
Evolution of the Degenerated Y-Chromosome of the Swamp Guppy, Micropoecilia picta

 , and
, and
Abstract
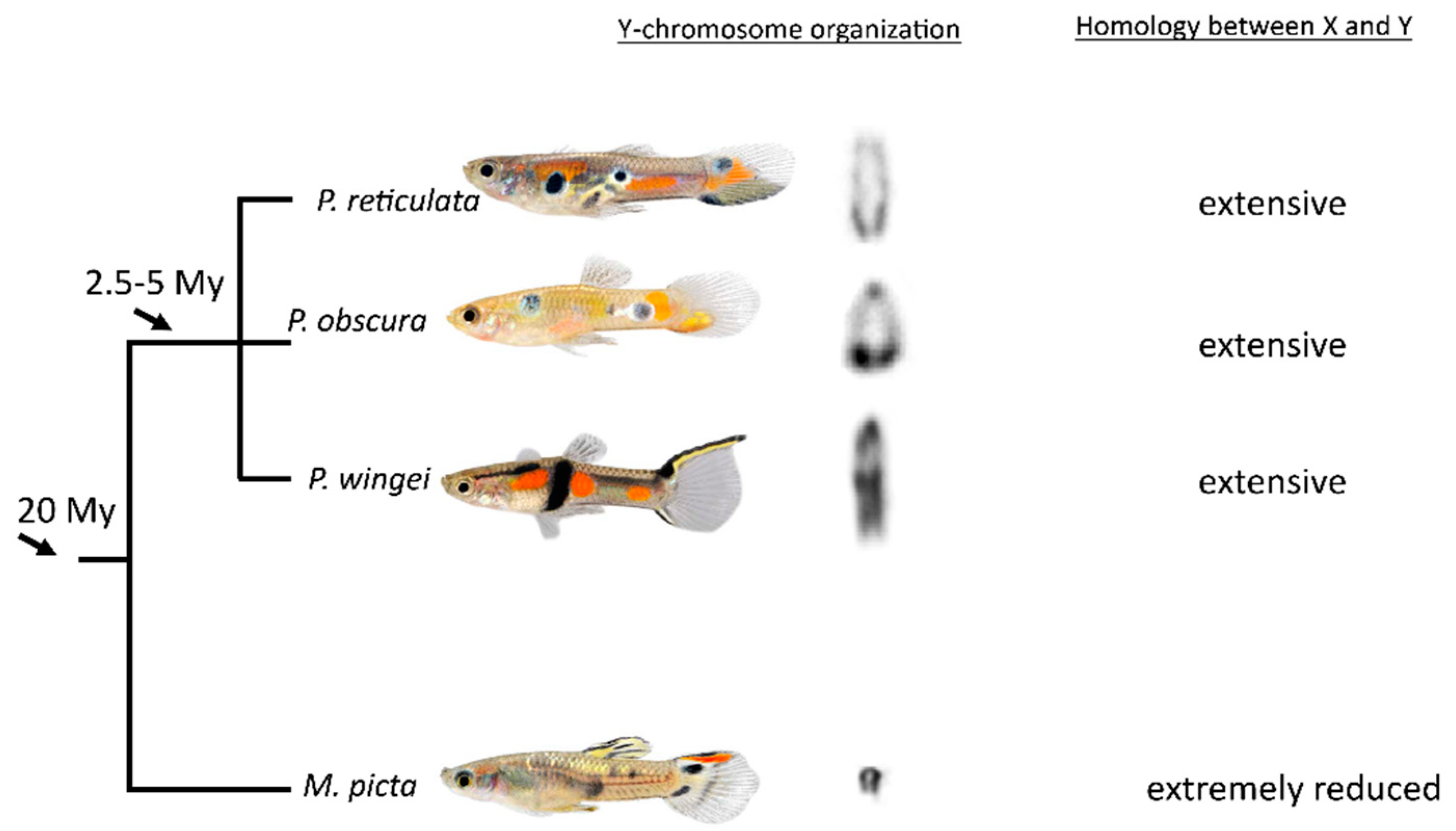
:1. Introduction
2. Materials and Methods
2.1. Animals
2.2. Mitotic Chromosome Preparation
2.3. Meiotic Chromosome Preparation
2.4. Immunostaining of Synaptonemal Complex and MLH1
2.5. Immunolocalization of Anti-5-Methylcytosine
2.6. Hormone Treatments
3. Results
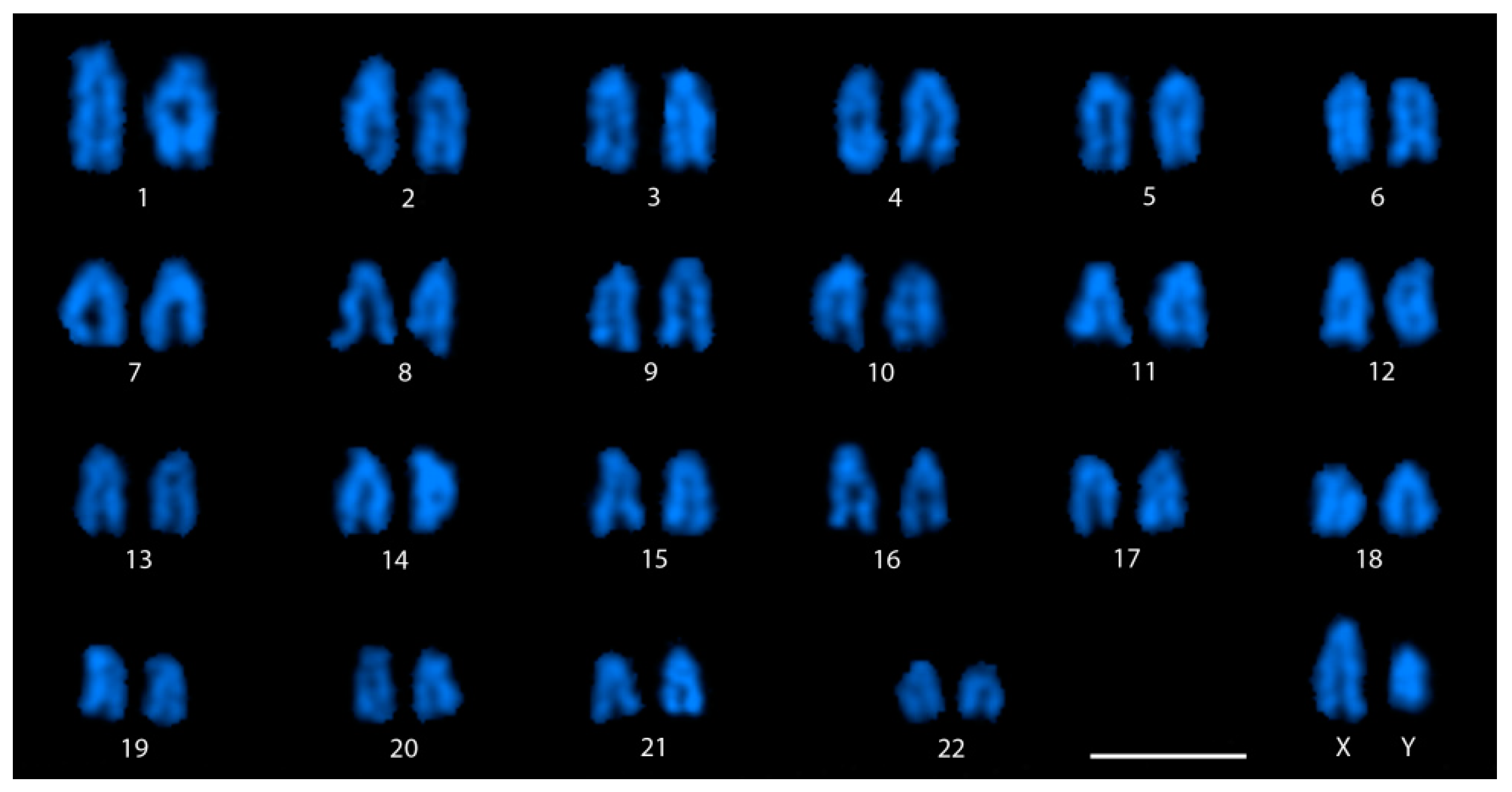
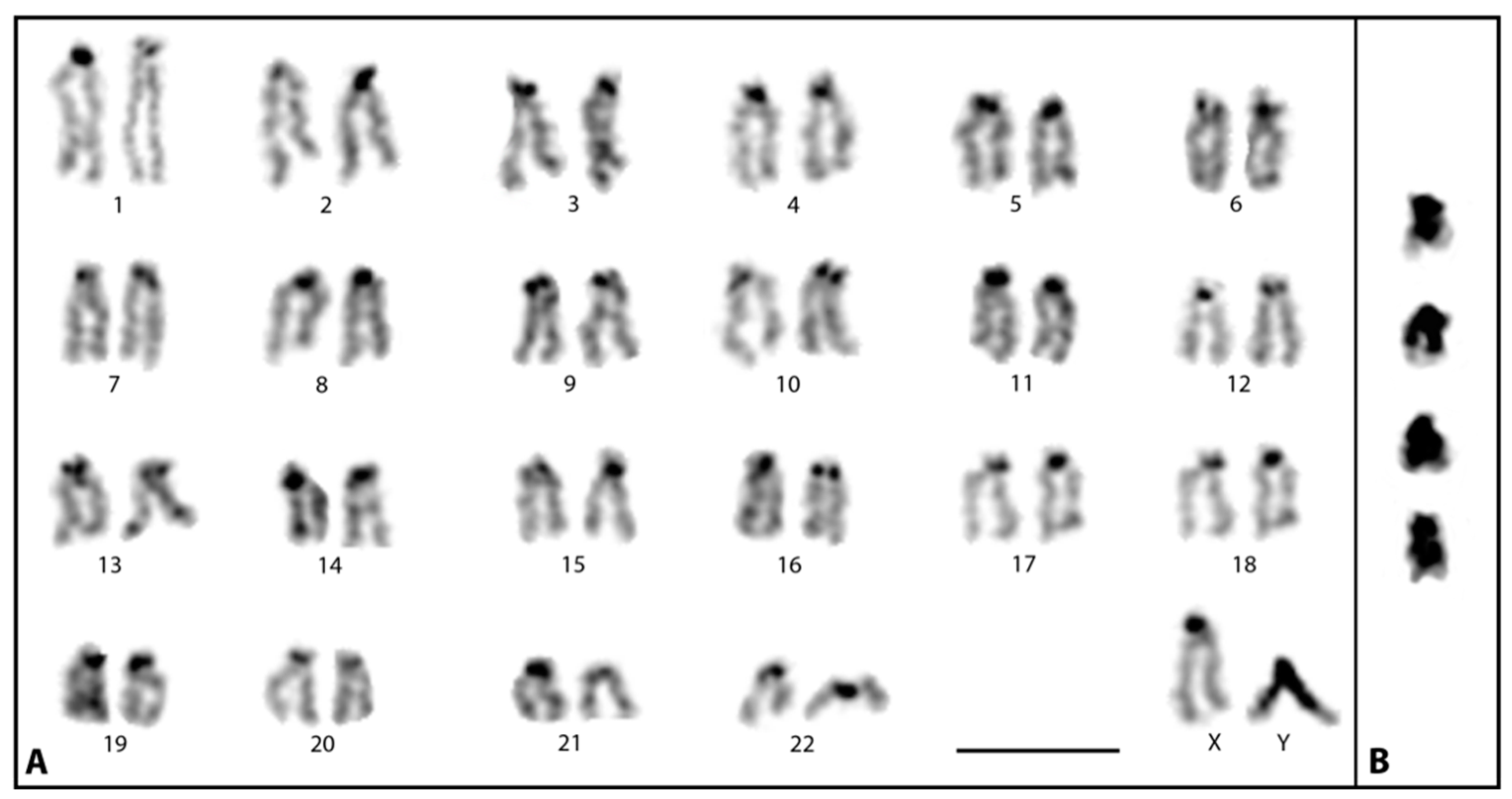
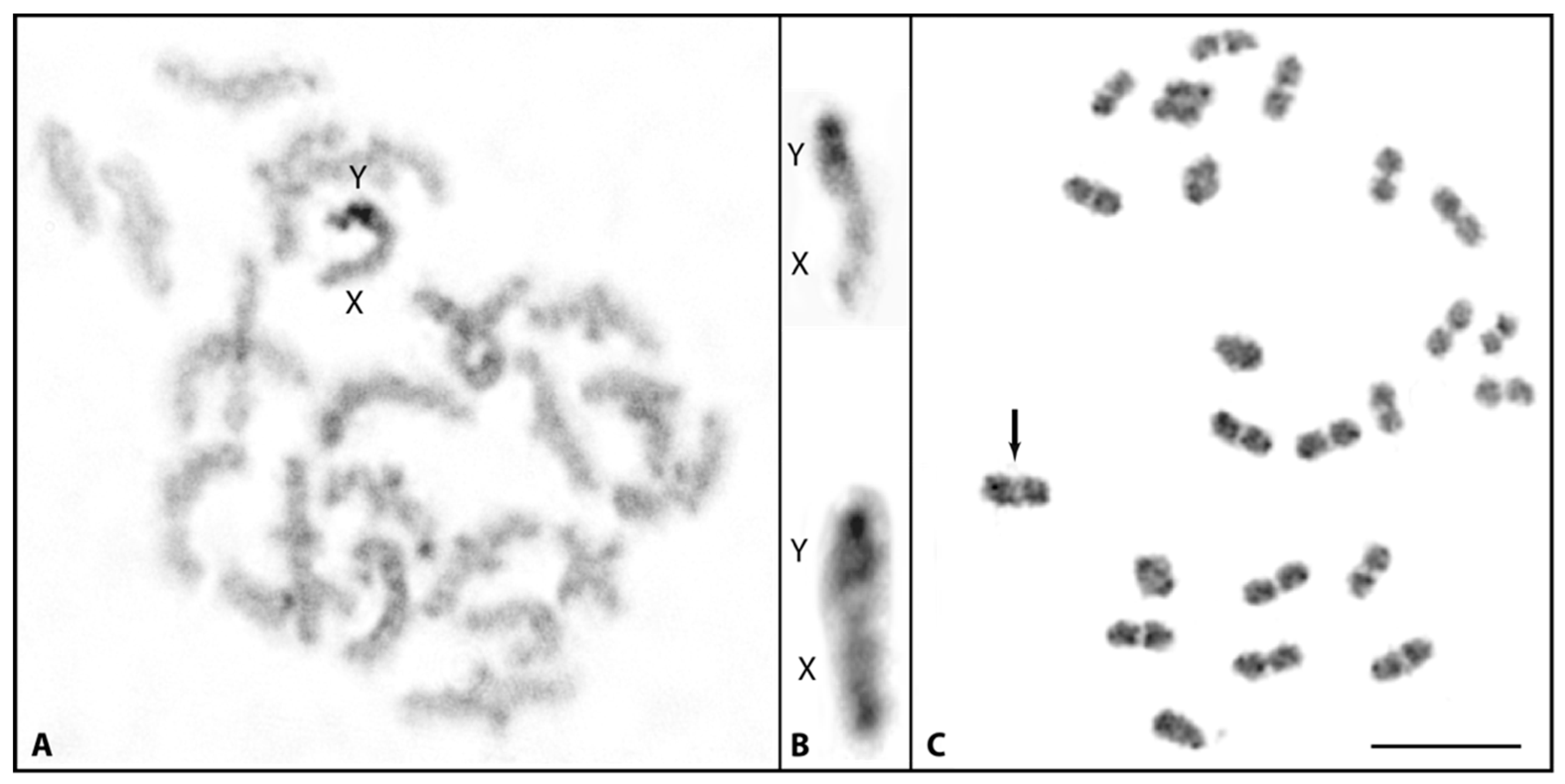
3.1. Cytological Identification of the M. picta Y Chromosome
3.2. Meiotic Association and Recombination between the X and Y
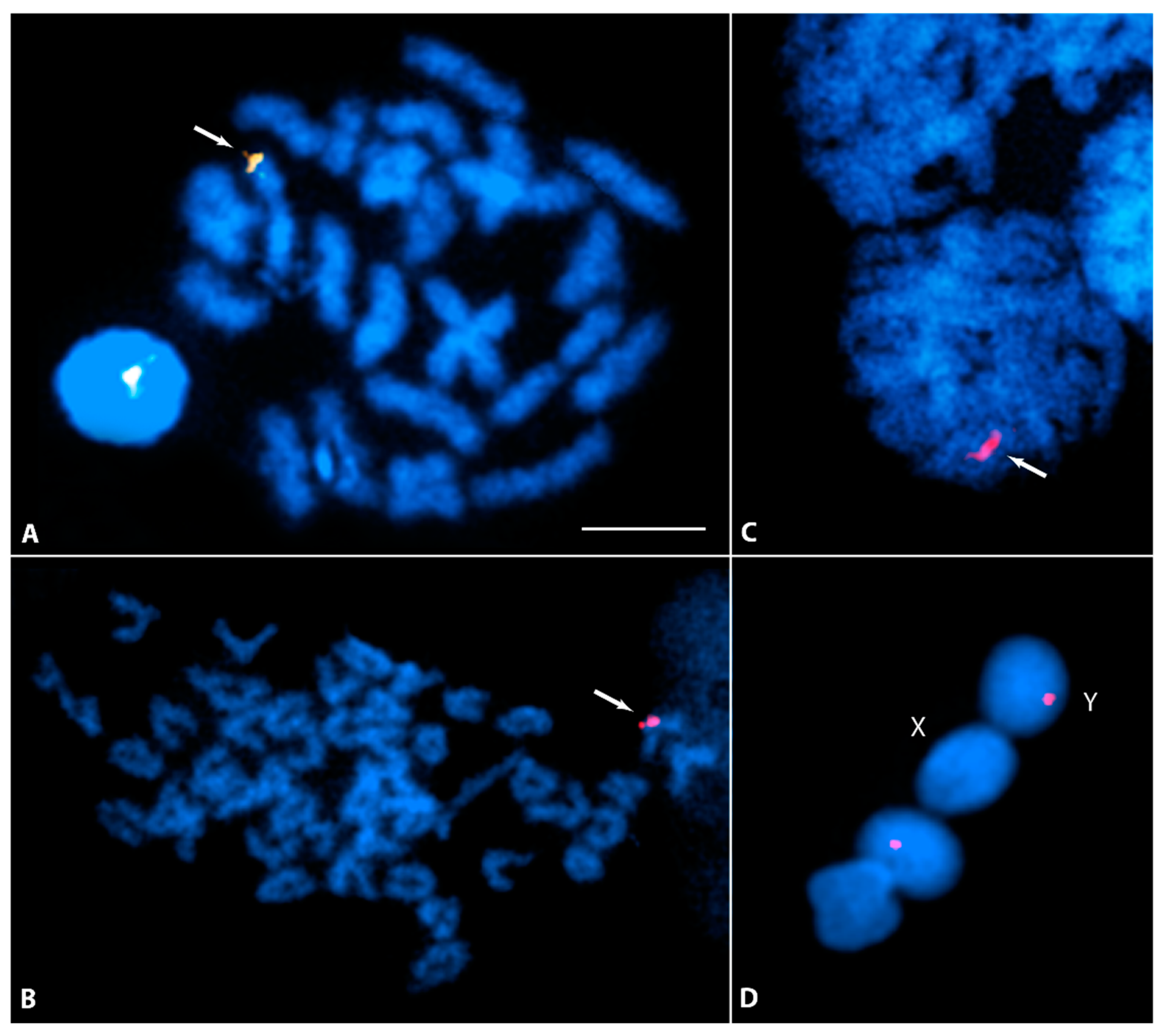
3.3. Y-Specific Hypermethylated Region
3.4. Y Chromosome-Linked Pigmentation Patterns
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| KCl | potassium chloride |
| 5-MeC | 5-methylcytosine |
| SC | synaptonemal complex |
| Ag-AS | silver-ammoniac silver |
| SNP | single-nucleotide polymorphism |
| CpG | 5′—cytosine—phosphate—guanine—3′ |
| LG12 | linkage group 12 |
| MSY- | male-specific region on Y |
| C-band- | centromeric band |
References
- Schartl, M. Sex chromosome evolution in non-mammalian vertebrates. Curr. Opin. Genet. Dev. 2004, 14, 634–641. [Google Scholar] [CrossRef] [PubMed]
