
Something Fishy about Siamese Fighting Fish (Betta splendens) Sex: Polygenic Sex Determination or a Newly Emerged Sex-Determining Region?

 ,
, 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Specimen Collection for Genome-Wide SNP Analysis and DNA Extraction
2.2. DArT Sequencing and Genotyping
2.3. Genetic Diversity of the Commercial Population Based on SNP Data Analysis
2.4. Marker Selection and DArT Sequencing Analysis
2.5. Comparison of Potential Sex-Linked Loci
2.6. In Silico Random Commercial Population Testing of Marker Selection and DArT Sequencing Analysis
DNA Marker Validation in Commercial Individuals
2.7. Validation of DNA Markers across Wild Siamese Fighting Fish Individuals
2.7.1. In Silico Chromosome Mapping
2.7.2. Homology Searching
2.8. Functional Annotation and Gene Ontology of Sex-Specific/Sex-Linked Loci
Determination of Repetitive Elements in Specific Regions
2.9. Comparative Genomics between Siamese Fighting Fish and Other Vertebrates
3. Results
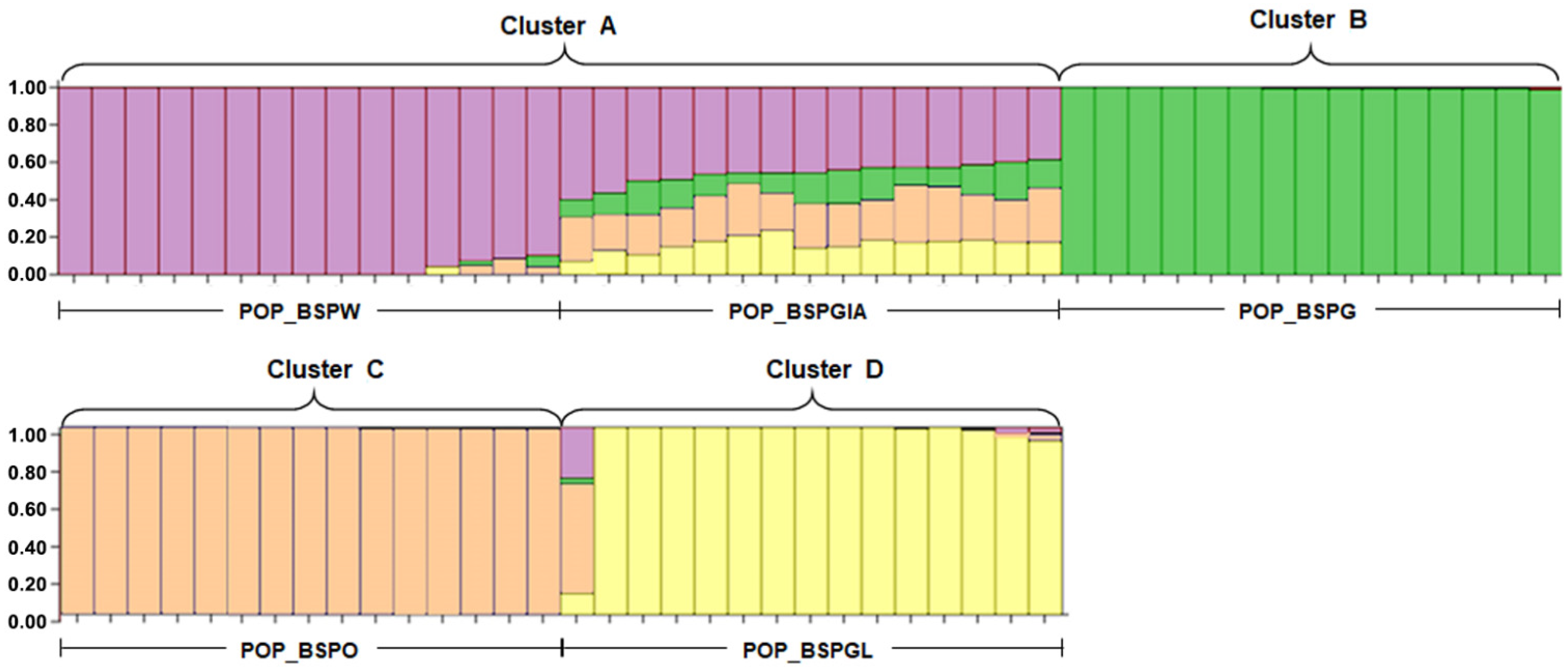
3.1. Population Structure and Genetic Diversity Analysis of Commercial Siamese Fighting Fish Populations
3.2. Determination of the Sex System and Identification of Sex-Specific/Sex-Linked Loci in Siamese Fighting Fish
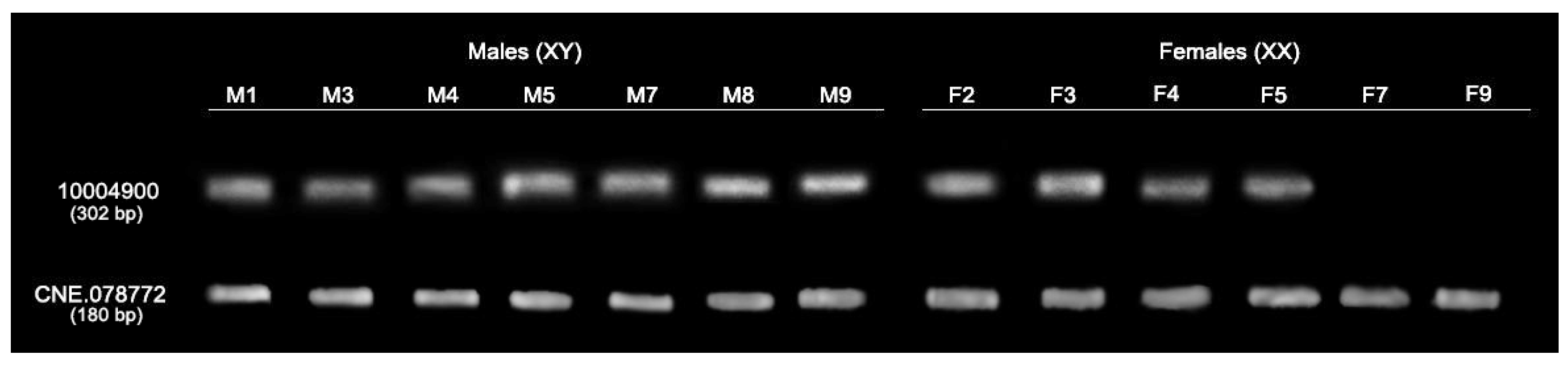
3.3. Validation of Sex-Specific Loci with DNA Markers
3.4. Genetic Variability across Wild Siamese Fighting Fish Individuals Based on Microsatellite Data
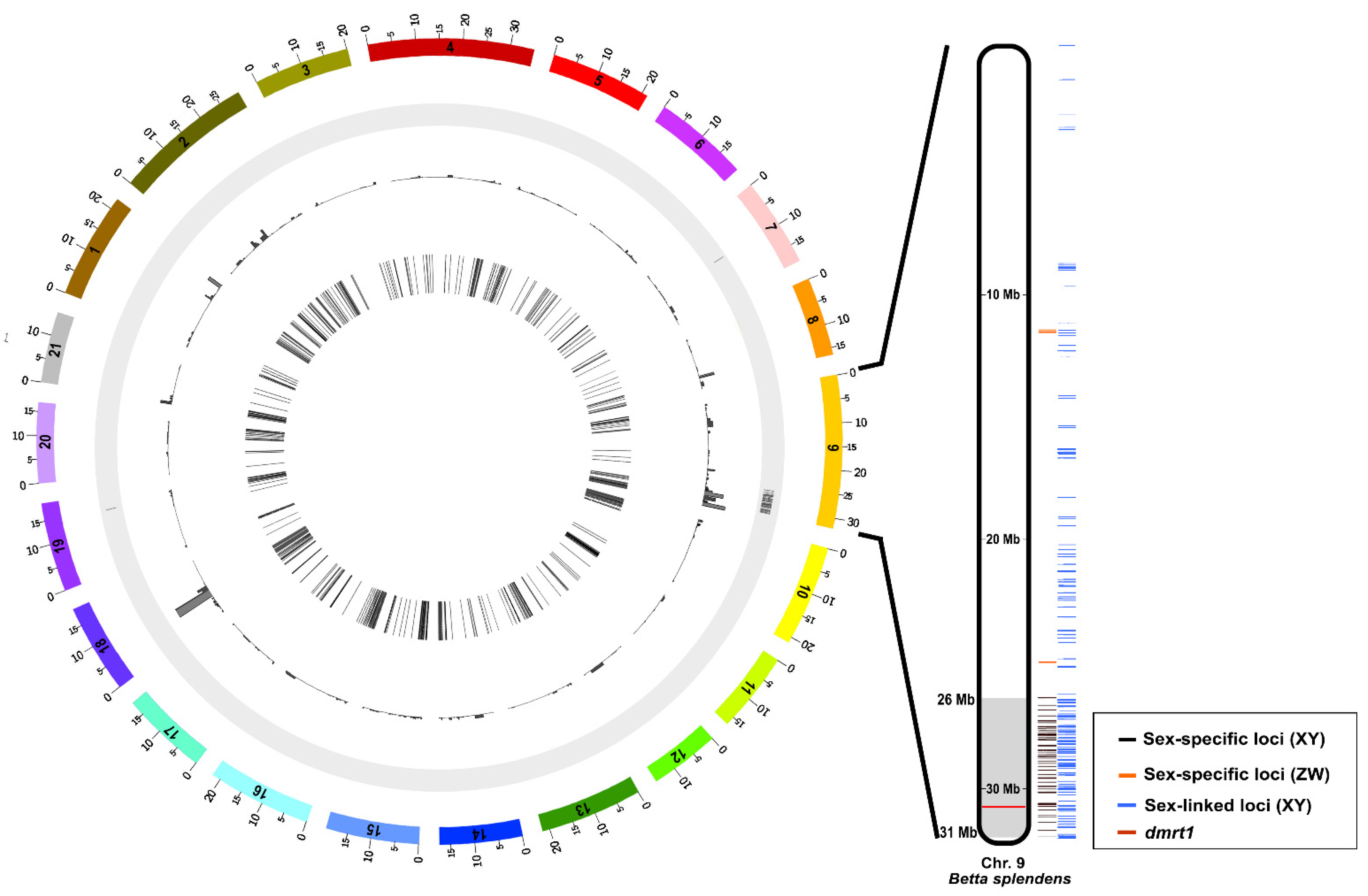
3.5. Chromosome Localization of Sex-Specific and Sex-Linked Loci
3.6. Homology of Putative Sex-Specific/Linked Loci
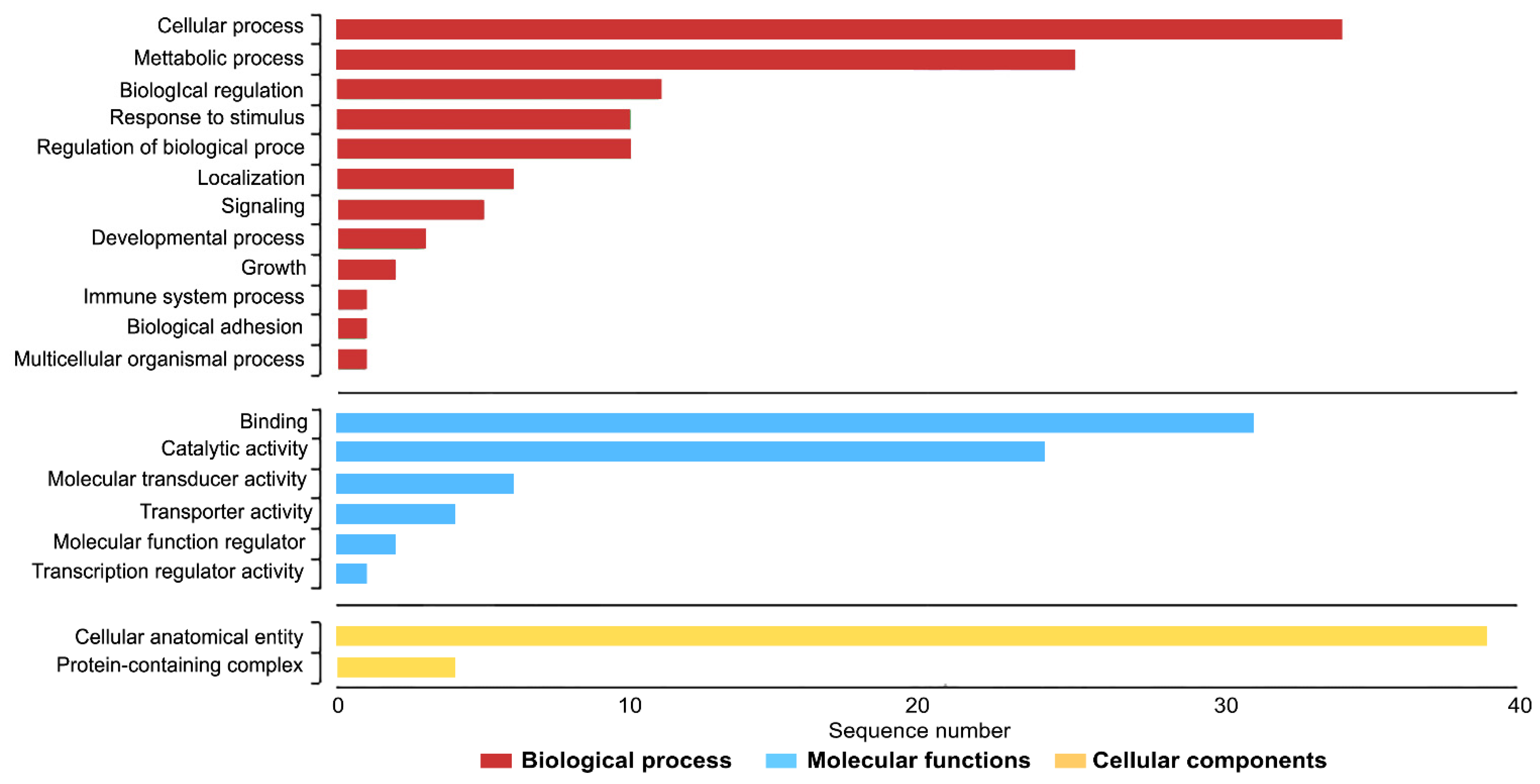
3.7. Functional Classification and Enrichment Analysis
3.8. Annotation of Repetitive Elements in the Specific Region
3.9. Linkage Homology of Siamese Fighting Fish and Other Vertebrates
4. Discussion
4.1. Is Only Chromosome 9 of Siamese Fighting Fish Involved in a SDR?
4.2. Co-Existence of XY and ZW Sex Determination in Siamese Fighting Fish
4.3. Genetic Admixture and Introgression of Siamese Fighting Fish Might Complicate Findings
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Regan, C.T. The Asiatic fishes of the family Anabantidae. Proc. Zool. Soc. Lond. 1910, B1909, 767–787. [Google Scholar]
- Rhoad, K.D.; Kalat, J.W.; Klopfer, P.H. Aggression and avoidance byBetta splendens toward natural and artificial stimuli. Learn. Behav. 1975, 3, 271–276. [Google Scholar] [CrossRef] [Green Version]
- Srikulnath, K.; Singchat, W.; Laopichienpong, N.; Ahmad, S.F.; Jehangir, M.; Subpayakom, N.; Suntronpong, A.; Jangtarwan, K.; Pongsanarm, T.; Panthum, T.; et al. Overview of the betta fish genome regarding species radiation, parental care, behavioral aggression, and pigmentation model relevant to humans. Genes Genom. 2021, 43, 91–104. [Google Scholar] [CrossRef]
- Smith, H.M. The freshwater fishes of Siam, or Thailand. Bull. US Natl. Mus. 1945, 188, 9. [Google Scholar]
- Lucas, G.A. A Study of Variation in the Siamese Fighting Fish, Betta Splendens, with Emphasis on Color Mutants and the Problem of Sex Determination; Iowa State University: Ames, IA, USA, 1968. [Google Scholar]
- Simpson, M. The Display of the Siamese Fighting Fish, Betta splendens. Anim. Behav. Monogr. 1968, 1, 1–73. [Google Scholar] [CrossRef]
- U.S. Fish and Wildlife Service. Siamese Fighting Fish (Betta splendens) Ecological Risk Screening Summary, USA. 2019. Available online: https://eol.org/pages/211488/articles (accessed on 6 January 2022).
- Department of Fisheries. Fisheries single window data system. In Fish Quarantine and Inspection Division; Department of Fisheries: Bangkok, Thailand, 2019. [Google Scholar]
