
Calmodulin as a Key Regulator of Exosomal Signal Peptides
 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Cells
2.2. Isolation of Exosomes from Conditioned Medium
2.3. Measurement of SEAP Activity
2.4. Nanoparticle Tracking Analysis (NTA)
2.5. Western Blotting
2.6. Immunocytochemical Staining
2.7. Immunoprecipitation
2.8. MALDI-TOF-MS
2.9. Statistical Analysis
3. Results
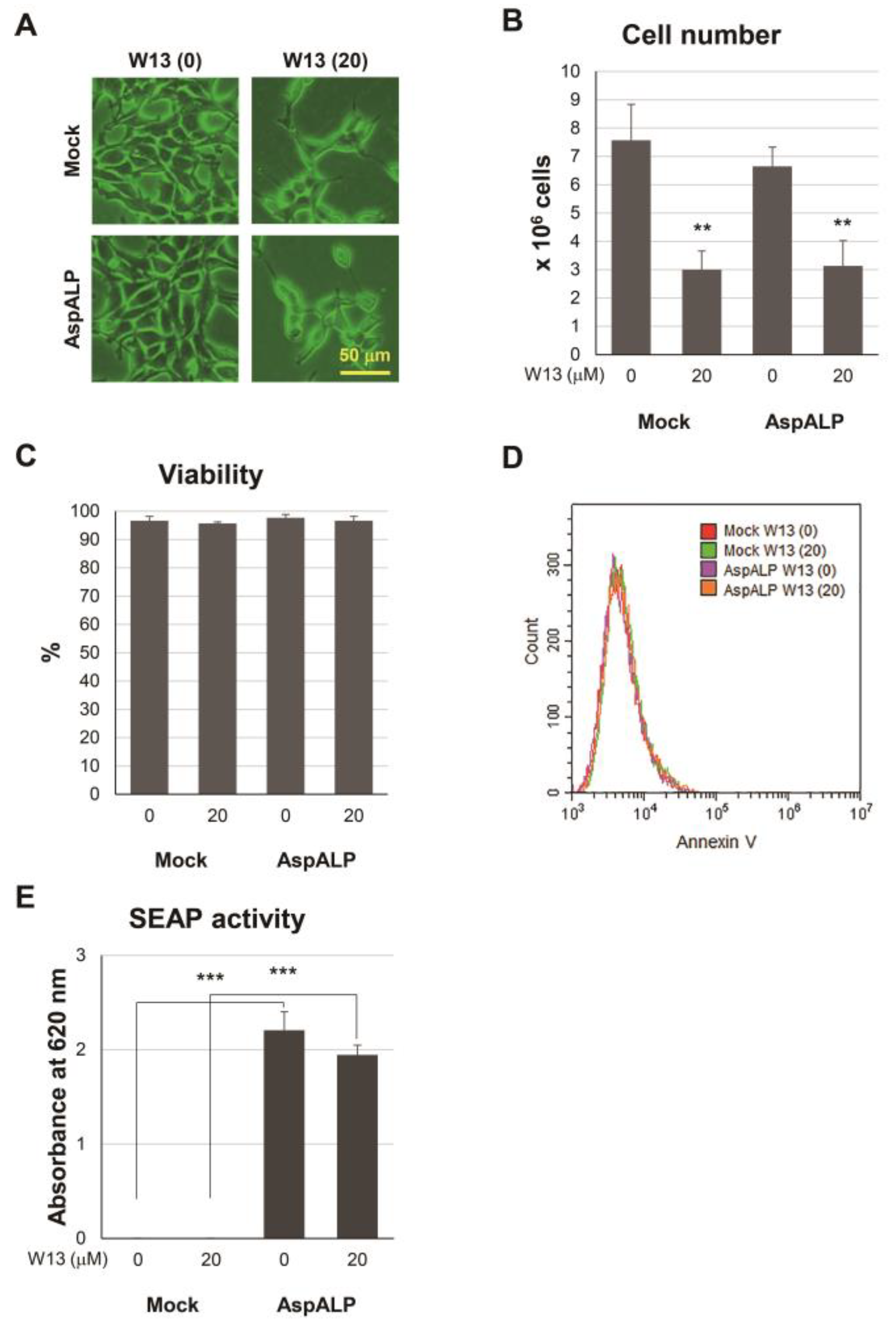
3.1. Effects of W13, a CaM Inhibitor, on T-REx Mock and T-REx AspALP Cells
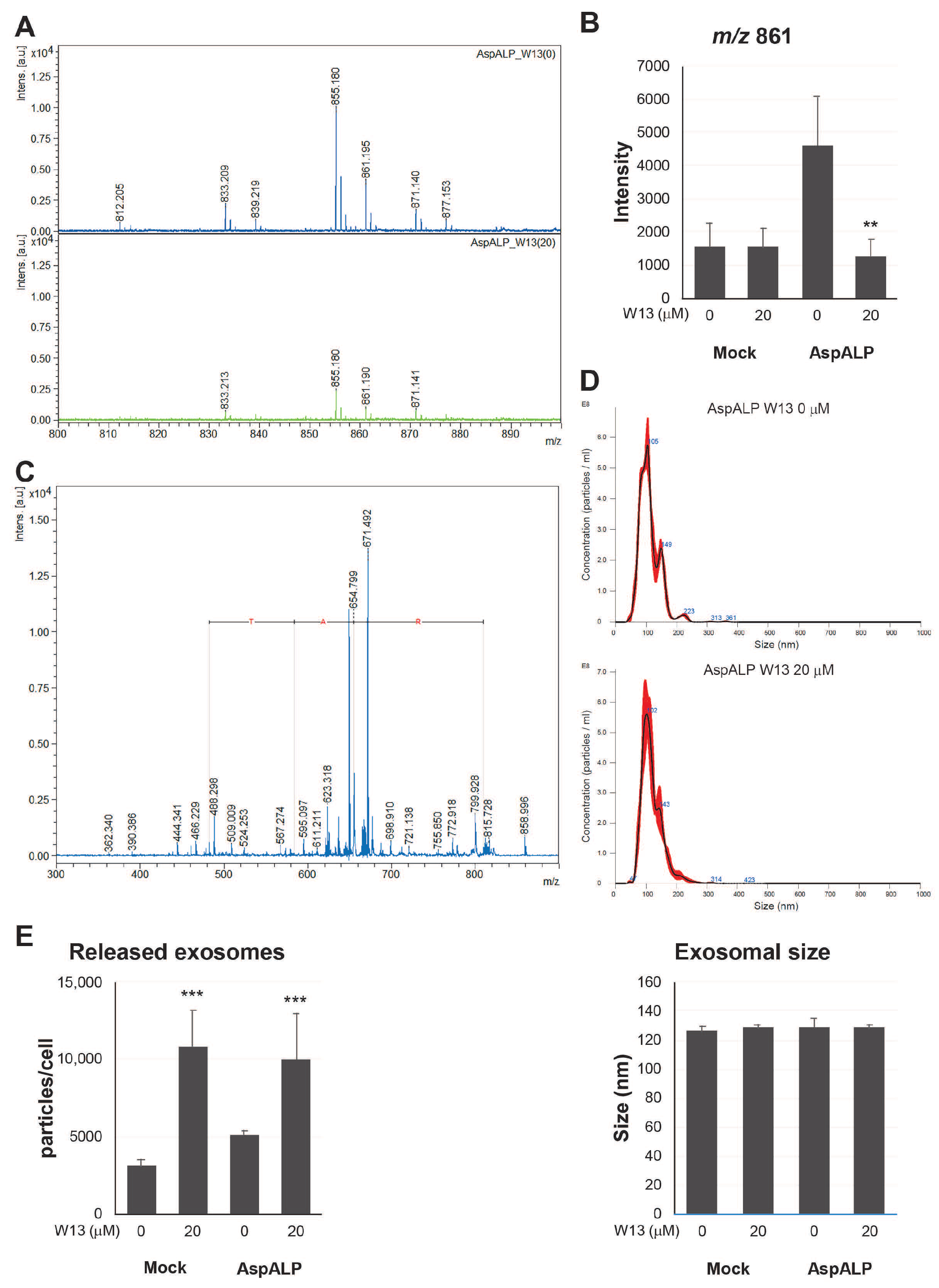
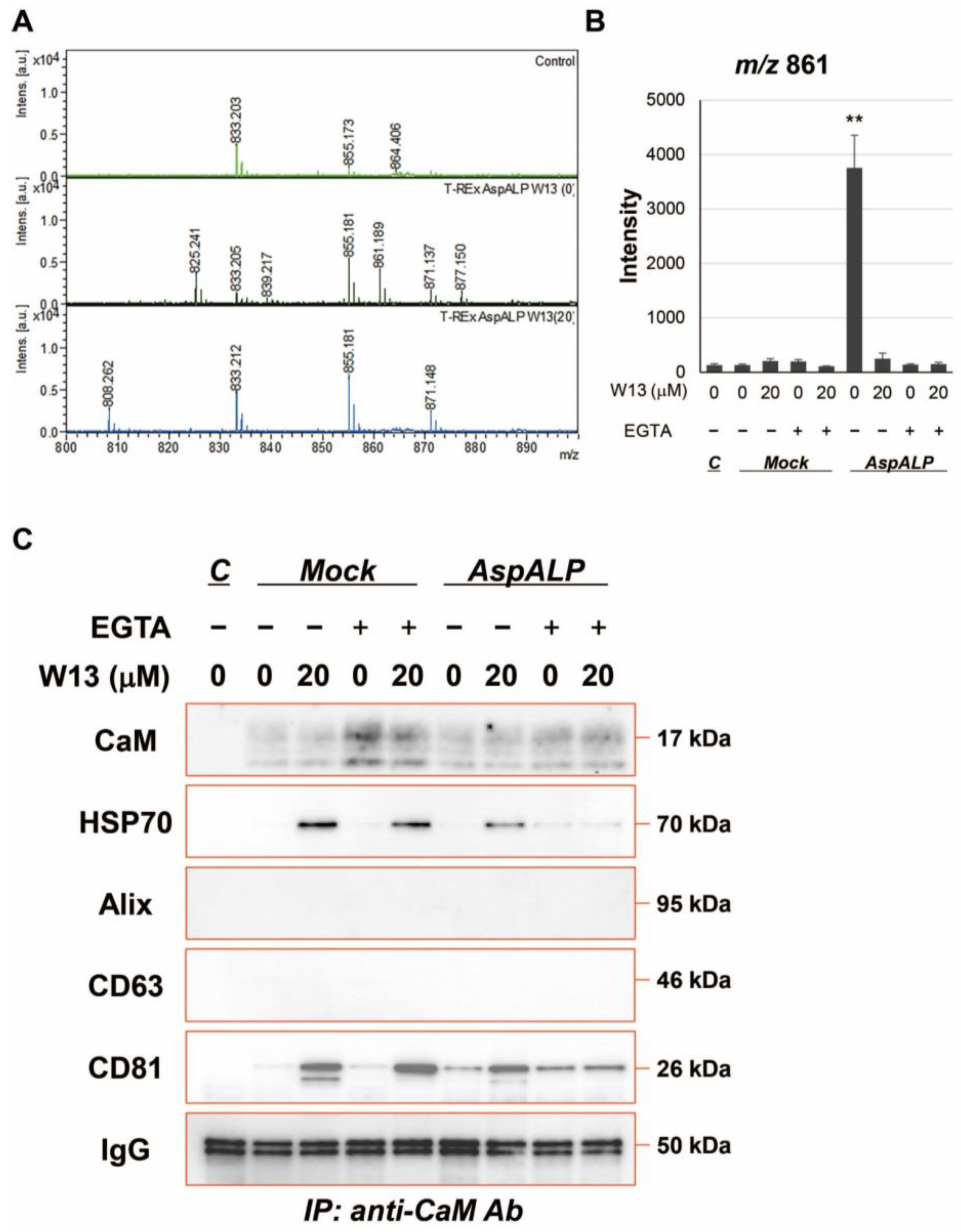
3.2. Effects of W13 on the Signal Peptide Content in Exosomes from T-REx AspALP Cells
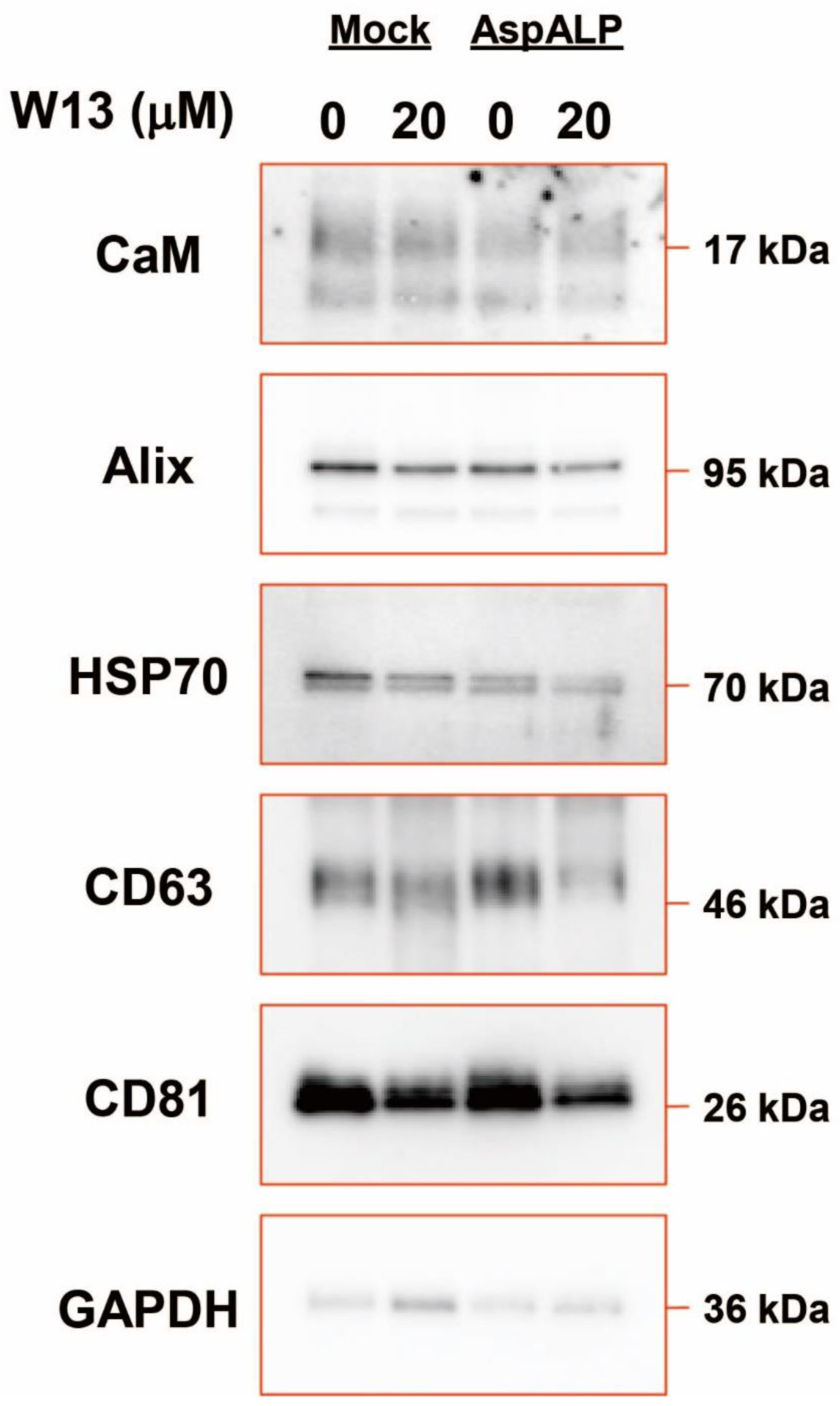
3.3. Changes in Exosomal Cargo Protein Abundance in T-REx Cells in the Presence or Absence of W13
3.4. Cargo Proteins in Exosomes from T-REx Cells
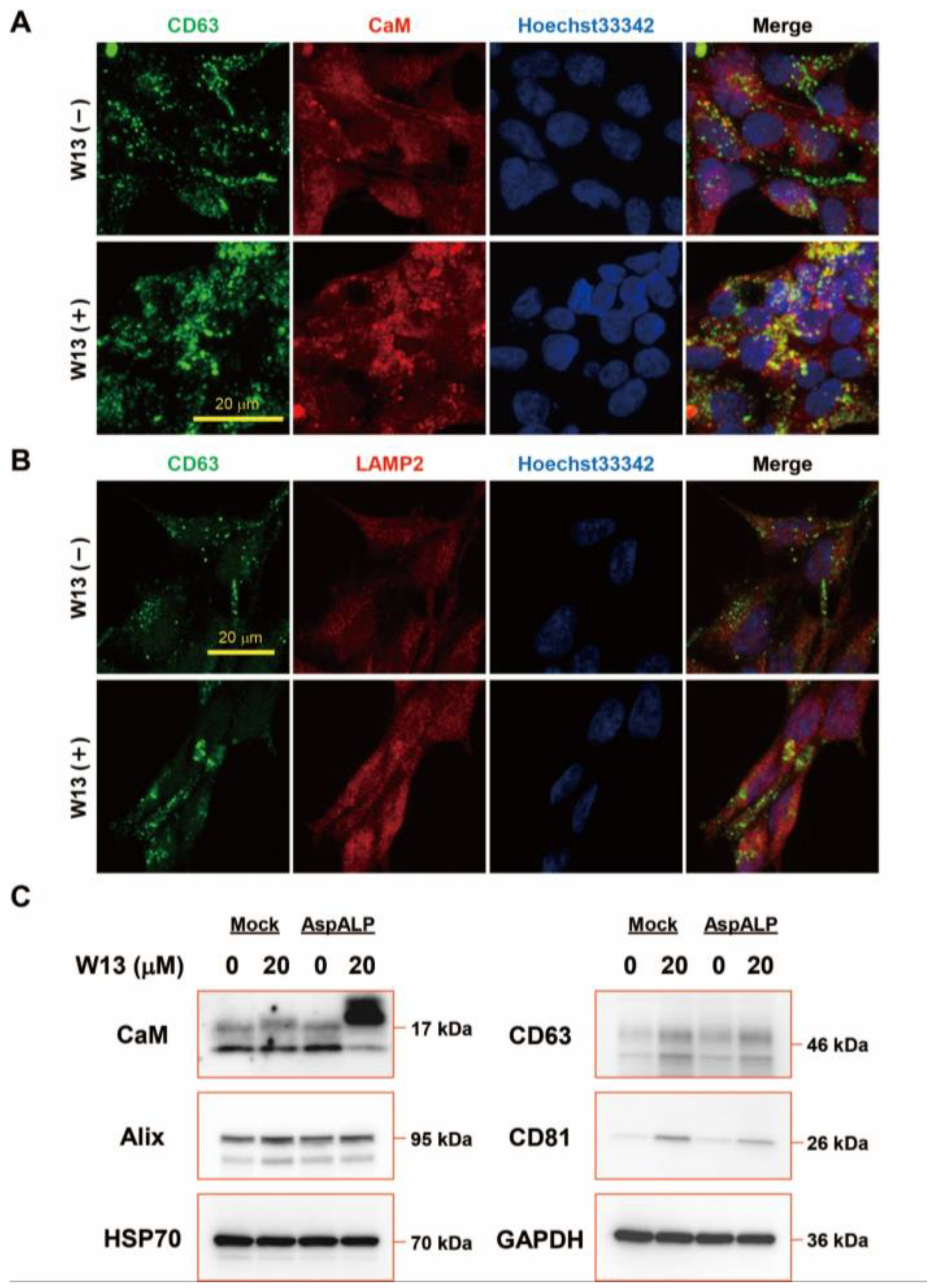
3.5. Interactions between CaM, Signal Peptides, and HSP70 and CD81 in T-REx Cells
4. Discussion
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Jackson, R.; Blobel, G. Post-translational cleavage of presecretory proteins with an extract of rough microsomes from dog pancreas containing signal peptidase activity. Proc. Natl. Acad. Sci. USA 1977, 74, 5598–5602. [Google Scholar] [CrossRef] [Green Version]
- Weihofen, A.; Binns, K.; Lemberg, M.; Ashman, K.; Martoglio, B. Identification of signal peptide peptidase, a presenilin-type aspartic protease. Science 2002, 296, 2215–2218. [Google Scholar] [CrossRef]
- Van Vliet, C.; Thomas, E.; Merino-Trigo, A.; Teasdale, R.; Gleeson, P. Intracellular sorting and transport of proteins. Prog. Biophys. Mol. Biol. 2003, 83, 1–45. [Google Scholar] [CrossRef]
