
The Protective Effects of Human Embryonic Stem Cell-Derived Mesenchymal Stem Cells in Noise-Induced Hearing Loss of Rats
 and
and
Abstract
:1. Introduction
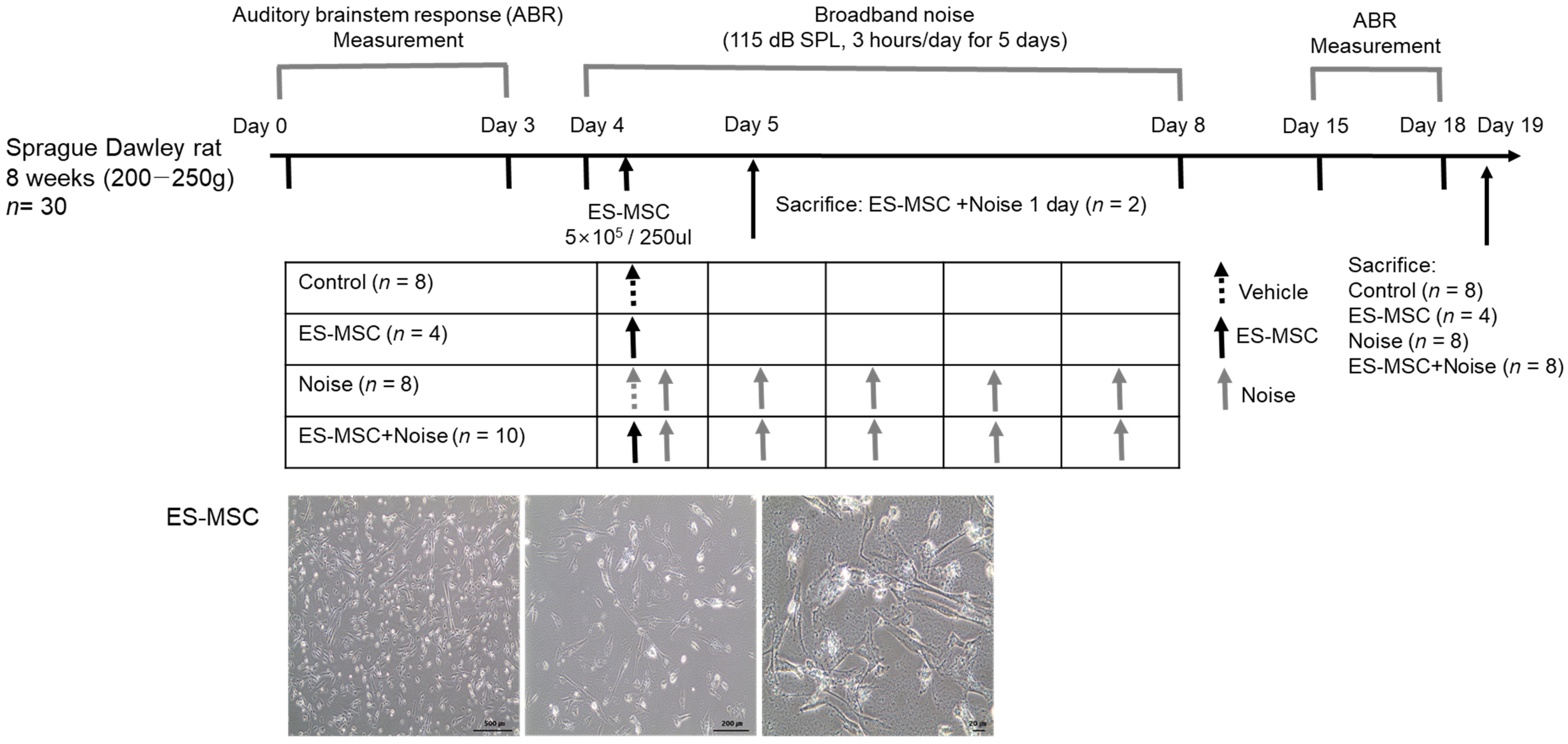
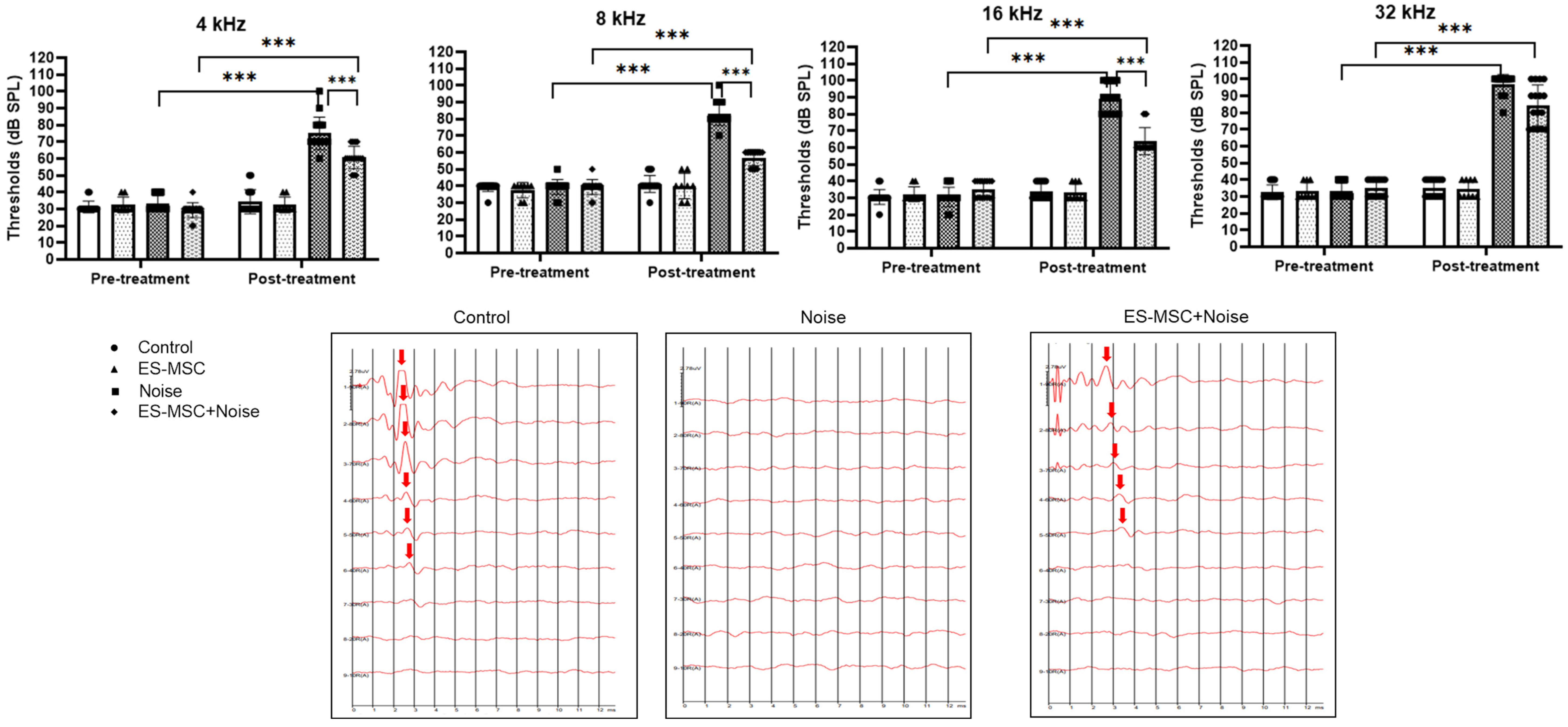
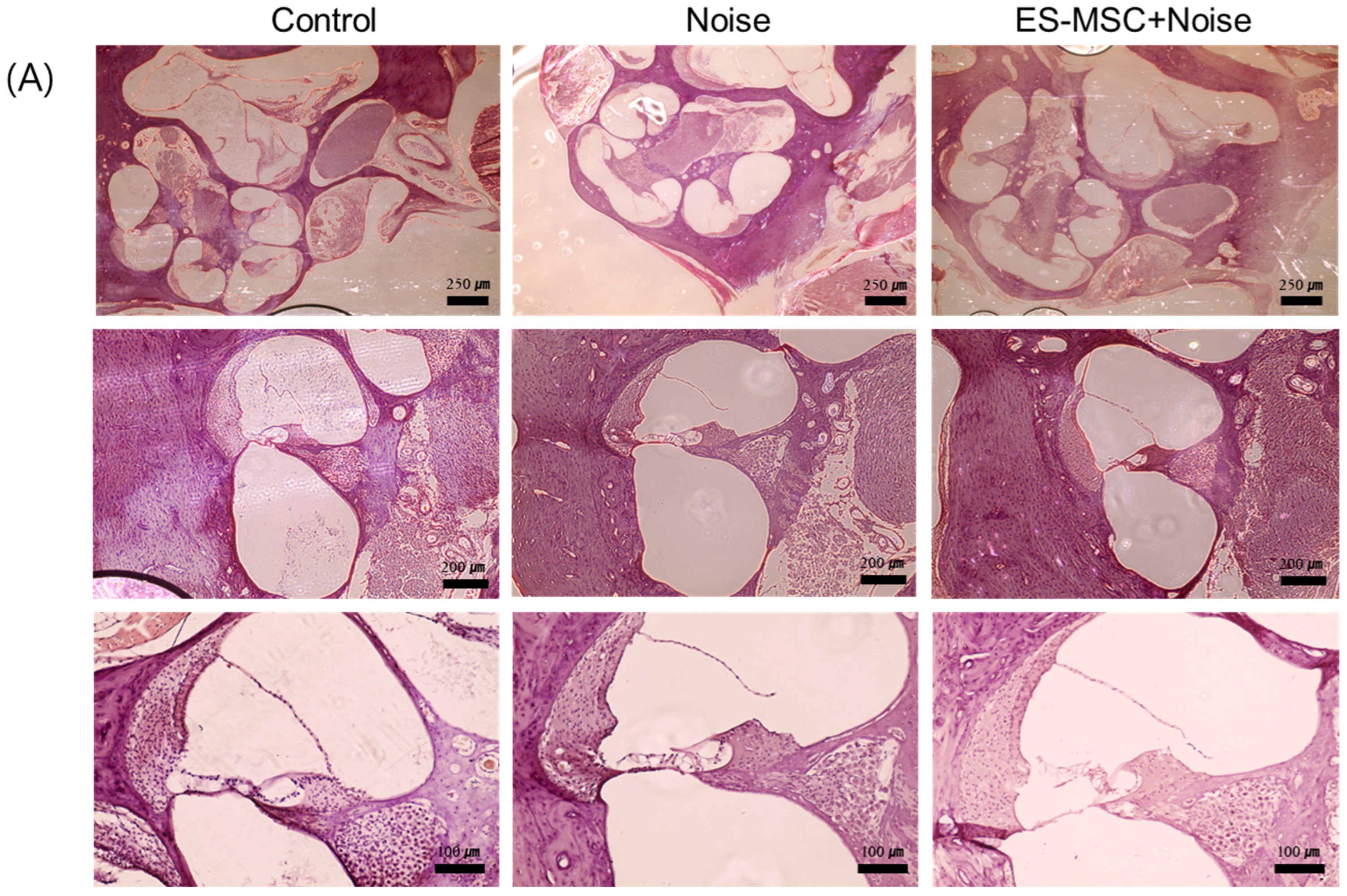
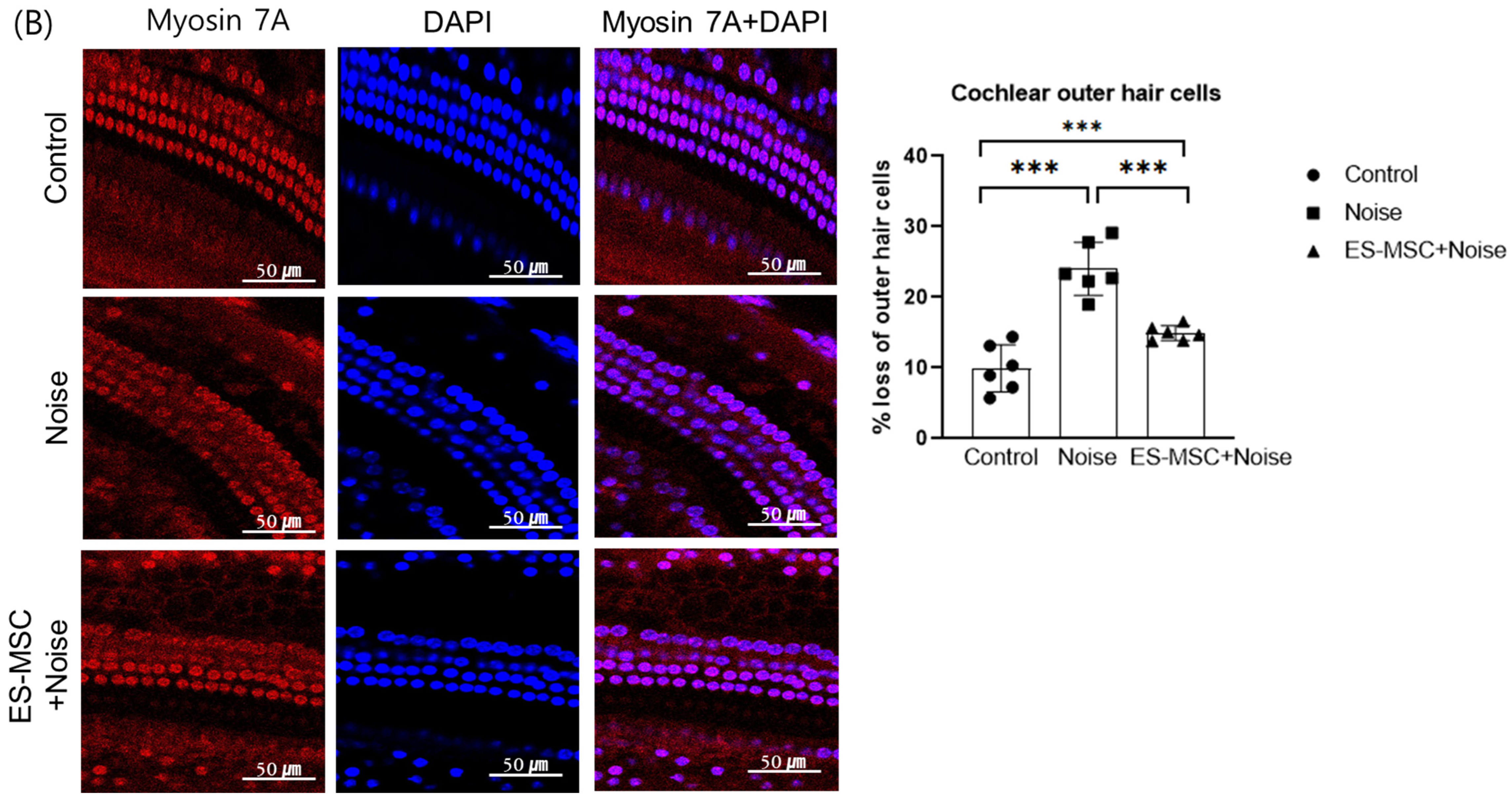
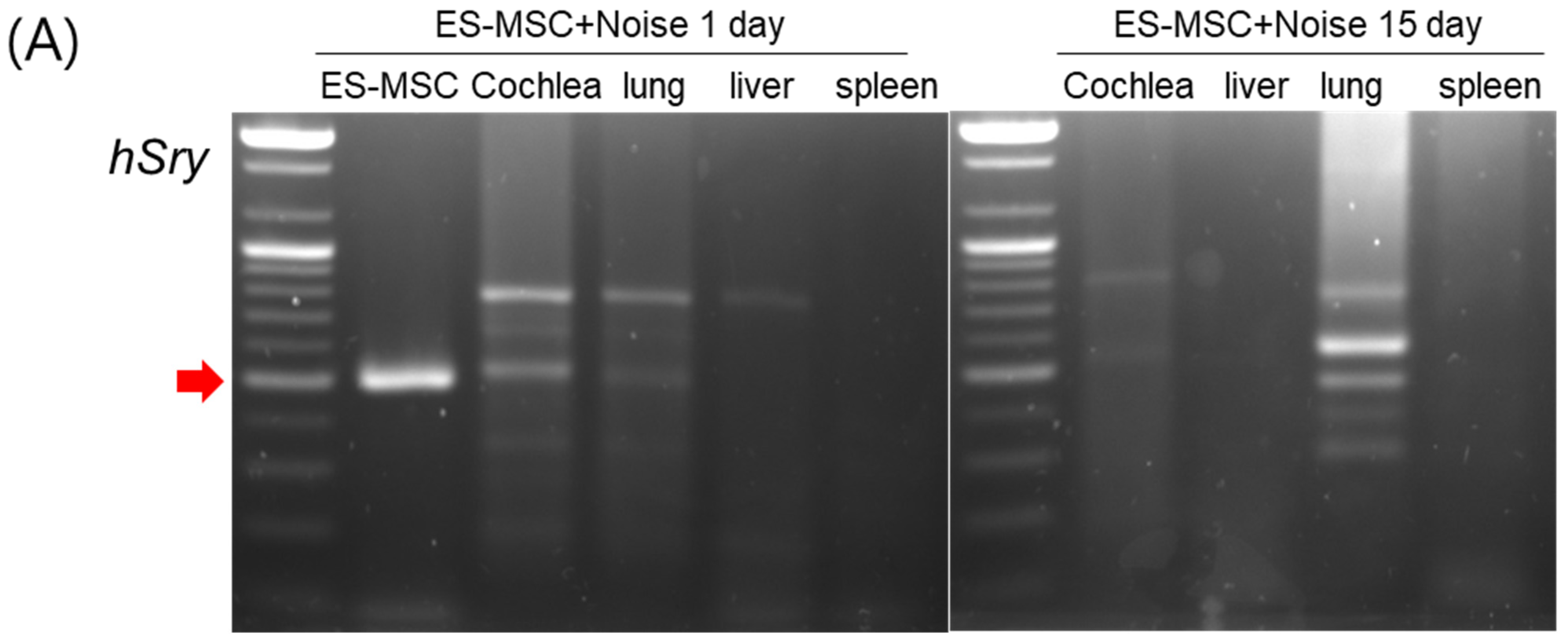
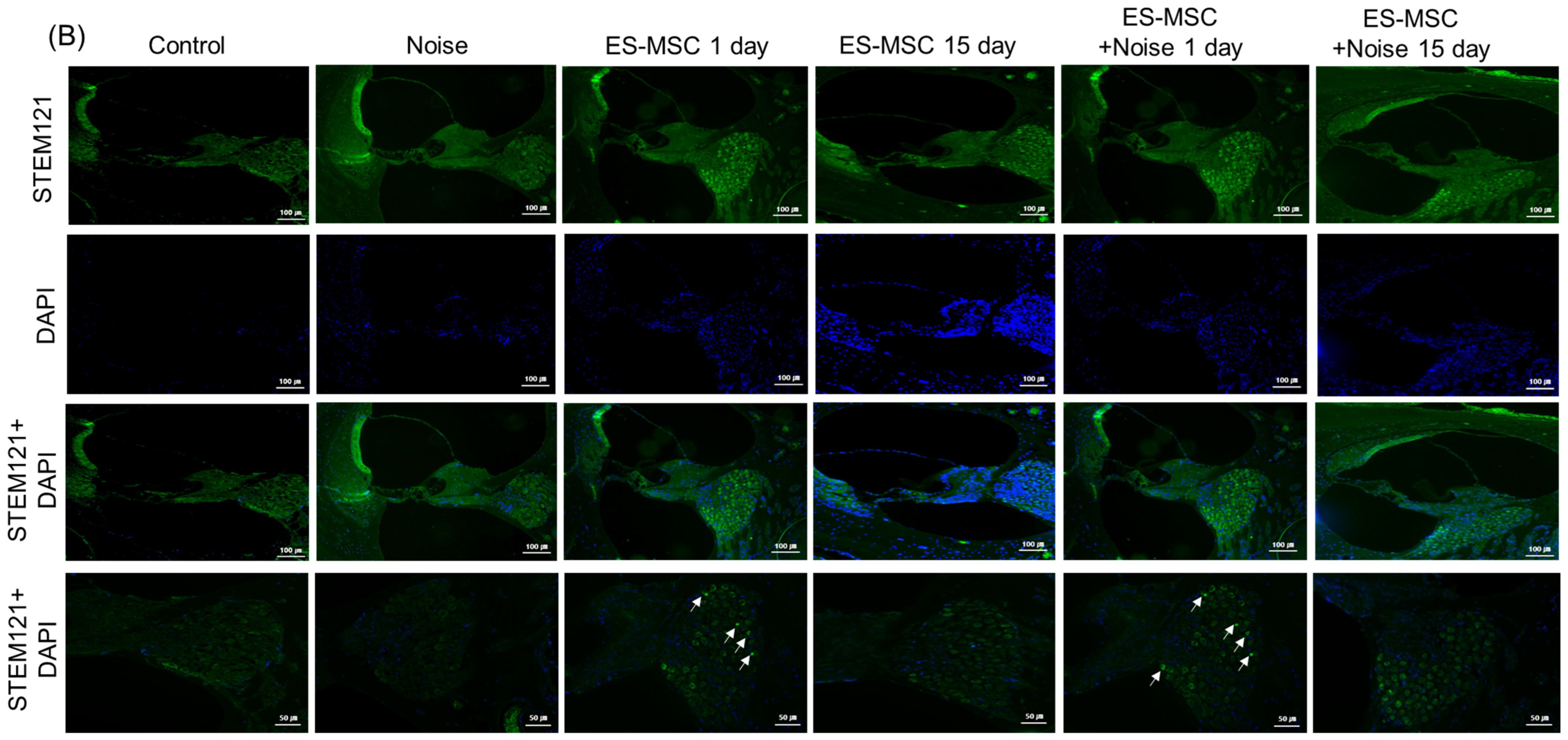
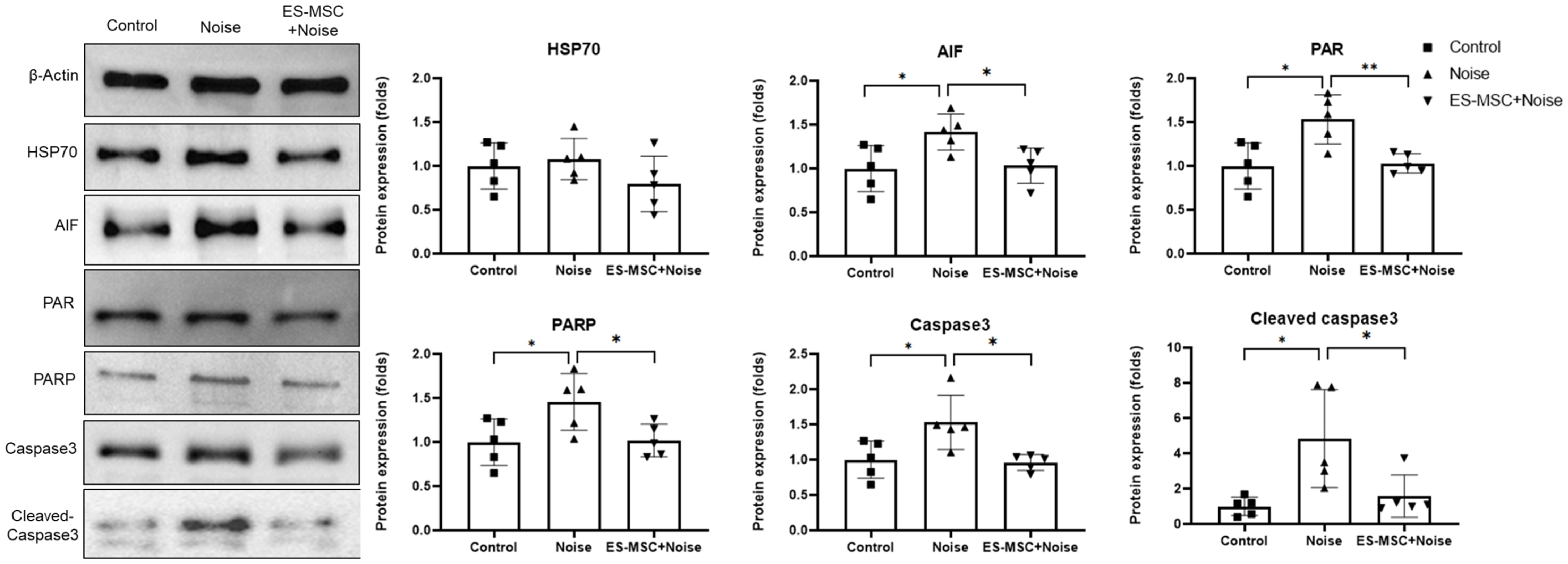
2. Results
3. Discussion
4. Materials and Methods
4.1. Ethical Approval
4.2. Preparation of Human Embryonic Stem-Derived Mesenchymal Stem Cells (ES-MSCs)
4.3. ES-MSC Injection and Noise Exposure
4.4. Examination of Auditory Evoked Potentials
4.5. Examination of Cochlear Histology
4.6. Distributions of Injected Human ES-MSCs
4.7. Expression Levels of Heat Shock Protein 70 (HSP70) and Apoptosis and Parthanatos-Related Markers
4.8. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Kim, S.Y.; Min, C.; Yoo, D.M.; Chang, J.; Lee, H.J.; Park, B.; Choi, H.G. Hearing Impairment Increases Economic Inequality. Clin. Exp. Otorhinolaryngol. 2021, 14, 278–286. [Google Scholar] [CrossRef] [PubMed]
- Wu, P.Z.; O’Malley, J.T.; de Gruttola, V.; Liberman, M.C. Primary Neural Degeneration in Noise-Exposed Human Cochleas: Correlations with Outer Hair Cell Loss and Word-Discrimination Scores. J. Neurosci. 2021, 41, 4439–4447. [Google Scholar] [CrossRef] [PubMed]
- Yamaguchi, T.; Yoneyama, M.; Ogita, K. Calpain inhibitor alleviates permanent hearing loss induced by intense noise by preventing disruption of gap junction-mediated intercellular communication in the cochlear spiral ligament. Eur. J. Pharmacol. 2017, 803, 187–194. [Google Scholar] [CrossRef] [PubMed]
