
Imaging Cataract-Specific Peptides in Human Lenses

Abstract
:1. Introduction
2. Materials and Methods
2.1. Tissue Preparation for Imaging Mass Spectrometry (IMS)
2.2. MALDI Data Acquisition
2.3. Peptide Identification and LC-MS/MS Analysis
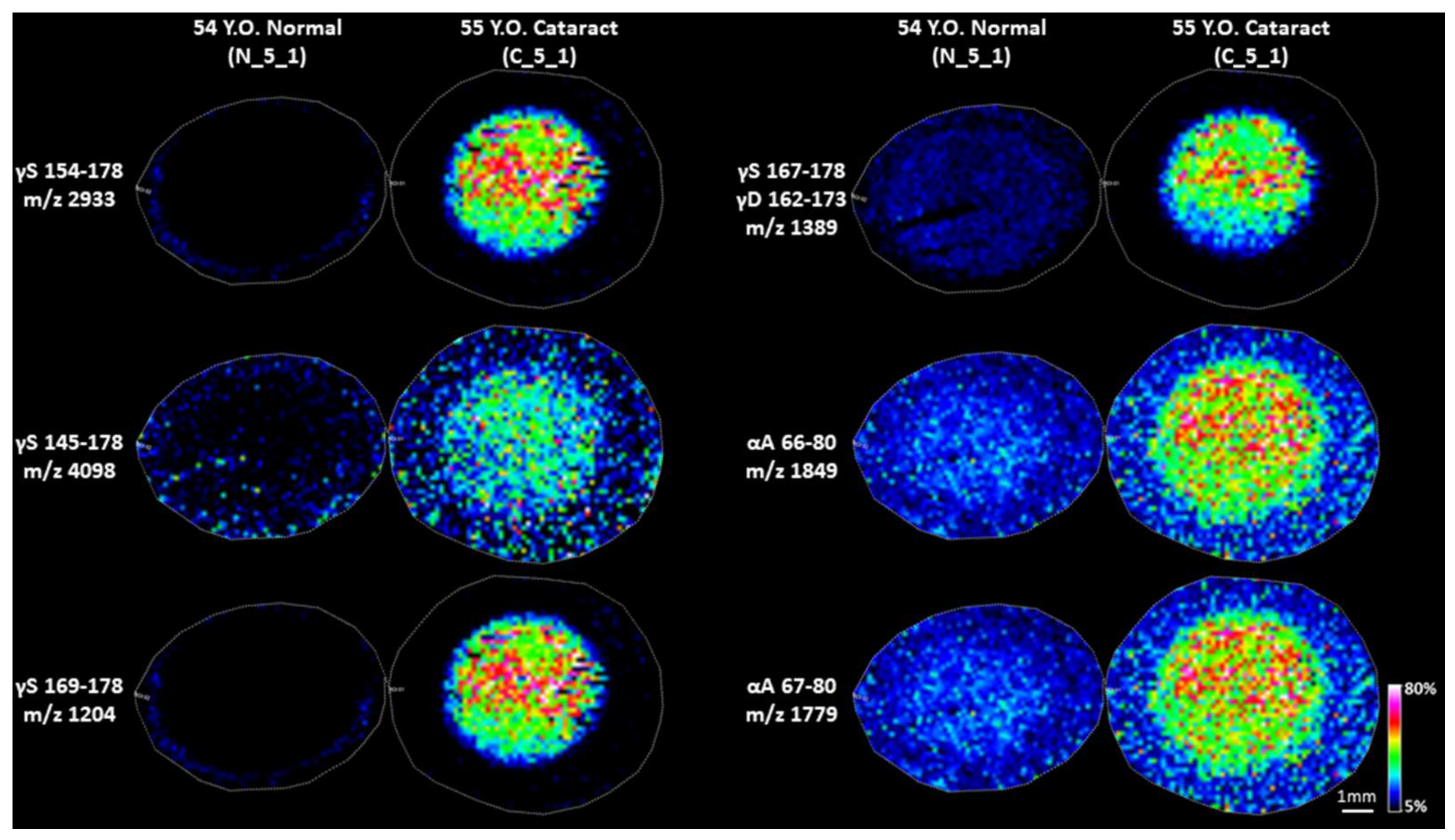
3. Results
3.1. Detection of Low Molecular Signals in Cataract Nucleus
3.2. Identification of Signals Detected by IMS and Further Confirmation of Cataract Specificity
3.3. Low Molecular Weight Signals in Normal Lenses
4. Discussion
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Congdon, N.; Vingerling, J.R.; Klein, B.E.K.; West, S.; Friedman, D.; Kempen, J.; O’Colmain, B.; Wu, S.-Y.; Taylor, H.R. Eye Diseases Prevalence Research Group Prevalence of Cataract and Pseudophakia/Aphakia Among Adults in the United States. Arch. Ophthalmol. 2004, 122, 487–494. [Google Scholar] [CrossRef] [PubMed]
- Truscott, R.J.; Schey, K.L.; Friedrich, M.G. Old Proteins in Man: A Field in its Infancy. Trends Biochem. Sci. 2016, 41, 654–664. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Takemoto, L. Quantitation of C-terminal modification of alpha-A crystallin during aging of the human lens. Exp. Eye Res. 1995, 60, 721–724. [Google Scholar] [PubMed]
- Wilmarth, P.A.; Tanner, S.; Dasari, S.; Nagalla, S.R.; Riviere, A.; Bafna, V.; Pevzner, P.A.; David, L.L. Age-related changes in human crystallins determined from comparative analysis of post-translational modifications in young and aged lens: Does deamidation contribute to crystallin insolubility? J. Proteome Res. 2006, 5, 2554–2566. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Grey, A.C.; Schey, K.L. Age-Related Changes in the Spatial Distribution of Human Lens α-Crystallin Products by MALDI Imaging Mass Spectrometry. Investig. Opthalmology Vis. Sci. 2009, 50, 4319–4329. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Takemoto, L.; Boyle, D. Increased deamidation of asparagine during human senile cataractogenesis. Mol. Vis. 2000, 6, 164–168. [Google Scholar]
- Hooi, M.Y.; Raftery, M.J.; Truscott, R.J. Racemization of two proteins over our lifespan: Deamidation of asparagine 76 in gammaS crystallin is greater in cataract than in normal lenses across the age range. Investig. Ophthalmol. Vis. Sci. 2012, 53, 3554–3561. [Google Scholar] [CrossRef]