- Pemberton, C.; Siriwardena, M.; Kleffmann, T.; Ruygrok, P.; Palmer, S.; Yandle, T.; Richards, A. First identification of circulating prepro-A-type natriuretic peptide (preproANP) signal peptide fragments in humans: Initial assessment as cardiovascular biomarkers. Clin. Chem. 2012, 58, 757–767. [Google Scholar] [CrossRef] [Green Version]
- Siriwardena, M.; Kleffmann, T.; Ruygrok, P.; Cameron, V.; Yandle, T.; Nicholls, M.; Richards, A.; Pemberton, C. B-type natriuretic peptide signal peptide circulates in human blood: Evaluation as a potential biomarker of cardiac ischemia. Circulation 2010, 122, 255–264. [Google Scholar] [CrossRef] [Green Version]
- Pemberton, C.; Siriwardena, M.; Kleffmann, T.; Richards, A. C-type natriuretic peptide (CNP) signal peptide fragments are present in the human circulation. Biochem. Biophys. Res. Commun. 2014, 449, 301–306. [Google Scholar] [CrossRef]
- Huang, H.; Vasilakis, P.; Zhong, X.; Shen, J.; Geronatsiou, K.; Papadaki, H.; Maragoudakis, M.; Gartaganis, S.; Vinores, S.; Tsopanoglou, N. Parstatin suppresses ocular neovascularization and inflammation. Investig. Ophthalmol. Vis. Sci. 2010, 51, 5825–5832. [Google Scholar] [CrossRef] [Green Version]
- Routhu, K.V.; Tsopanoglou, N.; Strande, J. Parstatin(1-26): The putative signal peptide of protease-activated receptor 1 confers potent protection from myocardial ischemia-reperfusion injury. J. Pharmacol. Exp. Ther. 2010, 332, 898–905. [Google Scholar] [CrossRef]
- Diamantopoulos, A.; Kyriazis, I.; Geronatsiou, K.; Papadaki, H.; Loudos, G.; Kagadis, G.; Katsanos, K.; Liatsikos, E.; Karnabatidis, D.; Siamblis, D.; et al. Parstatin prevents renal injury following ischemia/reperfusion and radiocontrast administration. Am. J. Nephrol. 2012, 36, 278–286. [Google Scholar] [CrossRef]
