
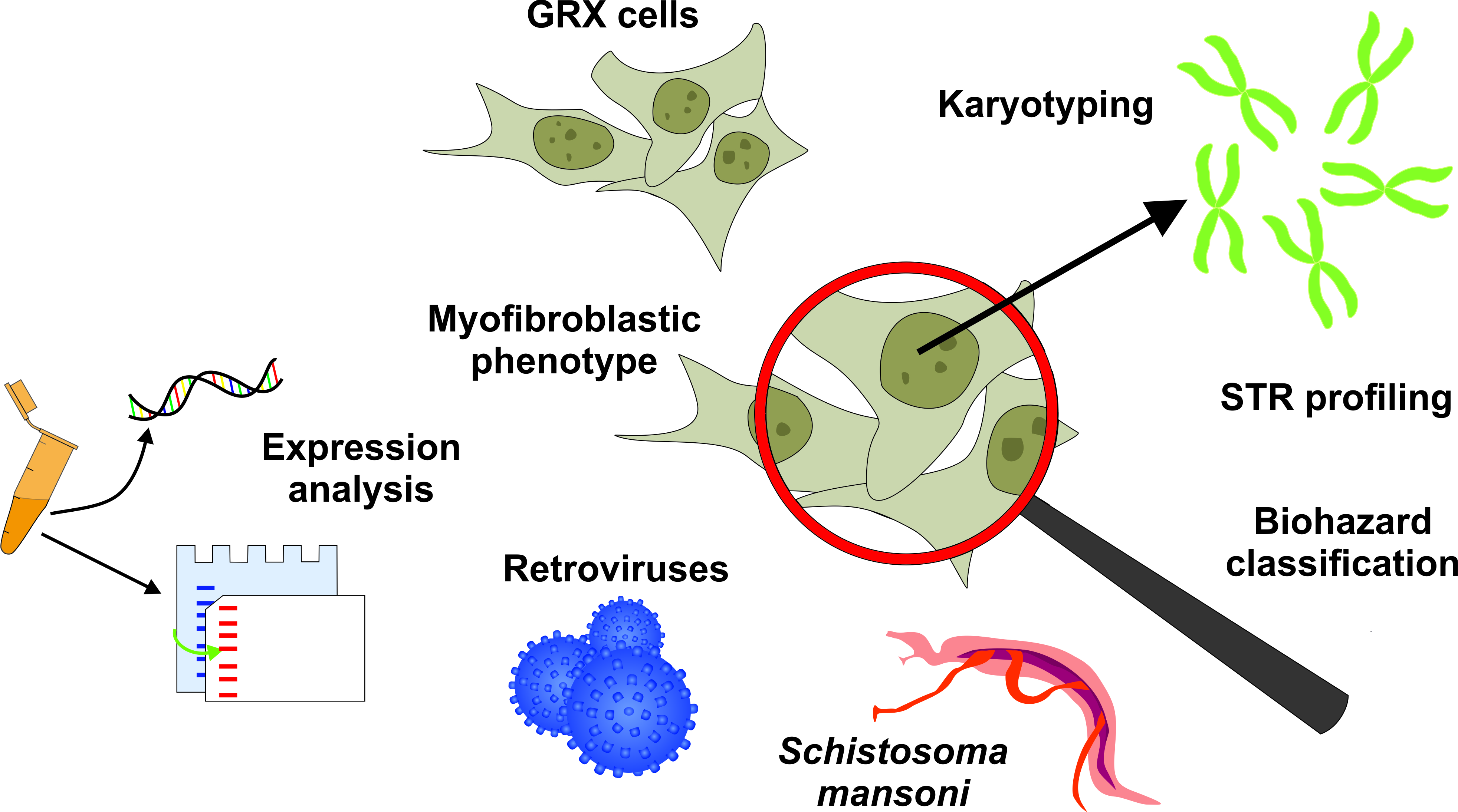
Genetic and Molecular Characterization of the Immortalized Murine Hepatic Stellate Cell Line GRX

 , , , , , , ,
, , , , , , , 
Abstract
:
1. Introduction
2. Materials and Methods
2.1. Literature Search
2.2. Cell Culture
2.3. Karyotyping
2.4. Short Tandem Repeat (STR) Profiling
2.5. Electron Microscopic Analysis
2.6. Analysis of Potential Schistosoma Mansoni Load
2.7. Next-Generation Sequencing
2.8. Western Blot Analysis
2.9. RNA Extraction and PCR Analysis
2.10. Rolling-Circle-Enhanced-Enzyme-Activity Detection (REEAD)
3. Results
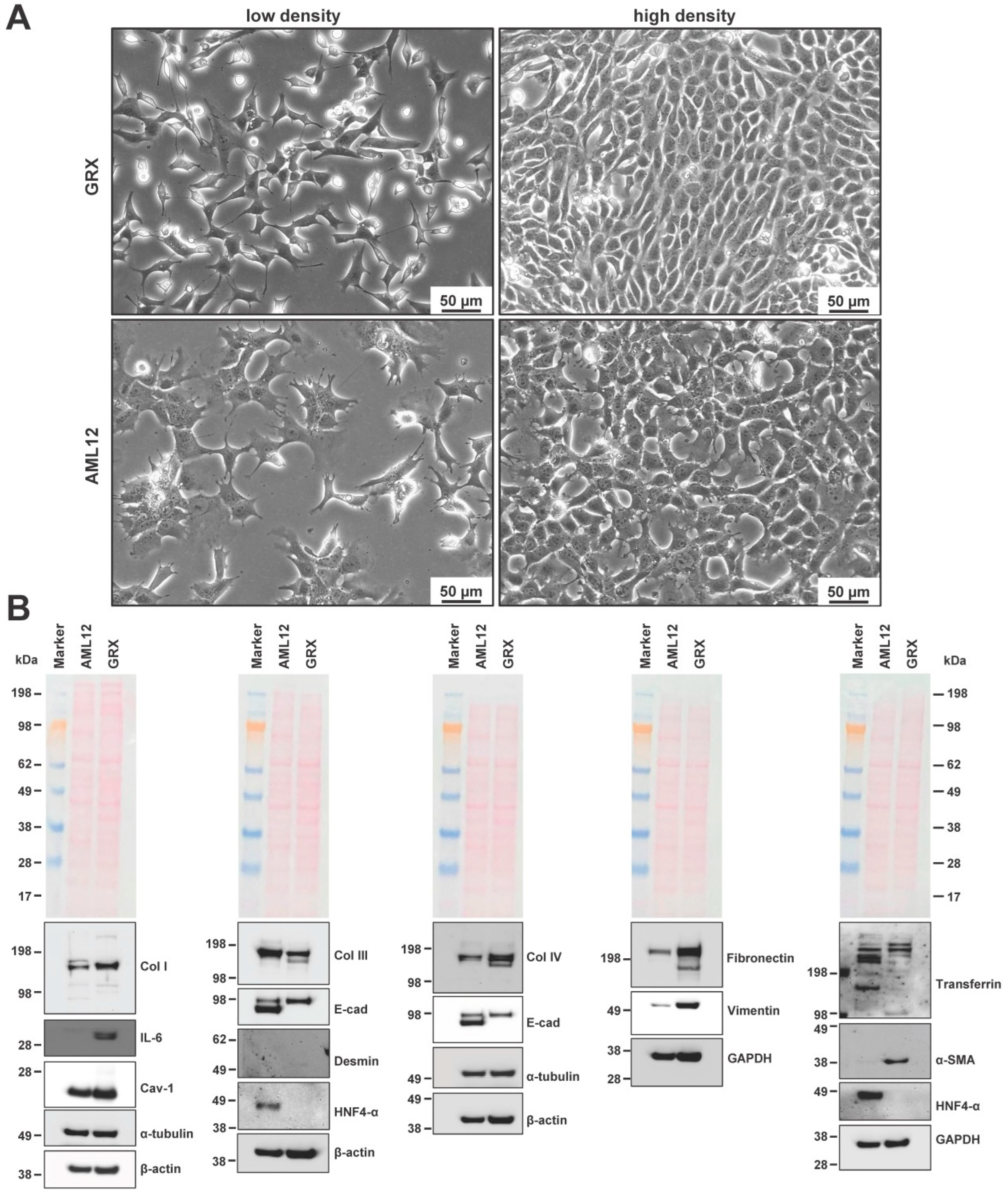
3.1. Phenotypic Characterization of GRX Cells
3.2. Determination of Virus Identity
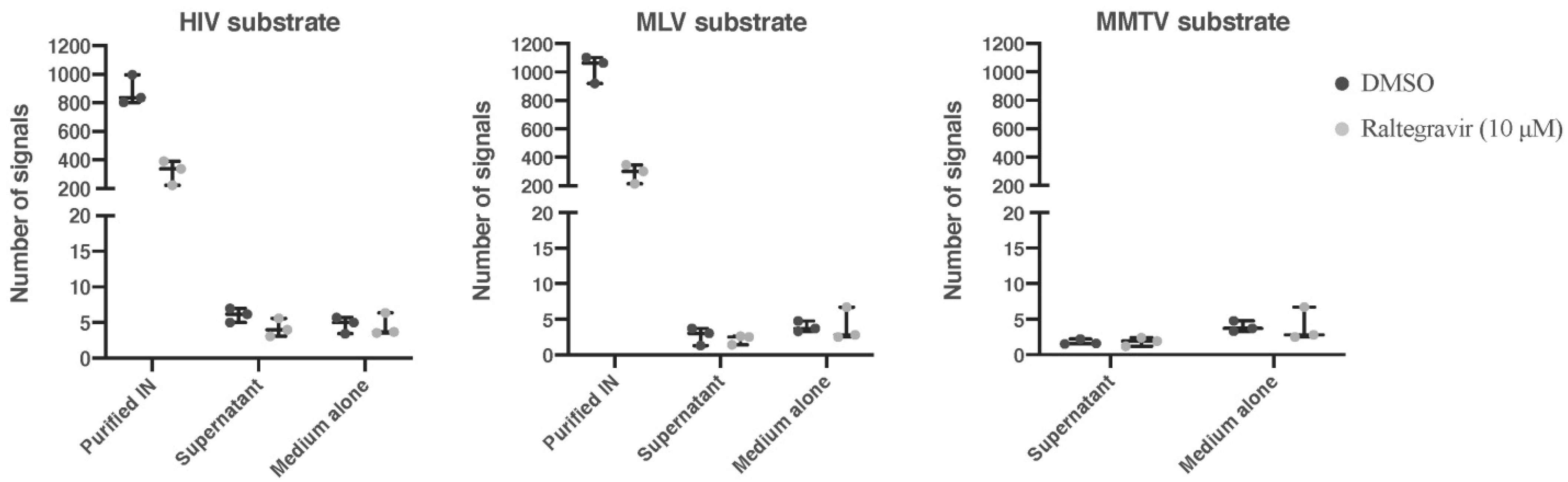
3.3. Analysis Retroviral Activity
3.4. Genotypic Characterization of GRX Cells
3.4.1. Karyogram Analysis of GRX Cells
3.4.2. Chromosomal Arrangements in GRX Cells
3.4.3. Short Tandem Repeat Analysis in GRX Cells
3.5. Considerations on Biological Safety Classification of GRX Cultures
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Weiskirchen, R.; Tacke, F. Cellular and molecular functions of hepatic stellate cells in inflammatory responses and liver immunology. Hepatobiliary Surg. Nutr. 2014, 3, 344–363. [Google Scholar] [CrossRef] [PubMed]
- Weiskirchen, R.; Weiskirchen, S.; Tacke, F. Recent advances in understanding liver fibrosis: Bridging basic science and individualized treatment concepts. F1000Research 2018, 7, 921. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Acharya, P.; Chouhan, K.; Weiskirchen, S.; Weiskirchen, R. Cellular mechanisms of liver fibrosis. Front. Pharmacol. 2021, 12, 671640. [Google Scholar] [CrossRef] [PubMed]