- Lyon, Y.A.; Sabbah, G.M.; Julian, R.R. Differences in α-crystallin isomerization reveal the activity of protein isoaspartyl methyltransferase (PIMT) in the nucleus and cortex of human lenses. Exp. Eye Res. 2018, 171, 131–141. [Google Scholar] [CrossRef]
- Seeley, E.H.; Caprioli, R.M. Molecular imaging of proteins in tissues by mass spectrometry. Proc. Natl. Acad. Sci. USA 2008, 105, 18126–18131. [Google Scholar] [CrossRef] [Green Version]
- Buchberger, A.R.; Delaney, K.; Johnson, J.; Jillian, J. Mass Spectrometry Imaging: A Review of Emerging Advancements and Future Insights. Anal. Chem. 2017, 90, 240–265. [Google Scholar] [CrossRef]
- Wenke, J.L.; Rose, K.L.; Spraggins, J.M.; Schey, K.L. MALDI Imaging Mass Specrometry Spatially Maps Age-Related Deami-dation and Truncation of Human Lens Aquaporin-0. Investig. Ophthalmol. Vis. Sci. 2015, 56, 7398–7405. [Google Scholar] [CrossRef] [PubMed]
- Anderson, D.M.; Nye-Wood, M.G.; Rose, K.L.; Donaldson, P.J.; Grey, A.C.; Schey, K.L. MALDI imaging mass spectrometry of β- and γ-crystallins in the ocular lens. J. Mass Spectrom. 2020, 55, e4473. [Google Scholar] [CrossRef] [PubMed]
- Santhoshkumar, P.; Udupa, P.; Murugesan, R.; Sharma, K.K. Significance of Interactions of Low Molecular Weight Crystallin Fragments in Lens Aging and Cataract Formation. J. Biol. Chem. 2008, 283, 8477–8485. [Google Scholar] [CrossRef] [Green Version]
- Su, S.-P.; McArthur, J.D.; Aquilina, J.A. Localization of low molecular weight crystallin peptides in the aging human lens using a MALDI mass spectrometry imaging approach. Exp. Eye Res. 2010, 91, 97–103. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Santhoshkumar, P.; Raju, M.; Sharma, K.K. αA-crystallin peptide SDRDKFVIFLDVKHF accumulating in aging lens impairs the function of α-crystallin and induces lens protein aggregation. PLoS ONE. 2011, 6, e19291. [Google Scholar] [CrossRef]
- Giblin, F.J.; Anderson, D.M.; Han, J.; Rose, K.L.; Wang, Z.; Schey, K.L. Acceleration of age-induced proteolysis in the guinea pig lens nucleus by in vivo exposure to hyperbaric oxygen: A mass spectrometry analysis. Exp. Eye Res. 2021, 210, 108697. [Google Scholar] [CrossRef]
- Schey, K.L.; Anderson, D.M.; Rose, K.L. Spatially-Directed Protein Identification from Tissue Sections by Top-Down LC-MS/MS with Electron Transfer Dissociation. Anal. Chem. 2013, 85, 6767–6774. [Google Scholar] [CrossRef] [Green Version]