- Duan, G.-F.; Ye, Y.; Xu, S.; Tao, W.; Zhao, S.; Jin, T.; Nicoll, R.; Shi, Y.; Sheng, N. Signal peptide represses GluK1 surface and synaptic trafficking through binding to amino-terminal domain. Nat. Commun. 2018, 9, 4879. [Google Scholar] [CrossRef]
- Ono, K.; Niwa, M.; Suzuki, H.; Kobayashi, N.; Yoshida, T.; Sawada, M. Secretion of signal peptides via extracellular vesicles. Biochem. Biophys. Res. Commun. 2021, 560, 21–26. [Google Scholar] [CrossRef] [PubMed]
- Ono, K.; Niwa, M.; Suzuki, H.; Kobayashi, N.; Yoshida, T.; Sawada, M. Signal Sequence-Dependent Orientation of Signal Peptide Fragments to Exosomes. Int. J. Mol. Sci. 2022, 23, 3137. [Google Scholar] [CrossRef]
- Martoglio, B.; Graf, R.; Dobberstein, B. Signal peptide fragments of preprolactin and HIV-1 p-gp160 interact with calmodulin. EMBO J. 1997, 16, 6636–6645. [Google Scholar] [CrossRef]
- Villalobo, A.; Ishida, H.; Vogel, H.; Berchtold, M. Calmodulin as a protein linker and a regulator of adaptor/scaffold proteins. Biochim. Biophys. Acta Mol. Cell Res. 2018, 1865, 507–521. [Google Scholar] [CrossRef]
- Skogberg, G.; Gudmundsdottir, J.; van der Post, S.; Sandström, K.; Bruhn, S.; Benson, M.; Mincheva-Nilsson, L.; Baranov, V.; Telemo, E.; Ekwall, O. Characterization of human thymic exosomes. PLoS ONE 2013, 8, e67554. [Google Scholar] [CrossRef]
- Huang, M.; Wei, J.; Peng, W.; Liang, J.; Zhao, C.; Qian, Y.; Dai, G.; Yuan, J.; Pan, F.; Xue, B.; et al. The association of CaM and Hsp70 regulates S-phase arrest and apoptosis in a spatially and temporally dependent manner in human cells. Cell Stress Chaperones 2009, 14, 343–353. [Google Scholar] [CrossRef] [Green Version]
