
Melatonin Protects Mitochondrial Function and Inhibits Oxidative Damage against the Decline of Human Oocytes Development Caused by Prolonged Cryopreservation
 and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Ethics Statement
2.2. Chemicals and Reagents
2.3. Immature Oocyte Collection and In Vitro Maturation
2.4. Study Design
2.5. Oocyte Vitrification and Warming
2.6. Developmental Competence
2.7. Single-Oocyte Proteomic Assay
2.8. Detection of ATP, ROS, and GSH Levels in Oocytes
2.9. Statistical Analysis
3. Results
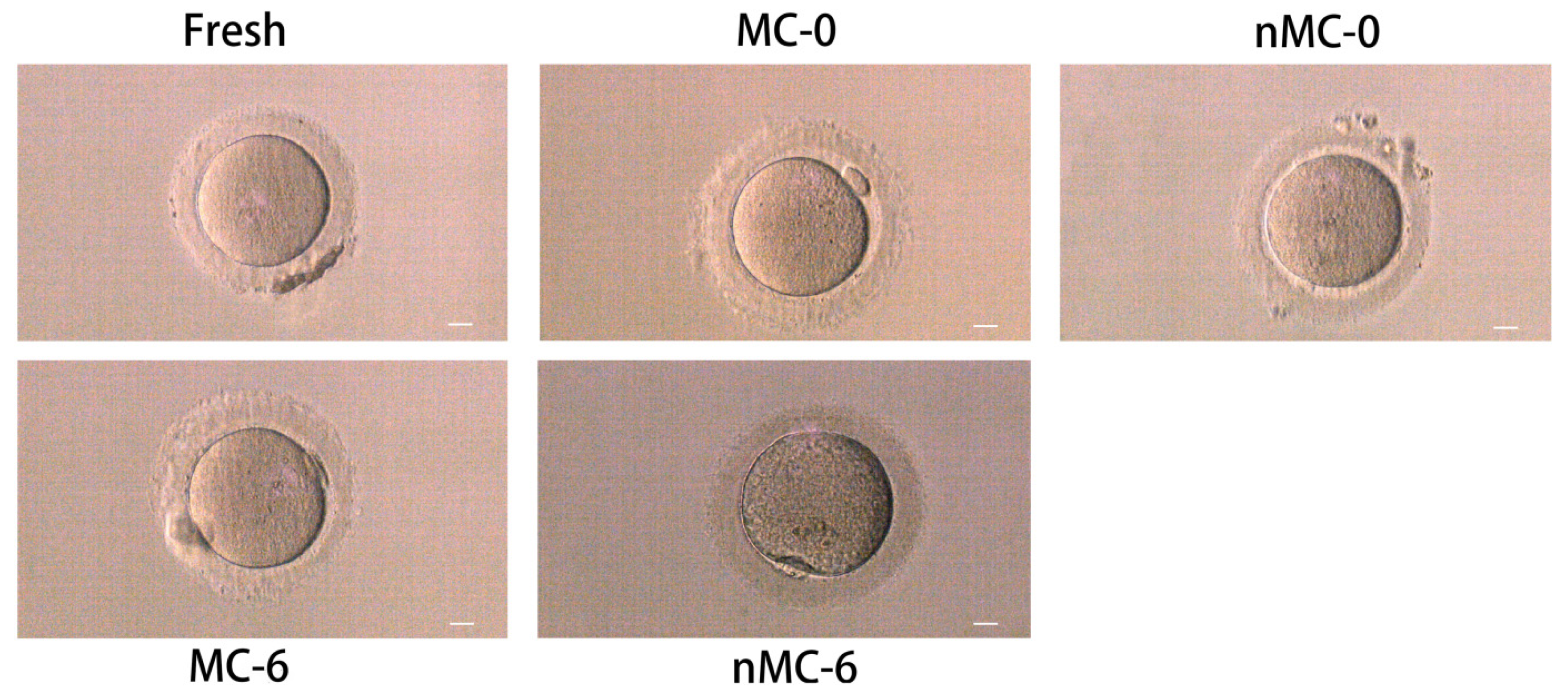
3.1. Morphological Observation of Vitrified–Warmed Human Oocytes
3.2. MT Protected the Developmental Competence of Human Oocytes in Prolonged Cryopreservation
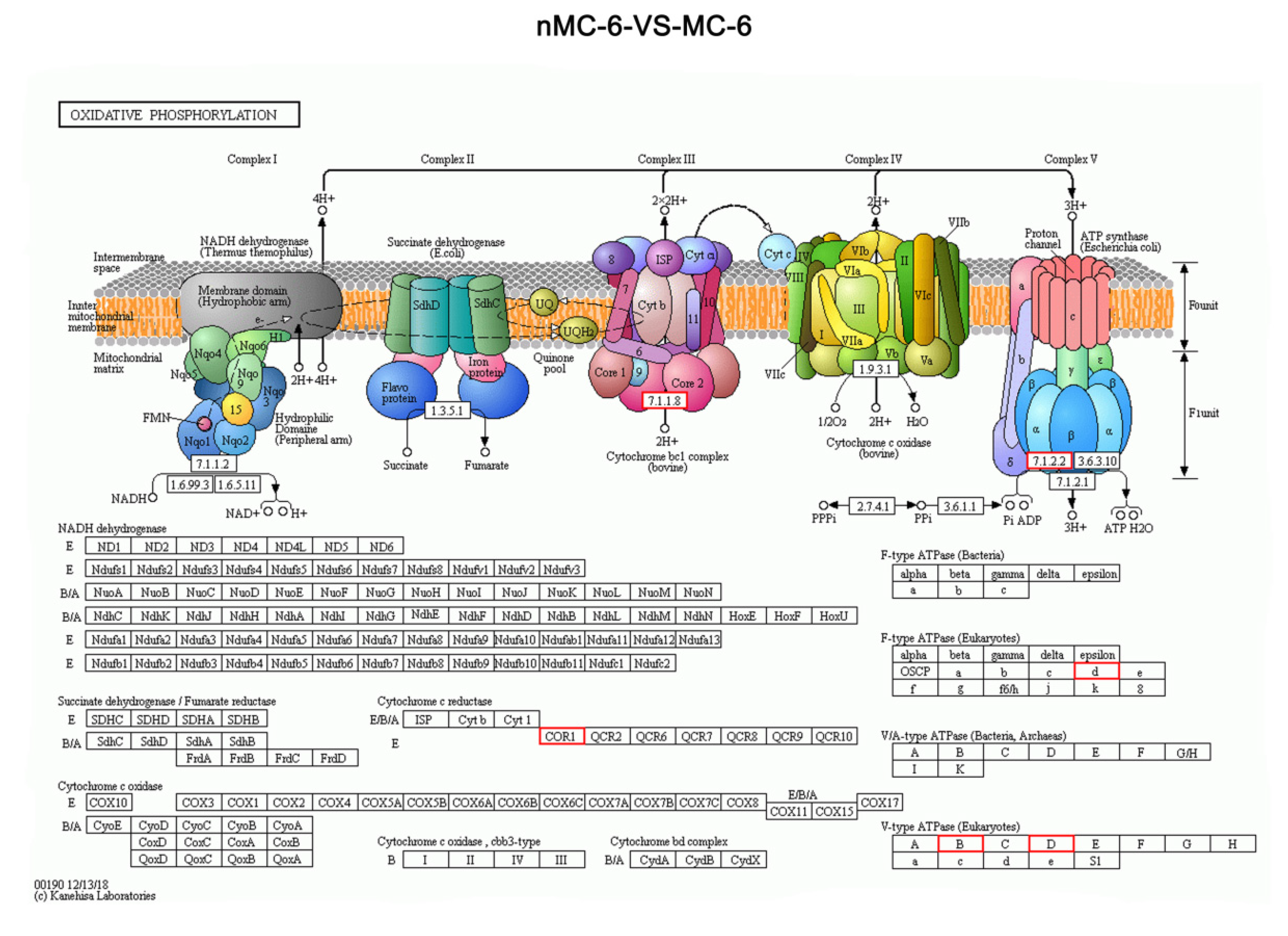
3.3. Micro-Sample Global Proteomic Expression Characteristics of Human Oocytes in Prolonged Cryopreservation
3.4. MT Increased ATP Production of Human Oocytes in PROLONGED CRYOPReservation
3.5. MT Inhibited the Aggravation of ROS Accumulation and Suppressed the Decline in GSH Production in Human Oocytes under Prolonged Cryopreservation
3.6. ROS/GSH Ratio in Prolonged Cryopreserved Human Oocytes
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Practice Committees of the American Society for Reproductive Medicine and the Society for Assisted Reproductive Technology. Mature oocyte cryopreservation: A guideline. Fertil. Steril. 2013, 99, 37–43. [Google Scholar] [CrossRef] [PubMed]
- Song, W.Y.; Peng, Z.F.; Chen, X.M.; Jin, H.X.; Yao, G.D.; Shi, S.L.; Yang, H.; Zhang, X.; Sun, Y.P. Effects of Vitrification on Outcomes of In Vivo-Mature, In Vitro-Mature and Immature Human Oocytes. Cell Physiol. Biochem. 2016, 38, 2053–2062. [Google Scholar] [CrossRef] [PubMed]
- Molina, I.; Gómez, J.; Balasch, S.; Pellicer, N.; Novella-Maestre, E. Osmotic-shock produced by vitrification solutions improves immature human oocytes in vitro maturation. Reprod. Biol. Endocrinol. 2016, 14, 27. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, L.; Xue, X.; Yan, J.; Yan, L.-Y.; Jin, X.-H.; Zhu, X.-H.; He, Z.-Z.; Liu, J.; Li, R.; Qiao, J. L-proline: A highly effective cryoprotectant for mouse oocyte vitrification. Sci. Rep. 2016, 6, 26326. [Google Scholar] [CrossRef]