- Mao, H.; Chen, Y. Noise-Induced Hearing Loss: Updates on Molecular Targets and Potential Interventions. Neural Plast. 2021, 2021, 4784385. [Google Scholar] [CrossRef]
- Kurabi, A.; Keithley, E.M.; Housley, G.D.; Ryan, A.F.; Wong, A.C. Cellular mechanisms of noise-induced hearing loss. Hear. Res. 2017, 349, 129–137. [Google Scholar] [CrossRef] [PubMed]
- Milon, B.; Shulman, E.D.; So, K.S.; Cederroth, C.R.; Lipford, E.L.; Sperber, M.; Sellon, J.B.; Sarlus, H.; Pregernig, G.; Shuster, B.; et al. A cell-type-specific atlas of the inner ear transcriptional response to acoustic trauma. Cell Rep. 2021, 36, 109758. [Google Scholar] [CrossRef]
- Forouzanfar, F.; Asgharzade, S. MicroRNAs in Noise-Induced Hearing Loss and their Regulation by Oxidative Stress and Inflammation. Curr. Drug. Targets 2020, 21, 1216–1224. [Google Scholar] [CrossRef]
- Shih, C.P.; Kuo, C.Y.; Lin, Y.Y.; Lin, Y.C.; Chen, H.K.; Wang, H.; Chen, H.C.; Wang, C.H. Inhibition of Cochlear HMGB1 Expression Attenuates Oxidative Stress and Inflammation in an Experimental Murine Model of Noise-Induced Hearing Loss. Cells 2021, 10, 810. [Google Scholar] [CrossRef]
- Fetoni, A.R.; Paciello, F.; Rolesi, R.; Paludetti, G.; Troiani, D. Targeting dysregulation of redox homeostasis in noise-induced hearing loss: Oxidative stress and ROS signaling. Free Radic. Biol. Med. 2019, 135, 46–59. [Google Scholar] [CrossRef]
- He, Z.; Ding, Y.; Mu, Y.; Xu, X.; Kong, W.; Chai, R.; Chen, X. Stem Cell-Based Therapies in Hearing Loss. Front. Cell. Dev. Biol. 2021, 9, 730042. [Google Scholar] [CrossRef]
- Sekiya, T.; Holley, M.C. Cell Transplantation to Restore Lost Auditory Nerve Function is a Realistic Clinical Opportunity. Cell Transplant. 2021, 30, 9636897211035076. [Google Scholar] [CrossRef] [PubMed]