- Yao, F.; Svensjö, T.; Winkler, T.; Lu, M.; Eriksson, C.; Eriksson, E. Tetracycline repressor, tetR, rather than the tetR-mammalian cell transcription factor fusion derivatives, regulates inducible gene expression in mammalian cells. Hum. Gene Ther. 1998, 9, 1939–1950. [Google Scholar] [CrossRef] [PubMed]
- Colomer, J.; Schmitt, A.; Toone, E.; Means, A. Identification and inhibitory properties of a novel Ca2+/ calmodulin antagonist. Biochemistry 2010, 49, 4244–4254. [Google Scholar] [CrossRef] [Green Version]
- Chafouleas, J.; Bolton, W.; Hidaka, H.; Boyd, A.; Means, A. Calmodulin and the cell cycle: Involvement in regulation of cell-cycle progression. Cell 1982, 28, 41–50. [Google Scholar] [CrossRef]
- Mruk, K.; Farley, B.; Ritacco, A.; Kobertz, W. Calmodulation meta-analysis: Predicting calmodulin binding via canonical motif clustering. J. Gen. Physiol. 2014, 144, 105–114. [Google Scholar] [CrossRef]
- Lee, K.; Lee, S.; Lee, W.; Yang, H.; Heo, W.D. Visualizing dynamic interaction between calmodulin and calmodulin-related kinases via a monitoring method in live mammalian cells. Proc. Natl. Acad. Sci. USA 2010, 107, 3412–3417. [Google Scholar] [CrossRef] [Green Version]
- Bosch, M.; Gil, J.; Bachs, O.; Agell, N. Calmodulin inhibitor W13 induces sustained activation of ERK2 and expression of p21(cip1). J. Biol. Chem. 1998, 273, 22145–22150. [Google Scholar] [CrossRef] [Green Version]
- Ma, Y.; Yoshida, T.; Matoba, K.; Kida, K.; Shintani, R.; Piao, Y.; Jin, J.; Nishino, T.; Hanayama, R. Identification of small compounds regulating the secretion of extracellular vesicles via a TIM4-affinity ELISA. Sci. Rep. 2021, 11, 13471. [Google Scholar] [CrossRef]
- Duangmano, S.; Sae-lim, P.; Suksamrarn, A.; Domann, F.; Patmasiriwat, P. Cucurbitacin B inhibits human breast cancer cell proliferation through disruption of microtubule polymerization and nucleophosmin/B23 translocation. BMC Complement. Altern. Med. 2012, 12, 185. [Google Scholar] [CrossRef] [Green Version]