- Shirazi, A.; Naderi, M.M.; Hassanpour, H.; Heidari, M.; Borjian, S.; Sarvari, A.; Akhondi, M.M. The effect of ovine oocyte vitrification on expression of subset of genes involved in epigenetic modifications during oocyte maturation and early embryo development. Theriogenology 2016, 86, 2136–2146. [Google Scholar] [CrossRef] [PubMed]
- Coello, A.; Campos, P.; Remohí, J.; Meseguer, M.; Cobo, A. A combination of hydroxypropyl cellulose and trehalose as supplementation for vitrification of human oocytes a retrospective cohort study. J. Assist. Reprod. Genet. 2016, 33, 413–421. [Google Scholar] [CrossRef] [Green Version]
- Testart, J.; Lassalle, B.; Forman, R.; Gazengel, A.; Belaisch-Allart, J.; Hazout, A.; Rainhorn, J.-D.; Frydman, R. Factors influencing the success rate of human embryo freezing in an in vitro fertilization and embryo transfer program. Fertil. Steril. 1987, 48, 107–112. [Google Scholar] [CrossRef]
- Riggs, R.; Mayer, J.; Dowling-Lacey, D.; Chi, T.-F.; Jones, E.; Oehninger, S. Does storage time influence postthaw survival and pregnancy outcome? An analysis of 11,768 cryopreserved human embryos. Fertil. Steril. 2008, 93, 109–115. [Google Scholar] [CrossRef]
- Goldman, K.N.; Kramer, Y.; Hodes-Wertz, B.; Noyes, N.; McCaffrey, C.; Grifo, J.A. Long-term cryopreservation of human oocytes does not increase embryonic aneuploidy. Fertil. Steril. 2015, 103, 662–668. [Google Scholar] [CrossRef]
- Stigliani, S.; Moretti, S.; Anserini, P.; Casciano, I.; Venturini, P.L.; Scaruffi, P. Storage time does not modify the gene expression profile of cryopreserved human metaphase II oocytes. Hum. Reprod. 2015, 30, 2519–2526. [Google Scholar] [CrossRef]
- Huo, Y.; Yuan, P.; Qin, Q.; Yan, Z.; Yan, L.; Liu, P.; Li, R.; Yan, J.; Qiao, J. Effects of vitrification and cryostorage duration on single-cell RNA-Seq profiling of vitrified-thawed human metaphase II oocytes. Front. Med. 2020, 15, 144–154. [Google Scholar] [CrossRef] [PubMed]
