
Adverse Crosstalk between Extracellular Matrix Remodeling and Ferroptosis in Basal Breast Cancer
 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Determination of Ferroptosis in Breast Cancer-Related Genes
2.2. Transcriptome Cohort of Breast Cancer for Patients Treated with Anthracyclin and Taxanes
2.3. Multi-Omics Validation Breast Cancer Cohort METABRIC
2.4. Transcriptome Dataset Testing Effect of Ferroptosis Inducers on Triple-Negative Breast Cancer Cell Models
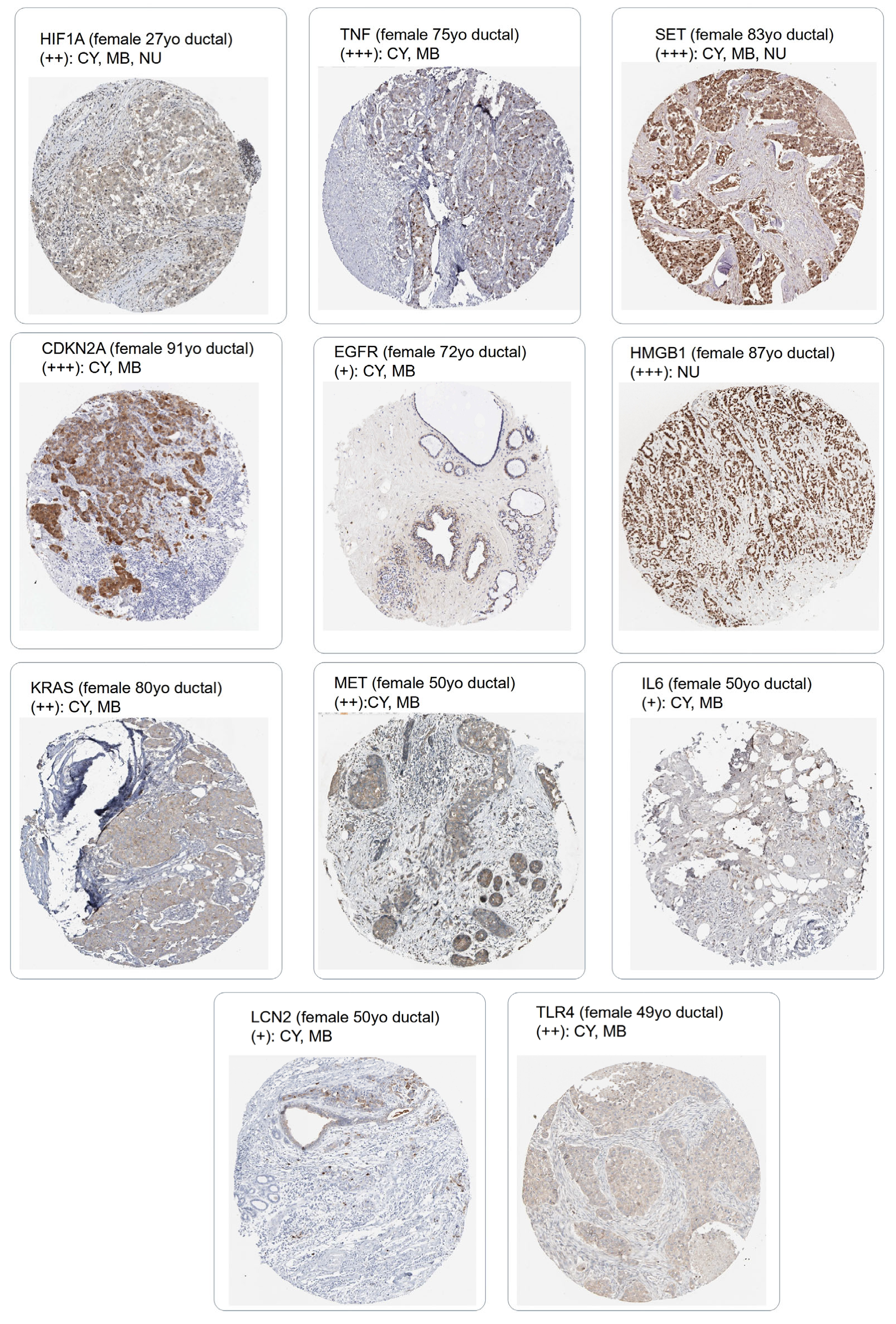
2.5. Immunohistochemistry Protein Level Expression of Eleven Markers
2.6. Gene Expression Analyses and Association to the Breast Cancer Prognosis
3. Results
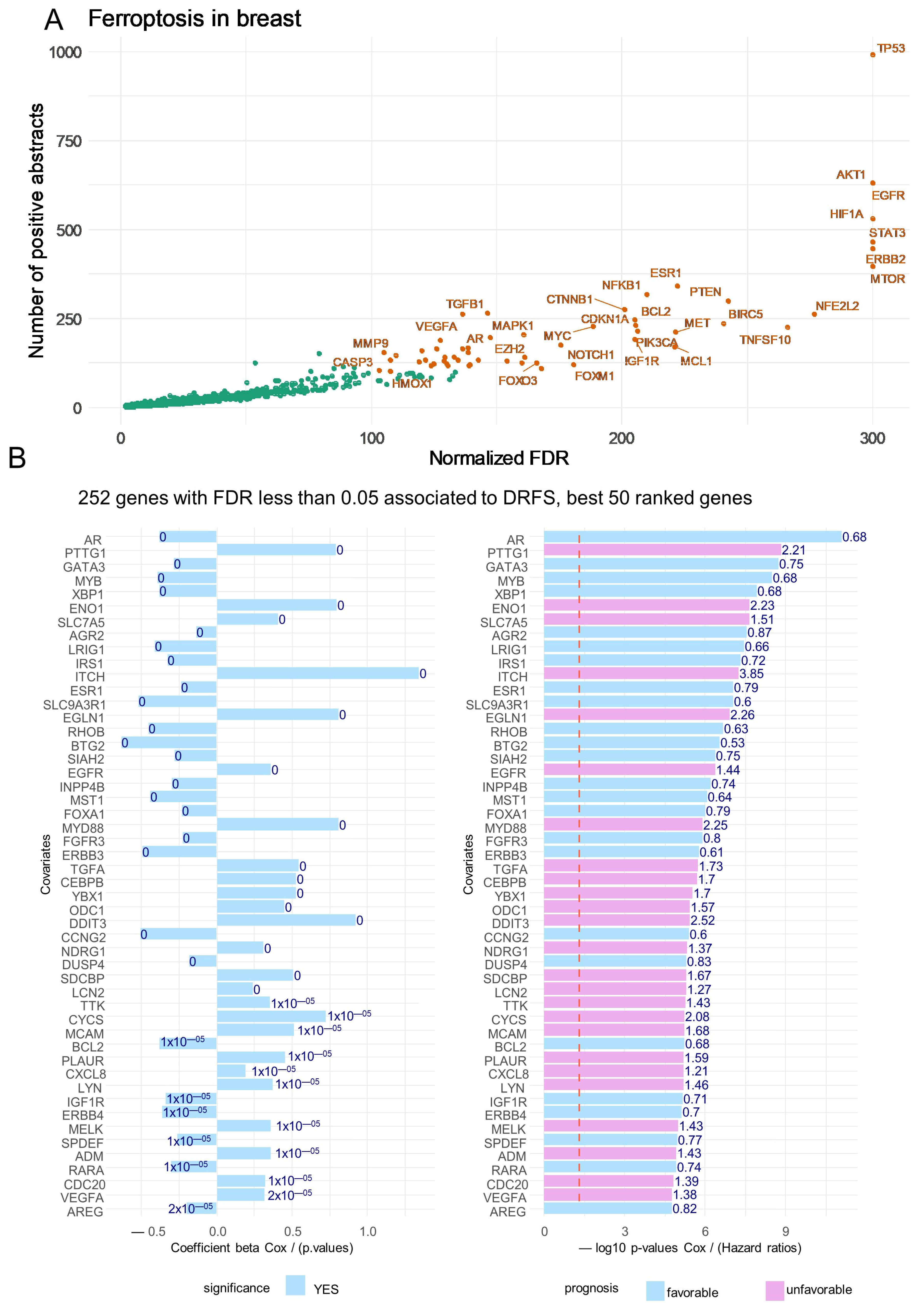
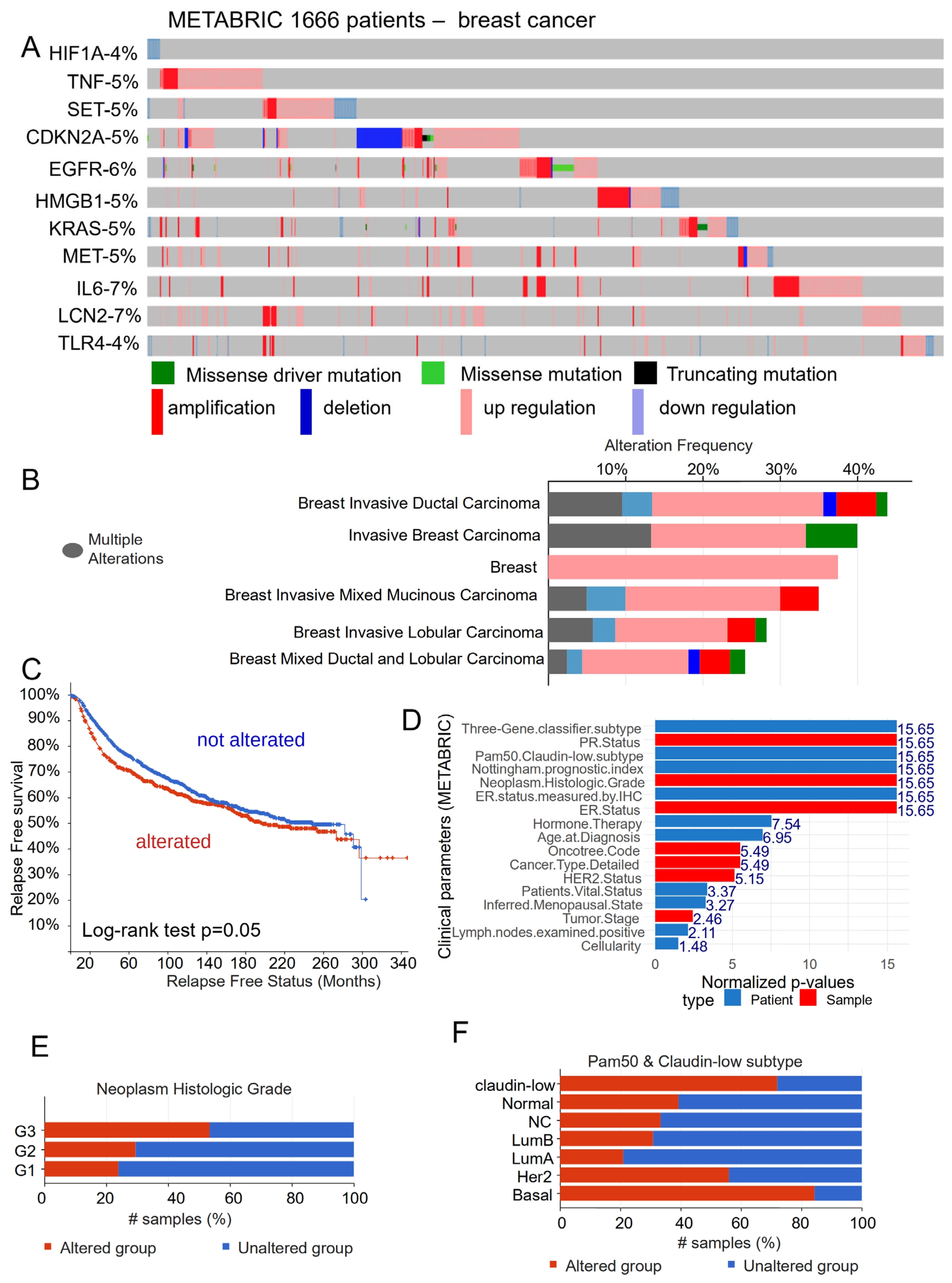
3.1. Ferroptosis Gene Expression Associated with the Prognosis of Patients with Breast Cancer
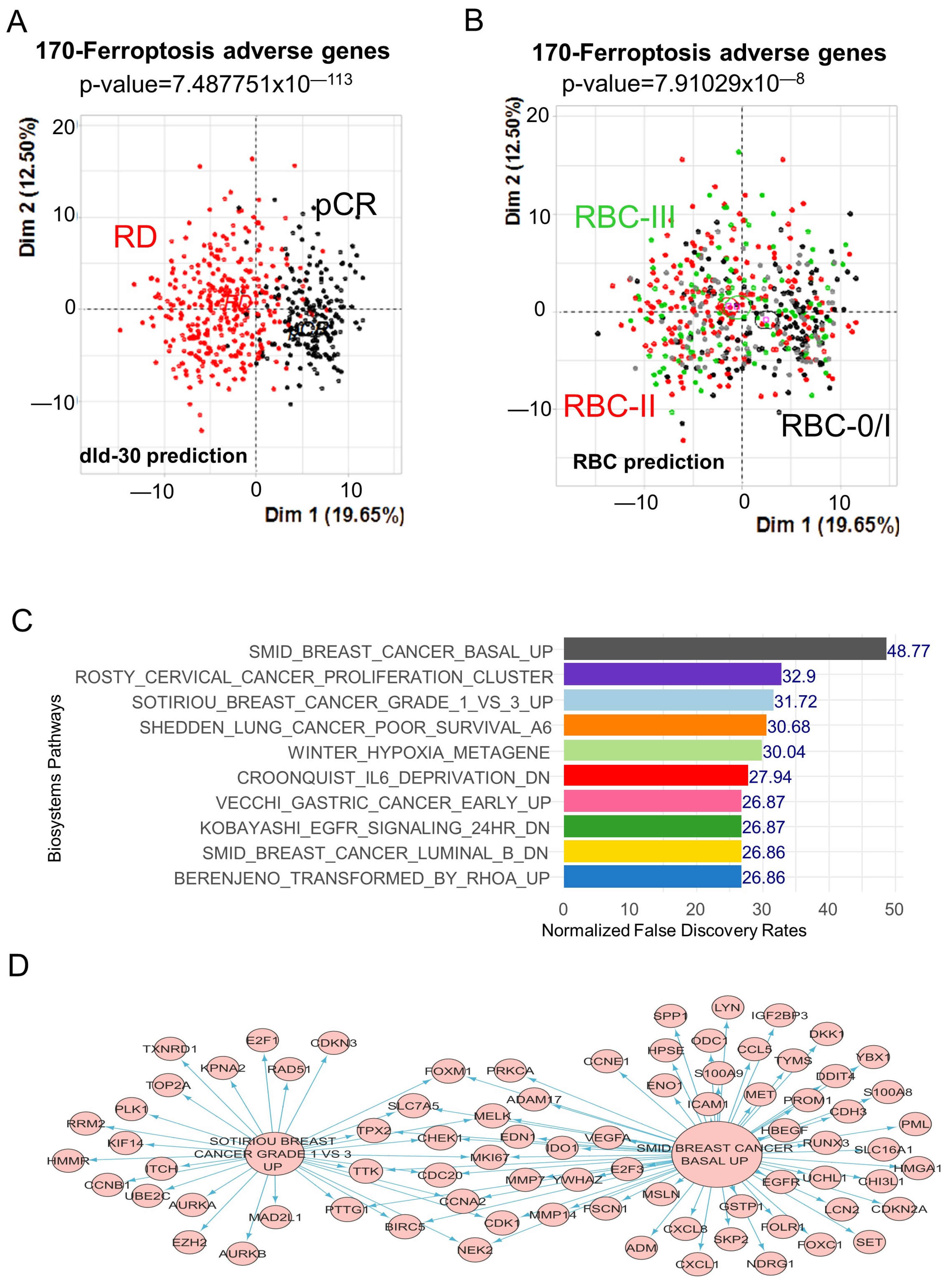
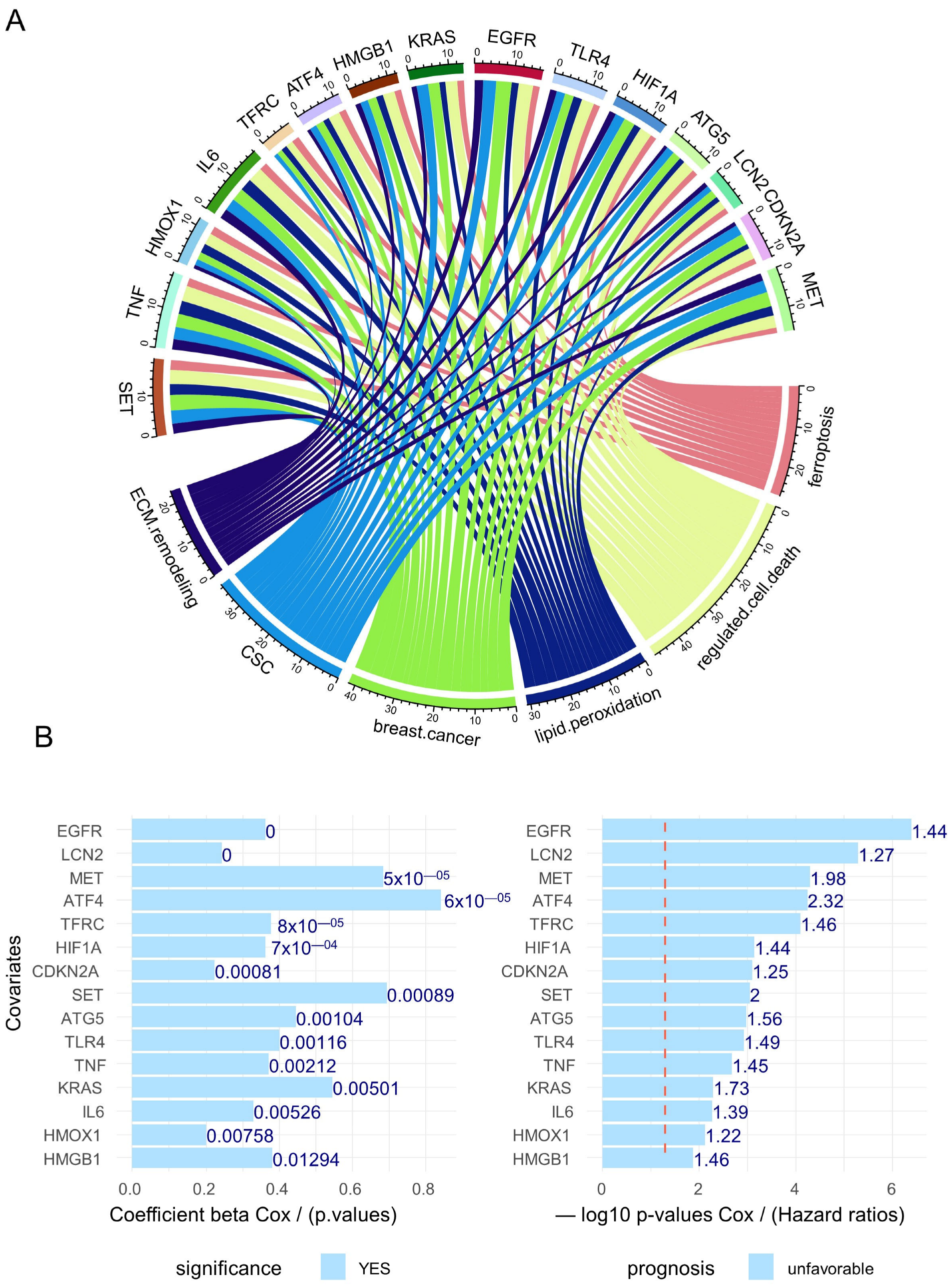
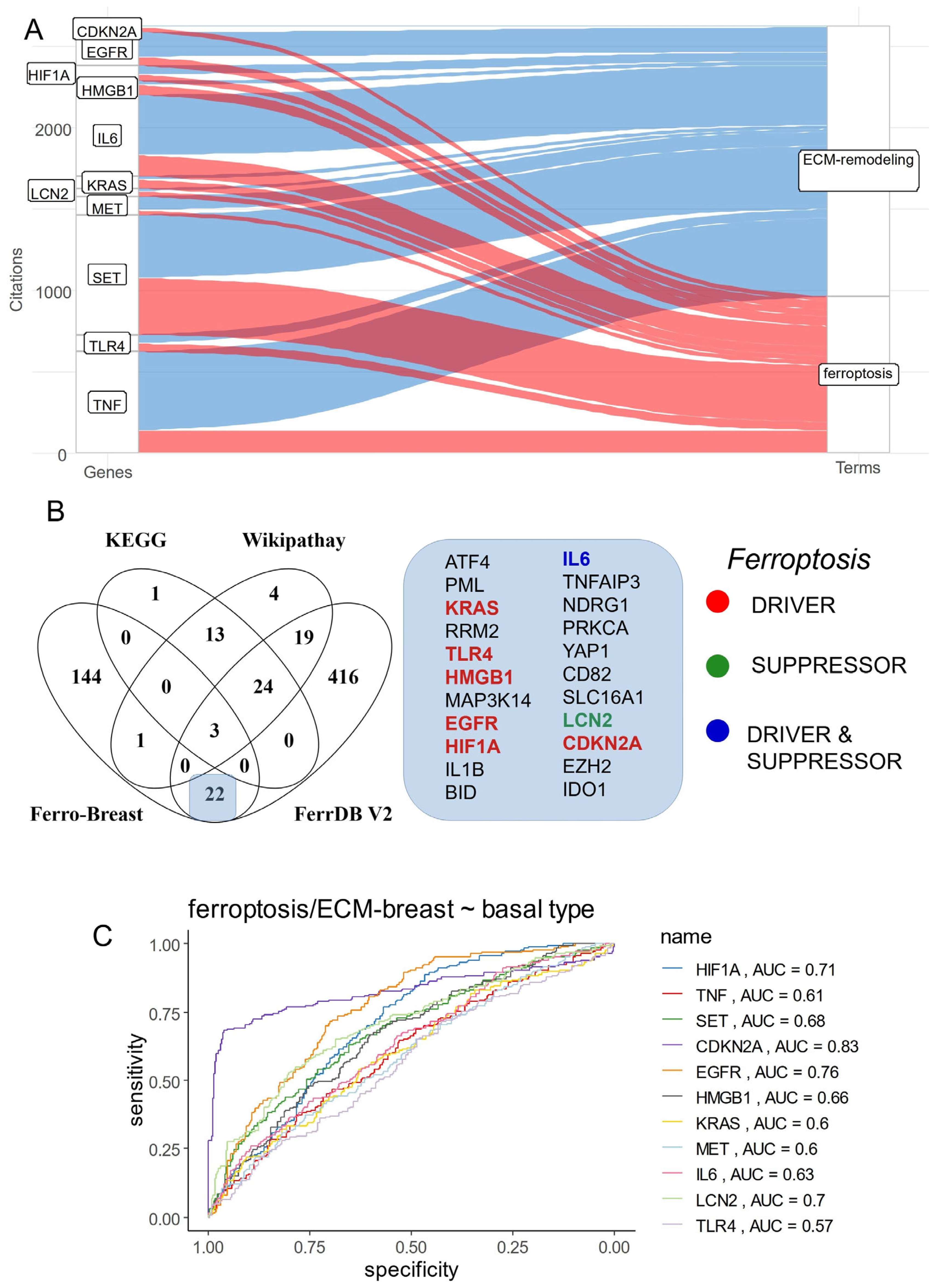
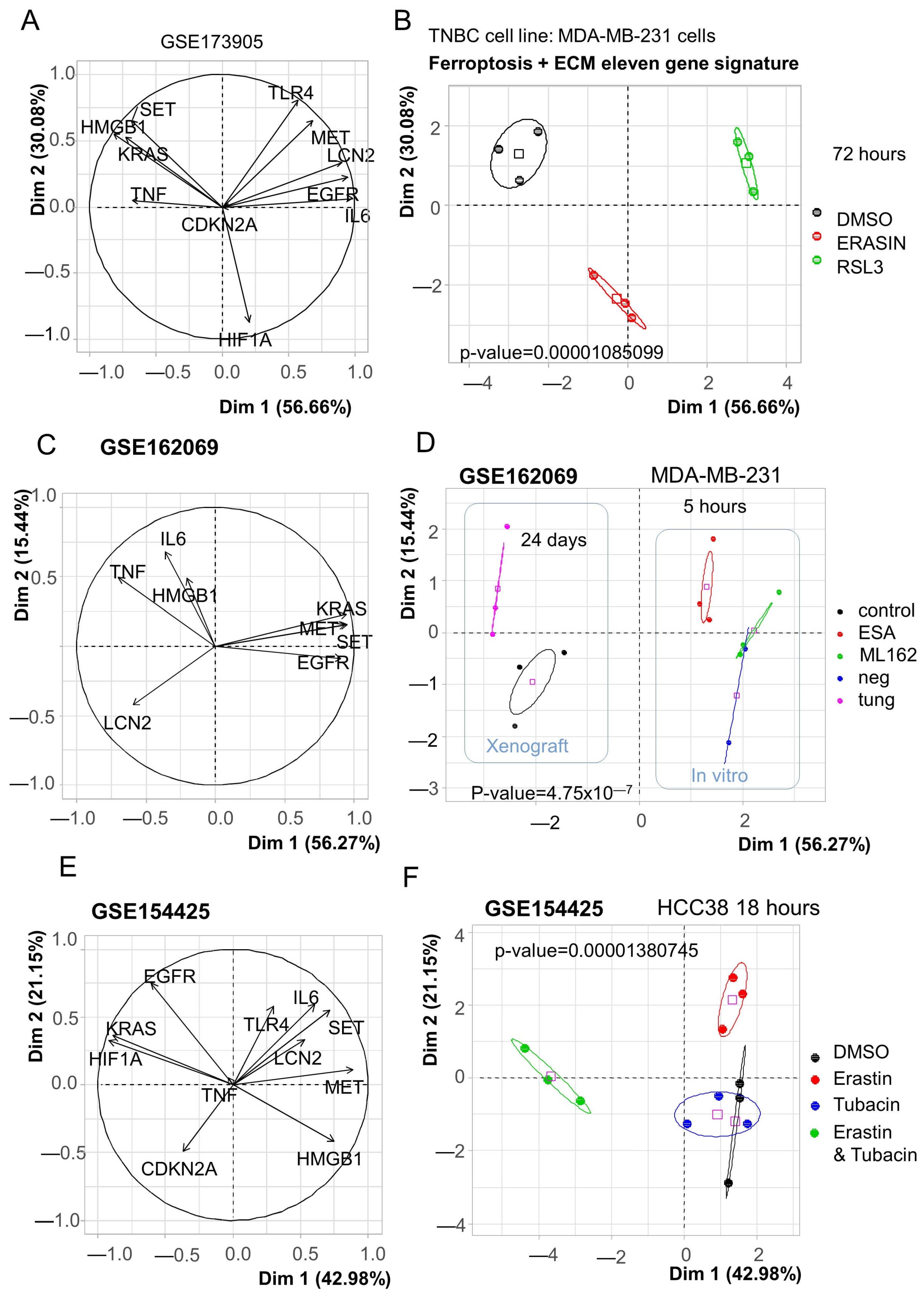
3.2. Breast Cancer Eleven-Gene Signature Implicated in Ferroptosis and Extracellular Matrix Remodeling
3.3. Ferroptosis/ECM Remodeling Signature Is Regulated by Ferroptosis Modifiers in Triple-Negative Breast Cancer Cells
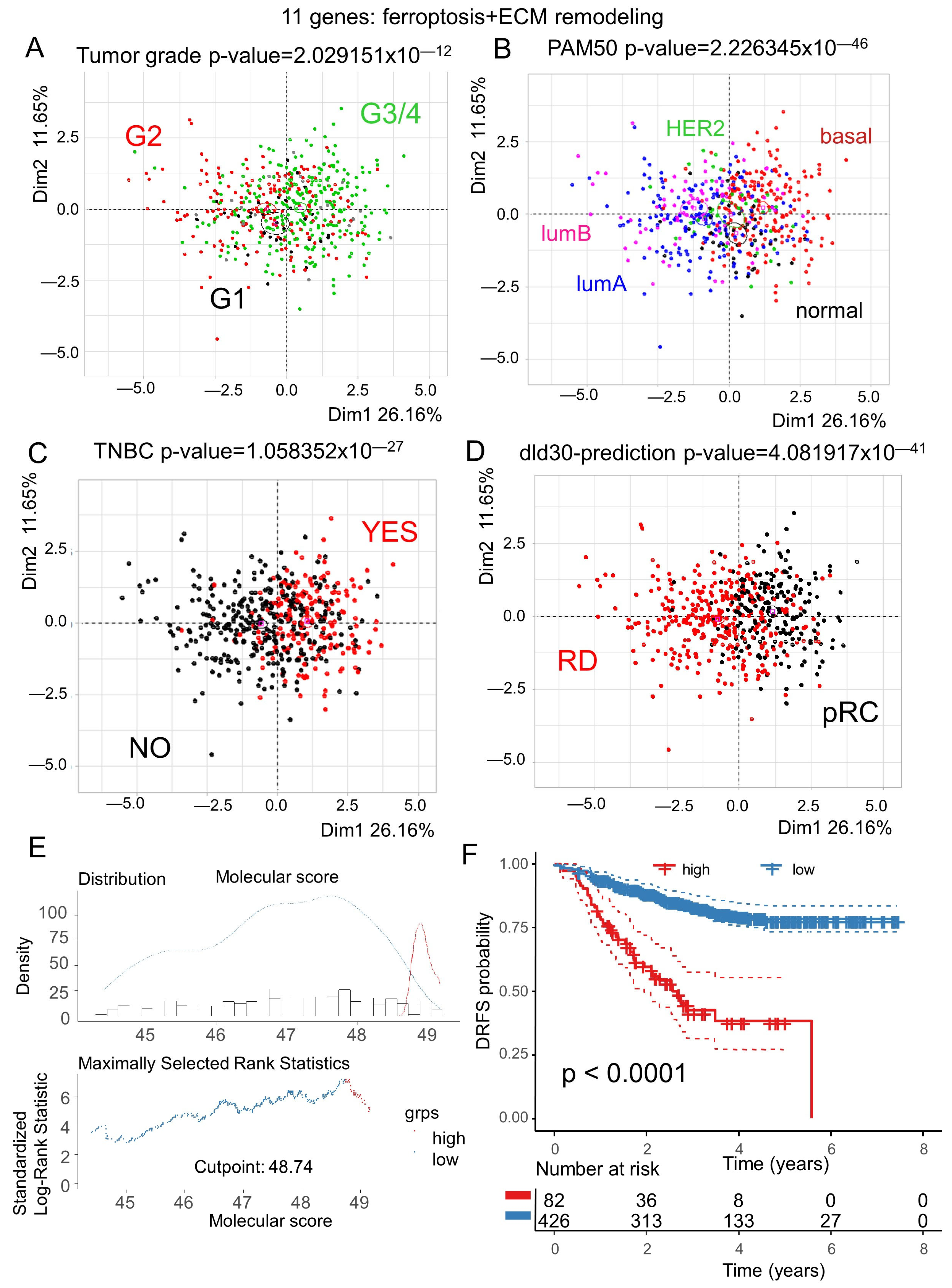
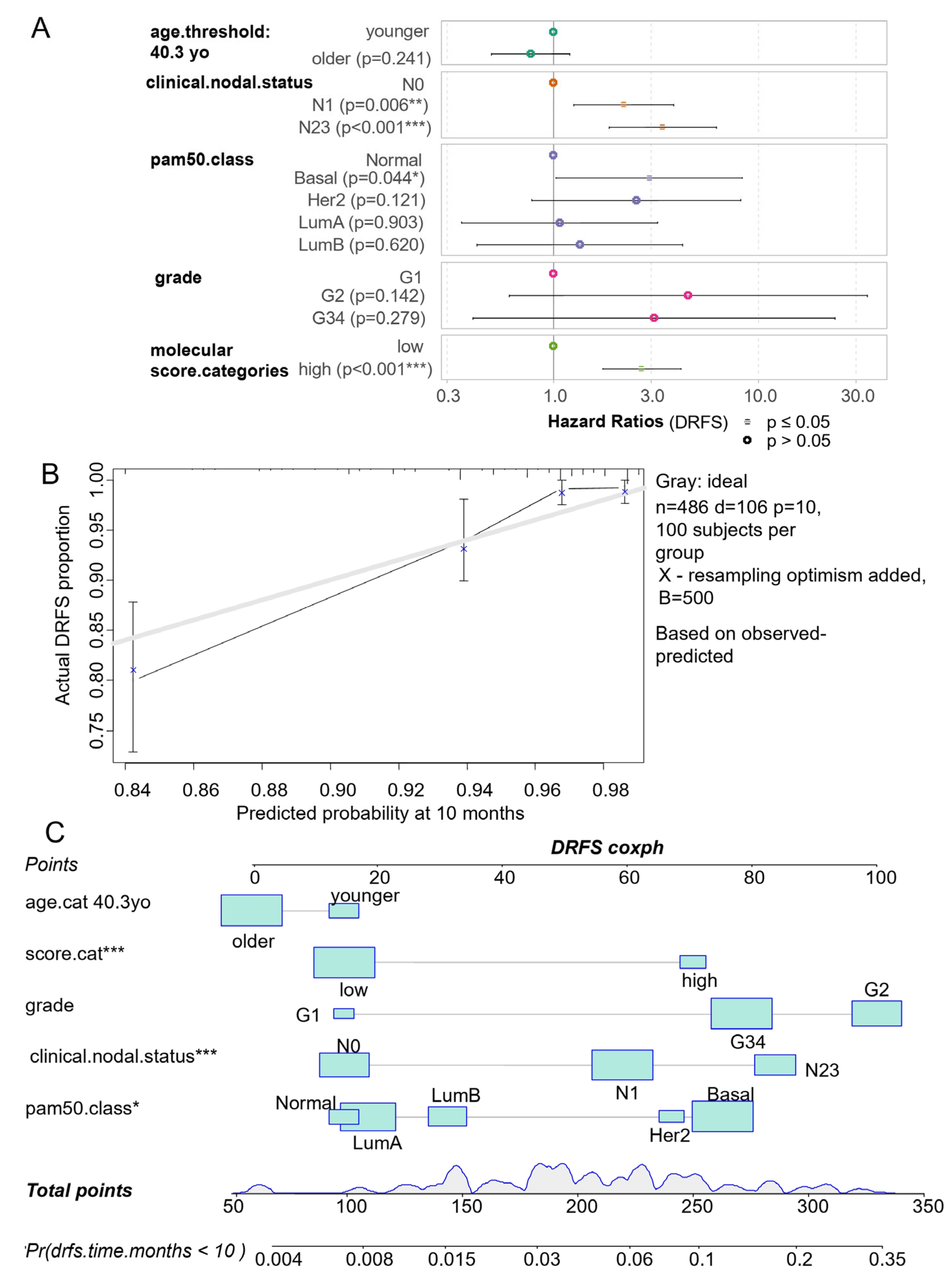
3.4. Ferroptosis/ECM Remodeling Molecular Score Is an Independent Adverse Parameter in the Prognosis of Breast Cancer
4. Discussion
5. Conclusions and Perspectives
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Siegel, R.L.; Miller, K.D.; Jemal, A. Cancer Statistics, 2020. CA Cancer J. Clin. 2020, 70, 7–30. [Google Scholar] [CrossRef]
- Perou, C.M.; Sørlie, T.; Eisen, M.B.; van de Rijn, M.; Jeffrey, S.S.; Rees, C.A.; Pollack, J.R.; Ross, D.T.; Johnsen, H.; Akslen, L.A.; et al. Molecular Portraits of Human Breast Tumours. Nature 2000, 406, 747–752. [Google Scholar] [CrossRef]
- Bastien, R.R.L.; Rodríguez-Lescure, Á.; Ebbert, M.T.W.; Prat, A.; Munárriz, B.; Rowe, L.; Miller, P.; Ruiz-Borrego, M.; Anderson, D.; Lyons, B.; et al. PAM50 Breast Cancer Subtyping by RT-QPCR and Concordance with Standard Clinical Molecular Markers. BMC Med. Genom. 2012, 5, 44. [Google Scholar] [CrossRef]
- Subhan, M.A.; Parveen, F.; Shah, H.; Yalamarty, S.S.K.; Ataide, J.A.; Torchilin, V.P. Recent Advances with Precision Medicine Treatment for Breast Cancer Including Triple-Negative Sub-Type. Cancers 2023, 15, 2204. [Google Scholar] [CrossRef]
- Liu, Y.; Hu, Y.; Jiang, Y.; Bu, J.; Gu, X. Targeting Ferroptosis, the Achilles’ Heel of Breast Cancer: A Review. Front. Pharmacol. 2022, 13, 1036140. [Google Scholar] [CrossRef]
- Dixon, S.J.; Lemberg, K.M.; Lamprecht, M.R.; Skouta, R.; Zaitsev, E.M.; Gleason, C.E.; Patel, D.N.; Bauer, A.J.; Cantley, A.M.; Yang, W.S.; et al. Ferroptosis: An Iron-Dependent Form of Nonapoptotic Cell Death. Cell 2012, 149, 1060–1072. [Google Scholar] [CrossRef]
- Tang, D.; Kroemer, G. Ferroptosis. Curr. Biol. 2020, 30, R1292–R1297. [Google Scholar] [CrossRef]
- Dolma, S.; Lessnick, S.L.; Hahn, W.C.; Stockwell, B.R. Identification of Genotype-Selective Antitumor Agents Using Synthetic Lethal Chemical Screening in Engineered Human Tumor Cells. Cancer Cell 2003, 3, 285–296. [Google Scholar] [CrossRef]
