
Icariin Treatment Rescues Diabetes Induced Bone Loss via Scavenging ROS and Activating Primary Cilia/Gli2/Osteocalcin Signaling Pathway

 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Reagents and Antibodies
2.2. T1DM SD Rats Model Construction
2.3. Diabetic Cell Model Constructionin Vitro
2.3.1. Primary Osteoblasts Isolation, Culture and Osteoblast Differentiation Induction
2.3.2. Cell Model Construction
2.3.3. Alizarin RED Staining Mineralized Nodules
2.3.4. Intracellular ROS Determination
2.3.5. ALP Activity Analysis
2.4. Immunofluorescence
2.4.1. Primary Cilia Immunofluorescence
2.4.2. Mitochondrial Mito-Tracker Green Immunofluorescence
2.5. Plasmids and Transfection
2.6. Immunohistochemistry
2.7. Micro-Computed Tomography (Micro-CT)
2.8. Western Blot
2.9. Transmission Electron Microscopy (TEM)
2.10. Statistical Analysis
3. Results
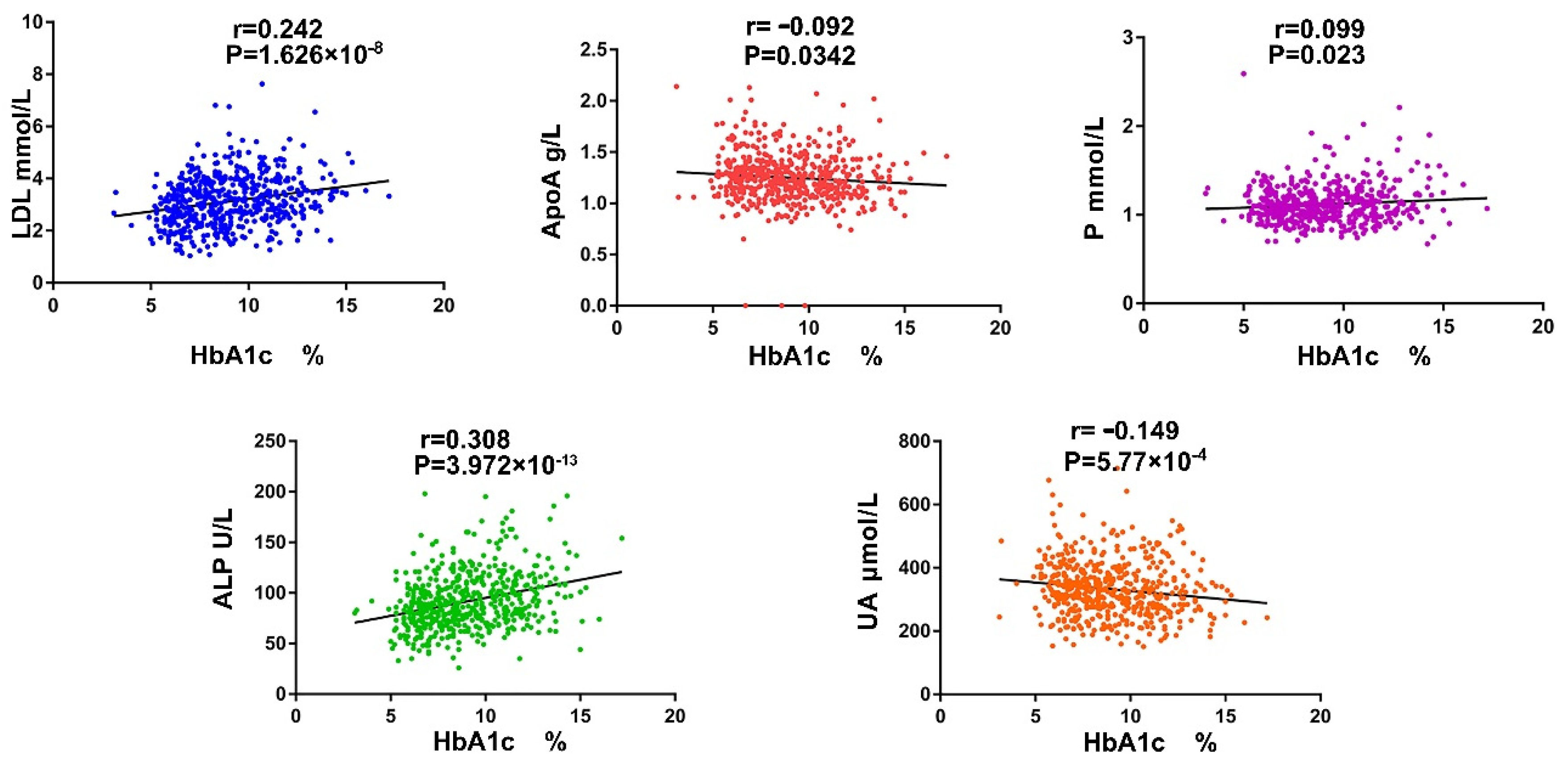
3.1. The Serum ALP and P Ion Are Important Indicators Reflecting Diabetic Bone Metabolism Disorders
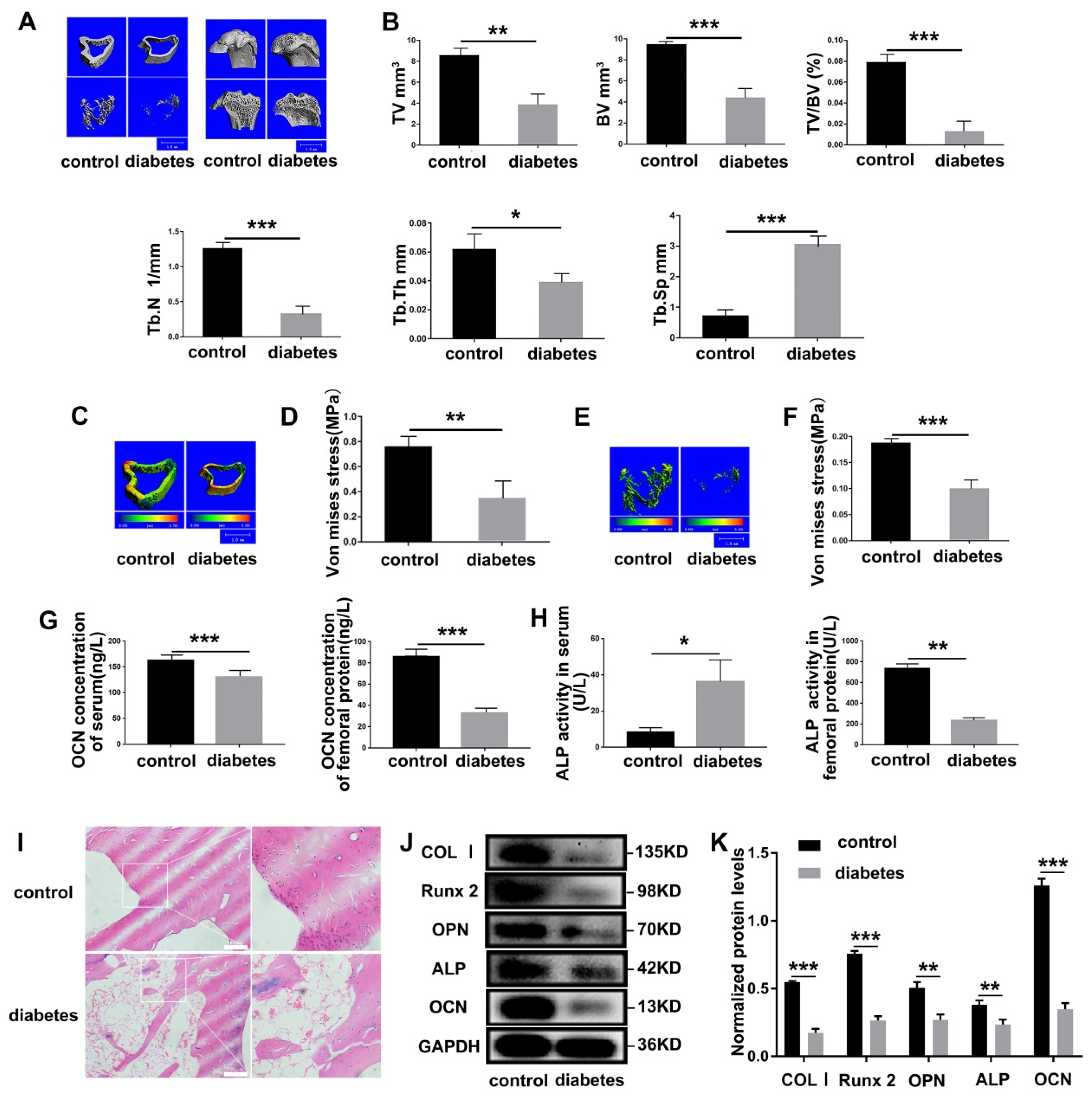