- Maharajan, N.; Cho, G.W.; Jang, C.H. Therapeutic Application of Mesenchymal Stem Cells for Cochlear Regeneration. In Vivo 2021, 35, 13–22. [Google Scholar] [CrossRef] [PubMed]
- Xu, Y.P.; Shan, X.D.; Liu, Y.Y.; Pu, Y.; Wang, C.Y.; Tao, Q.L.; Deng, Y.; Cheng, Y.; Fan, J.P. Olfactory epithelium neural stem cell implantation restores noise-induced hearing loss in rats. Neurosci. Lett. 2016, 616, 19–25. [Google Scholar] [CrossRef] [PubMed]
- Zhang, P.Z.; He, Y.; Jiang, X.W.; Chen, F.Q.; Chen, Y.; Shi, L.; Chen, J.; Chen, X.; Li, X.; Xue, T.; et al. Stem cell transplantation via the cochlear lateral wall for replacement of degenerated spiral ganglion neurons. Hear. Res. 2013, 298, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Hwang, S.H.; Gonzalez-Suarez, A.M.; Stybayeva, G.; Revzin, A. Prospects and Opportunities for Microsystems and Microfluidic Devices in the Field of Otorhinolaryngology. Clin. Exp. Otorhinolaryngol. 2021, 14, 29–42. [Google Scholar] [CrossRef] [PubMed]
- Peyvandi, A.A.; Roozbahany, N.A.; Peyvandi, H.; Abbaszadeh, H.A.; Majdinasab, N.; Faridan, M.; Niknazar, S. Critical role of SDF-1/CXCR4 signaling pathway in stem cell homing in the deafened rat cochlea after acoustic trauma. Neural Regen. Res. 2018, 13, 154–160. [Google Scholar] [CrossRef]
- Peyvandi, A.A.; Abbaszadeh, H.A.; Roozbahany, N.A.; Pourbakht, A.; Khoshsirat, S.; Niri, H.H.; Peyvandi, H.; Niknazar, S. Deferoxamine promotes mesenchymal stem cell homing in noise-induced injured cochlea through PI3K/AKT pathway. Cell Prolif. 2018, 51, e12434. [Google Scholar] [CrossRef] [Green Version]