- Srivastava, O.; Srivastava, K.; Chaves, J.; Gill, A. Post-translationally modified human lens crystallin fragments show aggregation in vitro. Biochem. Biophys. Rep. 2017, 10, 94–131. [Google Scholar] [CrossRef]
- Friedrich, M.G.; Lam, J.; Truscott, R.J. Degradation of an old human protein: Age-dependent cleavage of gammaS-crystallin generates a peptide that binds to cell membranes. J. Biol. Chem. 2012, 287, 39012–39020. [Google Scholar] [CrossRef] [Green Version]
- Quax-Jeuken, Y.; Driessen, H.; Leunissen, J.; Quax, W.; de Jong, W.; Bloemendal, H. beta s-Crystallin: Structure and evolution of a distinct member of the beta gamma-superfamily. EMBO J. 1985, 4, 2597–2602. [Google Scholar] [CrossRef]
- Thomson, J.A.; Augusteyn, R.C. Ontogeny of human lens crystallins. Exp. Eye Res. 1985, 40, 393–410. [Google Scholar] [CrossRef] [PubMed]
- Vendra, V.P.R.; Khan, I.; Chandani, S.; Muniyandi, A.; Balasubramanian, D. Gamma crystallins of the human eye lens. Biochim. Biophys. Acta (BBA)-Gen. Subj. 2016, 1860, 333–343. [Google Scholar] [CrossRef] [PubMed]
- Schey, K.L.; Wang, Z.; Friedrich, M.G.; Garland, D.L.; Truscott, R.J. Spatiotemporal changes in the human lens proteome: Critical insights into long-lived proteins. Prog. Retin. Eye Res. 2019, 76, 100802. [Google Scholar] [CrossRef] [PubMed]
- David, L.L.; Shearer, T.R. Role of proteolysis in lenses: A review. Lens. Eye Toxic. Res. 1989, 6, 725–747. [Google Scholar] [PubMed]
- Gupta, R.; Chen, J.; Srivastava, O.P. A serine-type protease activity of human lens betaA3-crystallin is responsible for its auto-degradation. Mol. Vis. 2010, 16, 2242–2252. [Google Scholar] [PubMed]
- Voorter, C.E.; de Haard-Hoekman, W.A.; Oetelaar, P.J.V.D.; Bloemendal, H.; De Jong, W.W. Spontaneous peptide bond cleavage in aging alpha-crystallin through a succinimide intermediate. J. Biol. Chem. 1988, 263, 19020–19023. [Google Scholar] [CrossRef]
- Friedrich, M.G.; Wang, Z.; Schey, K.L.; Truscott, R.J.W. Spontaneous Cleavage at Glu and Gln Residues in Long-Lived Proteins. ACS Chem. Biol. 2021, 16, 2244–2254. [Google Scholar] [CrossRef]