- Yang, W.S.; Stockwell, B.R. Synthetic Lethal Screening Identifies Compounds Activating Iron-Dependent, Nonapoptotic Cell Death in Oncogenic-RAS-Harboring Cancer Cells. Chem. Biol. 2008, 15, 234–245. [Google Scholar] [CrossRef]
- Luis, G.; Godfroid, A.; Nishiumi, S.; Cimino, J.; Blacher, S.; Maquoi, E.; Wery, C.; Collignon, A.; Longuespée, R.; Montero-Ruiz, L.; et al. Tumor Resistance to Ferroptosis Driven by Stearoyl-CoA Desaturase-1 (SCD1) in Cancer Cells and Fatty Acid Biding Protein-4 (FABP4) in Tumor Microenvironment Promote Tumor Recurrence. Redox Biol. 2021, 43, 102006. [Google Scholar] [CrossRef]
- Hassannia, B.; Vandenabeele, P.; Vanden Berghe, T. Targeting Ferroptosis to Iron Out Cancer. Cancer Cell 2019, 35, 830–849. [Google Scholar] [CrossRef]
- Sacco, A.; Battaglia, A.M.; Botta, C.; Aversa, I.; Mancuso, S.; Costanzo, F.; Biamonte, F. Iron Metabolism in the Tumor Microenvironment—Implications for Anti-Cancer Immune Response. Cells 2021, 10, 303. [Google Scholar] [CrossRef] [PubMed]
- Henke, E.; Nandigama, R.; Ergün, S. Extracellular Matrix in the Tumor Microenvironment and Its Impact on Cancer Therapy. Front. Mol. Biosci. 2020, 6, 160. [Google Scholar] [CrossRef]
- Fontaine, J.-F.; Priller, F.; Barbosa-Silva, A.; Andrade-Navarro, M.A. Génie: Literature-Based Gene Prioritization at Multi Genomic Scale. Nucleic Acids Res. 2011, 39, W455–W461. [Google Scholar] [CrossRef] [PubMed]
- Brancotte, B.; Biton, A.; Bernard-Pierrot, I.; Radvanyi, F.; Reyal, F.; Cohen-Boulakia, S. Gene List Significance At-a-Glance with GeneValorization. Bioinformatics 2011, 27, 1187–1189. [Google Scholar] [CrossRef] [PubMed]
- Sayers, E.W.; Barrett, T.; Benson, D.A.; Bryant, S.H.; Canese, K.; Chetvernin, V.; Church, D.M.; DiCuccio, M.; Edgar, R.; Federhen, S.; et al. Database Resources of the National Center for Biotechnology Information. Nucleic Acids Res. 2009, 37, D5–D15. [Google Scholar] [CrossRef] [PubMed]
- Hatzis, C. A Genomic Predictor of Response and Survival Following Taxane-Anthracycline Chemotherapy for Invasive Breast Cancer. JAMA 2011, 305, 1873. [Google Scholar] [CrossRef]
- Curtis, C.; Shah, S.P.; Chin, S.-F.; Turashvili, G.; Rueda, O.M.; Dunning, M.J.; Speed, D.; Lynch, A.G.; Samarajiwa, S.; Yuan, Y.; et al. The Genomic and Transcriptomic Architecture of 2000 Breast Tumours Reveals Novel Subgroups. Nature 2012, 486, 346–352. [Google Scholar] [CrossRef] [PubMed]
- Rueda, O.M.; Sammut, S.-J.; Seoane, J.A.; Chin, S.-F.; Caswell-Jin, J.L.; Callari, M.; Batra, R.; Pereira, B.; Bruna, A.; Ali, H.R.; et al. Dynamics of Breast-Cancer Relapse Reveal Late-Recurring ER-Positive Genomic Subgroups. Nature 2019, 567, 399–404. [Google Scholar] [CrossRef]
- Pereira, B.; Chin, S.-F.; Rueda, O.M.; Vollan, H.-K.M.; Provenzano, E.; Bardwell, H.A.; Pugh, M.; Jones, L.; Russell, R.; Sammut, S.-J.; et al. The Somatic Mutation Profiles of 2433 Breast Cancers Refines Their Genomic and Transcriptomic Landscapes. Nat. Commun. 2016, 7, 11479. [Google Scholar] [CrossRef]
- Gao, J.; Aksoy, B.A.; Dogrusoz, U.; Dresdner, G.; Gross, B.; Sumer, S.O.; Sun, Y.; Jacobsen, A.; Sinha, R.; Larsson, E.; et al. Integrative Analysis of Complex Cancer Genomics and Clinical Profiles Using the CBioPortal. Sci. Signal 2013, 6, pl1. [Google Scholar] [CrossRef] [PubMed]