- Ezaz, T.; Stiglec, R.; Veyrunes, F.; Marshall Graves, J.A. Relationships between vertebrate ZW and XY sex chromosome systems. Curr. Biol. 2006, 16, R736–R743. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gamble, T.; Zarkower, D. Sex determination. Curr. Biol. 2012, 22, R257–R262. [Google Scholar] [CrossRef] [Green Version]
- Haskins, C.P.; Haskins, E.F.; McLaughlin, J.J.A.; Hewitt, R.E. Polymorphism and population structure in Lebistes reticulatus, an ecological study. In Vertebrate Speciation; Blair, W.F., Ed.; University of Texas Press: Austin, TX, USA, 1961; pp. 320–395. [Google Scholar]
- Endler, J.A. Variation in the appearance of guppy color patterns to guppies and their predators under different visual conditions. Vis. Res. 1991, 31, 587–608. [Google Scholar] [CrossRef]
- Gordon, S.P.; Reznick, D.; Arendt, J.D.; Roughton, A.; Hernandez, M.N.O.; Bentzen, P.; Lopez-Sepulcre, A. Selection analysis on the rapid evolution of a secondary sexual trait. Proc. R. Soc. B Biol. Sci. 2015, 282, 20151244. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rice, W.R. The Accumulation of Sexually Antagonistic Genes as a Selective Agent Promoting the Evolution of Reduced Recombination between Primitive Sex-Chromosomes. Evolution 1987, 41, 911–914. [Google Scholar] [CrossRef]
- Winge, O.; Ditlevsen, E. Colour Inheritance and Sex Determination in Lebistes. Heredity 1947, 1, 65–83. [Google Scholar] [CrossRef]