- Tamura, H.; Jozaki, M.; Tanabe, M.; Shirafuta, Y.; Mihara, Y.; Shinagawa, M.; Tamura, I.; Maekawa, R.; Sato, S.; Taketani, T.; et al. Importance of Melatonin in Assisted Reproductive Technology and Ovarian Aging. Int. J. Mol. Sci. 2020, 21, 1135. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Reiter, R.J.; Sharma, R. Central and peripheral actions of melatonin on reproduction in seasonal and continuous breeding mammals. Gen. Comp. Endocrinol. 2020, 300, 113620. [Google Scholar] [CrossRef] [PubMed]
- Hao, T.; Xu, X.; Hao, H.; Du, W.; Pang, Y.; Zhao, S.; Zou, H.; Yang, S.; Zhu, H.; Yang, Y.; et al. Melatonin improves the maturation and developmental ability of bovine oocytes by up-regulating GJA4 to enhance gap junction intercellular communication. Reprod. Fertil. Dev. 2021, 33, 760. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.; Mu, Y.; Ding, D.; Zou, W.; Li, X.; Chen, B.; Leung, P.C.; Chang, H.; Zhu, Q.; Wang, K.; et al. Melatonin improves the effect of cryopreservation on human oocytes by suppressing oxidative stress and maintaining the permeability of the oolemma. J. Pineal Res. 2020, 70, e12707. [Google Scholar] [CrossRef] [PubMed]
- Reiter, R.J.; Tan, D.X.; Galano, A. Melatonin: Exceeding Expectations. Physiology 2014, 29, 325–333. [Google Scholar] [CrossRef] [Green Version]
- Hardeland, R. Melatonin and the theories of aging: A critical appraisal of melatonin’s role in antiaging mechanisms. J. Pineal Res. 2013, 55, 325–356. [Google Scholar] [CrossRef] [PubMed]
- Li, W.; Cheng, K.; Zhang, Y.; Meng, Q.; Zhu, S.; Zhou, G. No effect of exogenous melatonin on development of cryopreserved metaphase II oocytes in mouse. J. Anim. Sci. Biotechnol. 2015, 6, 42. [Google Scholar] [CrossRef] [Green Version]
- An, Q.; Peng, W.; Cheng, Y.; Lu, Z.; Zhou, C.; Zhang, Y.; Su, J. Melatonin supplementation during in vitro maturation of oocyte enhances subsequent development of bovine cloned embryos. J. Cell. Physiol. 2019, 234, 17370–17381. [Google Scholar] [CrossRef]
- Ortiz, A.; Espino, J.; Bejarano, I.; Lozano, G.M.; Monllor, F.; García, J.F.; Pariente, J.A.; Rodríguez, A.B. High endogenous melatonin concentrations enhance sperm quality and short-term in vitro exposure to melatonin improves aspects of sperm motility. J. Pineal Res. 2010, 50, 132–139. [Google Scholar] [CrossRef]
