
Cognitive Deficits in Aging Related to Changes in Basal Forebrain Neuronal Activity
 , and
, and 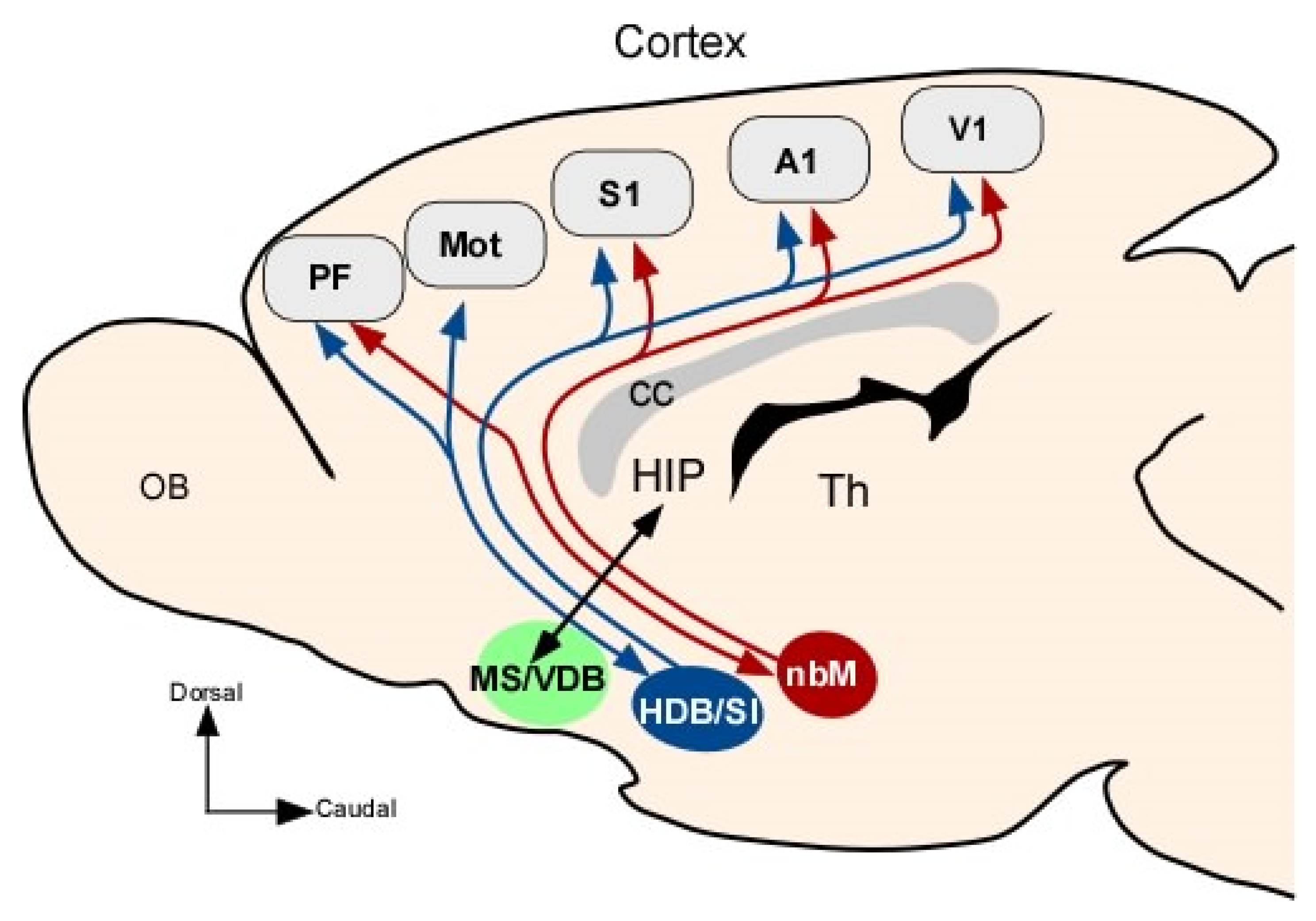{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Electroencephalographic Changes Provoked by Aging
3. The Basal Forebrain
4. Neuronal Changes during Aging
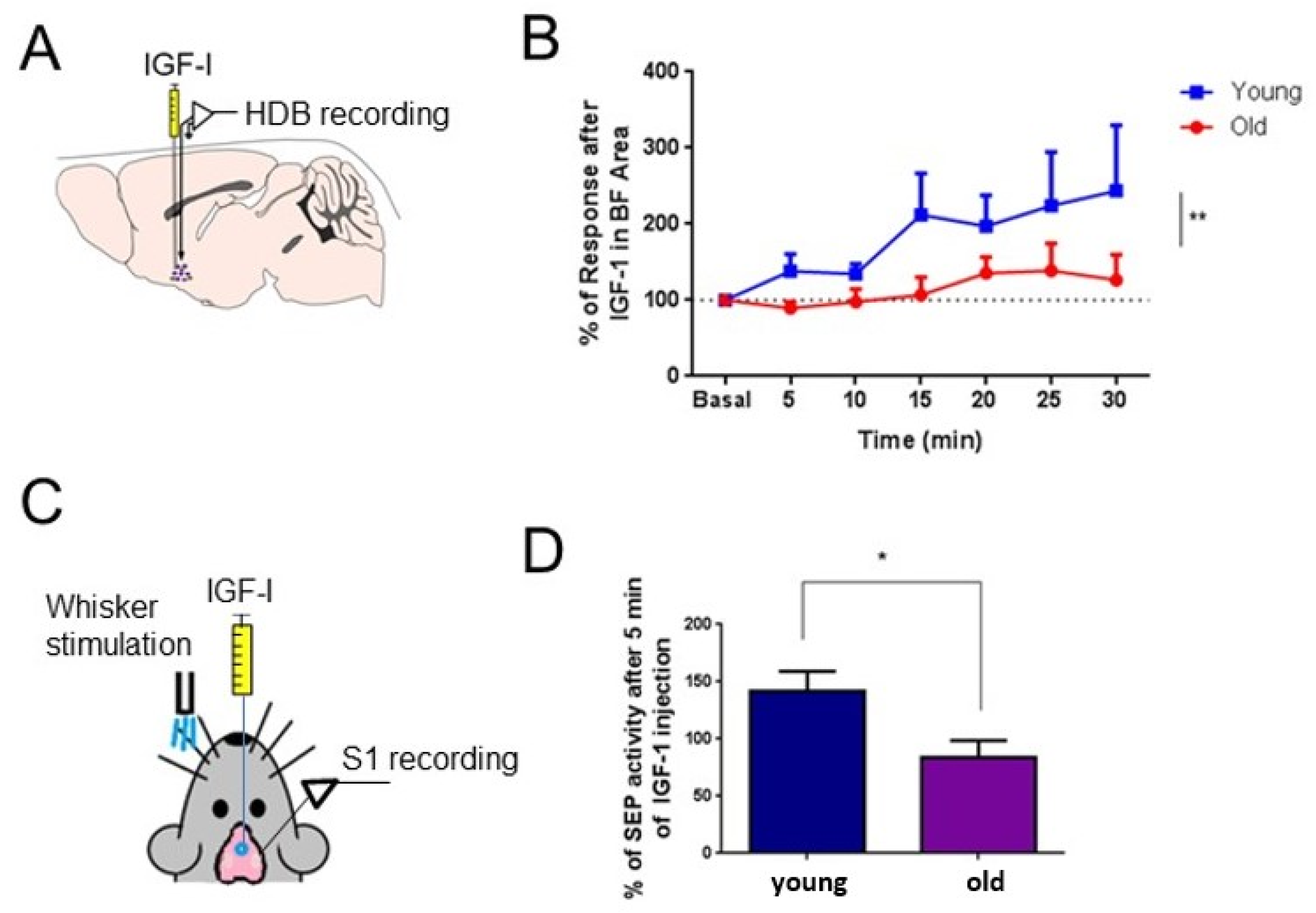
5. Insulin-like Growth Factor-I and the Aging Brain
6. Aging and Sleep
7. Neurodegenerative Diseases in Aging
8. Conclusions and Future Directions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Kenyon, C. A conserved regulatory system for aging. Cell 2001, 105, 165–168. [Google Scholar] [CrossRef]
- Smith, R.G.; Betancourt, L.; Sun, Y. Molecular endocrinology and physiology of the aging central nervous system. Endocr. Rev. 2005, 26, 203–250. [Google Scholar] [CrossRef]
- Burke, S.N.; Barnes, C.A. Neural plasticity in the ageing brain. Nat. Rev. Neurosci. 2006, 7, 30–40. [Google Scholar] [CrossRef]
- Morrison, J.H.; Baxter, M.G. The ageing cortical synapse: Hallmarks and implications for cognitive decline. Nat. Rev. Neurosci. 2012, 13, 240–250. [Google Scholar] [CrossRef]
- Petralia, R.S.; Mattson, M.P.; Yao, P.J. Communication breakdown: The impact of ageing on synapse structure. Ageing Res. Rev. 2014, 14, 31–42. [Google Scholar] [CrossRef]
- Masliah, E.; Mallory, M.; Hansen, L.; DeTeresa, R.; Terry, R.D. Quantitative synaptic alterations in the human neocortex during normal aging. Neurology 1993, 43, 192–197. [Google Scholar] [CrossRef]
- Enzinger, C.; Fazekas, F.; Matthews, P.M.; Ropele, S.; Schmidt, H.; Smith, S.; Schmidt, R. Risk factors for progression of brain atrophy in aging: Six-year follow-up of normal subjects. Neurology 2005, 64, 1704–1711. [Google Scholar] [CrossRef]
- Marner, L.; Nyengaard, J.R.; Tang, Y.; Pakkenberg, B. Marked loss of myelinated nerve fibers in the human brain with age. J. Comp. Neurol. 2003, 462, 144–152. [Google Scholar] [CrossRef]
- Ziegler, D.A.; Piguet, O.; Salat, D.H.; Prince, K.; Connally, E.; Corkin, S. Cognition in healthy aging is related to regional white matter integrity, but not cortical thickness. Neurobiol. Aging 2010, 31, 1912–1926. [Google Scholar] [CrossRef]
- Nunez, A. Unit activity of rat basal forebrain neurons: Relationship to cortical activity. Neuroscience 1996, 72, 757–766. [Google Scholar] [CrossRef]
- Manns, I.D.; Alonso, A.; Jones, B.E. Discharge profiles of juxtacellularly labeled and immunohistochemically identified GABAergic basal forebrain neurons recorded in association with the electroencephalogram in anesthetized rats. J. Neurosci. 2000, 20, 9252–9263. [Google Scholar] [CrossRef]
- Campos-Beltran, D.; Marshall, L. Changes in sleep EEG with aging in humans and rodents. Pflug. Arch. 2021, 473, 841–851. [Google Scholar] [CrossRef]
- Polich, J. On the relationship between EEG and P300: Individual differences, aging, and ultradian rhythms. Int. J. Psychophysiol. 1997, 26, 299–317. [Google Scholar] [CrossRef]
- Polich, J. EEG and ERP assessment of normal aging. Electroencephalogr. Clin. Neurophysiol. 1997, 104, 244–256. [Google Scholar] [CrossRef]
- Ishii, R.; Canuet, L.; Aoki, Y.; Hata, M.; Iwase, M.; Ikeda, S.; Nishida, K.; Ikeda, M. Healthy and Pathological Brain Aging: From the Perspective of Oscillations, Functional Connectivity, and Signal Complexity. Neuropsychobiology 2017, 75, 151–161. [Google Scholar] [CrossRef]
- Salat, D.H.; Tuch, D.S.; Greve, D.N.; van der Kouwe, A.J.; Hevelone, N.D.; Zaleta, A.K.; Rosen, B.R.; Fischl, B.; Corkin, S.; Rosas, H.D.; et al. Age-related alterations in white matter microstructure measured by diffusion tensor imaging. Neurobiol. Aging 2005, 26, 1215–1227. [Google Scholar] [CrossRef]
- Fjell, A.M.; Walhovd, K.B. Structural brain changes in aging: Courses, causes and cognitive consequences. Rev. Neurosci. 2010, 21, 187–221. [Google Scholar] [CrossRef]
- Thambisetty, M.; Wan, J.; Carass, A.; An, Y.; Prince, J.L.; Resnick, S.M. Longitudinal changes in cortical thickness associated with normal aging. Neuroimage 2010, 52, 1215–1223. [Google Scholar] [CrossRef]
- Mesulam, M.M.; Hersh, L.B.; Mash, D.C.; Geula, C. Differential cholinergic innervation within functional subdivisions of the human cerebral cortex: A choline acetyltransferase study. J. Comp. Neurol. 1992, 318, 316–328. [Google Scholar] [CrossRef]
- Mesulam, M.-M.; Mufson, E.J.; Wainer, B.H.; Levey, A.I. Central cholinergic pathways in the rat: An overview based on an alternative nomenclature (Ch1–Ch6). Neuroscience 1983, 10, 1185–1201. [Google Scholar] [CrossRef]