- Nanda, I.; Feichtinger, W.; Schmid, M.; Schroder, J.H.; Zischler, H.; Epplen, J.T. Simple Repetitive Sequences Are Associated with Differentiation of the Sex-Chromosomes in the Guppy Fish. J. Mol. Evol. 1990, 30, 456–462. [Google Scholar] [CrossRef]
- Tripathi, N.; Hoffmann, M.; Weigel, D.; Dreyer, C. Linkage Analysis Reveals the Independent Origin of Poeciliid Sex Chromosomes and a Case of Atypical Sex Inheritance in the Guppy (Poecilia reticulata). Genetics 2009, 182, 365–374. [Google Scholar] [CrossRef] [Green Version]
- Nanda, I.; Schories, S.; Tripathi, N.; Dreyer, C.; Haaf, T.; Schmid, M.; Schartl, M. Sex chromosome polymorphism in guppies. Chromosoma 2014, 123, 373–383. [Google Scholar] [CrossRef]
- Wright, A.E.; Darolti, I.; Bloch, N.I.; Oostra, V.; Sandkam, B.; Buechel, S.D.; Kolm, N.; Breden, F.; Vicoso, B.; Mank, J.E. Convergent recombination suppression suggests role of sexual selection in guppy sex chromosome formation. Nat. Commun. 2017, 8, 14251. [Google Scholar] [CrossRef] [PubMed]
- Morris, J.; Darolti, I.; Bloch, N.I.; Wright, A.E.; Mank, J.E. Shared and Species-Specific Patterns of Nascent Y Chromosome Evolution in Two Guppy Species. Genes 2018, 9, 238. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Darolti, I.; Wright, A.E.; Sandkam, B.A.; Morris, J.; Bloch, N.I.; Farre, M.; Fuller, R.C.; Bourne, G.R.; Larkin, D.M.; Breden, F.; et al. Extreme heterogeneity in sex chromosome differentiation and dosage compensation in livebearers. Proc. Natl. Acad. Sci. USA 2019, 116, 19031–19036. [Google Scholar] [CrossRef] [Green Version]