- Li, P.; Lin, Q.; Sun, S.; Yang, N.; Xia, Y.; Cao, S.; Zhang, W.; Li, Q.; Guo, H.; Zhu, M.; et al. Inhibition of Cannabinoid Receptor Type 1 Sensitizes Triple-Negative Breast Cancer Cells to Ferroptosis via Regulating Fatty Acid Metabolism. Cell Death Dis. 2022, 13, 808. [Google Scholar] [CrossRef] [PubMed]
- Kim, D.; Langmead, B.; Salzberg, S.L. HISAT: A Fast Spliced Aligner with Low Memory Requirements. Nat. Methods 2015, 12, 357–360. [Google Scholar] [CrossRef] [PubMed]
- Liao, Y.; Smyth, G.K.; Shi, W. FeatureCounts: An Efficient General Purpose Program for Assigning Sequence Reads to Genomic Features. Bioinformatics 2014, 30, 923–930. [Google Scholar] [CrossRef]
- Beatty, A.; Singh, T.; Tyurina, Y.Y.; Tyurin, V.A.; Samovich, S.; Nicolas, E.; Maslar, K.; Zhou, Y.; Cai, K.Q.; Tan, Y.; et al. Ferroptotic Cell Death Triggered by Conjugated Linolenic Acids Is Mediated by ACSL1. Nat. Commun. 2021, 12, 2244. [Google Scholar] [CrossRef]
- Trapnell, C.; Pachter, L.; Salzberg, S.L. TopHat: Discovering Splice Junctions with RNA-Seq. Bioinformatics 2009, 25, 1105–1111. [Google Scholar] [CrossRef]
- Trapnell, C.; Roberts, A.; Goff, L.; Pertea, G.; Kim, D.; Kelley, D.R.; Pimentel, H.; Salzberg, S.L.; Rinn, J.L.; Pachter, L. Differential Gene and Transcript Expression Analysis of RNA-Seq Experiments with TopHat and Cufflinks. Nat. Protoc. 2012, 7, 562–578. [Google Scholar] [CrossRef]
- Alothaim, T.; Charbonneau, M.; Tang, X. HDAC6 Inhibitors Sensitize Non-Mesenchymal Triple-Negative Breast Cancer Cells to Cysteine Deprivation. Sci. Rep. 2021, 11, 10956. [Google Scholar] [CrossRef]
- Lê, S.; Josse, J.; Husson, F. FactoMineR: An R Package for Multivariate Analysis. J. Stat. Softw. 2008, 25, 1–18. [Google Scholar] [CrossRef]
- Uhlen, M.; Zhang, C.; Lee, S.; Sjöstedt, E.; Fagerberg, L.; Bidkhori, G.; Benfeitas, R.; Arif, M.; Liu, Z.; Edfors, F.; et al. A Pathology Atlas of the Human Cancer Transcriptome. Science 2017, 357, eaan2507. [Google Scholar] [CrossRef]
- Kim, M.-S.; Pinto, S.M.; Getnet, D.; Nirujogi, R.S.; Manda, S.S.; Chaerkady, R.; Madugundu, A.K.; Kelkar, D.S.; Isserlin, R.; Jain, S.; et al. A Draft Map of the Human Proteome. Nature 2014, 509, 575–581. [Google Scholar] [CrossRef]
- Hudis, C.A.; Barlow, W.E.; Costantino, J.P.; Gray, R.J.; Pritchard, K.I.; Chapman, J.-A.W.; Sparano, J.A.; Hunsberger, S.; Enos, R.A.; Gelber, R.D.; et al. Proposal for Standardized Definitions for Efficacy End Points in Adjuvant Breast Cancer Trials: The STEEP System. JCO 2007, 25, 2127–2132. [Google Scholar] [CrossRef]
- Liberzon, A.; Birger, C.; Thorvaldsdóttir, H.; Ghandi, M.; Mesirov, J.P.; Tamayo, P. The Molecular Signatures Database (MSigDB) Hallmark Gene Set Collection. Cell Syst. 2015, 1, 417–425. [Google Scholar] [CrossRef]
- Chen, J.; Bardes, E.E.; Aronow, B.J.; Jegga, A.G. ToppGene Suite for Gene List Enrichment Analysis and Candidate Gene Prioritization. Nucleic Acids Res. 2009, 37, W305–W311. [Google Scholar] [CrossRef] [PubMed]
- Cline, M.S.; Smoot, M.; Cerami, E.; Kuchinsky, A.; Landys, N.; Workman, C.; Christmas, R.; Avila-Campilo, I.; Creech, M.; Gross, B.; et al. Integration of Biological Networks and Gene Expression Data Using Cytoscape. Nat. Protoc. 2007, 2, 2366–2382. [Google Scholar] [CrossRef] [PubMed]