- Budd, A.M.; Banh, Q.Q.; Domingos, J.A.; Jerry, D.R. Sex Control in Fish: Approaches, Challenges and Opportunities for Aquaculture. J. Mar. Sci. Eng. 2015, 3, 329–355. [Google Scholar] [CrossRef] [Green Version]
- Saekhow, S.; Thongprajukaew, K.; Phromkunthong, W. Blue aquarium background is appropriate for rearing male Siamese fighting fish (Betta splendens). Aquac. Int. 2019, 27, 891–903. [Google Scholar] [CrossRef]
- Svärdson, G.; Wickbom, T. The Chromosomes of Two Species of Anabantidae (Teleostei), with a New Case of Sex Reversal. Hereditas 2010, 28, 212–216. [Google Scholar] [CrossRef]
- Srikulnath, K.; Swatdipong, A.; Uno, Y.; Matsubara, K.; Peyachoknagul, S.; Matsuda, Y. Retroelement compartmentalization of Rex1, Rex3 and Rex6 in Siamese fighting fish genome. In Proceedings of the 17th National Genetic Conference, Chiang Mai, Thailand, 7–9 April 2011. [Google Scholar]
- Kwon, Y.M.; Vranken, N.; Hoge, C.; Lichak, M.R.; Norovich, A.L.; Francis, K.X.; Camacho-Garcia, J.; Bista, I.; Wood, J.; McCarthy, S.; et al. Genomic consequences of domestication of the Siamese fighting fish. Sci. Adv. 2022, 8, eabm4950. [Google Scholar] [CrossRef]
- Wang, L.; Sun, F.; Wan, Z.Y.; Yang, Z.; Tay, Y.X.; Lee, M.; Ye, B.; Wen, Y.; Meng, Z.; Fan, B.; et al. Transposon-induced epigenetic silencing in the X chromosome as a novel form of dmrt1 expression regulation during sex determination in the fighting fish. BMC Biol. 2022, 20, 5. [Google Scholar] [CrossRef]
- Purcell, C.M.; Seetharam, A.; Snodgrass, O.; Garcia, S.O.; Hyde, J.R.; Severin, A.J. Insights into teleost sex determination from the Seriola dorsalis genome assembly. BMC Genom. 2018, 19, 31. [Google Scholar] [CrossRef] [Green Version]
- Nguyen, D.H.M.; Panthum, T.; Ponjarat, J.; Laopichienpong, N.; Kraichak, E.; Singchat, W.; Ahmad, S.F.; Muangmai, N.; Peyachoknagul, S.; Na-Nakorn, U.; et al. An Investigation of ZZ/ZW and XX/XY Sex Determination Systems in North African Catfish (Clarias gariepinus, Burchell, 1822). Front. Genet. 2021, 11, 562856. [Google Scholar] [CrossRef]
- Nguyen, D.H.M.; Panthum, T.; Ponjarat, J.; Laopichienpong, N.; Kraichak, E.; Singchat, W.; Ahmad, S.F.; Muangmai, N.; Peyachoknagul, S.; Na-Nakorn, U.; et al. Genome-wide SNP analysis suggests male heterogamety in bighead catfish (Clarias macrocephalus, Günther, 1864). Aquaculture 2021, 543, 737005. [Google Scholar] [CrossRef]
- Nguyen, D.H.M.; Ponjarat, J.; Laopichienpong, N.; Panthum, T.; Singchat, W.; Ahmad, S.F.; Kraichak, E.; Muangmai, N.; Duengkae, P.; Peyachoknagul, S.; et al. Genome-Wide SNP Analysis of Hybrid Clariid Fish Reflects the Existence of Polygenic Sex-Determination in the Lineage. Front. Genet. 2022, 13, 789573. [Google Scholar] [CrossRef]
- Roberts, N.B.; Juntti, S.A.; Coyle, K.P.; Dumont, B.L.; Stanley, M.K.; Ryan, A.Q.; Fernald, R.D.; Roberts, R.B. Polygenic sex determination in the cichlid fish Astatotilapia burtoni. BMC Genom. 2016, 17, 835. [Google Scholar] [CrossRef] [Green Version]
- Von Schmidt, H. Geschlectsumwandlungen bei labyrinthfischen. Das Aquarium 1930, 9, 161–163. [Google Scholar]
- Noble, G.K. The experimental animal from the naturalist’s point of view. Am. Nat. 1939, 73, 113–126. [Google Scholar] [CrossRef]
- Kaiser, P.; Schmidt, E. Vollkommene Geschlechtsumwandlung beim weiblichen siamesischen Kampffisch Betta splendens. Zool. Anz. 1951, 146, 66–73. [Google Scholar]
- Schmidt, E. Sex reversal in spayed female bettas. Trop. Fish Hobbyist 1962, 21, 24. [Google Scholar]