- Darolti, I.; Wright, A.E.; Mank, J.E. Guppy Y Chromosome Integrity Maintained by Incomplete Recombination Suppression. Genome Biol. Evol. 2020, 12, 965–977. [Google Scholar] [CrossRef]
- Almeida, P.; Sandkam, B.A.; Morris, J.; Darolti, I.; Breden, F.; Mank, J.E. Divergence and Remarkable Diversity of the Y Chromosome in Guppies. Mol. Biol. Evol. 2021, 38, 619–633. [Google Scholar] [CrossRef] [PubMed]
- Bergero, R.; Gardner, J.; Bader, B.; Yong, L.; Charlesworth, D. Exaggerated heterochiasmy in a fish with sex-linked male coloration polymorphisms. Proc. Natl. Acad. Sci. USA 2019, 116, 6924–6931. [Google Scholar] [CrossRef] [Green Version]
- Charlesworth, D.; Bergero, R.; Graham, C.; Gardner, J.; Yong, L. Locating the Sex Determining Region of Linkage Group 12 of Guppy (Poecilia reticulata). G3-Genes Genom. Genet. 2020, 10, 3639–3649. [Google Scholar] [CrossRef]
- Charlesworth, D.; Zhang, Y.X.; Bergero, R.; Graham, C.; Gardner, J.; Yong, L. Using GC Content to Compare Recombination Patterns on the Sex Chromosomes and Autosomes of the Guppy, Poecilia reticulata, and Its Close Outgroup Species. Mol. Biol. Evol. 2020, 37, 3550–3562. [Google Scholar] [CrossRef]