- Metherate, R.; Cox, C.L.; Ashe, J.H. Cellular bases of neocortical activation—Modulation of neural oscillations by the nucleus basalis and endogenous acetylcholine. J. Neurosci. 1992, 12, 4701–4711. [Google Scholar] [CrossRef] [PubMed]
- Duque, A.; Balatoni, B.; Detari, L.; Zaborszky, L. EEG correlation of the discharge properties of identified neurons in the basal forebrain. J. Neurophysiol. 2000, 84, 1627–1635. [Google Scholar] [CrossRef] [PubMed]
- Celesia, G.G.; Jasper, H.H. Acetylcholine released from cerebral cortex in relation to state of activation. Neurology 1966, 16, 1053–1063. [Google Scholar] [CrossRef]
- Jasper, H.H.; Tessier, J. Acetylcholine liberation from cerebral cortex during paradoxical REM sleep. Science 1971, 172, 601–602. [Google Scholar] [CrossRef]
- Golmayo, L.; Nuñez, A.; Zaborszky, L. Electrophysiological evidence for the existence of a posterior cortical-prefrontal-basal forebrain circuitry in modulating sensory responses in visual and somatosensory rat cortical areas. Neuroscience 2003, 119, 597–609. [Google Scholar] [CrossRef]
- Fournier, G.N.; Semba, K.; Rasmusson, D.D. Modality- and region-specific acetylcholine release in the rat neocortex. Neuroscience 2004, 126, 257–262. [Google Scholar] [CrossRef]
- Semba, K. Multiple output pathways of the basal forebrain: Organization, chemical heterogeneity, and roles in vigilance. Behav. Brain Res. 2000, 115, 117–141. [Google Scholar] [CrossRef]
- Semba, K.; Fibiger, H.C. Organization of central cholinergic systems. Prog. Brain Res. 1989, 79, 37–63. [Google Scholar]
- Gombkoto, P.; Gielow, M.; Varsanyi, P.; Chavez, C.; Zaborszky, L. Contribution of the basal forebrain to corticocortical network interactions. Brain Struct. Funct. 2021, 226, 1803–1821. [Google Scholar] [CrossRef]
- Zaborszky, L.; Buhl, D.L.; Pobalashingham, S.; Bjaalie, J.G.; Nadasdy, Z. Three-dimensional chemoarchitecture of the basal forebrain: Spatially specific association of cholinergic and calcium binding protein-containing neurons. Neuroscience 2005, 136, 697–713. [Google Scholar] [CrossRef]
- Zaborszky, L.; Csordas, A.; Mosca, K.; Kim, J.; Gielow, M.R.; Vadasz, C.; Nadasdy, Z. Neurons in the Basal Forebrain Project to the Cortex in a Complex Topographic Organization that Reflects Corticocortical Connectivity Patterns: An Experimental Study Based on Retrograde Tracing and 3D Reconstruction. Cereb. Cortex 2015, 25, 118–137. [Google Scholar] [CrossRef] [PubMed]
- Rye, D.B.; Wainer, B.H.; Mesulam, M.M.; Mufson, E.J.; Saper, C.B. Cortical projections arising from the basal forebrain: A study of cholinergic and noncholinergic components employing combined retrograde tracing and immunohistochemical localization of choline acetyltransferase. Neuroscience 1984, 13, 627–643. [Google Scholar] [CrossRef] [PubMed]
- Freund, T.F.; Meskenaite, V. gamma-aminobutyric acid-containing basal forebrain neurons innervate inhibitory interneurons in the neocortex. Proc. Natl. Acad. Sci. USA 1992, 89, 738–742. [Google Scholar] [CrossRef] [PubMed]
- Henny, P.; Jones, B.E. Projections from basal forebrain to prefrontal cortex comprise cholinergic, GABAergic and glutamatergic inputs to pyramidal cells or interneurons. Eur. J. Neurosci. 2008, 27, 654–670. [Google Scholar] [CrossRef]
- Loopuijt, L.D.; Zahm, D.S. Synaptologic and fine structural features distinguishing a subset of basal forebrain cholinergic neurons embedded in the dense intrinsic fiber network of the caudal extended amygdala. J. Comp. Neurol. 2006, 498, 93–111. [Google Scholar] [CrossRef]
- Saper, C.B. Diffuse cortical projection systems: Anatomical organization and role in cortical function. In Handbook of Physiology: The Nervous System; Mountcastle, V.B., Plum, F., Geiger, S., Eds.; American Physiological Society: Bethesda, MD, USA, 1987; Volume 5, pp. 169–210. [Google Scholar]
- Woolf, N.J. Cholinergic systems in mammalian brain and spinal cord. Prog. Neurobiol. 1991, 37, 475–524. [Google Scholar] [CrossRef]
- Whalen, P.J.; Kapp, B.S.; Pascoe, J.P. Neuronal activity within the nucleus basalis and conditioned neocortical electroencephalographic activation. J. Neurosci. 1994, 14, 1623–1633. [Google Scholar] [CrossRef]
- Zaborszky, L. The modular organization of brain systems. Basal forebrain: The last frontier. Prog. Brain Res. 2002, 136, 359–372. [Google Scholar]
- Chaves-Coira, I.; Barros-Zulaica, N.; Rodrigo-Angulo, M.; Nunez, A. Modulation of Specific Sensory Cortical Areas by Segregated Basal Forebrain Cholinergic Neurons Demonstrated by Neuronal Tracing and Optogenetic Stimulation in Mice. Front. Neural Circuits 2016, 10, 28. [Google Scholar] [CrossRef]