- Herrmann, J.; Gressner, A.M.; Weiskirchen, R. Immortal hepatic stellate cell lines: Useful tools to study hepatic stellate cell biology and function? J. Cell. Mol. Med. 2007, 11, 704–722. [Google Scholar] [CrossRef] [Green Version]
- Borojevic, R.; Monteiro, A.N.; Vinhas, S.A.; Domont, G.B.; Mourão, P.A.; Emonard, H.; Grimaldi, G.J. Grimaud, J.A. Establishment of a continuous cell line from fibrotic schistosomal granulomas in mice livers. Vitr. Cell. Dev. Biol. 1985, 21, 382–390. [Google Scholar] [CrossRef]
- Margis, R.; Borojevic, R. Retinoid-mediated induction of the fat-storing phenotype in a liver connective tissue cell line (GRX). Biochim. Biophys. Acta 1989, 1011, 1–5. [Google Scholar] [CrossRef]
- Borojevic, R.; Guaragna, R.M.; Margis, R.; Dutra, H.S. In vitro induction of the fat-storing phenotype in a liver connective tissue cell line-GRX. Vitr. Cell. Dev. Biol. 1990, 26, 361–368. [Google Scholar] [CrossRef]
- Guaragna, R.M.; Trugo, L.; Borojevic, R. Neutral lipid synthesis and accumulation during in vitro induction of the lipocyte phenotype in hepatic connective tissue cells. Biochim. Biophys. Acta 1991, 1085, 29–34. [Google Scholar] [CrossRef]
- Margis, R.; Pinheiro-Margis, M.; da Silva, L.C.; Borojevic, R. Effects of retinol on proliferation, cell adherence and extracellular matrix synthesis in a liver myofibroblast or lipocyte cell line (GRX). Int. J. Exp. Pathol. 1992, 73, 125–135. [Google Scholar]