- Fraser, B.A.; Whiting, J.R.; Paris, J.R.; Weadick, C.J.; Parsons, P.J.; Charlesworth, D.; Bergero, R.; Bemm, F.; Hoffmann, M.; Kottler, V.A.; et al. Improved Reference Genome Uncovers Novel Sex-Linked Regions in the Guppy (Poecilia reticulata). Genome Biol. Evol. 2020, 12, 1789–1805. [Google Scholar] [CrossRef]
- Kirkpatrick, M.; Sardell, J.M.; Pinto, B.J.; Dixon, G.; Peichel, C.L.; Schartl, M. Evolution of the canonical sex chromosomes of the guppy and its relatives. G3-Genes Genom. Genet. 2022, 12, jkab435. [Google Scholar] [CrossRef]
- Schories, S.; Meyer, M.K.; Schartl, M. Description of Poecilia (Acanthophacelus) obscura n. sp., (Teleostei: Poeciliidae), a new guppy species from western Trinidad, with remarks on P. wingei and the status of the “Endler’s guppy”. Zootaxa 2009, 2266, 35–50. [Google Scholar] [CrossRef]
- Traut, W.; Winking, H. Meiotic chromosomes and stages of sex chromosome evolution in fish: Zebrafish, platyfish and guppy. Chromosome Res. 2001, 9, 659–672. [Google Scholar] [CrossRef] [PubMed]
- Lisachov, A.P.; Zadesenets, K.S.; Rubtsov, N.B.; Borodin, P.M. Sex Chromosome Synapsis and Recombination in Male Guppies. Zebrafish 2015, 12, 174–180. [Google Scholar] [CrossRef] [PubMed]
- Charlesworth, D.; Bergero, R.; Graham, C.; Gardner, J.; Keegan, K. How did the guppy Y chromosome evolve? PLoS Genet. 2021, 17, e1009704. [Google Scholar] [CrossRef]
- Kallman, K.D. The Platyfish, Xiphophorus maculatus. In Handbook of Genetics; King, R., Ed.; Springer: Boston, MA, USA, 1975; Volume 4, pp. 81–132. [Google Scholar]