- Chaithrashree, A.R.; Ingole, S.D.; Dighe, V.D.; Nagvekar, A.S.; Bharucha, S.; Dagli, N.R.; Kekan, P.M.; Kharde, S.D. Effect of melatonin on bovine sperm characteristics and ultrastructure changes following cryopreservation. Vet.-Med. Sci. 2019, 6, 177–186. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Malo, C.; Gil, L.; Gonzalez, N.; Martínez, F.; Cano, R.; de Blas, I.; Espinosa, E. Anti-oxidant supplementation improves boar sperm characteristics and fertility after cryopreservation: Comparison between cysteine and rosemary (Rosmarinus officinalis). Cryobiology 2010, 61, 142–147. [Google Scholar] [CrossRef] [PubMed]
- Fadl, A.M.; Ghallab, A.R.M.; Abou-Ahmed, M.M.; Moawad, A.R. Melatonin can improve viability and functional integrity of cooled and frozen/thawed rabbit spermatozoa. Reprod. Domest. Anim. 2020, 56, 103–111. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.J.; Zhang, Y.; Jia, G.X.; Meng, Q.G.; Bunch, T.D.; Liu, G.S.; Zhu, S.E.; Xhou, G.B. Effect of melatonin supplementation on cryopreserved sperm quality in mouse. Cryoletters 2016, 37, 115–122. [Google Scholar] [PubMed]
- Pariz, J.R.; Ranéa, C.; Monteiro, R.A.C.; Evenson, D.P.; Drevet, J.R.; Hallak, J. Melatonin and Caffeine Supplementation Used, Respectively, as Protective and Stimulating Agents in the Cryopreservation of Human Sperm Improves Survival, Viability, and Motility after Thawing compared to Traditional TEST-Yolk Buffer. Oxidative Med. Cell. Longev. 2019, 10, 6472945. [Google Scholar] [CrossRef]
- Zou, H.; Chen, B.; Ding, D.; Gao, M.; Chen, D.; Liu, Y.; Hao, Y.; Zou, W.; Ji, D.; Zhou, P.; et al. Melatonin promotes the development of immature oocytes from the COH cycle into healthy offspring by protecting mitochondrial function. J. Pineal Res. 2019, 68, e12621. [Google Scholar] [CrossRef]
- Zhang, Z.; Wang, T.; Hao, Y.; Panhwar, F.; Chen, Z.; Zou, W.; Ji, D.; Chen, B.; Zhou, P.; Zhao, G.; et al. Effects of trehalose vitrification and artificial oocyte activation on the development competence of human immature oocytes. Cryobiology 2017, 74, 43–49. [Google Scholar] [CrossRef]
- Moawad, A.R.; Tan, S.L.; Taketo, T. Beneficial effects of glutathione supplementation during vitrification of mouse oocytes at the germinal vesicle stage on their preimplantation development following maturation and fertilization in vitro. Cryobiology 2017, 76, 98–103. [Google Scholar] [CrossRef]