- Pinheiro-Margis, M.; Margis, R.; Borojevic, R. Collagen synthesis in an established liver connective tissue cell line (GRX) during induction of the fat-storing phenotype. Exp. Mol. Pathol. 1992, 56, 108–118. [Google Scholar] [CrossRef]
- Vicente, C.P.; Fortuna, V.A.; Margis, R.; Trugo, L.; Borojevic, R. Retinol uptake and metabolism, and cellular retinol binding protein expression in an in vitro model of hepatic stellate cells. Mol. Cell. Biochem. 1998, 187, 11–21. [Google Scholar] [CrossRef] [PubMed]
- Russell, W.M.S.; Burch, R. The Principles of Humane Experimental Technique; Methuen: London, UK, 1959. [Google Scholar]
- Liedtke, C.; Luedde, T.; Sauerbruch, T.; Scholten, D.; Streetz, K.; Tacke, F.; Tolba, R.; Trautwein, C.; Trebicka, J.; Weiskirchen, R. Experimental liver fibrosis research: Update on animal models, legal issues and translational aspects. Fibrogenesis Tissue Repair 2013, 6, 19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- National Library of Medicine. Available online: https://pubmed.ncbi.nlm.nih.gov/ (accessed on 30 March 2022).
- Google. Available online: https://www.google.com/ (accessed on 30 March 2022).
- Xu, L.; Hui, A.Y.; Albanis, E.; Arthur, M.J.; O’Byrne, S.M.; Blaner, W.S.; Mukherjee, P.; Friedman, S.L.; Eng, F.J. Human hepatic stellate cell lines, LX-1 and LX-2: New tools for analysis of hepatic fibrosis. Gut 2005, 54, 142–151. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vogel, S.; Piantedosi, R.; Frank, J.; Lalazar, A.; Rockey, D.C.; Friedman, S.L.; Blaner, W.S. An immortalized rat liver stellate cell line (HSC-T6): A new cell model for the study of retinoid metabolism in vitro. J. Lipid Res. 2000, 41, 882–893. [Google Scholar] [CrossRef]
- Greenwel, P.; Schwartz, M.; Rosas, M.; Peyrol, S.; Grimaud, J.A.; Rojkind, M. Characterization of fat-storing cell lines derived from normal and CCl4-cirrhotic livers. Differences in the production of interleukin-6. Lab. Invest. 1991, 65, 644–653. [Google Scholar]
- Greenwel, P.; Rubin, J.; Schwartz, M.; Hertzberg, E.L.; Rojkind, M. Liver fat-storing cell clones obtained from a CCl4-cirrhotic rat are heterogeneous with regard to proliferation, expression of extracellular matrix components, interleukin-6, and connexin 43. Lab. Invest. 1993, 69, 210–216. [Google Scholar]
- Wu, J.C.; Merlino, G.; Fausto, N. Establishment and characterization of differentiated, nontransformed hepatocyte cell lines derived from mice transgenic for transforming growth factor alpha. Proc. Natl. Acad. Sci. USA 1994, 91, 674–678. [Google Scholar] [CrossRef] [Green Version]
- Meurer, S.K.; Tezcan, O.; Lammers, T.; Weiskirchen, R. Differential regulation of Lipocalin 2 (LCN2) in doxorubicin-resistant 4T1 triple negative breast cancer cells. Cell. Signal. 2020, 74, 109731. [Google Scholar] [CrossRef]
- MGI. Guidelines for Nomenclature of Genes, Genetic Markers, Alleles, and Mutations in Mouse and Rat. Available online: http://www.informatics.jax.org/mgihome/nomen/gene.shtml (accessed on 30 March 2022).