- Wakabayashi, K.; Nagai, A.; Sheikh, A.M.; Shiota, Y.; Narantuya, D.; Watanabe, T.; Masuda, J.; Kobayashi, S.; Kim, S.U.; Yamaguchi, S. Transplantation of human mesenchymal stem cells promotes functional improvement and increased expression of neurotrophic factors in a rat focal cerebral ischemia model. J. Neurosci. Res. 2010, 88, 1017–1025. [Google Scholar] [CrossRef]
- Zhao, L.R.; Duan, W.M.; Reyes, M.; Keene, C.D.; Verfaillie, C.M.; Low, W.C. Human bone marrow stem cells exhibit neural phenotypes and ameliorate neurological deficits after grafting into the ischemic brain of rats. Exp. Neurol. 2002, 174, 11–20. [Google Scholar] [CrossRef] [Green Version]
- Choi, B.Y.; Song, J.J.; Chang, S.O.; Kim, S.U.; Oh, S.H. Intravenous administration of human mesenchymal stem cells after noise- or drug-induced hearing loss in rats. Acta Otolaryngol. 2012, 132 (Suppl. 1), S94–S102. [Google Scholar] [CrossRef]
- Warnecke, A.; Harre, J.; Shew, M.; Mellott, A.J.; Majewski, I.; Durisin, M.; Staecker, H. Successful Treatment of Noise-Induced Hearing Loss by Mesenchymal Stromal Cells: An RNAseq Analysis of Protective/Repair Pathways. Front. Cell. Neurosci. 2021, 15, 656930. [Google Scholar] [CrossRef] [PubMed]
- Xu, L.; Yuan, S.; Chen, W.; Ma, Y.; Luo, Y.; Guo, W.; Yang, S. Transplantation and Tracking of the Human Umbilical Cord Mesenchymal Stem Cell Labeled with Superparamagnetic Iron Oxide in Deaf Pigs. Anat. Rec. 2020, 303, 494–505. [Google Scholar] [CrossRef] [PubMed]
- Sun, G.W.; Fujii, M.; Matsunaga, T. Functional interaction between mesenchymal stem cells and spiral ligament fibrocytes. J. Neurosci. Res. 2012, 90, 1713–1722. [Google Scholar] [CrossRef]