- de Peyster, A.; Wang, Y. Genetic toxicity studies of gossypol. Mutat. Res. Genet. Toxicol. 1993, 297, 293–312. [Google Scholar] [CrossRef]
- Koehler, B.; Scherr, A.; Lorenz, S.; Elssner, C.; Kautz, N.; Welte, S.; Jaeger, D.; Urbanik, T.; Schulze-Bergkamen, H. Pan-Bcl-2 inhibitor obatoclax delays cell cycle progression and blocks migration of colorectal cancer cells. PLoS ONE 2014, 9, e106571. [Google Scholar] [CrossRef]
- Gonzalez-Begne, M.; Lu, B.; Han, X.; Hagen, F.; Hand, A.; Melvin, J.; Yates, J. Proteomic analysis of human parotid gland exosomes by multidimensional protein identification technology (MudPIT). J. Proteome Res. 2009, 8, 1304–1314. [Google Scholar] [CrossRef] [Green Version]
- Harfi, I.; D’Hondt, S.; Sariban, E. iPLA2 Activation Mediates Granular Exocytosis and Corrects Microbicidal Defects in ROS-Deficient and CGD Human Neutrophils. J. Clin. Immunol. 2019, 39, 486–493. [Google Scholar] [CrossRef]
- Baietti, M.; Zhang, Z.; Mortier, E.; Melchior, A.; Degeest, G.; Geeraerts, A.; Ivarsson, Y.; Depoortere, F.; Coomans, C.; Vermeiren, E.; et al. Syndecan–syntenin–ALIX regulates the biogenesis of exosomes. Nat. Cell Biol. 2012, 14, 677–685. [Google Scholar] [CrossRef]
- Cataldo, A.; Barnett, J.; Pieroni, C.; Nixon, R. Increased neuronal endocytosis and protease delivery to early endosomes in sporadic Alzheimer’s disease: Neuropathologic evidence for a mechanism of increased beta-amyloidogenesis. J. Neurosci. 1997, 17, 6142–6151. [Google Scholar] [CrossRef]
- Cataldo, A.; Peterhoff, C.; Troncoso, J.; Gomez-Isla, T.; Hyman, B.; Nixon, R. Endocytic pathway abnormalities precede amyloid beta deposition in sporadic Alzheimer’s disease and Down syndrome: Differential effects of APOE genotype and presenilin mutations. Am. J. Pathol. 2000, 157, 277–286. [Google Scholar] [CrossRef] [PubMed]