- Hess, K.R.; Anderson, K.; Symmans, W.F.; Valero, V.; Ibrahim, N.; Mejia, J.A.; Booser, D.; Theriault, R.L.; Buzdar, A.U.; Dempsey, P.J.; et al. Pharmacogenomic Predictor of Sensitivity to Preoperative Chemotherapy with Paclitaxel and Fluorouracil, Doxorubicin, and Cyclophosphamide in Breast Cancer. JCO 2006, 24, 4236–4244. [Google Scholar] [CrossRef] [PubMed]
- Symmans, W.F.; Peintinger, F.; Hatzis, C.; Rajan, R.; Kuerer, H.; Valero, V.; Assad, L.; Poniecka, A.; Hennessy, B.; Green, M.; et al. Measurement of Residual Breast Cancer Burden to Predict Survival after Neoadjuvant Chemotherapy. JCO 2007, 25, 4414–4422. [Google Scholar] [CrossRef] [PubMed]
- Smid, M.; Wang, Y.; Zhang, Y.; Sieuwerts, A.M.; Yu, J.; Klijn, J.G.M.; Foekens, J.A.; Martens, J.W.M. Subtypes of Breast Cancer Show Preferential Site of Relapse. Cancer Res. 2008, 68, 3108–3114. [Google Scholar] [CrossRef]
- Sotiriou, C.; Neo, S.-Y.; McShane, L.M.; Korn, E.L.; Long, P.M.; Jazaeri, A.; Martiat, P.; Fox, S.B.; Harris, A.L.; Liu, E.T. Breast Cancer Classification and Prognosis Based on Gene Expression Profiles from a Population-Based Study. Proc. Natl. Acad. Sci. USA 2003, 100, 10393–10398. [Google Scholar] [CrossRef]
- Niknafs, Y.S.; Pandian, B.; Gajjar, T.; Gaudette, Z.; Wheelock, K.; Maz, M.P.; Achar, R.K.; Song, M.; Massaro, C.; Cao, X.; et al. MiPanda: A Resource for Analyzing and Visualizing Next-Generation Sequencing Transcriptomics Data. Neoplasia 2018, 20, 1144–1149. [Google Scholar] [CrossRef] [PubMed]
- Zhou, N.; Yuan, X.; Du, Q.; Zhang, Z.; Shi, X.; Bao, J.; Ning, Y.; Peng, L. FerrDb V2: Update of the Manually Curated Database of Ferroptosis Regulators and Ferroptosis-Disease Associations. Nucleic Acids Res. 2023, 51, D571–D582. [Google Scholar] [CrossRef]
- Ogata, H.; Goto, S.; Sato, K.; Fujibuchi, W.; Bono, H.; Kanehisa, M. KEGG: Kyoto Encyclopedia of Genes and Genomes. Nucleic Acids Res. 1999, 27, 29–34. [Google Scholar] [CrossRef]
- Martens, M.; Ammar, A.; Riutta, A.; Waagmeester, A.; Slenter, D.N.; Hanspers, K.; A Miller, R.; Digles, D.; Lopes, E.N.; Ehrhart, F.; et al. WikiPathways: Connecting Communities. Nucleic Acids Res. 2021, 49, D613–D621. [Google Scholar] [CrossRef]
- Sharma, M.; Turaga, R.C.; Yuan, Y.; Satyanarayana, G.; Mishra, F.; Bian, Z.; Liu, W.; Sun, L.; Yang, J.; Liu, Z.-R. Simultaneously Targeting Cancer-Associated Fibroblasts and Angiogenic Vessel as a Treatment for TNBC. J. Exp. Med. 2021, 218, e20200712. [Google Scholar] [CrossRef] [PubMed]
- Doll, S.; Proneth, B.; Tyurina, Y.Y.; Panzilius, E.; Kobayashi, S.; Ingold, I.; Irmler, M.; Beckers, J.; Aichler, M.; Walch, A.; et al. ACSL4 Dictates Ferroptosis Sensitivity by Shaping Cellular Lipid Composition. Nat. Chem. Biol. 2017, 13, 91–98. [Google Scholar] [CrossRef] [PubMed]
- Timmerman, L.A.; Holton, T.; Yuneva, M.; Louie, R.J.; Padró, M.; Daemen, A.; Hu, M.; Chan, D.A.; Ethier, S.P.; van ‘t Veer, L.J.; et al. Glutamine Sensitivity Analysis Identifies the XCT Antiporter as a Common Triple-Negative Breast Tumor Therapeutic Target. Cancer Cell 2013, 24, 450–465. [Google Scholar] [CrossRef] [PubMed]
- Sun, L.-L.; Linghu, D.-L.; Hung, M.-C. Ferroptosis: A Promising Target for Cancer Immunotherapy. Am. J. Cancer Res. 2021, 11, 5856–5863. [Google Scholar]