3.2. Diabetes Results in Severe Impairment of Bone Mechanical Properties in SD Rats
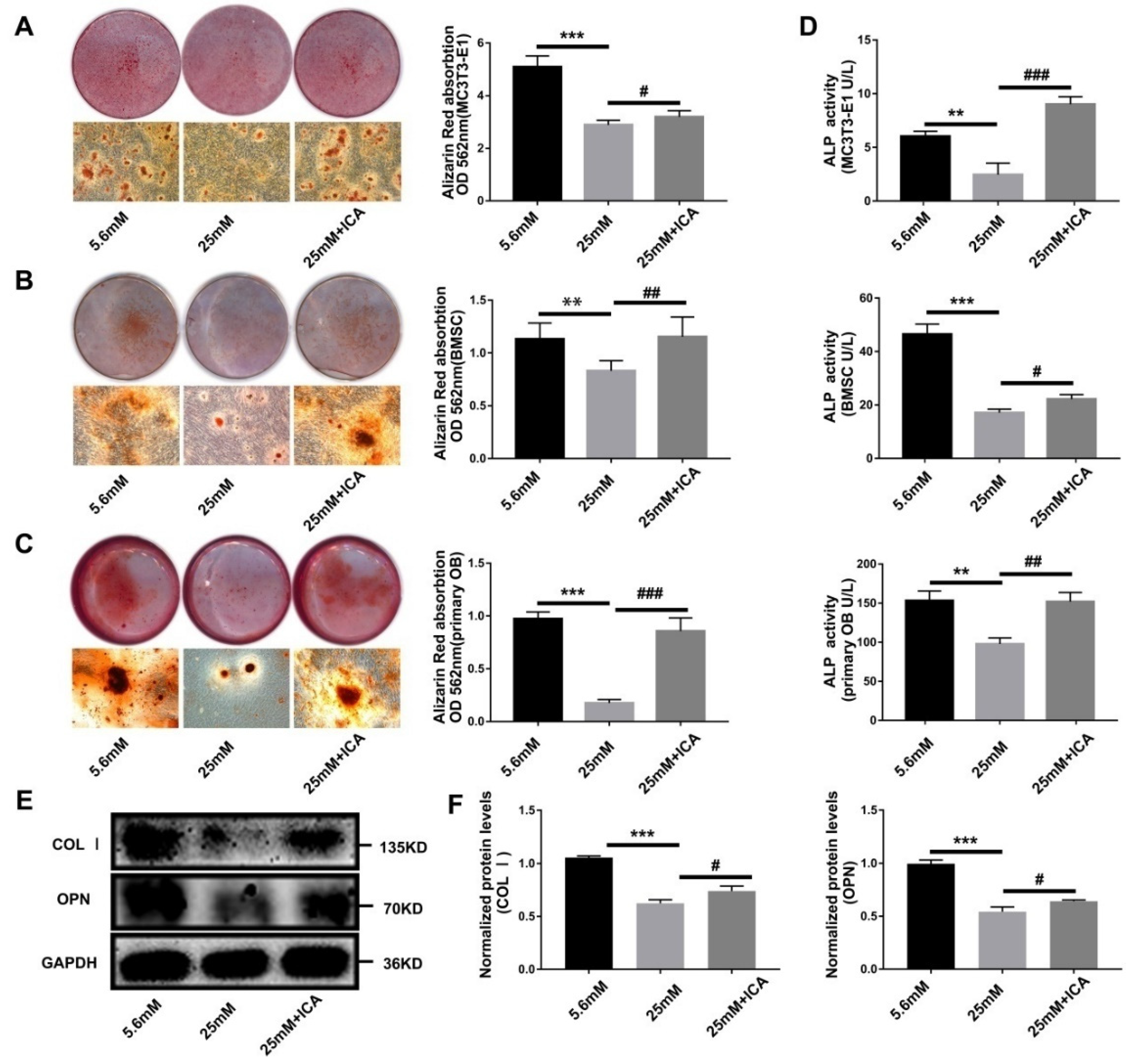
3.3. ICA Rescue Osteoblast Differentiation Inhibited by High Glucose
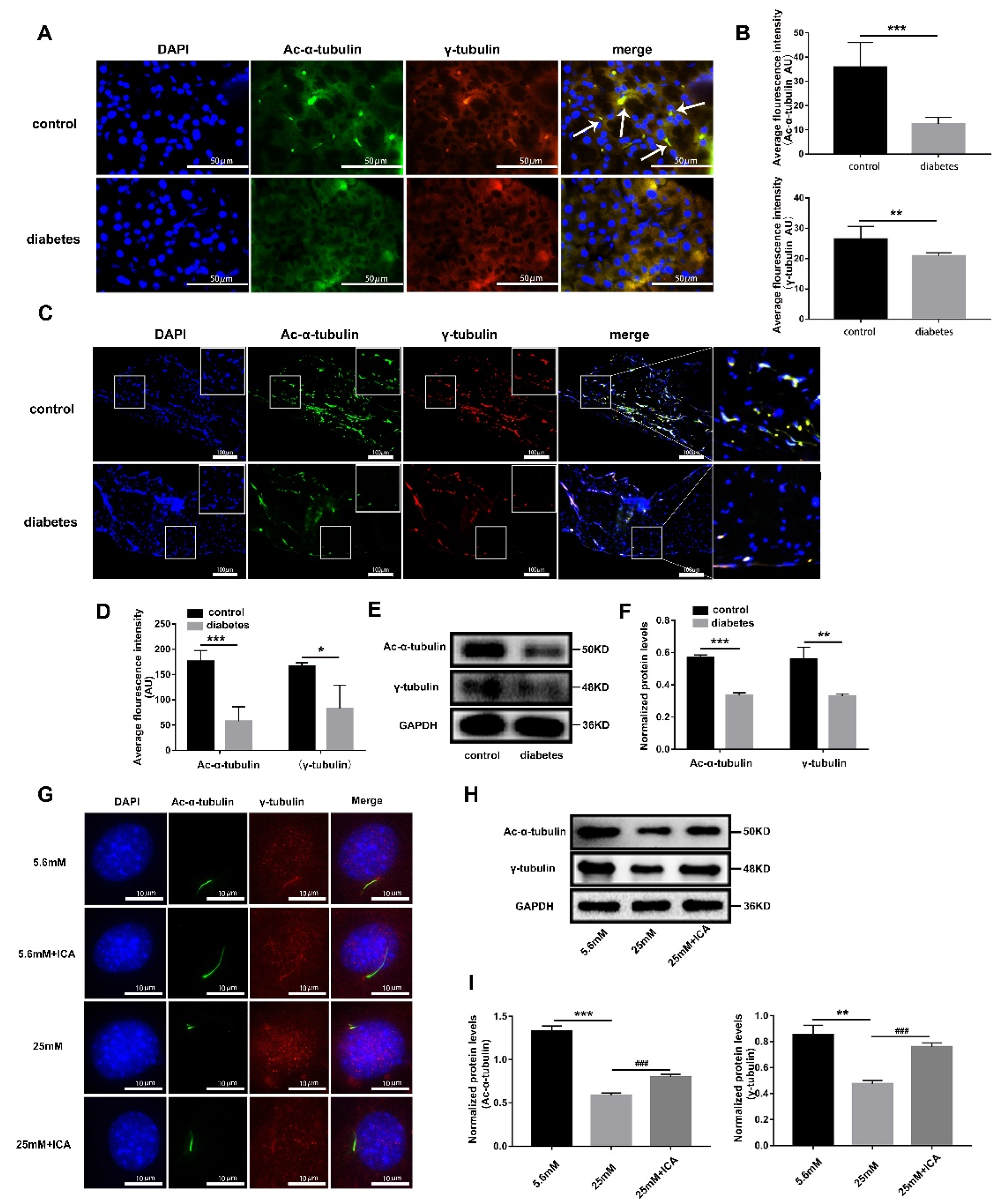
3.4. ICA Can Protect Osteoblast Primary Cilia from High Glucose Damage
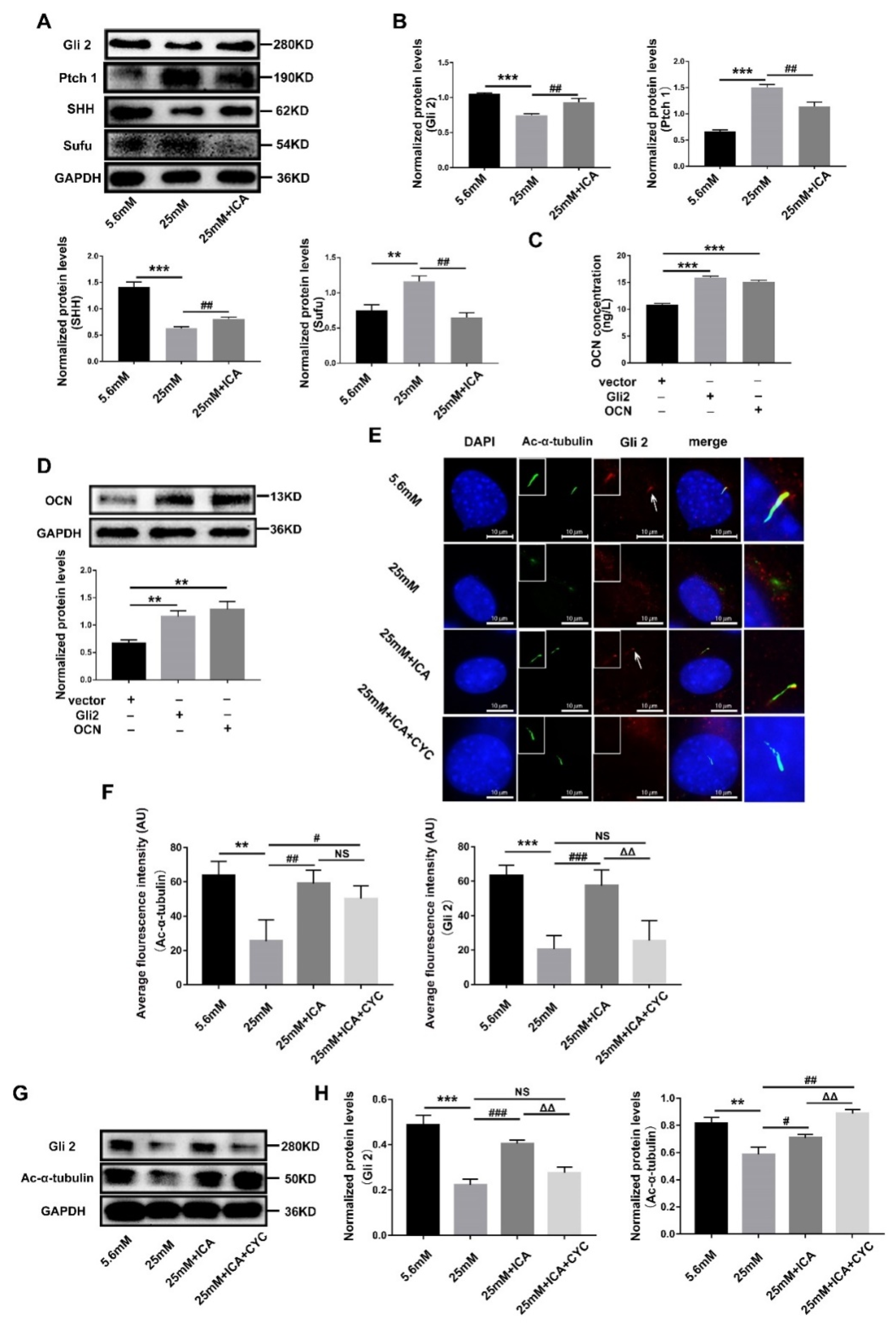
3.5. ICA Activates Hedgehog Signaling to Promote Osteoblast Differentiation by Maintaining Gli2’s Ciliary Localization
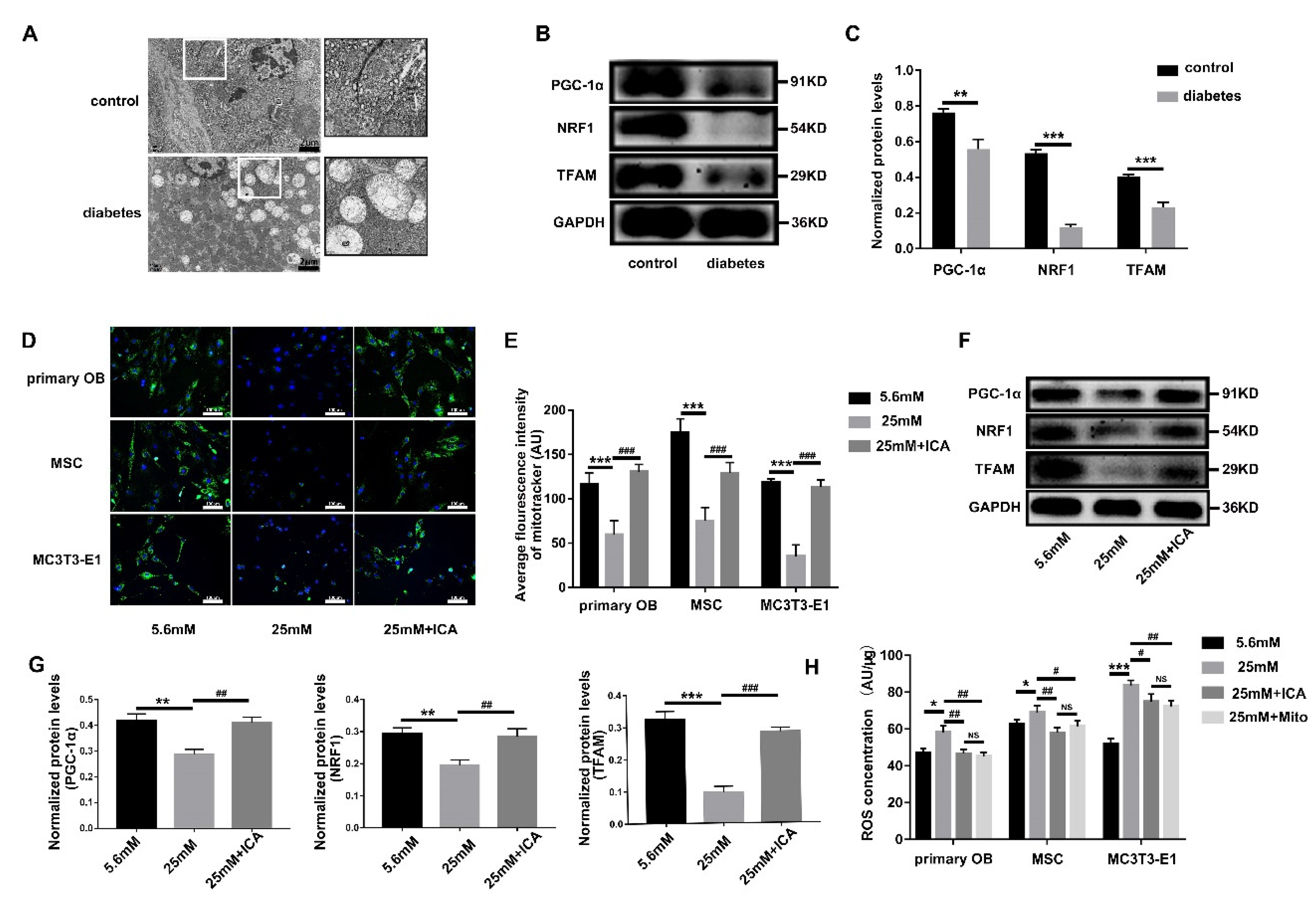
3.6. ICA Alleviates the ROS Threat to Mitochondria and Primary Cilia