- Gauthier, S.; Pérez-González, R.; Sharma, A.; Huang, F.; Alldred, M.; Pawlik, M.; Kaur, G.; Ginsberg, S.; Neubert, T.; Levy, E. Enhanced exosome secretion in Down syndrome brain—A protective mechanism to alleviate neuronal endosomal abnormalities. Acta Neuropathol. Commun. 2017, 5, 65. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tebar, F.; Villalonga, P.; Sorkina, T.; Agell, N.; Sorkin, A.; Enrich, C. Calmodulin regulates intracellular trafficking of epidermal growth factor receptor and the MAPK signaling pathway. Mol. Biol. Cell 2002, 13, 2057–2068. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Apodaca, G.; Enrich, C.; Mostov, K. Calmodulin antagonist, W-13, alters transcytosis, recycling, and the morphology of the endocytic pathway in Madin-Darby Canine Kidney cells. J. Biol. Chem. 1994, 269, 19005–19013. [Google Scholar] [CrossRef] [PubMed]
- Colombo, M.I.; Beron, W.; Stahl, P.D. Calmodulin regulates endosome fusion. J. Biol. Chem. 1997, 272, 7707–7712. [Google Scholar] [CrossRef] [Green Version]
- Sun, X.; Li, B.; Zhou, G.; Tang, W.; Bai, J.; Sun, D.; Zhou, R. Binding of the maize cytosolic Hsp70 to calmodulin, and identification of calmodulin-binding site in Hsp70. Plant Cell Physiol. 2000, 41, 804–810. [Google Scholar] [CrossRef] [Green Version]
- Mayer, M.P.; Bukau, B. Hsp70 chaperones: Cellular functions and molecular mechanism. Cell. Mol. Life Sci. 2005, 62, 670–684. [Google Scholar] [CrossRef] [Green Version]
- Liu, H.; Li, B.; Shang, Z.; Li, X.; Mu, R.; Sun, D.; Zhou, R. Calmodulin is involved in heat shock signal transduction in wheat. Plant Physiol. 2003, 132, 1186–1195. [Google Scholar] [CrossRef] [Green Version]