- Sumner, A.T. Simple Technique for Demonstrating Centromeric Heterochromatin. Exp. Cell Res. 1972, 75, 304–306. [Google Scholar] [CrossRef]
- Goodpasture, C.; Bloom, S.E. Visualization of Nucleolar Organizer Regions in Mammalian Chromosomes Using Silver Staining. Chromosoma 1975, 53, 37–50. [Google Scholar] [CrossRef]
- Baier, A.; Alsheimer, M.; Benavente, R. Synaptonemal complex protein SYCP3: Conserved polymerization properties among vertebrates. Biochim. Biophys. Acta 2007, 1774, 595–602. [Google Scholar] [CrossRef]
- Schmid, M.; Steinlein, C.; Yano, C.F.; Cioffi, M.B. Hypermethylated Chromosome Regions in Nine Fish Species with Heteromorphic Sex Chromosomes. Cytogenet. Genome Res. 2015, 147, 169–178. [Google Scholar] [CrossRef]
- Dor, L.; Shirak, A.; Kohn, Y.Y.; Gur, T.; Weller, J.I.; Zilberg, D.; Seroussi, E.; Ron, M. Mapping of the Sex Determining Region on Linkage Group 12 of Guppy (Poecilia reticulata). G3-Genes Genom. Genet. 2019, 9, 3867–3875. [Google Scholar] [CrossRef] [Green Version]
- Metzger, D.C.; Sandkam, B.A.; Darolti, I.; Mank, J.E. Rapid evolution of complete dosage compensation Poeciliids. BioRxiv 2021. [Google Scholar] [CrossRef]
- Barbin, A.; Montpellier, C.; Kokaljvokac, N.; Gibaud, A.; Niveleau, A.; Malfoy, B.; Dutrillaux, B.; Bourgeois, C.A. New Sites of Methylcytosine-Rich DNA Detected on Metaphase Chromosomes. Hum. Genet. 1994, 94, 684–692. [Google Scholar] [CrossRef] [PubMed]