- Ball, L.E.; Garland, D.L.; Crouch, R.K.; Schey, K.L. Post-translational Modifications of Aquaporin 0 (AQP0) in the Normal Human Lens: Spatial and Temporal Occurrence. Biochemistry 2004, 43, 9856–9865. [Google Scholar] [CrossRef]
- Geiger, T.; Clarke, S. Deamidation, isomerization, and racemization at asparaginyl and aspartyl residues in peptides. Suc-cinimide-linked reactions that contribute to protein degradation. J. Biol. Chem. 1987, 262, 785–794. [Google Scholar]
- Clarke, N.D. A proposed mechanism for the self-splicing of proteins. Proc. Natl. Acad. Sci. USA 1994, 91, 11084–11088. [Google Scholar] [CrossRef] [Green Version]
- Hains, P.G.; Truscott, R.J.W. Post-Translational Modifications in the Nuclear Region of Young, Aged, and Cataract Human Lenses. J. Proteome Res. 2007, 6, 3935–3943. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Protein | Predicted m/z a | Observed m/z | Error (ppm) |
|---|---|---|---|
| Gamma S 145–178 | 4096.1992 | 4096.1988 | 0.1 |
| Gamma S 147–178 | 3777.0347 | 3777.0319 | 0.74 |
| Gamma S 149–178 (pyroGlu) | 3546.8856 | 3546.8859 | 0.1 |
| Gamma S 150–178 | 3435.8536 | 3435.8524 | 0.35 |
| Gamma S 153-178 | 3046.6221 | 3046.6204 | 0.56 |
| Gamma S 154–178 | 2931.5952 | 2931.5942 | 0.35 |
| Gamma S 155–178 | 2803.5002 | 2803.4978 | 0.87 |
| Gamma S 157–178 | 2546.3627 | 2546.3614 | 0.52 |
| Gamma S 164–178 | 1587.8602 | 1587.8594 | 0.47 |
| Gamma S 167–178 | 1388.7645 | 1388.7641 | 0.25 |
| Gamma S 169–178 | 1204.6797 | 1204.6793 | 0.29 |
| Beta A3 188–215 | 3389.6747 | 3389.6704 | 1.3 |
| Beta A3 189–215 | 3252.6158 | 3252.6129 | 0.89 |
| Beta A3 190–215 | 3195.5943 | 3195.5924 | 0.6 |
| Beta A3 191–215 | 3138.5729 | 3138.5714 | 0.5 |
| Beta A3 196–215 | 2409.2494 | 2409.2462 | 1.3 |
| Gamma D 162–173 | 1388.8121 | 1388.8109 | 0.83 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Schey, K.L.; Wang, Z.; Rose, K.L.; Anderson, D.M.G. Imaging Cataract-Specific Peptides in Human Lenses. Cells 2022, 11, 4042. https://doi.org/10.3390/cells11244042
Schey KL, Wang Z, Rose KL, Anderson DMG. Imaging Cataract-Specific Peptides in Human Lenses. Cells. 2022; 11(24):4042. https://doi.org/10.3390/cells11244042
Chicago/Turabian StyleSchey, Kevin L., Zhen Wang, Kristie L. Rose, and David M. G. Anderson. 2022. "Imaging Cataract-Specific Peptides in Human Lenses" Cells 11, no. 24: 4042. https://doi.org/10.3390/cells11244042