- Becker, P. Experimentell Induzierter Geschlechtswechsel bei Anabantiden. Ph.D. Dissertation, University of Mainz, Mainz, Germany, 1969. [Google Scholar]
- Becker, P.; Roland, H.; Reinboth, R. An unusual approach to experimental sex inversion in the teleost fish Betta and Macropodus. In Intersexuality in the Animal Kingdom; Reinboth, R., Ed.; Springer: New York, NY, USA, 1975. [Google Scholar]
- Sutherland, B.J.; Rico, C.; Audet, C.; Bernatchez, L. Sex chromosome evolution, heterochiasmy, and physiological QTL in the salmonid brook charr Salvelinus fontinalis. G3 2017, 7, 2749–2762. [Google Scholar] [CrossRef] [Green Version]
- Koboldt, D.C.; Steinberg, K.M.; Larson, D.; Wilson, R.K.; Mardis, E.R. The Next-Generation Sequencing Revolution and Its Impact on Genomics. Cell 2013, 155, 27–38. [Google Scholar] [CrossRef] [Green Version]
- Koomgun, T.; Laopichienpong, N.; Singchat, W.; Panthum, T.; Phatcharakullawarawat, R.; Kraichak, E.; Sillapaprayoon, S.; Ahmad, S.F.; Muangmai, N.; Peyachoknagul, S.; et al. Genome Complexity Reduction High-Throughput Genome Sequencing of Green Iguana (Iguana iguana) Reveal a Paradigm Shift in Understanding Sex-Chromosomal Linkages on Homomorphic X and Y Sex Chromosomes. Front. Genet. 2020, 11, 556267. [Google Scholar] [CrossRef]
- Laopichienpong, N.; Kraichak, E.; Singchat, W.; Sillapaprayoon, S.; Muangmai, N.; Suntrarachun, S.; Baicharoen, S.; Peyachoknagul, S.; Chanhome, L.; Ezaz, T.; et al. Genome-wide SNP analysis of Siamese cobra (Naja kaouthia) reveals the molecular basis of transitions between Z and W sex chromosomes and supports the presence of an ancestral super-sex chromosome in amniotes. Genomics 2021, 113, 624–636. [Google Scholar] [CrossRef]
- Panthum, T.; Laopichienpong, N.; Kraichak, E.; Singchat, W.; Nguyen, D.H.M.; Ariyaraphong, N.; Ahmad, S.F.; Muangmai, N.; Duengkae, P.; Peyachoknagul, S.; et al. The Snakeskin Gourami (Trichopodus pectoralis) Tends to Exhibit XX/XY Sex Determination. Fishes 2021, 6, 43. [Google Scholar] [CrossRef]
- Suntronpong, A.; Panthum, T.; Laopichienpong, N.; Nguyen, D.H.M.; Kraichak, E.; Singchat, W.; Ariyaraphong, N.; Ahmad, S.F.; Muangmai, N.; Duengkae, P.; et al. Implications of genome-wide single nucleotide polymorphisms in jade perch (Scortum barcoo) reveals the putative XX/XY sex-determination system, facilitating a new chapter of sex control in aquaculture. Aquaculture 2021, 548, 737587. [Google Scholar] [CrossRef]
- Fan, G.; Chan, J.; Ma, K.; Yang, B.; Zhang, H.; Yang, X.; Shi, C.; Law, H.C.H.; Ren, Z.; Xu, Q.; et al. Chromosome-level reference genome of the Siamese fighting fish Betta splendens, a model species for the study of aggression. GigaScience 2018, 7, giy087. [Google Scholar] [CrossRef] [Green Version]
- Saint-Pé, K.; Blanchet, S.; Tissot, L.; Poulet, N.; Plasseraud, O.; Loot, G.; Veyssière, C.; Prunier, J.G. Genetic admixture between captive-bred and wild individuals affects patterns of dispersal in a brown trout (Salmo trutta) population. Conserv. Genet. 2018, 19, 1269–1279. [Google Scholar] [CrossRef] [Green Version]
- Beer, S.D.; Cornett, S.; Austerman, P.; Trometer, B.; Hoffman, T.; Bartron, M.L. Genetic diversity, admixture, and hatchery influence in Brook Trout (Salvelinus fontinalis) throughout western New York State. Ecol. Evol. 2019, 9, 7455–7479. [Google Scholar] [CrossRef] [Green Version]
- Witte, K.; Schmidt, J. Betta brownorum, a new species of anabantoids (Teleostei; Belontiidae) from Northwestern Borneo, with a key to the genus. Ichthyol. Explor. Freshw. 1992, 2, 305–330. [Google Scholar]