- Lehmann, B.D.; Bauer, J.A.; Chen, X.; Sanders, M.E.; Chakravarthy, A.B.; Shyr, Y.; Pietenpol, J.A. Identification of Human Triple-Negative Breast Cancer Subtypes and Preclinical Models for Selection of Targeted Therapies. J. Clin. Investig. 2011, 121, 2750–2767. [Google Scholar] [CrossRef]
- Yang, F.; Xiao, Y.; Ding, J.-H.; Jin, X.; Ma, D.; Li, D.-Q.; Shi, J.-X.; Huang, W.; Wang, Y.-P.; Jiang, Y.-Z.; et al. Ferroptosis Heterogeneity in Triple-Negative Breast Cancer Reveals an Innovative Immunotherapy Combination Strategy. Cell Metab. 2023, 35, 84–100.e8. [Google Scholar] [CrossRef] [PubMed]
- Jiang, Y.-Z.; Ma, D.; Suo, C.; Shi, J.; Xue, M.; Hu, X.; Xiao, Y.; Yu, K.-D.; Liu, Y.-R.; Yu, Y.; et al. Genomic and Transcriptomic Landscape of Triple-Negative Breast Cancers: Subtypes and Treatment Strategies. Cancer Cell 2019, 35, 428–440.e5. [Google Scholar] [CrossRef]
- Minami, J.K.; Morrow, D.; Bayley, N.A.; Fernandez, E.G.; Salinas, J.J.; Tse, C.; Zhu, H.; Su, B.; Plawat, R.; Jones, A.; et al. CDKN2A Deletion Remodels Lipid Metabolism to Prime Glioblastoma for Ferroptosis. Cancer Cell 2023, 41, 1048–1060.e9. [Google Scholar] [CrossRef]
- Al-Ansari, M.M.; Hendrayani, S.F.; Shehata, A.I.; Aboussekhra, A. P16(INK4A) Represses the Paracrine Tumor-Promoting Effects of Breast Stromal Fibroblasts. Oncogene 2013, 32, 2356–2364. [Google Scholar] [CrossRef]
- Shen, G.-M.; Zhao, Y.-Z.; Chen, M.-T.; Zhang, F.-L.; Liu, X.-L.; Wang, Y.; Liu, C.-Z.; Yu, J.; Zhang, J.-W. Hypoxia-Inducible Factor-1 (HIF-1) Promotes LDL and VLDL Uptake through Inducing VLDLR under Hypoxia. Biochem. J. 2012, 441, 675–683. [Google Scholar] [CrossRef]
- Bensaad, K.; Favaro, E.; Lewis, C.A.; Peck, B.; Lord, S.; Collins, J.M.; Pinnick, K.E.; Wigfield, S.; Buffa, F.M.; Li, J.-L.; et al. Fatty Acid Uptake and Lipid Storage Induced by HIF-1α Contribute to Cell Growth and Survival after Hypoxia-Reoxygenation. Cell Rep. 2014, 9, 349–365. [Google Scholar] [CrossRef] [PubMed]
- Yang, M.; Chen, P.; Liu, J.; Zhu, S.; Kroemer, G.; Klionsky, D.J.; Lotze, M.T.; Zeh, H.J.; Kang, R.; Tang, D. Clockophagy Is a Novel Selective Autophagy Process Favoring Ferroptosis. Sci. Adv. 2019, 5, eaaw2238. [Google Scholar] [CrossRef]
- Liu, J.; Yang, M.; Kang, R.; Klionsky, D.J.; Tang, D. Autophagic Degradation of the Circadian Clock Regulator Promotes Ferroptosis. Autophagy 2019, 15, 2033–2035. [Google Scholar] [CrossRef] [PubMed]
- Assidicky, R.; Tokat, U.M.; Tarman, I.O.; Saatci, O.; Ersan, P.G.; Raza, U.; Ogul, H.; Riazalhosseini, Y.; Can, T.; Sahin, O. Targeting HIF1-Alpha/MiR-326/ITGA5 Axis Potentiates Chemotherapy Response in Triple-Negative Breast Cancer. Breast Cancer Res. Treat. 2022, 193, 331–348. [Google Scholar] [CrossRef]
- Rebalka, I.A.; Monaco, C.M.F.; Varah, N.E.; Berger, T.; D’souza, D.M.; Zhou, S.; Mak, T.W.; Hawke, T.J. Loss of the Adipokine Lipocalin-2 Impairs Satellite Cell Activation and Skeletal Muscle Regeneration. Am. J. Physiol. Cell Physiol. 2018, 315, C714–C721. [Google Scholar] [CrossRef]
- Valashedi, M.R.; Roushandeh, A.M.; Tomita, K.; Kuwahara, Y.; Pourmohammadi-Bejarpasi, Z.; Kozani, P.S.; Sato, T.; Roudkenar, M.H. CRISPR/Cas9-Mediated Knockout of Lcn2 in Human Breast Cancer Cell Line MDA-MB-231 Ameliorates Erastin-Mediated Ferroptosis and Increases Cisplatin Vulnerability. Life Sci. 2022, 304, 120704. [Google Scholar] [CrossRef]