- Chaves-Coira, I.; Martin-Cortecero, J.; Nunez, A.; Rodrigo-Angulo, M.L. Basal Forebrain Nuclei Display Distinct Projecting Pathways and Functional Circuits to Sensory Primary and Prefrontal Cortices in the Rat. Front. Neuroanat. 2018, 12, 69. [Google Scholar] [CrossRef]
- Yuan, R.; Biswal, B.B.; Zaborszky, L. Functional Subdivisions of Magnocellular Cell Groups in Human Basal Forebrain: Test-Retest Resting-State Study at Ultra-high Field, and Meta-analysis. Cereb. Cortex 2019, 29, 2844–2858. [Google Scholar] [CrossRef] [PubMed]
- Radnikow, G.; Feldmeyer, D. Layer- and Cell Type-Specific Modulation of Excitatory Neuronal Activity in the Neocortex. Front. Neuroanat. 2018, 12, 1. [Google Scholar] [CrossRef] [PubMed]
- Vanderheyden, P.; Gies, J.P.; Ebinger, G.; De Keyser, J.; Landry, Y.; Vauquelin, G. Human M1-, M2- and M3-muscarinic cholinergic receptors: Binding characteristics of agonists and antagonists. J. Neurol. Sci. 1990, 97, 67–80. [Google Scholar] [CrossRef]
- Scarr, E.; Seo, M.S.; Aumann, T.D.; Chana, G.; Everall, I.P.; Dean, B. The distribution of muscarinic M1 receptors in the human hippocampus. J. Chem. Neuroanat. 2016, 77, 187–192. [Google Scholar] [CrossRef] [PubMed]
- Obermayer, J.; Verhoog, M.B.; Luchicchi, A.; Mansvelder, H.D. Cholinergic Modulation of Cortical Microcircuits Is Layer-Specific: Evidence from Rodent, Monkey and Human Brain. Front. Neural Circuits 2017, 11, 100. [Google Scholar] [CrossRef] [PubMed]
- Haam, J.; Yakel, J.L. Cholinergic modulation of the hippocampal region and memory function. J. Neurochem. 2017, 142 (Suppl. S2), 111–121. [Google Scholar] [CrossRef] [PubMed]
- Chaves-Coira, I.; Rodrigo-Angulo, M.L.; Nunez, A. Bilateral Pathways from the Basal Forebrain to Sensory Cortices May Contribute to Synchronous Sensory Processing. Front. Neuroanat. 2018, 12, 5. [Google Scholar] [CrossRef] [PubMed]
- Gallagher, M.; Colombo, P.J. Ageing: The cholinergic hypothesis of cognitive decline. Curr. Opin. Neurobiol. 1995, 5, 161–168. [Google Scholar] [CrossRef]
- Ballinger, E.C.; Ananth, M.; Talmage, D.A.; Role, L.W. Basal Forebrain Cholinergic Circuits and Signaling in Cognition and Cognitive Decline. Neuron 2016, 91, 1199–1218. [Google Scholar] [CrossRef]
- Grothe, M.; Zaborszky, L.; Atienza, M.; Gil-Neciga, E.; Rodriguez-Romero, R.; Teipel, S.J.; Amunts, K.; Suarez-Gonzalez, A.; Cantero, J.L. Reduction of basal forebrain cholinergic system parallels cognitive impairment in patients at high risk of developing Alzheimer’s disease. Cereb. Cortex 2010, 20, 1685–1695. [Google Scholar] [CrossRef]
- Hasselmo, M.E. Neuromodulation and the hippocampus: Memory function and dysfunction in a network simulation. Prog. Brain Res. 1999, 121, 3–18. [Google Scholar] [PubMed]
- Dumas, J.A.; Newhouse, P.A. The cholinergic hypothesis of cognitive aging revisited again: Cholinergic functional compensation. Pharmacol. Biochem. Behav. 2011, 99, 254–261. [Google Scholar] [CrossRef] [PubMed]
- Buzsaki, G.; Bickford, R.G.; Ponomareff, G.; Yhal, L.J.; Mandel, R.; Gage, F.H. Nucleus basalis and thalamic control of neocortical activity in the freely moving rat. Neuroscience 1988, 8, 4007–4026. [Google Scholar] [CrossRef]
- Cape, E.G.; Jones, B.E. Differential modulation of high-frequency gamma-electroencephalogram activity and sleep-wake state by noradrenaline and serotonin microinjections into the region of cholinergic basalis neurons. J. Neurosci. Off. J. Soc. Neurosci. 1998, 18, 2653–2666. [Google Scholar] [CrossRef] [PubMed]
- Burk, J.A.; Sarter, M. Dissociation between the attentional functions mediated via basal forebrain cholinergic and gabaergic neurons. Neuroscience 2001, 105, 899–909. [Google Scholar] [CrossRef]
- Kaur, S.; Junek, A.; Black, M.A.; Semba, K. Effects of ibotenate and 192IgG-saporin lesions of the nucleus basalis magnocellularis/substantia innominata on spontaneous sleep and wake states and on recovery sleep after sleep deprivation in rats. J. Neurosci. 2008, 28, 491–504. [Google Scholar] [CrossRef]
- Alenda, A.; Nuñez, A. Cholinergic modulation of sensory interference in rat primary somatosensory cortical neurons. Brain Res. 2007, 1133, 158–167. [Google Scholar] [CrossRef]
- Decker, M.W. The effects of aging on hippocampal and cortical projections of the forebrain cholinergic system. Brain Res. 1987, 434, 423–438. [Google Scholar] [CrossRef]
- Fischer, W.; Gage, F.H.; Bjorklund, A. Degenerative Changes in Forebrain Cholinergic Nuclei Correlate with Cognitive Impairments in Aged Rats. Eur. J. Neurosci. 1989, 1, 34–45. [Google Scholar] [CrossRef] [PubMed]