- Ramos, A.; Gonçalves, D. Artificial selection for male winners in the Siamese fighting fish Betta splendens correlates with high female aggression. Front. Zool. 2019, 16, 34. [Google Scholar] [CrossRef] [Green Version]
- Zhang, W.; Wang, H.; Brandt, D.Y.; Hu, B.; Sheng, J.; Wang, M.; Luo, H.; Guo, S.; Sheng, G.; Zeng, Q.; et al. The genetic architecture of phenotypic diversity in the betta fish (Betta splendens). bioRxiv 2021. [Google Scholar] [CrossRef]
- Ostrow, M. Bettas; T.F.H. Publications, Inc.: Neptune City, NJ, USA, 1989. [Google Scholar]
- Casselman, S.J.; Schulte-Hostedde, A.I. Reproductive roles predict sexual dimorphism in internal and external morphology of lake whitefish, Coregonus clupeaformis. Ecol. Freshw. Fish 2004, 13, 217–222. [Google Scholar] [CrossRef]
- Kitano, J.; Mori, S.; Peichel, C.L. Reduction of sexual dimorphism in stream-resident forms of three-spined stickleback Gasterosteus aculeatus. J. Fish Biol. 2012, 80, 131–146. [Google Scholar] [CrossRef]
- Supikamolseni, A.; Ngaoburanawit, N.; Sumontha, M.; Chanhome, L.; Suntrarachun, S.; Peyachoknagul, S.; Srikulnath, K. Molecular barcoding of venomous snakes and species-specific multiplex PCR assay to identify snake groups for which antivenom is available in Thailand. Genet. Mol. Res. 2015, 14, 13981–13997. [Google Scholar] [CrossRef]
- Jaccoud, D.; Peng, K.; Feinstein, D.; Kilian, A. Diversity Arrays: A solid state technology for sequence information independent genotyping. Nucleic Acids Res. 2001, 29, E25. [Google Scholar] [CrossRef] [Green Version]
- Kilian, A.; Wenzl, P.; Huttner, E.; Carling, J.; Xia, L.; Blois, H.; Caig, V.; Heller-Uszynska, K.; Jaccoud, D.; Hopper, C.; et al. Diversity arrays technology: A generic genome profiling technology on open platforms. Methods Mol. Biol. 2012, 888, 67–89. [Google Scholar] [CrossRef]
- Ren, R.; Ray, R.; Li, P.; Xu, J.; Zhang, M.; Liu, G.; Yao, X.; Kilian, A.; Yang, X. Construction of a high-density DArTseq SNP-based genetic map and identification of genomic regions with segregation distortion in a genetic population derived from a cross between feral and cultivated-type watermelon. Mol. Genet. Genom. 2015, 290, 1457–1470. [Google Scholar] [CrossRef]
- Pritchard, J.K.; Stephens, M.; Donnelly, P. Inference of population structure using multilocus genotype data. Genetics 2000, 155, 945–959. [Google Scholar] [CrossRef]
- Peakall, R.; Smouse, P.E. GenAlEx 6.5: Genetic analysis in Excel. Population genetic software for teaching and research—An update. Bioinformatics 2012, 28, 2537–2539. [Google Scholar] [CrossRef] [Green Version]
- Alam, M.; Neal, J.; O’Connor, K.; Kilian, A.; Topp, B. Ultra-high-throughput DArTseq-based silicoDArT and SNP markers for genomic studies in macadamia. PLoS ONE 2018, 13, e0203465. [Google Scholar] [CrossRef] [Green Version]
- Evanno, G.; Regnaut, S.; Goudet, J. Detecting the number of clusters of individuals using the software structure: A simulation study. Mol. Ecol. 2005, 14, 2611–2620. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Earl, D.A.; von Holdt, B.M. Structure Harvester: A website and program for visualizing Structure output and implementing the Evanno method. Conserv. Genet. Resour. 2012, 4, 359–361. [Google Scholar] [CrossRef]
- Jombart, T.; Ahmed, I. adegenet 1.3-1: New tools for the analysis of genome-wide SNP data. Bioinformatics 2011, 27, 3070–3071. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2021. [Google Scholar]
- Sven, K.; Klaus, R. HapEstXXR: Multi-Locus Stepwise Regression. 2019. Available online: https://rdrr.io/cran/HapEstXXR/man/coding.baseline.allele.html (accessed on 6 January 2022).