- Almeida, J.L.; Hill, C.R.; Cole, K.D. Mouse cell line authentication. Cytotechnology 2014, 66, 133–147. [Google Scholar] [CrossRef] [Green Version]
- Almeida, J.L.; Dakic, A.; Kindig, K.; Kone, M.; Letham, D.L.D.; Langdon, S.; Peat, R.; Holding-Pillai, J.; Hall, E.M.; Ladd, M.; et al. Interlaboratory study to validate a STR profiling method for intraspecies identification of mouse cell lines. PLoS ONE 2019, 14, e0218412. [Google Scholar] [CrossRef] [Green Version]
- Expasy Bioinformatics Resource Portal. Cellosaurus. CLASTR 1.4.4. The Cellosaurus STR Similarity Search Tool. Available online: https://web.expasy.org/cellosaurus-str-search (accessed on 30 March 2022).
- Deelder, A.M.; van Dam, G.J.; Kornelis, D.; Fillié, Y.E.; van Zeyl, R.J. Schistosoma: Analysis of monoclonal antibodies reactive with the circulating antigens CAA and CCA. Parasitology 1996, 112, 21–35. [Google Scholar] [CrossRef] [PubMed]
- Robijn, M.L.; Wuhrer, M.; Kornelis, D.; Deelder, A.M.; Geyer, R.; Hokke, C.H. Mapping fucosylated epitopes on glycoproteins and glycolipids of Schistosoma mansoni cercariae, adult worms and eggs. Parasitology 2005, 130, 67–77. [Google Scholar] [CrossRef] [PubMed]
- Schramm, G.; Falcone, F.H.; Gronow, A.; Haisch, K.; Mamat, U.; Doenhoff, M.J.; Oliveira, G.; Galle, J.; Dahinden, C.A.; Haas, H. Molecular characterization of an interleukin-4-inducing factor from Schistosoma mansoni eggs. J. Biol. Chem. 2003, 278, 18384–18392. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schramm, G.; Hamilton, J.V.; Balog, C.I.; Wuhrer, M.; Gronow, A.; Beckmann, S.; Wippersteg, V.; Grevelding, C.G.; Goldmann, T.; Weber, E.; et al. Molecular characterisation of kappa-5, a major antigenic glycoprotein from Schistosoma mansoni eggs. Mol. Biochem. Parasitol. 2009, 166, 4–14. [Google Scholar] [CrossRef]
- Fitzsimmons, C.M.; Schramm, G.; Jones, F.M.; Chalmers, I.W.; Hoffmann, K.F.; Grevelding, C.G.; Wuhrer, M.; Hokke, C.H.; Haas, H.; Doenhoff, M.J.; et al. Molecular characterization of omega-1: A hepatotoxic ribonuclease from Schistosoma mansoni eggs. Mol. Biochem. Parasitol. 2005, 144, 123–127. [Google Scholar] [CrossRef]
- Klüber, P.; Meurer, S.K.; Lambertz, J.; Schwarz, R.; Zechel-Gran, S.; Braunschweig, T.; Hurka, S.; Domann, E.; Weiskirchen, R. Depletion of Lipocalin 2 (LCN2) in mice leads to dysbiosis and persistent colonization with segmented filamentous bacteria. Int. J. Mol. Sci. 2021, 22, 13156. [Google Scholar] [CrossRef]
- Schmittgen, T.D.; Livak, K.J. Analyzing real-time PCR data by the comparative C(T) method. Nat. Protoc. 2008, 3, 1101–1108. [Google Scholar] [CrossRef]
- Wang, J.; Liu, J.; Thomsen, J.; Selnihhin, D.; Hede, M.S.; Kirsebom, F.C.; Franch, O.; Fjelstrup, S.; Stougaard, M.; Ho, Y.P.; et al. Novel DNA sensor system for highly sensitive and quantitative retrovirus detection using virus encoded integrase as a biomarker. Nanoscale 2017, 9, 440–448. [Google Scholar] [CrossRef]
- Beck-Engeser, G.B.; Eilat, D.; Harrer, T.; Jäck, H.M.; Wabl, M. Early onset of autoimmune disease by the retroviral integrase inhibitor raltegravir. Proc. Natl. Acad. Sci. USA 2009, 106, 20865–20870. [Google Scholar] [CrossRef] [Green Version]
- de Oliveira, C.M.; Martins, L.A.M.; de Sousa, A.C.; Moraes, K.D.S.; Costa, B.P.; Vieira, M.Q.; Coelho, B.P.; Borojevic, R.; de Oliveira, J.R.; Guma, F.C.R. Resveratrol increases the activation markers and changes the release of inflammatory cytokines of hepatic stellate cells. Mol. Cell. Biochem. 2021, 476, 649–661. [Google Scholar] [CrossRef]
- Mus Musculus Strain NOD/ShiLtJ Endogenous Virus Ecotropic Murine Leukemia Virus 30, Complete Sequence. Available online: https://www.ncbi.nlm.nih.gov/nuccore/KJ668269 (accessed on 30 March 2022).