- Murashita, H.; Tabuchi, K.; Hoshino, T.; Tsuji, S.; Hara, A. The effects of tempol, 3-aminobenzamide and nitric oxide synthase inhibitors on acoustic injury of the mouse cochlea. Hear. Res. 2006, 214, 1–6. [Google Scholar] [CrossRef]
- Marchetta, P.; Mohrle, D.; Eckert, P.; Reimann, K.; Wolter, S.; Tolone, A.; Lang, I.; Wolters, M.; Feil, R.; Engel, J.; et al. Guanylyl Cyclase A/cGMP Signaling Slows Hidden, Age- and Acoustic Trauma-Induced Hearing Loss. Front. Aging Neurosci. 2020, 12, 83. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.; An, R.; Umanah, G.K.; Park, H.; Nambiar, K.; Eacker, S.M.; Kim, B.; Bao, L.; Harraz, M.M.; Chang, C.; et al. A nuclease that mediates cell death induced by DNA damage and poly(ADP-ribose) polymerase-1. Science 2016, 354, aad6872. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Park, D.J.; Park, J.E.; Lee, S.H.; Eliceiri, B.P.; Choi, J.S.; Seo, Y.J. Protective effect of MSC-derived exosomes against cisplatin-induced apoptosis via heat shock protein 70 in auditory explant model. Nanomedicine 2021, 38, 102447. [Google Scholar] [CrossRef]
- Martins, L.F.; Costa, R.O.; Pedro, J.R.; Aguiar, P.; Serra, S.C.; Teixeira, F.G.; Sousa, N.; Salgado, A.J.; Almeida, R.D. Mesenchymal stem cells secretome-induced axonal outgrowth is mediated by BDNF. Sci. Rep. 2017, 7, 4153. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ahn, S.Y.; Sung, D.K.; Kim, Y.E.; Sung, S.; Chang, Y.S.; Park, W.S. Brain-derived neurotropic factor mediates neuroprotection of mesenchymal stem cell-derived extracellular vesicles against severe intraventricular hemorrhage in newborn rats. Stem Cells Transl. Med. 2021, 10, 374–384. [Google Scholar] [CrossRef]
- Blakley, B.W.; Seaman, M.; Alenezi, A. Brain-derived nerve growth factor in the cochlea—A reproducibility study. J. Otolaryngol. Head Neck Surg. 2020, 49, 37. [Google Scholar] [CrossRef]