- Gruber, B.; Georges, A. dartR: Importing and Analysing SNP and Silicodart Data Generated by Genome-Wide Restriction Fragment Analysis. 2021. Available online: https://cran.r-project.org/web/packages/dartR/index.html (accessed on 6 January 2022).
- Roff, D.A. Introduction to Computer-Intensive Methods of Data Analysis in Biology; Cambridge University Press: Cambridge, UK, 2006. [Google Scholar] [CrossRef]
- Kendall, M.G.; Buckland, W.G. A Dictionary of Statistical Terms, 4th ed.; Longmans: London, UK, 1982. [Google Scholar]
- Chailertrit, V.; Swatdipong, A.; Peyachoknagul, S.; Salaenoi, J.; Srikulnath, K. Isolation and characterization of novel microsatellite markers from Siamese fighting fish (Betta splendens, Osphronemidae, Anabantoidei) and their transferability to related species, B. smaragdina and B. imbellis. Genet. Mol. Res. 2014, 13, 7157–7162. [Google Scholar] [CrossRef]
- van Oosterhout, C.; Hutchinson, W.F.; Wills, D.P.M.; Shipley, P. micro-checker: Software for identifying and correcting genotyping errors in microsatellite data. Mol. Ecol. Notes 2004, 4, 535–538. [Google Scholar] [CrossRef]
- Park, S.D.E. Trypanotolerance in West African Cattle and the Population Genetic Effects of Selection. Ph.D. Thesis, University of Dublin, Dublin, Ireland, 2001. [Google Scholar]
- Excoffier, L.; Lischer, H.E.L. Arlequin suite ver 3.5: A new series of programs to perform population genetics analyses under Linux and Windows. Mol. Ecol. Resour. 2010, 10, 564–567. [Google Scholar] [CrossRef]
- Prost, S.; Petersen, M.; Grethlein, M.; Hahn, S.J.; Kuschik-Maczollek, N.; Olesiuk, M.E.; Reschke, J.-O.; Schmey, T.E.; Zimmer, C.; Gupta, D.K.; et al. Improving the Chromosome-Level Genome Assembly of the Siamese Fighting Fish (Betta splendens) in a University Master’s Course. G3 2020, 10, 2179–2183. [Google Scholar] [CrossRef]
- Naquin, D.; D’Aubenton-Carafa, Y.; Thermes, C.; Silvain, M. CIRCUS: A package for Circos display of structural genome variations from paired-end and mate-pair sequencing data. BMC Bioinform. 2014, 15, 198. [Google Scholar] [CrossRef] [Green Version]
- Parisseaux, B.; Bernardo, R. In silico mapping of quantitative trait loci in maize. Theor. Appl. Genet. 2004, 109, 508–514. [Google Scholar] [CrossRef]
- Temminck, C.J.; Schlegel, H. Oryzias Latipes (Temminck & Schlegel, 1846). Available online: https://www.gbif.org/species/2368377 (accessed on 6 January 2022).
- Kasahara, M.; Naruse, K.; Sasaki, S.; Nakatani, Y.; Qu, W.; Ahsan, B.; Yamada, T.; Nagayasu, Y.; Doi, K.; Kasai, Y.; et al. The medaka draft genome and insights into vertebrate genome evolution. Nature 2007, 447, 714–719. [Google Scholar] [CrossRef] [Green Version]
- Hamilton. Zebrafish (Danio rerio). 1822. Available online: https://www.gbif.org/species/9797255 (accessed on 6 January 2022).
- Broughton, R.E.; Milam, J.E.; Roe, B.A. The Complete Sequence of the Zebrafish (Danio rerio) Mitochondrial Genome and Evolutionary Patterns in Vertebrate Mitochondrial DNA. Genome Res. 2001, 11, 1958–1967. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Linnaeus. Chicken (Gallus gallus). 1758. Available online: https://www.gbif.org/species/9326020 (accessed on 6 January 2022).
- International Chicken Genome Sequencing Consortium. Sequence and comparative analysis of the chicken genome provide unique perspectives on vertebrate evolution. Nature 2004, 432, 695–716. [Google Scholar] [CrossRef] [PubMed]
- Bao, W.; Kojima, K.K.; Kohany, O. Repbase Update, a database of repetitive elements in eukaryotic genomes. Mob. DNA 2015, 6, 11. [Google Scholar] [CrossRef] [Green Version]
- O’Leary, N.A.; Wright, M.W.; Brister, J.R.; Ciufo, S.; Haddad, D.; McVeigh, R.; Rajput, B.; Robbertse, B.; Smith-White, B.; Ako-Adjei, D.; et al. Reference sequence (RefSeq) database at NCBI: Current status, taxonomic expansion, and functional annotation. Nucleic Acids Res. 2016, 44, D733–D745. [Google Scholar] [CrossRef] [Green Version]
- Lima, T.; Auchincloss, A.; Coudert, E.; Keller, G.; Michoud, K.; Rivoire, C.; Bulliard, V.; de Castro, E.; Lachaize, C.; Baratin, D.; et al. HAMAP: A database of completely sequenced microbial proteome sets and manually curated microbial protein families in UniProtKB/Swiss-Prot. Nucleic Acids Res. 2009, 37, D471–D478. [Google Scholar] [CrossRef] [PubMed]
- Al-Qurainy, F.; Alshameri, A.; Gaafar, A.-R.; Khan, S.; Nadeem, M.; Alameri, A.A.; Tarroum, M.; Ashraf, M. Comprehensive Stress-Based De Novo Transcriptome Assembly and Annotation of Guar (Cyamopsis tetragonoloba (L.) Taub.): An Important Industrial and Forage Crop. Int. J. Genom. 2019, 2019, 7295859. [Google Scholar] [CrossRef] [Green Version]
- Ou, S.; Su, W.; Liao, Y.; Chougule, K.; Agda, J.R.A.; Hellinga, A.J.; Lugo, C.S.B.; Elliott, T.A.; Ware, D.; Peterson, T.; et al. Benchmarking transposable element annotation methods for creation of a streamlined, comprehensive pipeline. Genome Biol. 2019, 20, 275. [Google Scholar] [CrossRef] [Green Version]
- Smit, A.F.A.; Hubley, R. RepeatMasker. 2008. Available online: http://www.repeatmasker.org (accessed on 6 January 2022).
- Manee, M.M.; Jackson, J.; Bergman, C.M. Conserved Noncoding Elements Influence the Transposable Element Landscape in Drosophila. Genome Biol. Evol. 2018, 10, 1533–1545. [Google Scholar] [CrossRef] [Green Version]
- Cabanettes, F.; Klopp, C. D-GENIES: Dot plot large genomes in an interactive, efficient and simple way. PeerJ 2018, 6, e4958. [Google Scholar] [CrossRef]
- Lee, J.; Hong, W.-Y.; Cho, M.; Sim, M.; Lee, D.; Ko, Y.; Kim, J. Synteny Portal: A web-based application portal for synteny block analysis. Nucleic Acids Res. 2016, 44, W35–W40. [Google Scholar] [CrossRef] [Green Version]
- Nguyen, N.T.T.; Vincens, P.; Crollius, H.R.; Louis, A. Genomicus 2018: Karyotype evolutionary trees and on-the-fly synteny computing. Nucleic Acids Res. 2018, 46, D816–D822. [Google Scholar] [CrossRef] [PubMed]
- Gray. The Western Clawed Frog (Xenopus tropicalis). 1864. Available online: https://www.gbif.org/species/8784293 (accessed on 6 January 2022).