- Chu, H.; Wang, J.J.; Qi, M.; Yoon, J.J.; Chen, X.; Wen, X.; Hammonds, J.; Ding, L.; Spearman, P. Tetherin/BST-2 is essential for the formation of the intracellular virus-containing compartment in HIV-infected macrophages. Cell Host Microbe 2012, 12, 360–372. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Han, Z.; Lv, M.; Shi, Y.; Yu, J.; Niu, J.; Yu, X.F.; Zhang, W. Mutation of glycosylation sites in BST-2 leads to its accumulation at intracellular CD63-positive vesicles without affecting its antiviral activity against multivesicular body-targeted HIV-1 and hepatitis B virus. Viruses 2016, 8, 62. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Waheed, A.A.; Gitzen, A.; Swiderski, M.; Freed, E.O. High-mannose but not complex-type glycosylation of tetherin is required for restriction of HIV-1 release. Viruses 2018, 10, 26. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ferreira, J.G.G.; Gava, S.G.; Oliveira, E.S.; Batista, I.C.A.; Fernandes, G.D.R.; Mourão, M.M.; Calzavara-Silva, C.E. Gene expression signatures in AML-12 hepatocyte cells upon Dengue virus infection and acetaminophen treatment. Viruses 2020, 12, 1284. [Google Scholar] [CrossRef]
- Mahauad-Fernandez, W.D.; DeMali, K.A.; Olivier, A.K.; Okeoma, C.M. Bone marrow stromal antigen 2 expressed in cancer cells promotes mammary tumor growth and metastasis. Breast Cancer Res. 2014, 16, 493. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Almeida, J.L.; Cole, K.D.; Plant, A.L. Standards for cell line authentication and beyond. PLoS Biol. 2016, 14, e1002476. [Google Scholar] [CrossRef]
- Nachman, M.W.; Searle, J.B. Why is the house mouse karyotype so variable? Trends Ecol. Evol. 1995, 10, 397–402. [Google Scholar] [CrossRef]
- Garagna, S.; Marziliano, N.; Zuccotti, M.; Searle, J.B.; Capanna, E.; Redi, C.A. Pericentromeric organization at the fusion point of mouse Robertsonian translocation chromosomes. Proc. Natl. Acad. Sci. USA 2001, 98, 171–175. [Google Scholar] [CrossRef] [Green Version]
- Cazaux, B.; Catalan, J.; Claude, J.; Britton-Davidian, J. Non-random occurrence of Robertsonian translocations in the house mouse (Mus musculus domesticus): Is it related to quantitative variation in the minor satellite? Cytogenet. Genome Res. 2014, 144, 124–130. [Google Scholar] [CrossRef]
- Bairoch, A. The Cellosaurus, a Cell-Line Knowledge Resource. J. Biomol. Tech. 2018, 29, 25–38. [Google Scholar] [CrossRef]
- Robin, T.; Capes-Davis, A.; Bairoch, A. CLASTR: The Cellosaurus STR similarity search tool-A precious help for cell line authentication. Int. J. Cancer 2020, 146, 1299–1306. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rechtsinformationssystem des Bundes. Bundesrecht Konsolidiert: Gesamte Rechtsvorschrift für Verordnung Biologische Arbeitsstoffe, Fassung vom 9.7. 2021. Available online: https://www.ris.bka.gv.at/GeltendeFassung.wxe?Abfrage=Bundesnormen&Gesetzesnummer=10009126 (accessed on 30 March 2022).
- Banco de Células do Rio de Janeiro (BCRJ). GRX. Available online: https://bcrj.org.br/celula/grx-liver-schistomiasis-mouse-fibroblast. (accessed on 30 March 2022).