- Ramekers, D.; Versnel, H.; Grolman, W.; Klis, S.F. Neurotrophins and their role in the cochlea. Hear. Res. 2012, 288, 19–33. [Google Scholar] [CrossRef]
- Han, Z.; Wang, C.P.; Cong, N.; Gu, Y.Y.; Ma, R.; Chi, F.L. Therapeutic value of nerve growth factor in promoting neural stem cell survival and differentiation and protecting against neuronal hearing loss. Mol. Cell. Biochem. 2017, 428, 149–159. [Google Scholar] [CrossRef]
- Tsai, S.C.; Lin, F.C.; Chang, K.H.; Li, M.C.; Chou, R.H.; Huang, M.Y.; Chen, Y.C.; Kao, C.Y.; Cheng, C.C.; Lin, H.C.; et al. The intravenous administration of skin-derived mesenchymal stem cells ameliorates hearing loss and preserves cochlear hair cells in cisplatin-injected mice: SMSCs ameliorate hearing loss and preserve outer hair cells in mice. Hear. Res. 2022, 413, 108254. [Google Scholar] [CrossRef]
- Jun, S.M.; Park, M.; Lee, J.Y.; Jung, S.; Lee, J.E.; Shim, S.H.; Song, H.; Lee, D.R. Single cell-derived clonally expanded mesenchymal progenitor cells from somatic cell nuclear transfer-derived pluripotent stem cells ameliorate the endometrial function in the uterus of a murine model with Asherman’s syndrome. Cell Prolif. 2019, 52, e12597. [Google Scholar] [CrossRef]
- Yoon, S.Y.; Yoon, J.A.; Park, M.; Shin, E.Y.; Jung, S.; Lee, J.E.; Eum, J.H.; Song, H.; Lee, D.R.; Lee, W.S.; et al. Recovery of ovarian function by human embryonic stem cell-derived mesenchymal stem cells in cisplatin-induced premature ovarian failure in mice. Stem Cell Res. Ther. 2020, 11, 255. [Google Scholar] [CrossRef]
