.; Phelan, K.; Mayhew, C.; Helmrath, M.; Takebe, T.; et al. Paracrine Signals Regulate Human Liver Organoid Maturation from Induced Pluripotent Stem Cells. Dev. Camb. 2017, 144, 1056–1064. [Google Scholar] [CrossRef]
- Pill, K.; Melke, J.; Mühleder, S.; Pultar, M.; Rohringer, S.; Priglinger, E.; Redl, H.R.; Hofmann, S.; Holnthoner, W. Microvascular Networks from Endothelial Cells and Mesenchymal Stromal Cells from Adipose Tissue and Bone Marrow: A Comparison. Front. Bioeng. Biotechnol. 2018, 6, 156. [Google Scholar] [CrossRef] [PubMed]
- Pettinato, G.; Lehoux, S.; Ramanathan, R.; Salem, M.M.; He, L.-X.; Muse, O.; Flaumenhaft, R.; Thompson, M.T.; Rouse, E.A.; Cummings, R.D.; et al. Generation of Fully Functional Hepatocyte-like Organoids from Human Induced Pluripotent Stem Cells Mixed with Endothelial Cells. Sci. Rep. 2019, 9, 8920. [Google Scholar] [CrossRef]
- Lin, Y.; Zhang, F.; Zhang, L.; Chen, L.; Zheng, S. Characteristics of SOX9-Positive Progenitor-like Cells during Cholestatic Liver Regeneration in Biliary Atresia. Stem Cell Res. Ther. 2022, 13, 114. [Google Scholar] [CrossRef]
- van Wenum, M.; Adam, A.A.A.; van der Mark, V.A.; Chang, J.C.; Wildenberg, M.E.; Hendriks, E.J.; Jongejan, A.; Moerland, P.D.; van Gulik, T.M.; Oude Elferink, R.P.; et al. Oxygen Drives Hepatocyte Differentiation and Phenotype Stability in Liver Cell Lines. J. Cell Commun. Signal. 2018, 12, 575–588. [Google Scholar] [CrossRef]
- Jung, H.R.; Kang, H.M.; Ryu, J.W.; Kim, D.S.; Noh, K.H.; Kim, E.S.; Lee, H.J.; Chung, K.S.; Cho, H.S.; Kim, N.S.; et al. Cell Spheroids with Enhanced Aggressiveness to Mimic Human Liver Cancer In Vitro and In Vivo. Sci. Rep. 2017, 7, 10499. [Google Scholar] [CrossRef]
- Takebe, T.; Sekine, K.; Enomura, M.; Koike, H.; Kimura, M.; Ogaeri, T.; Zhang, R.-R.; Ueno, Y.; Zheng, Y.-W.; Koike, N.; et al. Vascularized and Functional Human Liver from an IPSC-Derived Organ Bud Transplant. Nature 2013, 499, 481–484. [Google Scholar] [CrossRef]
- Takebe, T.; Zhang, R.-R.; Koike, H.; Kimura, M.; Yoshizawa, E.; Enomura, M.; Koike, N.; Sekine, K.; Taniguchi, H. Generation of a Vascularized and Functional Human Liver from an IPSC-Derived Organ Bud Transplant. Nat. Protoc. 2014, 9, 396–409. [Google Scholar] [CrossRef]
- Kratochwil, N.A.; Meille, C.; Fowler, S.; Klammers, F.; Ekiciler, A.; Molitor, B.; Simon, S.; Walter, I.; McGinnis, C.; Walther, J.; et al. Metabolic Profiling of Human Long-Term Liver Models and Hepatic Clearance Predictions from In Vitro Data Using Nonlinear Mixed-Effects Modeling. AAPS J. 2017, 19, 534–550. [Google Scholar] [CrossRef] [PubMed]
- Blau, B.J.; Miki, T. The Role of Cellular Interactions in the Induction of Hepatocyte Polarity and Functional Maturation in Stem Cell-Derived Hepatic Cells. Differentiation 2019, 106, 42–48. [Google Scholar] [CrossRef] [PubMed]
- Ma, X.; Qu, X.; Zhu, W.; Li, Y.-S.; Yuan, S.; Zhang, H.; Liu, J.; Wang, P.; Lai, C.S.E.; Zanella, F.; et al. Deterministically Patterned Biomimetic Human IPSC-Derived Hepatic Model via Rapid 3D Bioprinting. Proc. Natl. Acad. Sci. USA 2016, 113, 2206–2211. [Google Scholar] [CrossRef] [PubMed]