4. Discussion
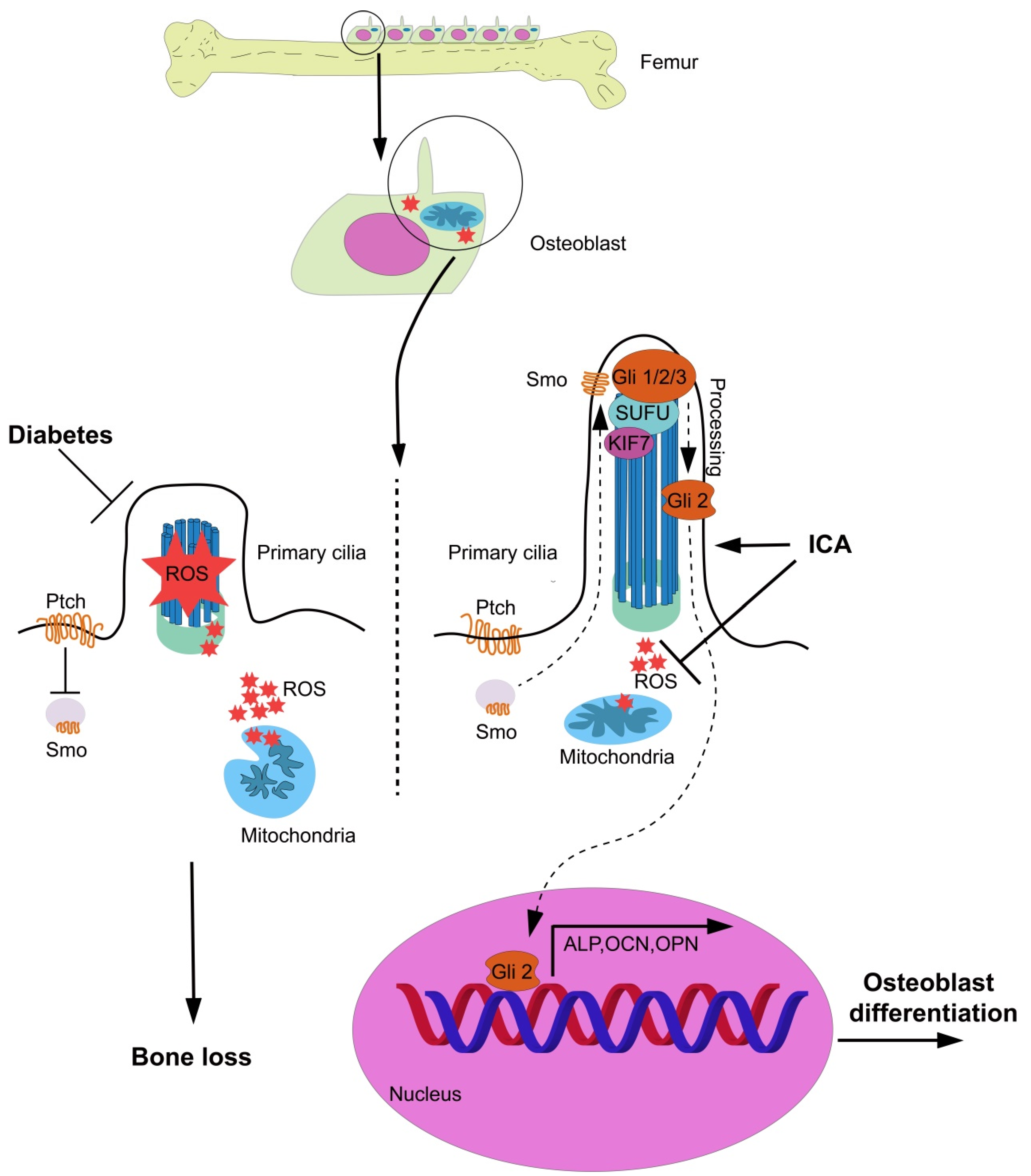
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Sun, H.; Saeedi, P.; Karuranga, S.; Pinkepank, M.; Ogurtsova, K.; Duncan, B.B.; Stein, C.; Basit, A.; Chan JC, N.; Mbanya, J.C.; et al. IDF Diabetes Atlas: Global, regional and country-level diabetes prevalence estimates for 2021 and projections for 2045. Diabetes Res. Clin. Pract. 2022, 183, 109119. [Google Scholar] [CrossRef] [PubMed]
- Q&A: Key points for IDF Diabetes Atlas 2017. Diabetes Res. Clin. Pract. 2018, 135, 235–236.