- Han, S.H.; McCool, B.A.; Murchison, D.; Nahm, S.S.; Parrish, A.R.; Griffith, W.H. Single-cell RT-PCR detects shifts in mRNA expression profiles of basal forebrain neurons during aging. Brain Res. Mol. Brain Res. 2002, 98, 67–80. [Google Scholar] [CrossRef]
- Brandeis, D.; Naylor, H.; Halliday, R.; Callaway, E.; Yano, L. Scopolamine effects on visual information processing, attention, and event-related potential map latencies. Psychophysiology 1992, 29, 315–336. [Google Scholar] [CrossRef]
- Hernandez, C.M.; Gearhart, D.A.; Parikh, V.; Hohnadel, E.J.; Davis, L.W.; Middlemore, M.L.; Warsi, S.P.; Waller, J.L.; Terry, A.V., Jr. Comparison of galantamine and donepezil for effects on nerve growth factor, cholinergic markers, and memory performance in aged rats. J. Pharmacol. Exp. Ther. 2006, 316, 679–694. [Google Scholar] [CrossRef]
- Aubert, I.; Rowe, W.; Meaney, M.J.; Gauthier, S.; Quirion, R. Cholinergic markers in aged cognitively impaired Long-Evans rats. Neuroscience 1995, 67, 277–292. [Google Scholar] [CrossRef]
- Stanley, E.M.; Fadel, J.R.; Mott, D.D. Interneuron loss reduces dendritic inhibition and GABA release in hippocampus of aged rats. Neurobiol. Aging 2012, 33, 431.e1. [Google Scholar] [CrossRef]
- Griffith, W.H.; Dubois, D.W.; Fincher, A.; Peebles, K.A.; Bizon, J.L.; Murchison, D. Characterization of age-related changes in synaptic transmission onto F344 rat basal forebrain cholinergic neurons using a reduced synaptic preparation. J. Neurophysiol. 2014, 111, 273–286. [Google Scholar] [CrossRef]
- Griffith, W.H.; Murchison, D.A. Enhancement of GABA-activated membrane currents in aged Fischer 344 rat basal forebrain neurons. J. Neurosci. 1995, 15, 2407–2416. [Google Scholar] [CrossRef]
- Lamour, Y.; Dutar, P.; Jobert, A. Septo-hippocampal neurons: Alterated properties in the aged rat. Brain Res. 1987, 416, 277–282. [Google Scholar] [CrossRef]
- Murchison, D.; Griffith, W.H. Low-voltage activated calcium currents increase in basal forebrain neurons from aged rats. J. Neurophysiol. 1995, 74, 876–887. [Google Scholar] [CrossRef]
- Murchison, D.; Griffith, W.H. High-voltage-activated calcium currents in basal forebrain neurons during aging. J. Neurophysiol. 1996, 76, 158–174. [Google Scholar] [CrossRef]
- Griffith, W.H.; Jasek, M.C.; Bain, S.H.; Murchison, D. Modification of ion channels and calcium homeostasis of basal forebrain neurons during aging. Behav. Brain Res. 2000, 115, 219–233. [Google Scholar] [CrossRef]
- Jasek, M.C.; Griffith, W.H. Pharmacological characterization of ionotropic excitatory amino acid receptors in young and aged rat basal forebrain. Neuroscience 1998, 82, 1179–1194. [Google Scholar] [CrossRef] [PubMed]
- Murchison, D.; Zawieja, D.C.; Griffith, W.H. Reduced mitochondrial buffering of voltage-gated calcium influx in aged rat basal forebrain neurons. Cell Calcium 2004, 36, 61–75. [Google Scholar] [CrossRef]
- Purdon, P.L.; Pavone, K.J.; Akeju, O.; Smith, A.C.; Sampson, A.L.; Lee, J.; Zhou, D.W.; Solt, K.; Brown, E.N. The Ageing Brain: Age-dependent changes in the electroencephalogram during propofol and sevoflurane general anaesthesia. Br. J. Anaesth. 2015, 115 (Suppl. S1), i46–i57. [Google Scholar] [CrossRef] [PubMed]
- Klimesch, W. EEG alpha and theta oscillations reflect cognitive and memory performance: A review and analysis. Brain Res. Rev. 1999, 29, 169–195. [Google Scholar] [CrossRef]
- Nunez, A.; Buno, W. The Theta Rhythm of the Hippocampus: From Neuronal and Circuit Mechanisms to Behavior. Front. Cell. Neurosci. 2021, 15, 649262. [Google Scholar] [CrossRef] [PubMed]
- Wimmer, M.E.; Rising, J.; Galante, R.J.; Wyner, A.; Pack, A.I.; Abel, T. Aging in mice reduces the ability to sustain sleep/wake states. PLoS ONE 2013, 8, e81880. [Google Scholar] [CrossRef]
- Cantero, J.L.; Atienza, M.; Madsen, J.R.; Stickgold, R. Gamma EEG dynamics in neocortex and hippocampus during human wakefulness and sleep. Neuroimage 2004, 22, 1271–1280. [Google Scholar] [CrossRef]
- Cantero, J.L.; Atienza, M.; Stickgold, R.; Kahana, M.J.; Madsen, J.R.; Kocsis, B. Sleep-dependent theta oscillations in the human hippocampus and neocortex. J. Neurosci. Off. J. Soc. Neurosci. 2003, 23, 10897–10903. [Google Scholar] [CrossRef]
- Buzsaki, G.; Draguhn, A. Neuronal oscillations in cortical networks. Science 2004, 304, 1926–1929. [Google Scholar] [CrossRef]
- Colgin, L.L. Rhythms of the hippocampal network. Nat. Rev. Neurosci. 2016, 17, 239–249. [Google Scholar] [CrossRef]