- Haaf, T. The effects of 5-azacytidine and 5-azadeoxycytidine on chromosome structure and function: Implications for methylation-associated cellular processes. Pharmacol. Ther. 1995, 65, 19–46. [Google Scholar] [CrossRef]
- Yoder, J.A.; Walsh, C.P.; Bestor, T.H. Cytosine methylation and the ecology of intragenomic parasites. Trends Genet. 1997, 13, 335–340. [Google Scholar] [CrossRef]
- Weber, M.; Hellmann, I.; Stadler, M.B.; Ramos, L.; Paabo, S.; Rebhan, M.; Schubeler, D. Distribution, silencing potential and evolutionary impact of promoter DNA methylation in the human genome. Nat. Genet. 2007, 39, 457–466. [Google Scholar] [CrossRef]
- Zakharyevich, K.; Tang, S.; Ma, Y.; Hunter, N. Delineation of joint molecule resolution pathways in meiosis identifies a crossover-specific resolvase. Cell 2012, 149, 334–347. [Google Scholar] [CrossRef] [Green Version]
- Baker, S.M.; Plug, A.W.; Prolla, T.A.; Bronner, C.E.; Harris, A.C.; Yao, X.; Christie, D.M.; Monell, C.; Arnheim, N.; Bradley, A.; et al. Involvement of mouse Mlh1 in DNA mismatch repair and meiotic crossing over. Nat. Genet. 1996, 13, 336–342. [Google Scholar] [CrossRef]
- Kirpichinikov, V.S. The genetics of aquarium fish selection. In Genetic Bases of Fish Selection; Springer: Berlin/Heidelberg, Germany; New York, NY, USA, 1981; pp. 77–103. [Google Scholar]