- Corstjens, P.L.; De Dood, C.J.; Kornelis, D.; Fat, E.M.; Wilson, R.A.; Kariuki, T.M.; Nyakundi, R.K.; Loverde, P.T.; Abrams, W.R.; Tanke, H.J.; et al. Tools for diagnosis, monitoring and screening of Schistosoma infections utilizing lateral-flow based assays and upconverting phosphor labels. Parasitology 2014, 141, 1841–1855. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Smit, C.H.; Homann, A.; van Hensbergen, V.P.; Schramm, G.; Haas, H.; van Diepen, A.; Hokke, C.H. Surface expression patterns of defined glycan antigens change during Schistosoma mansoni cercarial transformation and development of schistosomula. Glycobiology 2015, 25, 1465–1479. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- American Type Culture Collection Standards Development Organization Workgroup ASN-0002. Cell line misidentification: The beginning of the end. Nat. Rev. Cancer 2010, 10, 441–448. [Google Scholar] [CrossRef] [PubMed]
- Geraghty, R.J.; Capes-Davis, A.; Davis, J.M.; Downward, J.; Freshney, R.I.; Knezevic, I.; Lovell-Badge, R.; Masters, J.R.; Meredith, J.; Stacey, G.N.; et al. Guidelines for the use of cell lines in biomedical research. Br. J. Cancer 2014, 111, 1021–1046. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Weiskirchen, R.; Weimer, J.; Meurer, S.K.; Kron, A.; Seipel, B.; Vater, I.; Arnold, N.; Siebert, R.; Xu, L.; Friedman, S.L.; et al. Genetic characteristics of the human hepatic stellate cell line LX-2. PLoS ONE 2013, 8, e75692. [Google Scholar] [CrossRef] [Green Version]
- Walker, W.S.; Gatewood, J.; Olivas, E.; Askew, D.; Havenith, C.E. Mouse microglial cell lines differing in constitutive and interferon-gamma-inducible antigen-presenting activities for naive and memory CD4+ and CD8+ T cells. J. Neuroimmunol. 1995, 63, 163–174. [Google Scholar] [CrossRef]
- Littlefield, J.W. Three degrees of guanylic acid-inosinic acid pyrophosphorylase deficiency in mouse fibroblasts. Nature 1964, 203, 1142–1144. [Google Scholar] [CrossRef]
- Earle, W.R.; Schilling, E.L.; Stark, T.H.; Straus, N.P.; Brown, M.F.; Shelton, E. Production of malignancy in vitro. IV. The mouse fibroblast cultures and changes seen in the living cells. J. Nat. Cancer Inst. 1943, 4, 165–212. [Google Scholar] [CrossRef]
- Sandford, K.K.; Earle, W.R.; Likely, G.D. The growth in vitro of single isolated tissue cells. J. Natl. Cancer Inst. 1948, 9, 229–246. [Google Scholar]
- Sandford, K.K.; Hobbs, G.L.; Earle, W.R. The tumor-producing capacity of strain L mouse cells after 10 year in vitro. Cancer Res. 1956, 16, 162–166. [Google Scholar]
- McBurney, M.W.; Rogers, B.J. Isolation of male embryonal carcinoma cells and their chromosome replication patterns. Dev. Biol. 1982, 89, 503–508. [Google Scholar] [CrossRef]
- Nii, M.; Kayada, Y.; Yoshiga, K.; Takada, K.; Okamoto, T.; Yanagihara, K. Suppression of metastasis by tissue inhibitor of metalloproteinase-1 in a newly established human oral squamous cell carcinoma cell line. Int. J. Oncol. 2000, 16, 119–124. [Google Scholar] [CrossRef] [PubMed]
- ICLAC Register of Misidentified Cell Lines: Archive. Version 11 (8 June 2021). Available online: https://iclac.org/databases/cross-contaminations/archive/ (accessed on 30 March 2022).
- Wu, H.; Reizel, T.; Wang, Y.J.; Lapiro, J.L.; Kren, B.T.; Schug, J.; Rao, S.; Morgan, A.; Herman, A.; Shekels, L.L.; et al. A negative reciprocal regulatory axis between cyclin D1 and HNF4α modulates cell cycle progression and metabolism in the liver. Proc. Natl. Acad. Sci. USA 2020, 117, 17177–17186. [Google Scholar] [CrossRef] [PubMed]
- Kwon, M.; Leibowitz, M.L.; Lee, J.H. Small but mighty: The causes and consequences of micronucleus rupture. Exp. Mol. Med. 2020, 52, 1777–1786. [Google Scholar] [CrossRef]
- Hancks, D.C. A Role for retrotransposons in chromothripsis. Methods Mol. Biol. 2018, 1769, 169–181. [Google Scholar] [CrossRef]
- Callahan, R.; Smith, G.H. MMTV-induced mammary tumorigenesis: Gene discovery, progression to malignancy and cellular pathways. Oncogene 2000, 19, 992–1001. [Google Scholar] [CrossRef] [Green Version]
- Benoit, I.; Brownell, S.; Douville, R.N. Predicted Cellular Interactors of the Endogenous Retrovirus-K Integrase Enzyme. Microorganisms 2021, 9, 1509. [Google Scholar] [CrossRef]
- Milligan, J.N.; Jolly, E.R. Cercarial transformation and in vitro cultivation of Schistosoma mansoni schistosomules. J. Vis. Exp. 2011, 54, 3191. [Google Scholar] [CrossRef] [Green Version]
- van Noort, K.; Nguyen, D.L.; Kriechbaumer, V.; Hawes, C.; Hokke, C.H.; Schots, A.; Wilbers, R.H.P. Functional characterization of Schistosoma mansoni fucosyltransferases in Nicotiana benthamiana plants. Sci. Rep. 2020, 10, 18528. [Google Scholar] [CrossRef]
- Roderfeld, M.; Padem, S.; Lichtenberger, J.; Quack, T.; Weiskirchen, R.; Longerich, T.; Schramm, G.; Churin, Y.; Irungbam, K.; Tschuschner, A.; et al. Schistosoma mansoni egg-secreted antigens activate hepatocellular carcinoma-associated transcription factors c-Jun and STAT3 in hamster and human hepatocytes. Hepatology 2020, 72, 626–641. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cellosaurus–A Knowledge Resource on Cell Lines. Available online: https://web.expasy.org/cellosaurus/ (accessed on 30 March 2022).