- Colgin, L.L.; Denninger, T.; Fyhn, M.; Hafting, T.; Bonnevie, T.; Jensen, O.; Moser, M.B.; Moser, E.I. Frequency of gamma oscillations routes flow of information in the hippocampus. Nature 2009, 462, 353–357. [Google Scholar] [CrossRef] [PubMed]
- Colgin, L.L.; Moser, E.I. Gamma oscillations in the hippocampus. Physiology 2010, 25, 319–329. [Google Scholar] [CrossRef] [PubMed]
- Lisman, J.; Buzsaki, G.; Eichenbaum, H.; Nadel, L.; Ranganath, C.; Redish, A.D. Viewpoints: How the hippocampus contributes to memory, navigation and cognition. Nat. Neurosci. 2017, 20, 1434–1447. [Google Scholar] [CrossRef] [PubMed]
- Jacobson, T.K.; Howe, M.D.; Schmidt, B.; Hinman, J.R.; Escabi, M.A.; Markus, E.J. Hippocampal theta, gamma, and theta-gamma coupling: Effects of aging, environmental change, and cholinergic activation. J. Neurophysiol. 2013, 109, 1852–1865. [Google Scholar] [CrossRef] [PubMed]
- Jacobson, T.K.; Schmidt, B.; Hinman, J.R.; Escabi, M.A.; Markus, E.J. Age-related decrease in theta and gamma coherence across dorsal ca1 pyramidale and radiatum layers. Hippocampus 2015, 25, 1327–1335. [Google Scholar] [CrossRef]
- Brun, V.H.; Otnass, M.K.; Molden, S.; Steffenach, H.A.; Witter, M.P.; Moser, M.B.; Moser, E.I. Place cells and place recognition maintained by direct entorhinal-hippocampal circuitry. Science 2002, 296, 2243–2246. [Google Scholar] [CrossRef]
- Hafting, T.; Fyhn, M.; Molden, S.; Moser, M.B.; Moser, E.I. Microstructure of a spatial map in the entorhinal cortex. Nature 2005, 436, 801–806. [Google Scholar] [CrossRef]
- Tort, A.B.; Kramer, M.A.; Thorn, C.; Gibson, D.J.; Kubota, Y.; Graybiel, A.M.; Kopell, N.J. Dynamic cross-frequency couplings of local field potential oscillations in rat striatum and hippocampus during performance of a T-maze task. Proc. Natl. Acad. Sci. USA 2008, 105, 20517–20522. [Google Scholar] [CrossRef]
- Yordanova, J.; Falkenstein, M.; Kolev, V. Aging-related changes in motor response-related theta activity. Int. J. Psychophysiol. 2020, 153, 95–106. [Google Scholar] [CrossRef]
- Karlsson, A.E.; Lindenberger, U.; Sander, M.C. Out of Rhythm: Compromised Precision of Theta-Gamma Coupling Impairs Associative Memory in Old Age. J. Neurosci. 2022, 42, 1752–1764. [Google Scholar] [CrossRef]
- Mukherjee, A.; Wilson, E.M.; Rotwein, P. Insulin-like growth factor (IGF) binding protein-5 blocks skeletal muscle differentiation by inhibiting IGF actions. Mol. Endocrinol. 2008, 22, 206–215. [Google Scholar] [CrossRef] [PubMed]
- Gross, S.M.; Rotwein, P. Unraveling Growth Factor Signaling and Cell Cycle Progression in Individual Fibroblasts. J. Biol. Chem. 2016, 291, 14628–14638. [Google Scholar] [CrossRef]
- Torres-Aleman, I. Role of insulin-like growth factors in neuronal plasticity and neuroprotection. Adv. Exp. Med. Biol. 2005, 567, 243–258. [Google Scholar] [PubMed]
- Ishii, D.N.; Guertin, D.M.; Whalen, L.R. Reduced insulin-like growth factor-I mRNA content in liver, adrenal glands and spinal cord of diabetic rats. Diabetologia 1994, 37, 1073–1081. [Google Scholar] [CrossRef]
- Dyer, A.H.; Vahdatpour, C.; Sanfeliu, A.; Tropea, D. The role of Insulin-Like Growth Factor 1 (IGF-1) in brain development, maturation and neuroplasticity. Neuroscience 2016, 325, 89–99. [Google Scholar] [CrossRef] [PubMed]
- Al-Samerria, S.; Radovick, S. The Role of Insulin-like Growth Factor-1 (IGF-1) in the Control of Neuroendocrine Regulation of Growth. Cells 2021, 10, 2664. [Google Scholar] [CrossRef]
- Zegarra-Valdivia, J.A.; Chaves-Coira, I.; Fernandez de Sevilla, M.E.; Martinez-Rachadell, L.; Esparza, J.; Torres-Aleman, I.; Nunez, A. Reduced Insulin-Like Growth Factor-I Effects in the Basal Forebrain of Aging Mouse. Front. Aging Neurosci. 2021, 13, 682388. [Google Scholar] [CrossRef]
- Garcia-Magro, N.; Zegarra-Valdivia, J.A.; Troyas-Martinez, S.; Torres-Aleman, I.; Nunez, A. Response Facilitation Induced by Insulin-like Growth Factor-I in the Primary Somatosensory Cortex of Mice Was Reduced in Aging. Cells 2022, 11, 717. [Google Scholar] [CrossRef]
- Barros-Zulaica, N.; Villa, A.E.P.; Nunez, A. Response Adaptation in Barrel Cortical Neurons Facilitates Stimulus Detection during Rhythmic Whisker Stimulation in Anesthetized Mice. eNeuro 2019, 6, ENEURO.0471-18.2019. [Google Scholar] [CrossRef]
- Carro, E.; Nuñez, A.; Busiguina, S.; Torres-Aleman, I. Circulating insulin-like growth factor I mediates effects of exercise on the brain. J. Neurosci. 2000, 20, 2926–2933. [Google Scholar] [CrossRef]