- Liu, R.; Su, Z.; Zhou, H.; Huang, Q.; Fan, S.; Liu, C.; Han, Y. LsHSP70 is induced by high temperature to interact with calmodulin, leading to higher bolting resistance in lettuce. Sci. Rep. 2020, 10, 15155. [Google Scholar] [CrossRef]
- Li, J.; Soroka, J.; Buchner, J. The Hsp90 chaperone machinery: Conformational dynamics and regulation by co-chaperones. Biochim. Biophys. Acta Mol. Cell Res. 2012, 1823, 624–635. [Google Scholar] [CrossRef]
- Nishida, E.; Koyasu, S.; Sakai, H.; Yahara, I. Calmodulin-regulated binding of the 90-kDa heat shock protein to actin filaments. J. Biol. Chem. 1986, 261, 16033–16036. [Google Scholar] [CrossRef]
- Minami, Y.; Kawasaki, H.; Suzuki, K.; Yahara, I. The calmodulin-binding domain of the mouse 90-kDa heat shock protein. J. Biol. Chem. 1993, 268, 9604–9610. [Google Scholar] [CrossRef]
- Katoh, K.; Suzuki, H.; Terasawa, Y.; Mizuno, T.; Yasuda, J.; Shibata, H.; Maki, M. The penta-EF-hand protein ALG-2 interacts directly with the ESCRT-I component TSG101, and Ca2+-dependently co-localizes to aberrant endosomes with dominant-negative AAA ATPase SKD1/Vps4B. Biochem. J. 2005, 391, 677–685. [Google Scholar] [CrossRef] [Green Version]
- Andreu, Z.; Yáñez-Mó, M. Tetraspanins in extracellular vesicle formation and function. Front. Immunol. 2014, 5, 442. [Google Scholar] [CrossRef] [Green Version]
- Hemler, M.E. Targeting of tetraspanin proteins—Potential benefits and strategies. Nat. Rev. Drug Discov. 2008, 7, 747–758. [Google Scholar] [CrossRef] [Green Version]
- Luga, V.; Zhang, L.; Viloria-Petit, A.; Ogunjimi, A.; Inanlou, M.; Chiu, E.; Buchanan, M.; Hosein, A.; Basik, M.; Wrana, J. Exosomes mediate stromal mobilization of autocrine Wnt-PCP signaling in breast cancer cell migration. Cell 2012, 151, 1542–1556. [Google Scholar] [CrossRef] [Green Version]