- KDIGO 2020 Clinical Practice Guideline for Diabetes Management in Chronic Kidney Disease. Kidney Int. 2020, 98, S1–S115. [CrossRef] [PubMed]
- Klein, K.R.; Buse, J.B. The trials and tribulations of determining HbA(1c) targets for diabetes mellitus. Nat. Rev. Endocrinol. 2020, 16, 717–730. [Google Scholar] [CrossRef] [PubMed]
- Nielsen, J.J.; Low, S.A.; Ramseier, N.T.; Hadap, R.V.; Young, N.A.; Wang, M.; Low, P.S. Analysis of the bone fracture targeting properties of osteotropic ligands. J. Control. Release 2021, 329, 570–584. [Google Scholar] [CrossRef]
- Hofbauer, L.C.; Busse, B.; Eastell, R.; Ferrari, S.; Frost, M.; Müller, R.; Burden, A.M.; Rivadeneira, F.; Napoli, N.; Rauner, M. Bone fragility in diabetes: Novel concepts and clinical implications. Lancet Diabetes Endocrinol. 2022, 10, 207–220. [Google Scholar] [CrossRef]
- Sewing, L.; Potasso, L.; Baumann, S.; Schenk, D.; Gazozcu, F.; Lippuner, K.; Kraenzlin, M.; Zysset, P.; Meier, C. Bone Microarchitecture and Strength in Long-Standing Type 1 Diabetes. J. Bone Min. Res. 2022, 37, 837–847. [Google Scholar] [CrossRef]
- Napoli, N.; Schwartz, A.V.; Schafer, A.L.; Vittinghoff, E.; Cawthon, P.M.; Parimi, N.; Orwoll, E.; Strotmeyer, E.S.; Hoffman, A.R.; Barrett-Connor, E.; et al. Vertebral Fracture Risk in Diabetic Elderly Men: The MrOS Study. J. Bone Min. Res. 2018, 33, 63–69. [Google Scholar] [CrossRef] [Green Version]
- Sheu, A.; Greenfield, J.R.; White, C.P.; Center, J.R. Contributors to impaired bone health in type 2 diabetes. Trends Endocrinol. Metab. 2022, 23, S1043-2760(22)00201-6. [Google Scholar] [CrossRef]
- Cheng, K.; Guo, Q.; Yang, W.; Wang, Y.; Sun, Z.; Wu, H. Mapping Knowledge Landscapes and Emerging Trends of the Links Between Bone Metabolism and Diabetes Mellitus: A Bibliometric Analysis From 2000 to 2021. Front. Public Health 2022, 10, 918483. [Google Scholar] [CrossRef]
- Shahen, V.A.; Gerbaix, M.; Koeppenkastrop, S.; Lim, S.F.; Mcfarlane, K.E.; Nguyen AN, L.; Peng, X.Y.; Weiss, N.B.; Brennan-Speranza, T.C. Multifactorial effects of hyperglycaemia, hyperinsulinemia and inflammation on bone remodelling in type 2 diabetes mellitus. Cytokine Growth Factor Rev. 2020, 55, 109–118. [Google Scholar] [CrossRef]
- Napoli, N.; Chandran, M.; Pierroz, D.D.; Abrahamsen, B.; Schwartz, A.V.; Ferrari, S.L. Mechanisms of diabetes mellitus-induced bone fragility. Nat. Rev. Endocrinol. 2017, 13, 208–219. [Google Scholar] [CrossRef]
- Jiang, Y.; Zhang, Y.; Chen, W.; Liu, C.; Li, X.; Sun, D.; Liu, Z.; Xu, Y.; Mao, X.; Guo, Q.; et al. Correction to: Achyranthes bidentata extract exerts osteoprotective effects on steroid-induced osteonecrosis of the femoral head in rats by regulating RANKL/RANK/OPG signaling. J. Transl. Med. 2021, 19, 208. [Google Scholar] [CrossRef]
- Ahmad, W.; Ijaz, B.; Shabbiri, K.; Ahmed, F.; Rehman, S. Oxidative toxicity in diabetes and Alzheimer’s disease: Mechanisms behind ROS/ RNS generation. J. Biomed. Sci. 2017, 24, 76. [Google Scholar] [CrossRef]
- Dugan, L.L.; You, Y.H.; Ali, S.S.; Diamond-Stanic, M.; Miyamoto, S.; Decleves, A.E.; Andreyev, A.; Quach, T.; Ly, S.; Shekhtman, G.; et al. AMPK dysregulation promotes diabetes-related reduction of superoxide and mitochondrial function. J. Clin. Investig. 2013, 123, 4888–4899. [Google Scholar] [CrossRef] [Green Version]
- Li, D.; Chen, K.; Tang, H.; Hu, S.; Xin, L.; Jing, X.; He, Q.; Wang, S.; Song, J.; Mei, L.; et al. A Logic-Based Diagnostic and Therapeutic Hydrogel with Multistimuli Responsiveness to Orchestrate Diabetic Bone Regeneration. Adv. Mater. 2022, 34, e2108430. [Google Scholar] [CrossRef]
- Tao, Z.S.; Wang, H.S.; Li, T.L.; Wei, S. Silibinin-modified Hydroxyapatite coating promotes the osseointegration of titanium rods by activation SIRT1/SOD2 signaling pathway in diabetic rats. J. Mater. Sci. Mater Med. 2022, 33, 62. [Google Scholar] [CrossRef] [PubMed]
- Ma, H.; Wang, X.; Zhang, W.; Li, H.; Zhao, W.; Sun, J.; Yang, M. Melatonin Suppresses Ferroptosis Induced by High Glucose via Activation of the Nrf2/HO-1 Signaling Pathway in Type 2 Diabetic Osteoporosis. Oxid. Med. Cell. Longev. 2020, 2020, 9067610. [Google Scholar] [CrossRef] [PubMed]
- Tu, C.; Lu, H.; Zhou, T.; Zhang, W.; Deng, L.; Cao, W.; Yang, Z.; Wang, Z.; Wu, X.; Ding, J.; et al. Promoting the healing of infected diabetic wound by an anti-bacterial and nano-enzyme-containing hydrogel with inflammation-suppressing, ROS-scavenging, oxygen and nitric oxide-generating properties. Biomaterials 2022, 286, 121597. [Google Scholar] [CrossRef] [PubMed]
- Khosla, S.; Hofbauer, L.C. Osteoporosis treatment: Recent developments and ongoing challenges. Lancet Diabetes Endocrinol. 2017, 5, 898–907. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ebeling, P.R.; Nguyen, H.H.; Aleksova, J.; Vincent, A.J.; Wong, P.; Milat, F. Secondary Osteoporosis. Endocr. Rev. 2022, 43, 240–313. [Google Scholar] [CrossRef]