- Gazit, N.; Vertkin, I.; Shapira, I.; Helm, M.; Slomowitz, E.; Sheiba, M.; Mor, Y.; Rizzoli, S.; Slutsky, I. IGF-1 Receptor Differentially Regulates Spontaneous and Evoked Transmission via Mitochondria at Hippocampal Synapses. Neuron 2016, 89, 583–597. [Google Scholar] [CrossRef] [PubMed]
- Nuñez, A.; Carro, E.; Torres-Aleman, I. Insulin-like growth factor I modifies electrophysiological properties of rat brain stem neurons. J. Neurophysiol. 2003, 89, 3008–3017. [Google Scholar] [CrossRef] [PubMed]
- Nilsson, L.; Sara, V.R.; Nordberg, A. Insulin-like growth factor 1 stimulates the release of acetylcholine from rat cortical slices. Neurosci. Lett. 1988, 88, 221–226. [Google Scholar] [CrossRef]
- Araujo, D.M.; Lapchak, P.A.; Collier, B.; Chabot, J.G.; Quirion, R. Insulin-like growth factor-1 (somatomedin-C) receptors in the rat brain: Distribution and interaction with the hippocampal cholinergic system. Brain Res. 1989, 484, 130–138. [Google Scholar] [CrossRef] [PubMed]
- Castro-Alamancos, M.A.; Torres-Alemán, I. Learning of the conditioned eye-blink response is impaired by an antisense insuline-like growth factor I oligonucleotide. Proc. Natl. Acad. Sci. USA 1994, 91, 10203–10207. [Google Scholar] [CrossRef]
- Seto, D.; Zheng, W.H.; McNicoll, A.; Collier, B.; Quirion, R.; Kar, S. Insulin-like growth factor-I inhibits endogenous acetylcholine release from the rat hippocampal formation: Possible involvement of GABA in mediating the effects. Neuroscience 2002, 115, 603–612. [Google Scholar] [CrossRef]
- Maya-Vetencourt, J.F.; Baroncelli, L.; Viegi, A.; Tiraboschi, E.; Castren, E.; Cattaneo, A.; Maffei, L. IGF-1 restores visual cortex plasticity in adult life by reducing local GABA levels. Neural Plast. 2012, 2012, 250421. [Google Scholar] [CrossRef]
- Castro-Alamancos, M.A.; Torres-Aleman, I. Long-term depression of glutamate-induced gamma-aminobutyric acid release in cerebellum by insulin-like growth factor I. Proc. Natl. Acad. Sci. USA 1993, 90, 7386–7390. [Google Scholar] [CrossRef]
- Fernandez, A.M.; Torres-Aleman, I. The many faces of insulin-like peptide signalling in the brain. Nat. Rev. Neurosci. 2012, 13, 225–239. [Google Scholar] [CrossRef]
- Pardo, J.; Abba, M.C.; Lacunza, E.; Ogundele, O.M.; Paiva, I.; Morel, G.R.; Outeiro, T.F.; Goya, R.G. IGF-I Gene Therapy in Aging Rats Modulates Hippocampal Genes Relevant to Memory Function. J. Gerontol. A Biol. Sci. Med. Sci. 2018, 73, 459–467. [Google Scholar] [CrossRef]
- Ogundele, O.M.; Pardo, J.; Francis, J.; Goya, R.G.; Lee, C.C. A Putative Mechanism of Age-Related Synaptic Dysfunction Based on the Impact of IGF-1 Receptor Signaling on Synaptic CaMKIIalpha Phosphorylation. Front. Neuroanat. 2018, 12, 35. [Google Scholar] [CrossRef] [PubMed]
- Noriega-Prieto, J.A.; Maglio, L.E.; Zegarra-Valdivia, J.A.; Pignatelli, J.; Fernandez, A.M.; Martinez-Rachadell, L.; Fernandes, J.; Nunez, A.; Araque, A.; Torres-Aleman, I.; et al. Astrocytic IGF-IRs Induce Adenosine-Mediated Inhibitory Downregulation and Improve Sensory Discrimination. J. Neurosci. 2021, 41, 4768–4781. [Google Scholar] [CrossRef] [PubMed]
- Junnila, R.K.; List, E.O.; Berryman, D.E.; Murrey, J.W.; Kopchick, J.J. The GH/IGF-1 axis in ageing and longevity. Nat. Rev. Endocrinol. 2013, 9, 366–376. [Google Scholar] [CrossRef] [PubMed]
- Lee, C.H.; Ahn, J.H.; Park, J.H.; Yan, B.C.; Kim, I.H.; Lee, D.H.; Cho, J.H.; Chen, B.H.; Lee, J.C.; Cho, J.H.; et al. Decreased insulin-like growth factor-I and its receptor expression in the hippocampus and somatosensory cortex of the aged mouse. Neurochem. Res. 2014, 39, 770–776. [Google Scholar] [CrossRef]
- Muller, A.P.; Fernandez, A.M.; Haas, C.; Zimmer, E.; Portela, L.V.; Torres-Aleman, I. Reduced brain insulin-like growth factor I function during aging. Mol. Cell. Neurosci. 2012, 49, 9–12. [Google Scholar] [CrossRef]
- Ashpole, N.M.; Sanders, J.E.; Hodges, E.L.; Yan, H.; Sonntag, W.E. Growth hormone, insulin-like growth factor-1 and the aging brain. Exp. Gerontol. 2015, 68, 76–81. [Google Scholar] [CrossRef]
- Trejo, J.L.; Carro, E.; Lopez-Lopez, C.; Torres-Aleman, I. Role of serum insulin-like growth factor I in mammalian brain aging. Growth Horm. IGF Res. 2004, 14 (Suppl. A), S39–S43. [Google Scholar] [CrossRef]
- Piriz, J.; Muller, A.; Trejo, J.L.; Torres-Aleman, I. IGF-I and the aging mammalian brain. Exp. Gerontol. 2011, 46, 96–99. [Google Scholar] [CrossRef]
- Szymusiak, R. Magnocellular nuclei of the basal forebrain: Substrates of sleep and arousal regulation. Sleep 1995, 18, 478–500. [Google Scholar] [CrossRef]
- Jones, B.E. From waking to sleeping: Neuronal and chemical substrates. Trends Pharmacol. Sci. 2005, 26, 578–586. [Google Scholar] [CrossRef]