- Gamble, T.; Coryell, J.; Ezaz, T.; Lynch, J.; Scantlebury, D.P.; Zarkower, D. Restriction Site-Associated DNA Sequencing (RAD-seq) Reveals an Extraordinary Number of Transitions among Gecko Sex-Determining Systems. Mol. Biol. Evol. 2015, 32, 1296–1309. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sember, A.; Pappová, M.; Forman, M.; Nguyen, P.; Marec, F.; Dalíková, M.; Divišová, K.; Doležálková-Kaštánková, M.; Zrzavá, M.; Sadílek, D.; et al. Patterns of Sex Chromosome Differentiation in Spiders: Insights from Comparative Genomic Hybridisation. Genes 2020, 11, 849. [Google Scholar] [CrossRef]
- Ezaz, T.; Srikulnath, K.; Graves, J.A.M. Origin of Amniote Sex Chromosomes: An Ancestral Super-Sex Chromosome, or Common Requirements? J. Hered. 2016, 108, 94–105. [Google Scholar] [CrossRef] [PubMed]
- Singchat, W.; Ahmad, S.F.; Sillapaprayoon, S.; Muangmai, N.; Duengkae, P.; Peyachoknagul, S.; O’Connor, R.E.; Griffin, D.K.; Srikulnath, K. Partial Amniote Sex Chromosomal Linkage Homologies Shared on Snake W Sex Chromosomes Support the Ancestral Super-Sex Chromosome Evolution in Amniotes. Front. Genet. 2020, 11, 948. [Google Scholar] [CrossRef] [PubMed]
- Singchat, W.; Ahmad, S.F.; Laopichienpong, N.; Suntronpong, A.; Panthum, T.; Griffin, D.K.; Srikulnath, K. Snake W Sex Chromosome: The Shadow of Ancestral Amniote Super-Sex Chromosome. Cells 2020, 9, 2386. [Google Scholar] [CrossRef] [PubMed]
- Linnaeus. Atlantic Salmon (Salmo salar). 1758. Available online: https://www.gbif.org/species/7595433 (accessed on 21 January 2022).
- Gabián, M.; Morán, P.; Fernández, A.I.; Villanueva, B.; Chtioui, A.; Kent, M.P.; Covelo-Soto, L.; Fernández, A.; Saura, M. Identification of genomic regions regulating sex determination in Atlantic salmon using high density SNP data. BMC Genom. 2019, 20, 764. [Google Scholar] [CrossRef]
- Linnaeus. Nile Tilapia (Oreochromis niloticus). 1758. Available online: https://www.gbif.org/species/4285694 (accessed on 6 January 2022).
- Triay, C.; Conte, M.A.; Baroiller, J.-F.; Bezault, E.; Clark, F.E.; Penman, D.J.; Kocher, T.D.; D’Cotta, H. Structure and Sequence of the Sex Determining Locus in Two Wild Populations of Nile Tilapia. Genes 2020, 11, 1017. [Google Scholar] [CrossRef]
- Zhou, Q. A swimy locus on Y chromosome of the platyfish (Xiphophorus maculatus) is derived from a novel DNA transposon Zisupton. Gene 2012, 503, 254–259. [Google Scholar] [CrossRef]
- Allshire, R.; Madhani, H.D. Ten principles of heterochromatin formation and function. Nat. Rev. Mol. Cell Biol. 2018, 19, 229–244. [Google Scholar] [CrossRef]
- Srikulnath, K.; Ahmad, S.F.; Singchat, W.; Panthum, T. Do Ty3/Gypsy Transposable Elements Play Preferential Roles in Sex Chromosome Differentiation? Life 2022, 12, 522. [Google Scholar] [CrossRef] [PubMed]
- Kuznetsova, I.S.; Thevasagayam, N.M.; Sridatta, P.S.R.; Komissarov, A.S.; Saju, J.M.; Ngoh, S.Y.; Jiang, J.; Shen, X.; Orbãn, L.; Orban, L. Primary analysis of repeat elements of the Asian seabass (Lates calcarifer) transcriptome and genome. Front. Genet. 2014, 5, 223. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lesson, R.P. Catalogue des Reptiles qui font partie d’une Collection zoologique recueille dans l’Inde continentale ou en Afrique, et apportée en France par M. Lamare-Piquot. Bull. Sci. Nat. Geol. 1831, 25, 119–123. [Google Scholar]
- Thongchum, R.; Singchat, W.; Laopichienpong, N.; Tawichasri, P.; Kraichak, E.; Prakhongcheep, O.; Sillapaprayoon, S.; Muangmai, N.; Baicharoen, S.; Suntrarachun, S.; et al. Diversity of PBI-DdeI satellite DNA in snakes correlates with rapid independent evolution and different functional roles. Sci. Rep. 2019, 9, 15459. [Google Scholar] [CrossRef] [Green Version]
- Günther, A.C.L.G. Catalogue of the fishes in the British Museum. In Catalogue of the Physostomi, Containing the Families Siluridae, Characinidae, Haplochitonidae, Sternoptychidae, Scopelidae, Stomiatidae in the Collection of the British Museum; Taylor & Francis: London, UK, 1864; Volume 5, pp. 1–455. [Google Scholar]
- Schartl, M.; Schories, S.; Wakamatsu, Y.; Nagao, Y.; Hashimoto, H.; Bertin, C.; Mourot, B.; Schmidt, C.; Wilhelm, D.; Centanin, L.; et al. Sox5 is involved in germ-cell regulation and sex determination in medaka following co-option of nested transposable elements. BMC Biol. 2018, 16, 16. [Google Scholar] [CrossRef] [Green Version]
- Nanda, I.; Kondo, M.; Hornung, U.; Asakawa, S.; Winkler, C.; Shimizu, A.; Shan, Z.; Haaf, T.; Shimizu, N.; Shima, A.; et al. A duplicated copy of DMRT1 in the sex-determining region of the Y chromosome of the medaka, Oryzias latipes. Proc. Natl. Acad. Sci. USA 2002, 99, 11778–11783. [Google Scholar] [CrossRef] [Green Version]
- Heckel, J.J. Eine neue Gattung von Poecilien mit rochenartigem Anklammerungs-Organe. Sitz. Kais. Akad. Wiss. Wien. 1848, 1, 289–303. [Google Scholar]
- Linnaeus. Papaya (Carica papaya). 1758. Available online: https://www.gbi.f.org/species/164003120 (accessed on 6 January 2022).