- Yong, E.L.; Cheong, W.F.; Huang, Z.; Thu WP, P.; Cazenave-Gassiot, A.; Seng, K.Y.; Logan, S. Randomized, double-blind, placebo-controlled trial to examine the safety, pharmacokinetics and effects of Epimedium prenylflavonoids, on bone specific alkaline phosphatase and the osteoclast adaptor protein TRAF6 in post-menopausal women. Phytomedicine 2021, 91, 153680. [Google Scholar] [CrossRef]
- Wisanwattana, W.; Wongkrajang, K.; Cao, D.Y.; Shi, X.K.; Zhang, Z.H.; Zhou, Z.Y.; Li, F.; Mei, Q.G.; Wang, C.; Suksamrarn, A.; et al. Inhibition of Phosphodiesterase 5 Promotes the Aromatase-Mediated Estrogen Biosynthesis in Osteoblastic Cells by Activation of cGMP/PKG/SHP2 Pathway. Front. Endocrinol. 2021, 12, 636784. [Google Scholar] [CrossRef]
- Wu, Y.; Cao, L.; Xia, L.; Wu, Q.; Wang, J.; Wang, X.; Xu, L.; Zhou, Y.; Xu, Y.; Jiang, X. Evaluation of Osteogenesis and Angiogenesis of Icariin in Local Controlled Release and Systemic Delivery for Calvarial Defect in Ovariectomized Rats. Sci. Rep. 2017, 7, 5077. [Google Scholar] [CrossRef] [Green Version]
- Zhou, L.; Poon, C.C.; Wong, K.Y.; Cao, S.; Dong, X.; Zhang, Y.; Wong, M.S. Icariin ameliorates estrogen-deficiency induced bone loss by enhancing IGF-I signaling via its crosstalk with non-genomic ERα signaling. Phytomedicine 2021, 82, 153413. [Google Scholar] [CrossRef]
- Kim, B.; Lee, K.Y.; Park, B. Icariin abrogates osteoclast formation through the regulation of the RANKL-mediated TRAF6/NF-κB/ERK signaling pathway in Raw264.7 cells. Phytomedicine 2018, 51, 181–190. [Google Scholar] [CrossRef]
- Yang, A.; Yu, C.; Lu, Q.; Li, H.; Li, Z.; He, C. Mechanism of Action of Icariin in Bone Marrow Mesenchymal Stem Cells. Stem Cells Int. 2019, 2019, 5747298. [Google Scholar] [CrossRef] [Green Version]
- Xia, J.; Hu, J.N.; Zhang, R.B.; Liu, W.; Zhang, H.; Wang, Z.; Jiang, S.; Wang, Y.P.; Li, W. Icariin exhibits protective effects on cisplatin-induced cardiotoxicity via ROS-mediated oxidative stress injury in vivo and in vitro. Phytomedicine 2022, 104, 154331. [Google Scholar] [CrossRef]
- Zhao, H.; Zhao, T.; Yang, J.; Huang, Q.; Wu, H.; Pan, Y.; Wang, H.; Qian, Y. Epimedium protects against dyszoospermia in mice with Pex3 knockout by exerting antioxidant effects and regulating the expression level of P16. Cell Death Dis. 2022, 13, 69. [Google Scholar] [CrossRef]
- Ding, X.; Zhao, H.; Qiao, C. Icariin protects podocytes from NLRP3 activation by Sesn2-induced mitophagy through the Keap1-Nrf2/HO-1 axis in diabetic nephropathy. Phytomedicine 2022, 99, 154005. [Google Scholar] [CrossRef]
- Shi, W.; Gao, Y.; Wang, Y.; Zhou, J.; Wei, Z.; Ma, X.; Ma, H.; Xian, C.J.; Wang, J.; Chen, K. The flavonol glycoside icariin promotes bone formation in growing rats by activating the cAMP signaling pathway in primary cilia of osteoblasts. J. Biol. Chem. 2017, 292, 20883–20896. [Google Scholar] [CrossRef] [PubMed]
- Yuan, X.; Serra, R.A.; Yang, S. Function and regulation of primary cilia and intraflagellar transport proteins in the skeleton. Ann. N. Y. Acad. Sci. 2015, 1335, 78–99. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kluth, O.; Stadion, M.; Gottmann, P.; Aga, H.; Jähnert, M.; Scherneck, S.; Vogel, H.; Krus, U.; Seelig, A.; Ling, C.; et al. Decreased Expression of Cilia Genes in Pancreatic Islets as a Risk Factor for Type 2 Diabetes in Mice and Humans. Cell. Rep. 2019, 26, 3027–3036.e3023. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, S.Y.; Long, F. Notch signaling suppresses glucose metabolism in mesenchymal progenitors to restrict osteoblast differentiation. J. Clin. Investig. 2018, 128, 5573–5586. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ehnert, S.; Sreekumar, V.; Aspera-Werz, R.H.; Sajadian, S.O.; Wintermeyer, E.; Sandmann, G.H.; Bahrs, C.; Hengstler, J.G.; Godoy, P.; Nussler, A.K. TGF-beta1 impairs mechanosensation of human osteoblasts via HDAC6-mediated shortening and distortion of primary cilia. J. Mol. Med. 2017, 95, 653–663. [Google Scholar] [CrossRef]
- Esteves-Monteiro, M.; Menezes-Pinto, D.; Ferreira-Duarte, M.; Dias-Pereira, P.; Morato, M.; Duarte-Araújo, M. Histomorphometry Changes and Decreased Reactivity to Angiotensin II in the Ileum and Colon of Streptozotocin-Induced Diabetic Rats. Int. J. Mol. Sci. 2022, 23, 13233. [Google Scholar] [CrossRef]
- Cheng, R.X.; Feng, Y.; Liu, D.; Wang, Z.H.; Zhang, J.T.; Chen, L.H.; Su, C.J.; Wang, B.; Huang, Y.; Ji, R.R.; et al. The role of Nav1.7 and methylglyoxal-mediated activation of TRPA1 in itch and hypoalgesia in a murine model of type 1 diabetes. Theranostics 2019, 9, 4287–4307. [Google Scholar] [CrossRef]
- Xu, J.; Deng, X.; Wu, X.; Zhu, H.; Zhu, Y.; Liu, J.; Chen, Q.; Yuan, C.; Liu, G.; Wang, C. Primary cilia regulate gastric cancer-induced bone loss via cilia/Wnt/β-catenin signaling pathway. Aging 2021, 13, 8989–9010. [Google Scholar] [CrossRef]
- Xu, D.; Liu, J.; Zheng, W.; Gao, Q.; Gao, Y.; Leng, X. Identification of Polysaccharides From Dipsacus asperoides and Their Effects on Osteoblast Proliferation and Differentiation in a High-Glucose Environment. Front. Pharmacol. 2022, 13, 851956. [Google Scholar] [CrossRef]