- Lee, Y.; Jung, W.H.; Koo, J.S. Adipocytes Can Induce Epithelial-Mesenchymal Transition in Breast Cancer Cells. Breast Cancer Res. Treat. 2015, 153, 323–335. [Google Scholar] [CrossRef]
- Dirat, B.; Bochet, L.; Dabek, M.; Daviaud, D.; Dauvillier, S.; Majed, B.; Wang, Y.Y.; Meulle, A.; Salles, B.; Le Gonidec, S.; et al. Cancer-Associated Adipocytes Exhibit an Activated Phenotype and Contribute to Breast Cancer Invasion. Cancer Res. 2011, 71, 2455–2465. [Google Scholar] [CrossRef]
- Zhao, C.; Wu, M.; Zeng, N.; Xiong, M.; Hu, W.; Lv, W.; Yi, Y.; Zhang, Q.; Wu, Y. Cancer-Associated Adipocytes: Emerging Supporters in Breast Cancer. J. Exp. Clin. Cancer Res. 2020, 39, 156. [Google Scholar] [CrossRef] [PubMed]
- Gyamfi, J.; Eom, M.; Koo, J.-S.; Choi, J. Multifaceted Roles of Interleukin-6 in Adipocyte-Breast Cancer Cell Interaction. Transl. Oncol. 2018, 11, 275–285. [Google Scholar] [CrossRef]
- Li, H.; Yang, P.; Wang, J.; Zhang, J.; Ma, Q.; Jiang, Y.; Wu, Y.; Han, T.; Xiang, D. HLF Regulates Ferroptosis, Development and Chemoresistance of Triple-Negative Breast Cancer by Activating Tumor Cell-Macrophage Crosstalk. J. Hematol. Oncol. 2022, 15, 2. [Google Scholar] [CrossRef]
- Liu, X.; Adorno-Cruz, V.; Chang, Y.-F.; Jia, Y.; Kawaguchi, M.; Dashzeveg, N.K.; Taftaf, R.; Ramos, E.K.; Schuster, E.J.; El-Shennawy, L.; et al. EGFR Inhibition Blocks Cancer Stem Cell Clustering and Lung Metastasis of Triple Negative Breast Cancer. Theranostics 2021, 11, 6632–6643. [Google Scholar] [CrossRef] [PubMed]
- Wu, X.; Sheng, H.; Zhao, L.; Jiang, M.; Lou, H.; Miao, Y.; Cheng, N.; Zhang, W.; Ding, D.; Li, W. Co-Loaded Lapatinib/PAB by Ferritin Nanoparticles Eliminated ECM-Detached Cluster Cells via Modulating EGFR in Triple-Negative Breast Cancer. Cell Death Dis. 2022, 13, 557. [Google Scholar] [CrossRef] [PubMed]
- Kyriakopoulou, K.; Kefali, E.; Piperigkou, Z.; Riethmüller, C.; Greve, B.; Franchi, M.; Götte, M.; Karamanos, N.K. EGFR Is a Pivotal Player of the E2/Erβ—Mediated Functional Properties, Aggressiveness, and Stemness in Triple-Negative Breast Cancer Cells. FEBS J. 2022, 289, 1552–1574. [Google Scholar] [CrossRef] [PubMed]
- Wood, G.E.; Hockings, H.; Hilton, D.M.; Kermorgant, S. The Role of MET in Chemotherapy Resistance. Oncogene 2021, 40, 1927–1941. [Google Scholar] [CrossRef]
- Matsumoto, K.; Nakamura, T. Roles of HGF as a Pleiotropic Factor in Organ Regeneration. EXS 1993, 65, 225–249. [Google Scholar]
- Tovar, E.A.; Graveel, C.R. MET in Human Cancer: Germline and Somatic Mutations. Ann. Transl. Med. 2017, 5, 205. [Google Scholar] [CrossRef]
- Wu, Q.; Song, L.; Guo, Y.; Liu, S.; Wang, W.; Liu, H.; Gong, A.; Liao, X.; Zhu, H.; Wang, D. Activated Stellate Cell Paracrine HGF Exacerbated Pancreatic Cancer Cell Ferroptosis Resistance. Oxidative Med. Cell Longev. 2022, 2022, 2985249. [Google Scholar] [CrossRef] [PubMed]
- Chen, R.; Zou, J.; Kang, R.; Tang, D. The Redox Protein High-Mobility Group Box 1 in Cell Death and Cancer. Antioxid. Redox Signal 2023. [Google Scholar] [CrossRef] [PubMed]
- Wu, Y.; Zhao, Y.; Yang, H.-Z.; Wang, Y.-J.; Chen, Y. HMGB1 Regulates Ferroptosis through Nrf2 Pathway in Mesangial Cells in Response to High Glucose. Biosci. Rep. 2021, 41, BSR20202924. [Google Scholar] [CrossRef]