- Lindholm, A.; Breden, F. Sex chromosomes and sexual selection in poeciliid fishes. Am. Nat. 2002, 160, S214–S224. [Google Scholar] [CrossRef]
- Gordon, S.P.; Lopez-Sepulcre, A.; Rumbo, D.; Reznick, D.N. Rapid Changes in the Sex Linkage of Male Coloration in Introduced Guppy Populations. Am. Nat. 2017, 189, 196–200. [Google Scholar] [CrossRef] [Green Version]
- Van Doorn, G.S.; Kirkpatrick, M. Turnover of sex chromosomes induced by sexual conflict. Nature 2007, 449, 909–912. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Nanda, I.; Schories, S.; Simeonov, I.; Adolfi, M.C.; Du, K.; Steinlein, C.; Alsheimer, M.; Haaf, T.; Schartl, M. Evolution of the Degenerated Y-Chromosome of the Swamp Guppy, Micropoecilia picta. Cells 2022, 11, 1118. https://doi.org/10.3390/cells11071118
Nanda I, Schories S, Simeonov I, Adolfi MC, Du K, Steinlein C, Alsheimer M, Haaf T, Schartl M. Evolution of the Degenerated Y-Chromosome of the Swamp Guppy, Micropoecilia picta. Cells. 2022; 11(7):1118. https://doi.org/10.3390/cells11071118
Chicago/Turabian StyleNanda, Indrajit, Susanne Schories, Ivan Simeonov, Mateus Contar Adolfi, Kang Du, Claus Steinlein, Manfred Alsheimer, Thomas Haaf, and Manfred Schartl. 2022. "Evolution of the Degenerated Y-Chromosome of the Swamp Guppy, Micropoecilia picta" Cells 11, no. 7: 1118. https://doi.org/10.3390/cells11071118