- De Waard, E.A.C.; De Jong, J.J.A.; Koster, A.; Savelberg, H.; Van Geel, T.A.; Houben, A.; Schram, M.T.; Dagnelie, P.C.; Van Der Kallen, C.J.; Sep, S.J.S.; et al. The association between diabetes status, HbA1c, diabetes duration, microvascular disease, and bone quality of the distal radius and tibia as measured with high-resolution peripheral quantitative computed tomography-The Maastricht Study. Osteoporos. Int. 2018, 29, 2725–2738. [Google Scholar] [CrossRef] [Green Version]
- Morgan, E.F.; Unnikrisnan, G.U.; Hussein, A.I. Bone Mechanical Properties in Healthy and Diseased States. Ann. Rev. Biomed. Eng. 2018, 20, 119–143. [Google Scholar] [CrossRef]
- Zhang, J.; Zhang, W.; Dai, J.; Wang, X.; Shen, S.G. Overexpression of Dlx2 enhances osteogenic differentiation of BMSCs and MC3T3-E1 cells via direct upregulation of Osteocalcin and Alp. Int. J. Oral. Sci. 2019, 11, 12. [Google Scholar] [CrossRef] [Green Version]
- Anvarian, Z.; Mykytyn, K.; Mukhopadhyay, S.; Pedersen, L.B.; Christensen, S.T. Cellular signalling by primary cilia in development, organ function and disease. Nat. Rev. Nephrol. 2019, 15, 199–219. [Google Scholar] [CrossRef]
- Moore, E.R.; Zhu, Y.X.; Ryu, H.S.; Jacobs, C.R. Periosteal progenitors contribute to load-induced bone formation in adult mice and require primary cilia to sense mechanical stimulation. Stem. Cell Res. 2018, 9, 190. [Google Scholar]
- Bangs, F.; Anderson, K.V. Primary Cilia and Mammalian Hedgehog Signaling. Cold Spring Harb. Perspect. Biol. 2017, 9, a028175. [Google Scholar] [CrossRef] [Green Version]
- Tsushima, K.; Bugger, H.; Wende, A.R.; Soto, J.; Jenson, G.A.; Tor, A.R.; Mcglauflin, R.; Kenny, H.C.; Zhang, Y.; Souvenir, R.; et al. Mitochondrial Reactive Oxygen Species in Lipotoxic Hearts Induce Post-Translational Modifications of AKAP121, DRP1, and OPA1 That Promote Mitochondrial Fission. Circ. Res. 2018, 122, 58–73. [Google Scholar] [CrossRef]
- Ruegsegger, G.N.; Creo, A.L.; Cortes, T.M.; Dasari, S.; Nair, K.S. Altered mitochondrial function in insulin-deficient and insulin-resistant states. J. Clin. Investig. 2018, 128, 3671–3681. [Google Scholar] [CrossRef] [Green Version]
- Moruzzi, N.; Valladolid-Acebes, I.; Kannabiran, S.A.; Bulgaro, S.; Burtscher, I.; Leibiger, B.; Leibiger, I.B.; Berggren, P.O.; Brismar, K. Mitochondrial impairment and intracellular reactive oxygen species alter primary cilia morphology. Life Sci. Alliance 2022, 5, e202201505. [Google Scholar] [CrossRef]
- Volta, F.; Scerbo, M.J.; Seelig, A.; Wagner, R.; O’brien, N.; Gerst, F.; Fritsche, A.; Häring, H.U.; Zeigerer, A.; Ullrich, S.; et al. Glucose homeostasis is regulated by pancreatic β-cell cilia via endosomal EphA-processing. Nat. Commun. 2019, 10, 5686. [Google Scholar] [CrossRef] [Green Version]
- Cho, J.H.; Li, Z.A.; Zhu, L.; Muegge, B.D.; Roseman, H.F.; Lee, E.Y.; Utterback, T.; Woodhams, L.G.; Bayly, P.V.; Hughes, J.W. Islet primary cilia motility controls insulin secretion. Sci. Adv. 2022, 8, eabq8486. [Google Scholar] [CrossRef]
- Chinipardaz, Z.; Liu, M.; Graves, D.T.; Yang, S. Role of Primary Cilia in Bone and Cartilage. J. Dent. Res. 2022, 101, 253–260. [Google Scholar] [CrossRef] [PubMed]
- Kawata, K.; Narita, K.; Washio, A.; Kitamura, C.; Nishihara, T.; Kubota, S.; Takeda, S. Odontoblast differentiation is regulated by an interplay between primary cilia and the canonical Wnt pathway. Bone 2021, 150, 116001. [Google Scholar] [CrossRef] [PubMed]
- Chinipardaz, Z.; Liu, M.; Graves, D.; Yang, S. Diabetes impairs fracture healing through disruption of cilia formation in osteoblasts. Bone 2021, 153, 116176. [Google Scholar] [CrossRef] [PubMed]
- Zhang, C.; Feinberg, D.; Alharbi, M.; Ding, Z.; Lu, C.; O’connor, J.P.; Graves, D.T. Chondrocytes Promote Vascularization in Fracture Healing Through a FOXO1-Dependent Mechanism. J. Bone Min. Res. 2019, 34, 547–556. [Google Scholar] [CrossRef] [PubMed]
- Lu, Y.; Alharbi, M.; Zhang, C.; O’connor, J.P.; Graves, D.T. Deletion of FOXO1 in chondrocytes rescues the effect of diabetes on mechanical strength in fracture healing. Bone 2019, 123, 159–167. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Graves, D.T. Keratinocyte Function in Normal and Diabetic Wounds and Modulation by FOXO1. J. Diabetes Res. 2020, 2020, 3714704. [Google Scholar] [CrossRef]
- Sun, J.; Shin, D.Y.; Eiseman, M.; Yallowitz, A.R.; Li, N.; Lalani, S.; Li, Z.; Cung, M.; Bok, S.; Debnath, S.; et al. SLITRK5 is a negative regulator of hedgehog signaling in osteoblasts. Nat. Commun. 2021, 12, 4611. [Google Scholar] [CrossRef]
- Wu, J.; Wang, R.; Kan, X.; Zhang, J.; Sun, W.; Goltzman, D.; Miao, D. A Sonic Hedgehog-Gli-Bmi1 signaling pathway plays a critical role in p27 deficiency induced bone anabolism. Int. J. Biol. Sci. 2022, 18, 956–969. [Google Scholar] [CrossRef]
- Santos, N.; Reiter, J.F. A central region of Gli2 regulates its localization to the primary cilium and transcriptional activity. J. Cell Sci. 2014, 127, 1500–1510. [Google Scholar] [CrossRef] [Green Version]