- Shin, E.Y.; Kim, D.S.; Lee, M.J.; Lee, A.R.; Shim, S.H.; Baek, S.W.; Han, D.K.; Lee, D.R. Prevention of chemotherapy-induced premature ovarian insufficiency in mice by scaffold-based local delivery of human embryonic stem cell-derived mesenchymal progenitor cells. Stem Cell Res. Ther. 2021, 12, 431. [Google Scholar] [CrossRef] [PubMed]
- Jung, S.K.; Lee, J.E.; Lee, C.W.; Shim, S.H.; Lee, D.R. Rapid Production and Genetic Stability of Human Mesenchymal Progenitor Cells Derived from Human Somatic Cell Nuclear Transfer-Derived Pluripotent Stem Cells. Int. J. Mol. Sci. 2021, 22, 9238. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.Y.; Heo, H.; Kim, D.H.; Kim, H.J.; Oh, S.H. Neural Plastic Changes in the Subcortical Auditory Neural Pathway after Single-Sided Deafness in Adult Mice: A MEMRI Study. Biomed. Res. Int. 2018, 2018, 8624745. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alvarado, J.C.; Fuentes-Santamaria, V.; Jareno-Flores, T.; Blanco, J.L.; Juiz, J.M. Normal variations in the morphology of auditory brainstem response (ABR) waveforms: A study in Wistar rats. Neurosci. Res. 2012, 73, 302–311. [Google Scholar] [CrossRef]
- Kim, S.Y.; Jung, G.; Shim, Y.J.; Koo, J.W. The Novel Peptide Vaccine GV1001 Protects Hearing in a Kanamycin-induced Ototoxicity Mouse Model. Otol. Neurotol. 2018, 39, e731–e737. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Author [Year] | Species | Type of Mesenchymal Stem Cells (MSCs) | Noise Exposure | Route of Delivery | Dosage (Cells) | Time | Outcome |
|---|---|---|---|---|---|---|---|
| SY Kim et al. [2022] | SD rat (8 weeks) | Human ES-MSCs | 115 dB SPL, 3 h/day for 5 days | Tail vein | 5 × 105 | pretreatment | 4, 8, 16 kHz improved |
| Athanasia Warnecke et al. [2021] | C57BL/6 mice (4 weeks) | Human WJ-MSCs | 118 dB SPL, 4 h | PSC | 1 × 103 | posttreatment (48 h) | 4, 8, 32 kHz improved |