- Koui, Y.; Kido, T.; Ito, T.; Oyama, H.; Chen, S.-W.; Katou, Y.; Shirahige, K.; Miyajima, A. An In Vitro Human Liver Model by IPSC-Derived Parenchymal and Non-Parenchymal Cells. Stem Cell Rep. 2017, 9, 490–498. [Google Scholar] [CrossRef] [PubMed]
- Cortesi, M.; Giordano, E. Non-Destructive Monitoring of 3D Cell Cultures: New Technologies and Applications. PeerJ 2022, 10, e13338. [Google Scholar] [CrossRef]
- James-Bhasin, M.; Siegel, P.M.; Nazhat, S.N. A Three-Dimensional Dense Collagen Hydrogel to Model Cancer Cell/Osteoblast Interactions. J. Funct. Biomater. 2018, 9, 72. [Google Scholar] [CrossRef]
- Pan, Y.; Jiang, D.; Gu, C.; Qiu, Y.; Wan, H.; Wang, P. 3D Microgroove Electrical Impedance Sensing to Examine 3D Cell Cultures for Antineoplastic Drug Assessment. Microsyst. Nanoeng. 2020, 6, 23. [Google Scholar] [CrossRef]
- Goodenough, D.A.; Paul, D.L. Gap Junctions. Cold Spring Harb. Perspect. Biol. 2009, 1, a002576. [Google Scholar] [CrossRef]
- Van Rijen, H.V.M.; Van Kempen, M.J.A.; Analbers, L.J.S.; Rook, M.B.; Van Ginneken, C.G.; Gros, D.; Jongsmal, H.J.; Rijen, V.; van Kempen, M.J.A.; Analbers, U.J.S.; et al. Gap Junctions in Human Umbilical Cord Endothelial Cells Contain Multiple Connexins. Am. J. Physiol.-Cell Physiol. 1997, 272, C117–C130. [Google Scholar] [CrossRef]
- Willebrords, J.; Crespo Yanguas, S.; Maes, M.; Decrock, E.; Wang, N.; Leybaert, L.; da Silva, T.C.; Veloso Alves Pereira, I.; Jaeschke, H.; Cogliati, B.; et al. Structure, Regulation and Function of Gap Junctions in Liver. Cell Commun. Adhes. 2016, 22, 29–37. [Google Scholar] [CrossRef]
- Halonen, S.; Ovissi, A.; Boyd, S.; Kari, J.; Kronström, K.; Kosunen, J.; Laurén, H.; Numminen, K.; Sievänen, H.; Hyttinen, J. Human In Vivo Liver and Tumor Bioimpedance Measured with Biopsy Needle. Physiol. Meas. 2022, 43, 015006. [Google Scholar] [CrossRef] [PubMed]
- Ogawa, R.; Hallas-Potts, A.; Wu, H.; Jia, J.; Bagnaninchi, P.O. Measuring 3D Cell Culture Viability in Multiple 3D Printed Scaffolds Within a Single Miniature Electrical Impedance Tomography Sensor. Adv. Eng. Mater. 2021, 23, 2100338. [Google Scholar] [CrossRef]
- Ahmed, T.A.E.; Griffith, M.; Hincke, M. Characterization and Inhibition of Fibrin Hydrogel–Degrading Enzymes during Development of Tissue Engineering Scaffolds. Tissue Eng. 2007, 13, 1469–1477. [Google Scholar] [CrossRef]
- Banihashemi, M.; Mohkam, M.; Safari, A.; Nezafat, N.; Negahdaripour, M.; Mohammadi, F.; Kianpour, S.; Ghasemi, Y. Optimization of Three Dimensional Culturing of the HepG2 Cell Line in Fibrin Scaffold. Hepat. Mon. 2015, 15, 22731. [Google Scholar] [CrossRef]
- Chhabra, A.; Song, H.H.G.; Grzelak, K.A.; Polacheck, W.J.; Fleming, H.E.; Chen, C.S.; Bhatia, S.N. A Vascularized Model of the Human Liver Mimics Regenerative Responses. Proc. Natl. Acad. Sci. USA 2022, 119, e2115867119. [Google Scholar] [CrossRef]
- Danoy, M.; Bernier, M.L.; Kimura, K.; Poulain, S.; Kato, S.; Mori, D.; Kido, T.; Plessy, C.; Kusuhara, H.; Miyajima, A.; et al. Optimized Protocol for the Hepatic Differentiation of Induced Pluripotent Stem Cells in a Fluidic Microenvironment. Biotechnol. Bioeng. 2019, 116, 1762–1776. [Google Scholar] [CrossRef] [PubMed]