- Wang, L.; Kang, F.-B.; Wang, J.; Yang, C.; He, D.-W. Downregulation of MiR-205 Contributes to Epithelial-Mesenchymal Transition and Invasion in Triple-Negative Breast Cancer by Targeting HMGB1-RAGE Signaling Pathway. Anticancer Drugs 2019, 30, 225–232. [Google Scholar] [CrossRef]
- Sánchez-Muñoz, A.; Gallego, E.; de Luque, V.; Pérez-Rivas, L.G.; Vicioso, L.; Ribelles, N.; Lozano, J.; Alba, E. Lack of Evidence for KRAS Oncogenic Mutations in Triple-Negative Breast Cancer. BMC Cancer 2010, 10, 136. [Google Scholar] [CrossRef]
- Kim, R.-K.; Suh, Y.; Yoo, K.-C.; Cui, Y.-H.; Kim, H.; Kim, M.-J.; Gyu Kim, I.; Lee, S.-J. Activation of KRAS Promotes the Mesenchymal Features of Basal-Type Breast Cancer. Exp. Mol. Med. 2015, 47, e137. [Google Scholar] [CrossRef]
- Dai, E.; Han, L.; Liu, J.; Xie, Y.; Kroemer, G.; Klionsky, D.J.; Zeh, H.J.; Kang, R.; Wang, J.; Tang, D. Autophagy-Dependent Ferroptosis Drives Tumor-Associated Macrophage Polarization via Release and Uptake of Oncogenic KRAS Protein. Autophagy 2020, 16, 2069–2083. [Google Scholar] [CrossRef]
- Rajput, S.; Volk-Draper, L.D.; Ran, S. TLR4 Is a Novel Determinant of the Response to Paclitaxel in Breast Cancer. Mol. Cancer Ther. 2013, 12, 1676–1687. [Google Scholar] [CrossRef]
- Zhu, K.; Zhu, X.; Sun, S.; Yang, W.; Liu, S.; Tang, Z.; Zhang, R.; Li, J.; Shen, T.; Hei, M. Inhibition of TLR4 Prevents Hippocampal Hypoxic-Ischemic Injury by Regulating Ferroptosis in Neonatal Rats. Exp. Neurol. 2021, 345, 113828. [Google Scholar] [CrossRef]
- Feng, R.; Xiong, Y.; Lei, Y.; Huang, Q.; Liu, H.; Zhao, X.; Chen, Z.; Chen, H.; Liu, X.; Wang, L.; et al. Lysine-Specific Demethylase 1 Aggravated Oxidative Stress and Ferroptosis Induced by Renal Ischemia and Reperfusion Injury through Activation of TLR4/NOX4 Pathway in Mice. J. Cell Mol. Med. 2022, 26, 4254–4267. [Google Scholar] [CrossRef] [PubMed]
- Liu, C.-Y.; Huang, T.-T.; Chen, Y.-T.; Chen, J.-L.; Chu, P.-Y.; Huang, C.-T.; Wang, W.-L.; Lau, K.-Y.; Dai, M.-S.; Shiau, C.-W.; et al. Targeting SET to Restore PP2A Activity Disrupts an Oncogenic CIP2A-Feedforward Loop and Impairs Triple Negative Breast Cancer Progression. eBioMedicine 2019, 40, 263–275. [Google Scholar] [CrossRef] [PubMed]
- Katanov, C.; Lerrer, S.; Liubomirski, Y.; Leider-Trejo, L.; Meshel, T.; Bar, J.; Feniger-Barish, R.; Kamer, I.; Soria-Artzi, G.; Kahani, H.; et al. Regulation of the Inflammatory Profile of Stromal Cells in Human Breast Cancer: Prominent Roles for TNF-α and the NF-ΚB Pathway. Stem Cell Res. Ther. 2015, 6, 87. [Google Scholar] [CrossRef] [PubMed]
- Yin, J.; Meng, X.; Peng, L.; Xie, W.; Liu, X.; He, W.; Li, S. Ferroptosis and Cancer Immunotherapy. Curr. Mol. Med. 2023, 23, 401–409. [Google Scholar] [CrossRef] [PubMed]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Variable | Level | Low (n = 426) | High (n = 82) | Total (n = 508) | p-Value |
|---|---|---|---|---|---|
| age.years | mean (sd) | 49.7 (10.4) | 50.5 (10.8) | 49.8 (10.5) | 0.524161 |
| age.categories (40.3 yo) | younger | 79 (18.5) | 20 (24.4) | 99 (19.5) | |
| older | 347 (81.5) | 62 (75.6) | 409 (80.5) | 0.283927 | |
| er.status.ihc | Negative | 144 (34.1) | 61 (76.2) | 205 (40.8) | |
| Positive | 278 (65.9) | 19 (23.8) | 297 (59.2) | <0.0001 | |
| missing | 4 | 2 | 6 | ||