| Liangwei Xu et al. [2020] | Bama minipig (4 weeks) | Human WJ-MSCs | 80 times continuous impulse noise | Subarachnoid cavity | 1 × 108 | posttreatment (1 weeks) | 1, 2, 4, 8, 16, 24 kHz improved |
| Ali Asghar Peyvandi et al. [2017] | Wistar rat (12 weeks) | Rat BM-MSCs | 115 dB SPL, 6 h/day for 5 days | RW niche | 1 × 105 | posttreatment | DFO pre-conditioning increases homing ability of MSCs into injured ear |
| Ali Asghar Peyvandi et al. [2017] | Wistar rat (8–12 weeks) | Rat BM-MSCs | 110 dB SPL, 6 h/day for 5 days | RW niche | 1 × 105 | posttreatment | CXCR4 antagonist (AMD3100) decreases homing ability of MSCs into injured ear |
| BY Choi et al. [2012] | SD rat (7–8 weeks) | Human BM-MSCs | 120 dB SPL, 9 h | Tail vein | 4 × 106 | posttreatment (48 h) | No statistically significant difference |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kim, S.Y.; Lee, J.E.; Kang, S.H.; Lee, S.M.; Jeon, J.; Lee, D.R. The Protective Effects of Human Embryonic Stem Cell-Derived Mesenchymal Stem Cells in Noise-Induced Hearing Loss of Rats. Cells 2022, 11, 3524. https://doi.org/10.3390/cells11213524
Kim SY, Lee JE, Kang SH, Lee SM, Jeon J, Lee DR. The Protective Effects of Human Embryonic Stem Cell-Derived Mesenchymal Stem Cells in Noise-Induced Hearing Loss of Rats. Cells. 2022; 11(21):3524. https://doi.org/10.3390/cells11213524
Chicago/Turabian StyleKim, So Young, Jeoung Eun Lee, Sung Hun Kang, So Min Lee, Jiwon Jeon, and Dong Ryul Lee. 2022. "The Protective Effects of Human Embryonic Stem Cell-Derived Mesenchymal Stem Cells in Noise-Induced Hearing Loss of Rats" Cells 11, no. 21: 3524. https://doi.org/10.3390/cells11213524