- Xu, M.; Chung, S.; Zhang, S.; Zhong, P.; Ma, C.; Chang, W.C.; Weissbourd, B.; Sakai, N.; Luo, L.; Nishino, S.; et al. Basal forebrain circuit for sleep-wake control. Nat. Neurosci. 2015, 18, 1641–1647. [Google Scholar] [CrossRef]
- Landolt, H.P.; Dijk, D.J.; Achermann, P.; Borbely, A.A. Effect of age on the sleep EEG: Slow-wave activity and spindle frequency activity in young and middle-aged men. Brain Res. 1996, 738, 205–212. [Google Scholar] [CrossRef]
- Luca, G.; Haba Rubio, J.; Andries, D.; Tobback, N.; Vollenweider, P.; Waeber, G.; Marques Vidal, P.; Preisig, M.; Heinzer, R.; Tafti, M. Age and gender variations of sleep in subjects without sleep disorders. Ann. Med. 2015, 47, 482–491. [Google Scholar] [CrossRef] [PubMed]
- Mander, B.A.; Winer, J.R.; Walker, M.P. Sleep and Human Aging. Neuron 2017, 94, 19–36. [Google Scholar] [CrossRef] [PubMed]
- Hasan, S.; Dauvilliers, Y.; Mongrain, V.; Franken, P.; Tafti, M. Age-related changes in sleep in inbred mice are genotype dependent. Neurobiol. Aging 2012, 33, 195.e13. [Google Scholar] [CrossRef] [PubMed]
- Panagiotou, M.; Vyazovskiy, V.V.; Meijer, J.H.; Deboer, T. Differences in electroencephalographic non-rapid-eye movement sleep slow-wave characteristics between young and old mice. Sci. Rep. 2017, 7, 43656. [Google Scholar] [CrossRef] [PubMed]
- Panagiotou, M.; Papagiannopoulos, K.; Rohling, J.H.T.; Meijer, J.H.; Deboer, T. How Old Is Your Brain? Slow-Wave Activity in Non-rapid-eye-movement Sleep as a Marker of Brain Rejuvenation After Long-Term Exercise in Mice. Front. Aging Neurosci. 2018, 10, 233. [Google Scholar] [CrossRef]
- McKillop, L.E.; Fisher, S.P.; Cui, N.; Peirson, S.N.; Foster, R.G.; Wafford, K.A.; Vyazovskiy, V.V. Effects of Aging on Cortical Neural Dynamics and Local Sleep Homeostasis in Mice. J. Neurosci. 2018, 38, 3911–3928. [Google Scholar] [CrossRef]
- Carrier, J.; Viens, I.; Poirier, G.; Robillard, R.; Lafortune, M.; Vandewalle, G.; Martin, N.; Barakat, M.; Paquet, J.; Filipini, D. Sleep slow wave changes during the middle years of life. Eur. J. Neurosci. 2011, 33, 758–766. [Google Scholar] [CrossRef]
- Steriade, M.; Nuñez, A.; Amzica, F. A novel slow (<1 Hz) oscillation of neocortical neurons in vivo—Depolarizing and hyperpolarizing components. J. Neurosci. 1993, 13, 3252–3265. [Google Scholar]
- Shaw, M.E.; Sachdev, P.S.; Anstey, K.J.; Cherbuin, N. Age-related cortical thinning in cognitively healthy individuals in their 60s: The PATH Through Life study. Neurobiol. Aging 2016, 39, 202–209. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Schwam, J.; Chen, Q. Senescence-Associated Cell Transition and Interaction (SACTAI): A Proposed Mechanism for Tissue Aging, Repair, and Degeneration. Cells 2022, 11, 1089. [Google Scholar] [CrossRef] [PubMed]
- Whitehouse, P.J.; Price, D.L.; Clark, A.W.; Coyle, J.T.; DeLong, M.R. Alzheimer disease: Evidence for selective loss of cholinergic neurons in the nucleus basalis. Ann. Neurol. 1981, 10, 122–126. [Google Scholar] [CrossRef] [PubMed]
- McGeer, P.L.; McGeer, E.G.; Suzuki, J.; Dolman, C.E.; Nagai, T. Aging, Alzheimer’s disease, and the cholinergic system of the basal forebrain. Neurology 1984, 34, 741–745. [Google Scholar] [CrossRef] [PubMed]
- Arendt, T.; Bigl, V.; Tennstedt, A.; Arendt, A. Neuronal loss in different parts of the nucleus basalis is related to neuritic plaque formation in cortical target areas in Alzheimer’s disease. Neuroscience 1985, 14, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Vogels, O.J.; Broere, C.A.; ter Laak, H.J.; ten Donkelaar, H.J.; Nieuwenhuys, R.; Schulte, B.P. Cell loss and shrinkage in the nucleus basalis Meynert complex in Alzheimer’s disease. Neurobiol. Aging 1990, 11, 3–13. [Google Scholar] [CrossRef]
- Nagai, T.; McGeer, P.L.; Peng, J.H.; McGeer, E.G.; Dolman, C.E. Choline acetyltransferase immunohistochemistry in brains of Alzheimer’s disease patients and controls. Neurosci. Lett. 1983, 36, 195–199. [Google Scholar] [CrossRef]
- Hasselmo, M.E. Neuromodulation: Acetylcholine and memory consolidation. Trends Cogn. Sci. 1999, 3, 351–359. [Google Scholar] [CrossRef]
- Blennow, K.; Hampel, H. CSF markers for incipient Alzheimer’s disease. Lancet Neurol. 2003, 2, 605–613. [Google Scholar] [CrossRef]
- Hampel, H.; Blennow, K.; Shaw, L.M.; Hoessler, Y.C.; Zetterberg, H.; Trojanowski, J.Q. Total and phosphorylated tau protein as biological markers of Alzheimer’s disease. Exp. Gerontol. 2010, 45, 30–40. [Google Scholar] [CrossRef]