- Müller-Höcker, J.; Schäfer, S.; Weis, S.; Münscher, C.; Strowitzki, T. Morphological-cytochemical and molecular genetic analyses of mitochondria in isolated human oocytes in the reproductive age. Mol. Hum. Reprod. 1996, 2, 951–958. [Google Scholar] [CrossRef]
- Brieger, K.; Schiavone, S.; Miller, F.J., Jr.; Krause, K.-H. Reactive oxygen species: From health to disease. Swiss Med. Wkly. 2012, 142, w13659. [Google Scholar] [CrossRef]
- Dai, J.; Wu, C.; Muneri, C.W.; Niu, Y.; Zhang, S.; Rui, R.; Zhang, D. Changes in mitochondrial function in porcine vitrified MII-stage oocytes and their impacts on apoptosis and developmental ability. Cryobiology 2015, 71, 291–298. [Google Scholar] [CrossRef] [PubMed]
- Hales, K.G. The machinery of mitochondrial fusion, division, and distribution, and emerging connections to apoptosis. Mitochondrion 2004, 4, 285–308. [Google Scholar] [CrossRef] [PubMed]
- Barros, F.D.D.A.; Adona, P.R.; Guemra, S.; Damião, B.C.M. Oxidative homeostasis in oocyte competence for in vitro embryo development. Anim. Sci. J. 2019, 90, 1343–1349. [Google Scholar] [CrossRef] [PubMed]
- Liu, T.; Sun, L.; Zhang, Y.; Wang, Y.; Zheng, J. Imbalanced GSH/ROS and sequential cell death. J. Biochem. Mol. Toxicol. 2021, 36, e22942. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Li, W.; Ma, Y.; Wang, D.; Zhao, X.; Zeng, C.; Zhang, M.; Zeng, X.; Meng, Q.; Zhou, G. Improved development by melatonin treatment after vitrification of mouse metaphase II oocytes. Cryobiology 2016, 73, 335–342. [Google Scholar] [CrossRef]
- Bianchi, V.; Coticchio, G.; Fava, L.; Flamigni, C.; Borini, A. Meiotic spindle imaging in human oocytes frozen with a slow freezing procedure involving high sucrose concentration. Hum. Reprod. 2005, 20, 1078–1083. [Google Scholar] [CrossRef] [Green Version]
- Zhao, X.-M.; Du, W.-H.; Wang, D.; Hao, H.-S.; Liu, Y.; Qin, T.; Zhu, H.-B. Effect of cyclosporine pretreatment on mitochondrial function in vitrified bovine mature oocytes. Fertil. Steril. 2011, 95, 2786–2788. [Google Scholar] [CrossRef]