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| STR Marker | GRX | EOC 20 | HNOS | A-9 | P19 | NCTC Clone 929 |
|---|---|---|---|---|---|---|
| 1-1 | 10 | 10 | 10 | 10 | 10 | 10 |
| 1-2 | 16 | 16 | 16 | 17, 18 | 16, 17 | 17 |
| 2-1 | 9 | 9 | 9, 16 | 9 | 9 | 9 |
| 3-2 | 14 | 14 | 14 | 14 | 14 | 13, 14 |
| 4-2 | 19.3 | 19.3 | 20.3 | 20.3, 21.3 | 13, 21.3 | 20.3 |
| 5-5 | 15 | 15 | 15 | 14, 15 | 14 | 14 |
| 6-4 | 19 | 19 | 18 | 17, 18 | 18 | 17, 18 |
| 6-7 | 12 | 12 | 12, 16 | 12 | 12 | 12 |
| 7-1 | 26 | 26 | 26.2 | 26, 27 | 26 | 25, 26, 27 |
| 8-1 | 16 | 16, 17 | 16 | 15, 16, 17 | 16 | 16 |
| 11-2 | 16 | 17 | 16 | 16 | 16 | 15, 16 |
| 12-1 | 16 | 16 | 16,17 | 16 | 16 | 16 |
| 13-1 | 17 | 17 | 17 | 17 | 17 | 17 |
| 15-3 | 25.3 | 25.3 | 22.3, 25.3 | 25.3 | 26.3 | 24.3, 25.3, 26.3 |
| 17-2 | 16 | 16 | 15, 16 | 15 | 15 | 15 |
| 18-3 | 16 | 16 | 16 | 16 | 16, 17 | 16 |
| 19-2 | 12 | 12 | 12, 13 | 12 | 12, 14 | 12 |
| X-1 | 26, 27 | 27 | 27 | 26, 27 | 26 | 26, 27 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Schröder, S.K.; Schüler, H.M.; Petersen, K.V.; Tesauro, C.; Knudsen, B.R.; Pedersen, F.S.; Krus, F.; Buhl, E.M.; Roeb, E.; Roderfeld, M.; et al. Genetic and Molecular Characterization of the Immortalized Murine Hepatic Stellate Cell Line GRX. Cells 2022, 11, 1504. https://doi.org/10.3390/cells11091504
Schröder SK, Schüler HM, Petersen KV, Tesauro C, Knudsen BR, Pedersen FS, Krus F, Buhl EM, Roeb E, Roderfeld M, et al. Genetic and Molecular Characterization of the Immortalized Murine Hepatic Stellate Cell Line GRX. Cells. 2022; 11(9):1504. https://doi.org/10.3390/cells11091504
Chicago/Turabian StyleSchröder, Sarah K., Herdit M. Schüler, Kamilla V. Petersen, Cinzia Tesauro, Birgitta R. Knudsen, Finn S. Pedersen, Frederike Krus, Eva M. Buhl, Elke Roeb, Martin Roderfeld, and et al. 2022. "Genetic and Molecular Characterization of the Immortalized Murine Hepatic Stellate Cell Line GRX" Cells 11, no. 9: 1504. https://doi.org/10.3390/cells11091504