- Burger nee Buch, K.; Padberg, F.; Nolde, T.; Teipel, S.J.; Stubner, S.; Haslinger, A.; Schwarz, M.J.; Sunderland, T.; Arai, H.; Rapoport, S.I.; et al. Cerebrospinal fluid tau protein shows a better discrimination in young old (<70 years) than in old old patients with Alzheimer’s disease compared with controls. Neurosci. Lett. 1999, 277, 21–24. [Google Scholar] [CrossRef] [PubMed]
- Rubio, A.; Perez, M.; de Lecea, L.; Avila, J. Effect of cortistatin on tau phosphorylation at Ser262 site. J. Neurosci. Res. 2008, 86, 2462–2475. [Google Scholar] [CrossRef] [PubMed]
- Cantero, J.L.; Moreno-Lopez, B.; Portillo, F.; Rubio, A.; Hita-Yanez, E.; Avila, J. Role of tau protein on neocortical and hippocampal oscillatory patterns. Hippocampus 2011, 21, 827–834. [Google Scholar] [CrossRef] [PubMed]
- Llorens-Martin, M.; Blazquez-Llorca, L.; Benavides-Piccione, R.; Rabano, A.; Hernandez, F.; Avila, J.; DeFelipe, J. Selective alterations of neurons and circuits related to early memory loss in Alzheimer’s disease. Front. Neuroanat. 2014, 8, 38. [Google Scholar] [CrossRef] [PubMed]
- Cantero, J.L.; Atienza, M.; Ramos-Cejudo, J.; Fossati, S.; Wisniewski, T.; Osorio, R.S. Plasma tau predicts cerebral vulnerability in aging. Aging 2020, 12, 21004–21022. [Google Scholar] [CrossRef]
- Haass, C.; Schlossmacher, M.G.; Hung, A.Y.; Vigo-Pelfrey, C.; Mellon, A.; Ostaszewski, B.L.; Lieberburg, I.; Koo, E.H.; Schenk, D.; Teplow, D.B.; et al. Amyloid beta-peptide is produced by cultured cells during normal metabolism. Nature 1992, 359, 322–325. [Google Scholar] [CrossRef]
- Haass, C.; Koo, E.H.; Mellon, A.; Hung, A.Y.; Selkoe, D.J. Targeting of cell-surface beta-amyloid precursor protein to lysosomes: Alternative processing into amyloid-bearing fragments. Nature 1992, 357, 500–503. [Google Scholar] [CrossRef]
- Baker-Nigh, A.; Vahedi, S.; Davis, E.G.; Weintraub, S.; Bigio, E.H.; Klein, W.L.; Geula, C. Neuronal amyloid-beta accumulation within cholinergic basal forebrain in ageing and Alzheimer’s disease. Brain 2015, 138, 1722–1737. [Google Scholar] [CrossRef]
- Marks, S.M.; Lockhart, S.N.; Baker, S.L.; Jagust, W.J. Tau and beta-Amyloid Are Associated with Medial Temporal Lobe Structure, Function, and Memory Encoding in Normal Aging. J. Neurosci. 2017, 37, 3192–3201. [Google Scholar] [CrossRef]
- Lesne, S.E.; Sherman, M.A.; Grant, M.; Kuskowski, M.; Schneider, J.A.; Bennett, D.A.; Ashe, K.H. Brain amyloid-beta oligomers in ageing and Alzheimer’s disease. Brain 2013, 136, 1383–1398. [Google Scholar] [CrossRef]
- Burrinha, T.; Guimas Almeida, C. Aging impact on amyloid precursor protein neuronal trafficking. Curr. Opin. Neurobiol. 2022, 73, 102524. [Google Scholar] [CrossRef] [PubMed]
- McKinney, M.; Jacksonville, M.C. Brain cholinergic vulnerability: Relevance to behavior and disease. Biochem. Pharmacol. 2005, 70, 1115–1124. [Google Scholar] [CrossRef] [PubMed]
- Geula, C.; Nagykery, N.; Nicholas, A.; Wu, C.K. Cholinergic neuronal and axonal abnormalities are present early in aging and in Alzheimer disease. J. Neuropathol. Exp. Neurol. 2008, 67, 309–318. [Google Scholar] [CrossRef] [PubMed]
- Gouras, G.K.; Tsai, J.; Naslund, J.; Vincent, B.; Edgar, M.; Checler, F.; Greenfield, J.P.; Haroutunian, V.; Buxbaum, J.D.; Xu, H.; et al. Intraneuronal Abeta42 accumulation in human brain. Am. J. Pathol. 2000, 156, 15–20. [Google Scholar] [CrossRef]
- Geula, C.; Nagykery, N.; Wu, C.K.; Bu, J. Loss of calbindin-D28K from aging human cholinergic basal forebrain: Relation to plaques and tangles. J. Neuropathol. Exp. Neurol. 2003, 62, 605–616. [Google Scholar] [CrossRef]
- Riascos, D.; de Leon, D.; Baker-Nigh, A.; Nicholas, A.; Yukhananov, R.; Bu, J.; Wu, C.K.; Geula, C. Age-related loss of calcium buffering and selective neuronal vulnerability in Alzheimer’s disease. Acta Neuropathol. 2011, 122, 565–576. [Google Scholar] [CrossRef]


Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chaves-Coira, I.; García-Magro, N.; Zegarra-Valdivia, J.; Torres-Alemán, I.; Núñez, Á. Cognitive Deficits in Aging Related to Changes in Basal Forebrain Neuronal Activity. Cells 2023, 12, 1477. https://doi.org/10.3390/cells12111477
Chaves-Coira I, García-Magro N, Zegarra-Valdivia J, Torres-Alemán I, Núñez Á. Cognitive Deficits in Aging Related to Changes in Basal Forebrain Neuronal Activity. Cells. 2023; 12(11):1477. https://doi.org/10.3390/cells12111477
Chicago/Turabian StyleChaves-Coira, Irene, Nuria García-Magro, Jonathan Zegarra-Valdivia, Ignacio Torres-Alemán, and Ángel Núñez. 2023. "Cognitive Deficits in Aging Related to Changes in Basal Forebrain Neuronal Activity" Cells 12, no. 11: 1477. https://doi.org/10.3390/cells12111477