- Rocha-Frigoni, N.A.; Leão, B.C.; Dall’Acqua, P.C.; Mingoti, G.Z. Improving the cytoplasmic maturation of bovine oocytes matured in vitro with intracellular and/or extracellular antioxidants is not associated with increased rates of embryo development. Theriogenology 2016, 86, 1897–1905. [Google Scholar] [CrossRef] [Green Version]
- Zhao, X.-M.; Hao, H.-S.; Du, W.-H.; Zhao, S.-J.; Wang, H.-Y.; Wang, N.; Wang, D.; Liu, Y.; Qin, T.; Zhu, H.-B. Melatonin inhibits apoptosis and improves the developmental potential of vitrified bovine oocytes. J. Pineal Res. 2015, 60, 132–141. [Google Scholar] [CrossRef]
- Jou, M.-J. Melatonin preserves the transient mitochondrial permeability transition for protection during mitochondrial Ca2+ stress in astrocyte. J. Pineal Res. 2011, 50, 427–435. [Google Scholar] [CrossRef]
- Zhu, Q.; Wang, K.; Zhang, C.; Chen, B.; Zou, H.; Zou, W.; Xue, R.; Ji, D.; Yu, Z.; Rao, B.; et al. Effect of melatonin on the clinical outcome of patients with repeated cycles after failed cycles of in vitro fertilization and intracytoplasmic sperm injection. Zygote 2022, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Tomas, C.; Orava, M.; Tuomivaara, L.; Martikainen, H. Low pregnancy rate is achieved in patients treated with intracytoplasmic sperm injection due to previous low or failed fertilization in in-vitro fertilization. Hum. Reprod. 1998, 13, 65–70. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gardner, D.; Schoolcraft, W.B. Culture and transfer of human blastocysts. Curr. Opin. Obstet. Gynecol. 1999, 11, 307–311. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| F | nMC-0 | MC-0 | nMC-6 | MC-6 | |
|---|---|---|---|---|---|
| IVM-MII (n) | 50 | 58 | 60 | 55 | 57 |
| Fertilization rate (%) | 86.00% (43/50) a,c | 62.07% (36/58) | 83.33% (50/60) b | 58.18% (32/55) | 64.91% (37/57) |
| Cleavage rate (%) | 86.05% (37/43) | 80.56% (29/36) | 84.00% (42/50) | 68.75% (22/32) | 67.57% (25/37) |
| High-quality cleavage embryo rate (%) | 64.86% (24/37) b,c | 34.48% (10/29) | 61.90% (26/42) b | 18.18% (4/22) | 48.00% (12/25) |
| Blastocyst rate (%) | 51.35% (19/37) b,c | 17.24% (5/29) | 40.48% (17/42) b | 9.09% (2/22) | 28.00% (7/25) |
| F vs. nMC-0 | F vs. MC-0 | F vs. nMC-6 | F vs. MC-6 | nMC-0 vs. MC-0 | nMC-6 vs. MC-6 | nMC-0 vs. nMC-6 | MC-0 vs. MC-6 | |
|---|---|---|---|---|---|---|---|---|