- Ohba, S. Hedgehog Signaling in Skeletal Development: Roles of Indian Hedgehog and the Mode of Its Action. Int. J. Mol. Sci. 2020, 21, 6665. [Google Scholar] [CrossRef]
- Goodpaster, B.H.; Sparks, L.M. Metabolic Flexibility in Health and Disease. Cell Metab. 2017, 25, 1027–1036. [Google Scholar] [CrossRef] [Green Version]
- Ji, Y.; Chae, S.; Lee, H.K.; Park, I.; Kim, C.; Ismail, T.; Kim, Y.; Park, J.W.; Kwon, O.S.; Kang, B.S.; et al. Peroxiredoxin5 Controls Vertebrate Ciliogenesis by Modulating Mitochondrial Reactive Oxygen Species. Antioxid. Redox Signal. 2019, 30, 1731–1745. [Google Scholar] [CrossRef]
- Eftekharpour, E.; Fernyhough, P. Oxidative Stress and Mitochondrial Dysfunction Associated with Peripheral Neuropathy in Type 1 Diabetes. Antioxid. Redox Signal. 2022, 37, 578–596. [Google Scholar] [CrossRef]
- Bhatti, J.S.; Sehrawat, A.; Mishra, J.; Sidhu, I.S.; Navik, U.; Khullar, N.; Kumar, S.; Bhatti, G.K.; Reddy, P.H. Oxidative stress in the pathophysiology of type 2 diabetes and related complications: Current therapeutics strategies and future perspectives. Free Radic. Biol. Med. 2022, 184, 114–134. [Google Scholar] [CrossRef]
- An, Y.; Zhang, H.; Wang, C.; Jiao, F.; Xu, H.; Wang, X.; Luan, W.; Ma, F.; Ni, L.; Tang, X.; et al. Activation of ROS/MAPKs/NF-κB/NLRP3 and inhibition of efferocytosis in osteoclast-mediated diabetic osteoporosis. Faseb J. 2019, 33, 12515–12527. [Google Scholar] [CrossRef] [Green Version]
- Liu, Y.; Palanivel, R.; Rai, E.; Park, M.; Gabor, T.V.; Scheid, M.P.; Xu, A.; Sweeney, G. Adiponectin stimulates autophagy and reduces oxidative stress to enhance insulin sensitivity during high-fat diet feeding in mice. Diabetes 2015, 64, 36–48. [Google Scholar] [CrossRef] [Green Version]
- Hughes, J.W.; Cho, J.H.; Conway, H.E.; Digruccio, M.R.; Ng, X.W.; Roseman, H.F.; Abreu, D.; Urano, F.; Piston, D.W. Primary cilia control glucose homeostasis via islet paracrine interactions. Proc. Natl. Acad. Sci. USA 2020, 117, 8912–8923. [Google Scholar] [CrossRef] [Green Version]
- Kong, M.J.; Bak, S.H.; Han, K.H.; Kim, J.I.; Park, J.W.; Park, K.M. Fragmentation of kidney epithelial cell primary cilia occurs by cisplatin and these cilia fragments are excreted into the urine. Redox Biol. 2019, 20, 38–45. [Google Scholar] [CrossRef]
- Panchapakesan, U.; Pollock, C. The primary cilia in diabetic kidney disease: A tubulocentric view? Int. J. Biochem. Cell Biol. 2020, 122, 105718. [Google Scholar] [CrossRef]
- Zhu, Y.; Ye, L.; Cai, X.; Li, Z.; Fan, Y.; Yang, F. Icariin-Loaded Hydrogel Regulates Bone Marrow Mesenchymal Stem Cell Chondrogenic Differentiation and Promotes Cartilage Repair in Osteoarthritis. Front. Bioeng. Biotechnol. 2022, 10, 755260. [Google Scholar] [CrossRef]
- Ho-Shui-Ling, A.; Bolander, J.; Rustom, L.E.; Johnson, A.W.; Luyten, F.P.; Picart, C. Bone regeneration strategies: Engineered scaffolds, bioactive molecules and stem cells current stage and future perspectives. Biomaterials 2018, 180, 143–162. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Chang, X.; Ma, Q.; Sun, B.; Li, H.; Zhou, J.; Hu, Y.; Yang, X.; Li, J.; Chen, X.; et al. Bioinspired drug-delivery system emulating the natural bone healing cascade for diabetic periodontal bone regeneration. Bioact. Mater. 2022, 21, 324–339. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Model | Unstandardized Coefficients | Standardized Coefficients | t | Significance | Collinearity Statistics | ||
|---|---|---|---|---|---|---|---|
| B | Std. Error | Beta | Tolerance | VIF | |||
| (Constant) | 6.849 | 0.882 | 7.761 | 0.000 | |||
| LDL | 0.635 | 0.100 | 0.253 | 6.369 | 0.000 | 0.971 | 1.030 |
| ApoA | −1.421 | 0.396 | −0.142 | −3.584 | 0.000 | 0.968 | 1.033 |
| P | 1.315 | 0.445 | 0.116 | 2.953 | 0.003 | 0.985 | 1.015 |
| ALP | 0.025 | 0.003 | 0.284 | 7.249 | 0.000 | 0.992 | 1.008 |
| UA | −0.005 | 0.001 | −0.191 | −4.791 | 0.000 | 0.959 | 1.043 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liu, J.; Cheng, Q.; Wu, X.; Zhu, H.; Deng, X.; Wang, M.; Yang, S.; Xu, J.; Chen, Q.; Li, M.; et al. Icariin Treatment Rescues Diabetes Induced Bone Loss via Scavenging ROS and Activating Primary Cilia/Gli2/Osteocalcin Signaling Pathway. Cells 2022, 11, 4091. https://doi.org/10.3390/cells11244091
Liu J, Cheng Q, Wu X, Zhu H, Deng X, Wang M, Yang S, Xu J, Chen Q, Li M, et al. Icariin Treatment Rescues Diabetes Induced Bone Loss via Scavenging ROS and Activating Primary Cilia/Gli2/Osteocalcin Signaling Pathway. Cells. 2022; 11(24):4091. https://doi.org/10.3390/cells11244091
Chicago/Turabian StyleLiu, Jie, Qingfeng Cheng, Xiangmei Wu, Huifang Zhu, Xiaoyan Deng, Maorong Wang, Shengyong Yang, Jie Xu, Qian Chen, Mengxue Li, and et al. 2022. "Icariin Treatment Rescues Diabetes Induced Bone Loss via Scavenging ROS and Activating Primary Cilia/Gli2/Osteocalcin Signaling Pathway" Cells 11, no. 24: 4091. https://doi.org/10.3390/cells11244091