- Linnaeus, C. Systema naturae sive regna tria naturae, secundum classes, ordines, genera, species, cum characteribus, differentiis, synonymis, locis. Laurentii Salvii Holmiae. Ed. Duodecima Reformata 1758, 1, 823. [Google Scholar]
- Franchini, P.; Jones, J.C.; Xiong, P.; Kneitz, S.; Gompert, Z.; Warren, W.C.; Walter, R.B.; Meyer, A.; Schartl, M. Long-term experimental hybridisation results in the evolution of a new sex chromosome in swordtail fish. Nat. Commun. 2018, 9, 5136. [Google Scholar] [CrossRef]
- Gamble, T. Using RAD-seq to recognize sex-specific markers and sex chromosome systems. Mol. Ecol. 2016, 25, 2114–2116. [Google Scholar] [CrossRef] [Green Version]
- Jiang, S.; Ma, X.; Li, T.; Zhu, C.; You, X. Developing Single Nucleotide Polymorphisms for Identification of Cod Products by RAD-Seq. Animals 2020, 10, 423. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Harris, A.; Siggers, P.; Corrochano, S.; Warr, N.; Sagar, D.; Grimes, D.T.; Suzuki, M.; Burdine, R.D.; Cong, F.; Koo, B.-K.; et al. ZNRF3 functions in mammalian sex determination by inhibiting canonical WNT signaling. Proc. Natl. Acad. Sci. USA 2018, 115, 5474–5479. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Günther, A. On a collection of fishes from Chefoo, north China. Ann. Mag. Nat. Hist. 1873, 12, 377–380. [Google Scholar] [CrossRef] [Green Version]
- Nanda, I.; Shan, Z.; Schartl, M.; Burt, D.; Koehler, M.; Nothwang, H.-G.; Grutzner, F.; Paton, I.R.; Windsor, D.; Dunn, I.; et al. 300 million years of conserved synteny between chicken Z and human chromosome 9. Nat. Genet. 1999, 21, 258–259. [Google Scholar] [CrossRef] [PubMed]
- Matsuda, M.; Nagahama, Y.; Shinomiya, A.; Sato, T.; Matsuda, C.; Kobayashi, T.; Morrey, C.E.; Shibata, N.; Asakawa, S.; Shimizu, N. DMY is a Y-specific DM-domain gene required for male development in the medaka fish. Nature 2002, 417, 559–563. [Google Scholar] [CrossRef] [PubMed]
- Cui, Z.; Liu, Y.; Wang, W.; Wang, Q.; Zhang, N.; Lin, F.; Wang, N.; Shao, C.; Dong, Z.; Li, Y.; et al. Genome editing reveals dmrt1 as an essential male sex-determining gene in Chinese tongue sole (Cynoglossus semilaevis). Sci. Rep. 2017, 7, 42213. [Google Scholar] [CrossRef]
- Moore, E.C.; Ciccotto, P.J.; Peterson, E.N.; Lamm, M.S.; Albertson, R.C.; Roberts, R.B. Polygenic sex determination produces modular sex polymorphism in an African cichlid fish. Proc. Natl. Acad. Sci. USA 2022, 119, e2118574119. [Google Scholar] [CrossRef]
- Bachtrog, D.; Mank, J.E.; Peichel, C.L.; Kirkpatrick, M.; Otto, S.P.; Ashman, T.-L.; Hahn, M.W.; Kitano, J.; Mayrose, I.; Ming, R.; et al. Sex Determination: Why So Many Ways of Doing It? PLoS Biol. 2014, 12, e1001899. [Google Scholar] [CrossRef] [Green Version]
- Linnaeus. Northern Pike (Esox lucius). 1758. Available online: https://www.gbif.org/species/113226615 (accessed on 6 January 2022).
- Pan, Q.; Anderson, J.L.; Bertho, S.; Herpin, A.; Wilson, C.; Postlethwait, J.H.; Schartl, M.; Guiguen, Y. Vertebrate sex-determining genes play musical chairs. Comptes Rendus Biol. 2016, 339, 258–262. [Google Scholar] [CrossRef]
- Ogita, Y.; Mawaribuchi, S.; Nakasako, K.; Tamura, K.; Matsuda, M.; Katsumura, T.; Oota, H.; Watanabe, G.; Yoneda, S.; Takamatsu, N.; et al. Parallel Evolution of Two dmrt1-Derived Genes, dmy and dm-W, for Vertebrate Sex Determination. iScience 2020, 23, 100757. [Google Scholar] [CrossRef]
- Foster, J.W.; Graves, J.A. An SRY-related sequence on the marsupial X chromosome:implications for the evolution of the mammalian testis-determininggene. Proc. Natl. Acad. Sci. USA 1994, 91, 1927–1931. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sutton, E.; Hughes, J.; White, S.; Sekido, R.; Tan, J.; Arboleda, V.; Rogers, N.; Knower, K.; Rowley, L.; Eyre, H.; et al. Identification of SOX3 as an XX male sex reversal gene in mice and humans. J. Clin. Investig. 2011, 121, 328–341. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wong, E.S.; Schmitt, B.M.; Kazachenka, A.; Thybert, D.; Redmond, A.; Connor, F.; Rayner, T.F.; Feig, C.; Ferguson-Smith, A.C.; Marioni, J.C.; et al. Interplay of cis and trans mechanisms driving transcription factor binding and gene expression evolution. Nat. Commun. 2017, 8, 1092. [Google Scholar] [CrossRef] [PubMed]
- Tanaka, K.; Takehana, Y.; Naruse, K.; Hamaguchi, S.; Sakaizumi, M. Evidence for Different Origins of Sex Chromosomes in Closely Related Oryzias Fishes: Substitution of the Master Sex-Determining Gene. Genetics 2007, 177, 2075–2081. [Google Scholar] [CrossRef] [Green Version]
- Charlesworth, D. Evolution of recombination rates between sex chromosomes. Philos. Trans. R. Soc. B Biol. Sci. 2017, 372, 20160456. [Google Scholar] [CrossRef] [Green Version]
- Liew, W.C.; Bartfai, R.; Lim, Z.; Sreenivasan, R.; Siegfried, K.; Orban, L. Polygenic Sex Determination System in Zebrafish. PLoS ONE 2012, 7, e34397. [Google Scholar] [CrossRef] [Green Version]
- Kallman, K.D. Evidence for the Existence of Transformer Genes for Sex in the Teleost Xiphophorus maculatus. Genetics 1968, 60, 811–828. [Google Scholar] [CrossRef]
- Seehausen, O.; Van Alphen, J.; Lande, R. Color polymorphism and sex ratio distortion in a cichlid fish as an incipient stage in sympatric speciation by sexual selection. Ecol. Lett. 1999, 2, 367–378. [Google Scholar] [CrossRef]
- Böhne, A.; Weber, A.A.-T.; Rajkov, J.; Rechsteiner, M.; Riss, A.; Egger, B.; Salzburger, W. Repeated Evolution Versus Common Ancestry: Sex Chromosome Evolution in the Haplochromine Cichlid Pseudocrenilabrus philander. Genome Biol. Evol. 2019, 11, 439–458. [Google Scholar] [CrossRef] [Green Version]
- Ser, J.R.; Roberts, R.B.; Kocher, T.D. Multiple Interacting Loci Control Sex Determination in Lake Malawi Cichlid Fish. Evolution 2010, 64, 486–501. [Google Scholar] [CrossRef] [Green Version]
- Peterson, E.N.; Cline, M.E.; Moore, E.C.; Roberts, N.B.; Roberts, R.B. Genetic sex determination in Astatotilapia calliptera, a prototype species for the Lake Malawi cichlid radiation. Naturwissenschaften 2017, 104, 41. [Google Scholar] [CrossRef] [PubMed]
- Gammerdinger, W.J.; Conte, M.A.; Sandkam, B.A.; Penman, D.J.; Kocher, T.D. Characterization of sex chromosomes in three deeply diverged species of Pseudocrenilabrinae (Teleostei: Cichlidae). Hydrobiologia 2018, 832, 397–408. [Google Scholar] [CrossRef]