| Number of differential proteins | 70 | 133 | 148 | 17 | 58 | 160 | 53 | 212 |
| Number of differential proteins associated with mitochondrial function | 14 | 20 | 21 | 1 | 11 | 19 | 2 | 32 |
| Up | 0 | 0 | 0 | 2 | 8 | 19 | 1 | 32 |
| Down | 14 | 20 | 21 | 0 | 3 | 0 | 2 | 0 |
| Primary_protein_ID | Mean Intensity (nMT-6) | Mean Intensity (MT-6) | Ratio | p-Value | Class | Description |
|---|---|---|---|---|---|---|
| p|Q9Y5K8|VATD_HUMAN | 57,736,499.74 | 155,072,081.7 | 2.685858728 | 0.012634037 | Up | V-type proton ATPase subunit D OS = Homo sapiens OX = 9606 GN = ATP6V1D PE = 1 SV = 1 |
| sp|O75947|ATP5H_HUMAN | 757,186,306.8 | 2,746,405,384 | 3.627119718 | 0.027221848 | Up | ATP synthase subunit d, mitochondrial OS = Homo sapiens OX = 9606 GN = ATP5PD PE = 1 SV = 3 |
| sp|P21281|VATB2_HUMAN | 1,576,694,555 | 2,656,550,766 | 1.684886117 | 0.043175449 | Up | V-type proton ATPase subunit B, brain isoform OS = Homo sapiens OX = 9606 GN = ATP6V1B2 PE = 1 SV=3 |
| sp|P31930|QCR1_HUMAN | 1,021,503,942 | 2,837,296,332 | 2.777567676 | 0.045073322 | Up | Cytochrome b-c1 complex subunit 1, mitochondrial OS=Homo sapiens OX = 9606 GN = UQCRC1 PE = 1 SV = 3 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhu, Q.; Ding, D.; Yang, H.; Zou, W.; Yang, D.; Wang, K.; Zhang, C.; Chen, B.; Ji, D.; Hao, Y.; et al. Melatonin Protects Mitochondrial Function and Inhibits Oxidative Damage against the Decline of Human Oocytes Development Caused by Prolonged Cryopreservation. Cells 2022, 11, 4018. https://doi.org/10.3390/cells11244018
Zhu Q, Ding D, Yang H, Zou W, Yang D, Wang K, Zhang C, Chen B, Ji D, Hao Y, et al. Melatonin Protects Mitochondrial Function and Inhibits Oxidative Damage against the Decline of Human Oocytes Development Caused by Prolonged Cryopreservation. Cells. 2022; 11(24):4018. https://doi.org/10.3390/cells11244018
Chicago/Turabian StyleZhu, Qi, Ding Ding, Han Yang, Weiwei Zou, Dandan Yang, Kaijuan Wang, Chao Zhang, Beili Chen, Dongmei Ji, Yan Hao, and et al. 2022. "Melatonin Protects Mitochondrial Function and Inhibits Oxidative Damage against the Decline of Human Oocytes Development Caused by Prolonged Cryopreservation" Cells 11, no. 24: 4018. https://doi.org/10.3390/cells11244018