- van Niel, G.; Charrin, S.; Simoes, S.; Romao, M.; Rochin, L.; Saftig, P.; Marks, M.; Rubinstein, E.; Raposo, G. The tetraspanin CD63 regulates ESCRT-independent and -dependent endosomal sorting during melanogenesis. Dev. Cell. 2011, 21, 708–721. [Google Scholar] [CrossRef] [Green Version]
- Edrington, T.; Yeagle, P.; Gretzula, C.; Boesze-Battaglia, K. Calcium-Dependent Association of Calmodulin with the C-Terminal Domain of the Tetraspanin Protein Peripherin/rds. Biochemistry 2007, 46, 3862. [Google Scholar] [CrossRef] [Green Version]
- Catalano, M.; O’Driscoll, L. Inhibiting extracellular vesicles formation and release: A review of EV inhibitors. J. Extracell. Vesicles 2020, 9, 1703244. [Google Scholar] [CrossRef] [Green Version]
- Talukder, M.S.U.; Pervin, M.S.; Tanvir, M.; Fujimoto, K.; Tanaka, M.; Itoh, G.; Yumura, S. Ca2+-Calmodulin Dependent Wound Repair in Dictyostelium Cell Membrane. Cells 2020, 9, 1058. [Google Scholar] [CrossRef]





Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ono, K.; Niwa, M.; Suzuki, H.; Kobayashi, N.B.; Yoshida, T.; Sawada, M. Calmodulin as a Key Regulator of Exosomal Signal Peptides. Cells 2023, 12, 158. https://doi.org/10.3390/cells12010158
Ono K, Niwa M, Suzuki H, Kobayashi NB, Yoshida T, Sawada M. Calmodulin as a Key Regulator of Exosomal Signal Peptides. Cells. 2023; 12(1):158. https://doi.org/10.3390/cells12010158
Chicago/Turabian StyleOno, Kenji, Mikio Niwa, Hiromi Suzuki, Nahoko Bailey Kobayashi, Tetsuhiko Yoshida, and Makoto Sawada. 2023. "Calmodulin as a Key Regulator of Exosomal Signal Peptides" Cells 12, no. 1: 158. https://doi.org/10.3390/cells12010158