- Leclerc, E.; Kimura, K.; Shinohara, M.; Danoy, M.; Le Gall, M.; Kido, T.; Miyajima, A.; Fujii, T.; Sakai, Y. Comparison of the Transcriptomic Profile of Hepatic Human Induced Pluripotent Stem like Cells Cultured in Plates and in a 3D Microscale Dynamic Environment. Genomics 2017, 109, 16–26. [Google Scholar] [CrossRef]
- Camp, J.G.; Sekine, K.; Gerber, T.; Loeffler-Wirth, H.; Binder, H.; Gac, M.; Kanton, S.; Kageyama, J.; Damm, G.; Seehofer, D.; et al. Multilineage Communication Regulates Human Liver Bud Development from Pluripotency. Nature 2017, 546, 533. [Google Scholar] [CrossRef]
- Takagi, C.; Yagi, H.; Hieda, M.; Tajima, K.; Hibi, T.; Abe, Y.; Kitago, M.; Shinoda, M.; Itano, O.; Kitagawa, Y. Mesenchymal Stem Cells Contribute to Hepatic Maturation of Human Induced Pluripotent Stem Cells. Eur. Surg. Res. 2017, 58, 27–39. [Google Scholar] [CrossRef]
- Raasch, M.; Fritsche, E.; Kurtz, A.; Bauer, M.; Mosig, A.S. Microphysiological Systems Meet HiPSC Technology—New Tools for Disease Modeling of Liver Infections in Basic Research and Drug Development. Adv. Drug Deliv. Rev. 2018, 140, 51–67. [Google Scholar] [CrossRef]












{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Exp | Cell Type | No of Chips | Seeding Day | Biomaterial | Time Points | Medium |
|---|---|---|---|---|---|---|
| 1 | iPSC-HLC | 5 | d10 | Geltrex (3:2) | d8, d20 | HCM |
| 2 | iPSC-HLC | 4 | d10 | Geltrex (2:1) | d8, d20 | HCM |
| 3 | iPSC-HLC | 5 | d10 | fibrin | d8 (term) | HCM |
| 4 | iPSC-HLC | 9 | d11 | fibrin | d8, d20 | HCM+apr |
| 5 | iPSC-HLC | 6 | d11 | col I | d8, d20 | HCM |
| 6 | iPSC-HLC | 10 | d11 | fibrin-col I | d8, d20 | HCM+apr |
| 7 | iPSC-HLC+ HUVECs in media channels | 2 | d10 | Geltrex 3:2 | d8, d20 | HCM:EGM |
| 8 | iPSC-HLC+ HUVECs in media channels | 2 | d10 | Geltrex 2:1 | d8, d20 | HCM:EGM |
| 9 | iPSC-HLC + GFP- HUVECs mixed in gel channel | 6 | d10 | Geltrex 2:1 | d8, d20 | HCM:EGM |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Suominen, S.; Hyypijev, T.; Venäläinen, M.; Yrjänäinen, A.; Vuorenpää, H.; Lehti-Polojärvi, M.; Räsänen, M.; Seppänen, A.; Hyttinen, J.; Miettinen, S.; et al. Improvements in Maturity and Stability of 3D iPSC-Derived Hepatocyte-like Cell Cultures. Cells 2023, 12, 2368. https://doi.org/10.3390/cells12192368
Suominen S, Hyypijev T, Venäläinen M, Yrjänäinen A, Vuorenpää H, Lehti-Polojärvi M, Räsänen M, Seppänen A, Hyttinen J, Miettinen S, et al. Improvements in Maturity and Stability of 3D iPSC-Derived Hepatocyte-like Cell Cultures. Cells. 2023; 12(19):2368. https://doi.org/10.3390/cells12192368
Chicago/Turabian StyleSuominen, Siiri, Tinja Hyypijev, Mari Venäläinen, Alma Yrjänäinen, Hanna Vuorenpää, Mari Lehti-Polojärvi, Mikko Räsänen, Aku Seppänen, Jari Hyttinen, Susanna Miettinen, and et al. 2023. "Improvements in Maturity and Stability of 3D iPSC-Derived Hepatocyte-like Cell Cultures" Cells 12, no. 19: 2368. https://doi.org/10.3390/cells12192368