- Veller, C.; Muralidhar, P.; Constable, G.W.A.; Nowak, M.A. Drift-Induced Selection Between Male and Female Heterogamety. Genetics 2017, 207, 711–727. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vuilleumier, S.; Lande, R.; van Alphen, J.J.M.; Seehausen, O. Invasion and fixation of sex-reversal genes. J. Evol. Biol. 2007, 20, 913–920. [Google Scholar] [CrossRef] [Green Version]
- Myosho, T.; Otake, H.; Masuyama, H.; Matsuda, M.; Kuroki, Y.; Fujiyama, A.; Naruse, K.; Hamaguchi, S.; Sakaizumi, M. Tracing the Emergence of a Novel Sex-Determining Gene in Medaka, Oryzias luzonensis. Genetics 2012, 191, 163–170. [Google Scholar] [CrossRef] [Green Version]
- Roco, Á.S.; Olmstead, A.W.; Degitz, S.J.; Amano, T.; Zimmerman, L.B.; Bullejos, M. Coexistence of Y, W, and Z sex chromosomes in Xenopus tropicalis. Proc. Natl. Acad. Sci. USA 2015, 112, E4752–E4761. [Google Scholar] [CrossRef] [Green Version]
- Baroiller, J.-F.; D’Cotta, H. The Reversible Sex of Gonochoristic Fish: Insights and Consequences. Sex. Dev. 2016, 10, 242–266. [Google Scholar] [CrossRef]
- Gammerdinger, W.J.; Kocher, T.D. Unusual Diversity of Sex Chromosomes in African Cichlid Fishes. Genes 2018, 9, 480. [Google Scholar] [CrossRef] [Green Version]
- Ieda, R.; Hosoya, S.; Tajima, S.; Atsumi, K.; Kamiya, T.; Nozawa, A.; Aoki, Y.; Tasumi, S.; Koyama, T.; Nakamura, O.; et al. Identification of the sex-determining locus in grass puffer (Takifugu niphobles) provides evidence for sex-chromosome turnover in a subset of Takifugu species. PLoS ONE 2018, 13, e0190635. [Google Scholar] [CrossRef] [Green Version]
- Rüber, L.; Britz, R.; Zardoya, R. Molecular Phylogenetics and Evolutionary Diversification of Labyrinth Fishes (Perciformes: Anabantoidei). Syst. Biol. 2006, 55, 374–397. [Google Scholar] [CrossRef] [Green Version]
- Grazyna, F.-S.; Fopp-Bayat, D.; Jankun, M.; Krejszeff, S.; Mamcarz, A. Note on the karyotype and NOR location of Siamese fighting fish Betta splendens (Perciformes, Osphronemidae). Caryologia 2008, 614, 349–353. [Google Scholar] [CrossRef]
- Kikuchi, K.; Hamaguchi, S. Novel sex-determining genes in fish and sex chromosome evolution. Dev. Dyn. 2013, 242, 339–353. [Google Scholar] [CrossRef] [PubMed]
- Stauffer, J.R., Jr.; Bowers, N.J.; Kellogg, K.A.; McKaye, K.R. A revision of the blue-black Pseudotropheus zebra (Teleostei: Cichlidae) complex from Lake Malaŵi, Africa, with a description of a new genus and ten new species. Proc. Acad. Nat. Sci. USA 1997, 148, 189–230. [Google Scholar]
- Kottler, V.A.; Schartl, M. The Colorful Sex Chromosomes of Teleost Fish. Genes 2018, 9, 233. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Panijpan, B.; Sriwattanarothai, N.; Laosinchai, P. Wild Betta fighting fish species in Thailand and other Southeast Asian countries. ScienceAsia 2020, 46, 382–391. [Google Scholar] [CrossRef]
- Zhu, B.; Ge, W. Genome editing in fishes and their applications. Gen. Comp. Endocrinol. 2018, 257, 3–12. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| XX/XY Sex Determination Type | ZZ/ZW Sex Determination Type | ||||
|---|---|---|---|---|---|
| 0:100 Female:Male | 100:0 Female:Male | ||||
| SNP | PA | SNP | PA | ||
| POP_BSPG | Sex-specific loci | 2 | 76 | – | – |
| Overall mean distance between males and females | 1.00 ± 0.000 | 0.710 ± 0.020 | – | – | |
| Overall mean distance within females | 0.502 ± 0.042 | 0.734 ± 0.043 | – | – | |
| Overall mean distance within males | 0.00 ± 0.00 | 0.520 ± 0.042 | – | – | |
| CATT test | χ2 = 12.451 p < 0.001 | χ2 = 6.503–14.711 p < 0.001 | – | – | |
| POP_BSPGIA | Sex-specific loci | 1 | 3 | – | 3 |
| Overall mean distance between males and females | 1.00 ± 0.00 | 1.00 ± 0.00 | – | 1.00 ± 0.00 | |
| Overall mean distance within females | 0.00 ± 0.00 | 0.952 ± 0.048 | – | 0.00 ± 0.00 | |
| Overall mean distance within males | 0.00 ± 0.00 | 0.00 ± 0.00 | – | 0.30 ± 0.05 | |
| CATT test | χ2 = 10.452 p < 0.001 | χ2 = 8.012 p < 0.001 | – | χ2 = 8.400 p < 0.001 | |
| POP_BSPGL | Sex-specific loci | 1 | – | – | – |
| Overall mean distance between males and females | 1.00 ± 0.00 | – | – | – | |
| Overall mean distance within females | 0.00 ± 0.00 | – | – | – | |
| Overall mean distance within males | 0.00 ± 0.00 | – | – | – | |
| CATT test | χ2 = 11.258 p < 0.001 | – | – | – | |
| POP_BSPO | Sex-specific loci | – | – | – | – |
| Overall mean distance between males and females | – | – | – | – | |
| Overall mean distance within females | – | – | – | – | |
| Overall mean distance within males | – | – | – | – | |
| CATT test | – | – | – | – | |
| POP_BSPW | Sex-specific loci | – | 1 | – | – |
| Overall mean distance between males and females | – | 1.00 ± 0.00 | – | – | |
| Overall mean distance within females | – | 0.00 ± 0.00 | – | – | |
| Overall mean distance within males | – | 0.00 ± 0.00 | – | – | |
| CATT test | – | χ2 = 13.258 p < 0.001 | – | – | |
| Repeat Class | Fragments | Percentage (%) |
|---|---|---|
| LTR retrotransposon | 1158 | 48.70 |
| Ty3/Gypsy | 807 | 33.94 |
| Ty1/Copia | 351 | 14.76 |
| Non-LTR retrotransposon | 55 | 2.31 |
| Long interspersed nuclear elements (LINEs) | 55 | 2.31 |
| DNA transposon | 645 | 27.12 |
| Helitron | 512 | 21.53 |
| Polinton | 24 | 17.09 |
| Terminal inverted repeat (TIR) | 109 | 13.57 |
| Repeat region (unclassified) | 520 | 21.87 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Panthum, T.; Jaisamut, K.; Singchat, W.; Ahmad, S.F.; Kongkaew, L.; Wongloet, W.; Dokkaew, S.; Kraichak, E.; Muangmai, N.; Duengkae, P.; et al. Something Fishy about Siamese Fighting Fish (Betta splendens) Sex: Polygenic Sex Determination or a Newly Emerged Sex-Determining Region? Cells 2022, 11, 1764. https://doi.org/10.3390/cells11111764
Panthum T, Jaisamut K, Singchat W, Ahmad SF, Kongkaew L, Wongloet W, Dokkaew S, Kraichak E, Muangmai N, Duengkae P, et al. Something Fishy about Siamese Fighting Fish (Betta splendens) Sex: Polygenic Sex Determination or a Newly Emerged Sex-Determining Region? Cells. 2022; 11(11):1764. https://doi.org/10.3390/cells11111764
Chicago/Turabian StylePanthum, Thitipong, Kitipong Jaisamut, Worapong Singchat, Syed Farhan Ahmad, Lalida Kongkaew, Wongsathit Wongloet, Sahabhop Dokkaew, Ekaphan Kraichak, Narongrit Muangmai, Prateep Duengkae, and et al. 2022. "Something Fishy about Siamese Fighting Fish (Betta splendens) Sex: Polygenic Sex Determination or a Newly Emerged Sex-Determining Region?" Cells 11, no. 11: 1764. https://doi.org/10.3390/cells11111764