| pr.status.ihc | Negative | 192 (45.6) | 66 (82.5) | 258 (51.5) | |
| Positive | 229 (54.4) | 14 (17.5) | 243 (48.5) | <0.0001 | |
| missing | 5 | 2 | 7 | ||
| TNBC | no | 289 (70.3) | 22 (28.2) | 311 (63.6) | |
| YES | 122 (29.7) | 56 (71.8) | 178 (36.4) | <0.0001 | |
| missing | 15 | 4 | 19 | ||
| pam50.class | Normal | 42 (9.9) | 2 (2.4) | 44 (8.7) | |
| Basal | 123 (28.9) | 66 (80.5) | 189 (37.2) | ||
| Her2 | 33 (7.7) | 4 (4.9) | 37 (7.3) | ||
| LumA | 152 (35.7) | 8 (9.8) | 160 (31.5) | ||
| LumB | 76 (17.8) | 2 (2.4) | 78 (15.4) | <0.0001 | |
| clinical.tumor.stage | T-0,1,2 | 247 (58.0) | 41 (50.0) | 288 (56.7) | |
| T-3 | 119 (27.9) | 26 (31.7) | 145 (28.5) | ||
| T-4 | 60 (14.1) | 15 (18.3) | 75 (14.8) | 0.379010 | |
| clinical.nodal.status | N-0 | 140 (32.9) | 17 (20.7) | 157 (30.9) | |
| N-1 | 205 (48.1) | 39 (47.6) | 244 (48.0) | ||
| N-2,3 | 81 (19.0) | 26 (31.7) | 107 (21.1) | 0.013986 | |
| clinical.ajcc.stage | IIB | 131 (30.8) | 20 (24.4) | 151 (29.7) | |
| IIIA | 99 (23.2) | 22 (26.8) | 121 (23.8) | ||
| IIIB | 63 (14.8) | 17 (20.7) | 80 (15.7) | ||
| IIA | 109 (25.6) | 12 (14.6) | 121 (23.8) | ||
| IIIC | 16 (3.8) | 7 (8.5) | 23 (4.5) | ||
| Inflammatory | 2 (0.5) | 2 (2.4) | 4 (0.8) | ||
| I | 6 (1.4) | 2 (2.4) | 8 (1.6) | 0.033976 | |
| grade | G-1 | 32 (7.8) | 0 (0.0) | 32 (6.6) | |
| G-2 | 167 (40.8) | 13 (16.9) | 180 (37.0) | ||
| G-3,4 | 210 (51.3) | 64 (83.1) | 274 (56.4) | <0.0001 | |
| missing | 17 | 5 | 22 | ||
| drfs.status | 1 | 70 (16.4) | 41 (50.0) | 111 (21.9) | |
| 0 | 356 (83.6) | 41 (50.0) | 397 (78.1) | <0.0001 | |
| drfs.time.years | mean (sd) | 3.2 (1.6) | 2 (1.2) | 3 (1.6) | <0.0001 |
| type.taxane | Taxotere | 78 (45.6) | 14 (51.9) | 92 (46.5) | |
| Taxol | 93 (54.4) | 13 (48.1) | 106 (53.5) | 0.691854 | |
| missing | 255 | 55 | 310 |
| Variables | Hazard Ratios | Confidence-Low | Confidence-High | p-Value |
|---|---|---|---|---|
| age.cat older | 0.770 | 0.498 | 1.192 | 2.41 × 10−1 |
| clinical.nodal.statusN1 | 2.202 | 1.255 | 3.862 | 5.91 × 10−3 |
| clinical.nodal.statusN23 | 3.395 | 1.857 | 6.205 | 7.15 × 10−5 |
| pam50.classBasal | 2.924 | 1.027 | 8.327 | 4.45 × 10−2 |
| pam50.classHer2 | 2.530 | 0.783 | 8.177 | 1.21 × 10−1 |
| pam50.classLumA | 1.071 | 0.356 | 3.221 | 9.03 × 10−1 |
| pam50.classLumB | 1.339 | 0.422 | 4.252 | 6.20 × 10−1 |
| Grade.G2 | 4.526 | 0.603 | 33.950 | 1.42 × 10−1 |
| Grade.G34 | 3.085 | 0.402 | 23.693 | 2.79 × 10−1 |
| score.high | 2.689 | 1.728 | 4.185 | 1.17 × 10−5 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Desterke, C.; Cosialls, E.; Xiang, Y.; Elhage, R.; Duruel, C.; Chang, Y.; Hamaï, A. Adverse Crosstalk between Extracellular Matrix Remodeling and Ferroptosis in Basal Breast Cancer. Cells 2023, 12, 2176. https://doi.org/10.3390/cells12172176
Desterke C, Cosialls E, Xiang Y, Elhage R, Duruel C, Chang Y, Hamaï A. Adverse Crosstalk between Extracellular Matrix Remodeling and Ferroptosis in Basal Breast Cancer. Cells. 2023; 12(17):2176. https://doi.org/10.3390/cells12172176
Chicago/Turabian StyleDesterke, Christophe, Emma Cosialls, Yao Xiang, Rima Elhage, Clémence Duruel, Yunhua Chang, and Ahmed Hamaï. 2023. "Adverse Crosstalk between Extracellular Matrix Remodeling and Ferroptosis in Basal Breast Cancer" Cells 12, no. 17: 2176. https://doi.org/10.3390/cells12172176