
Bioinformatics Analysis and Validation of the Role of Lnc-RAB11B-AS1 in the Development and Prognosis of Hepatocellular Carcinoma
Abstract
:1. Introduction
2. Materials and Methods
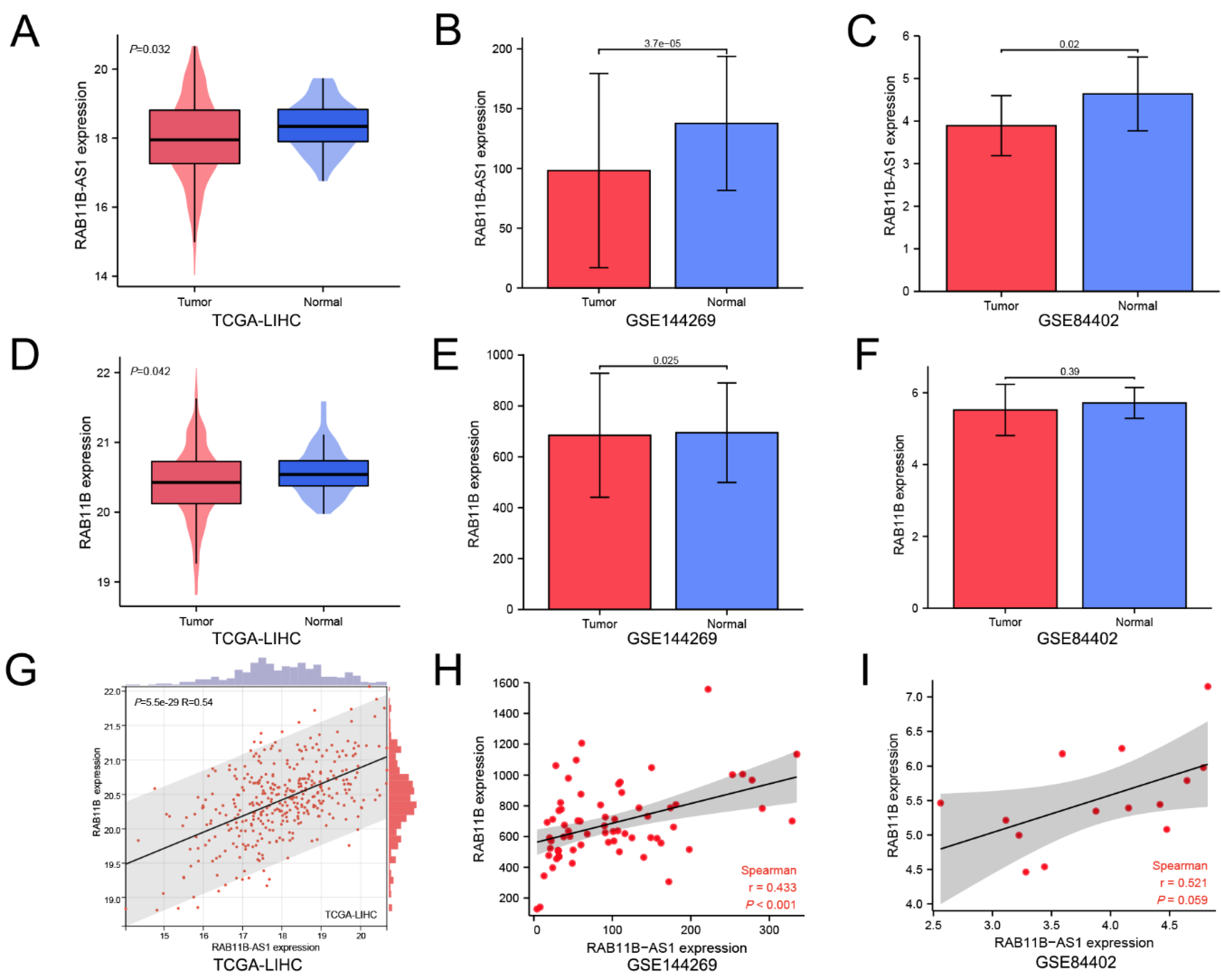
2.1. Data Acquisition and Preprocessing
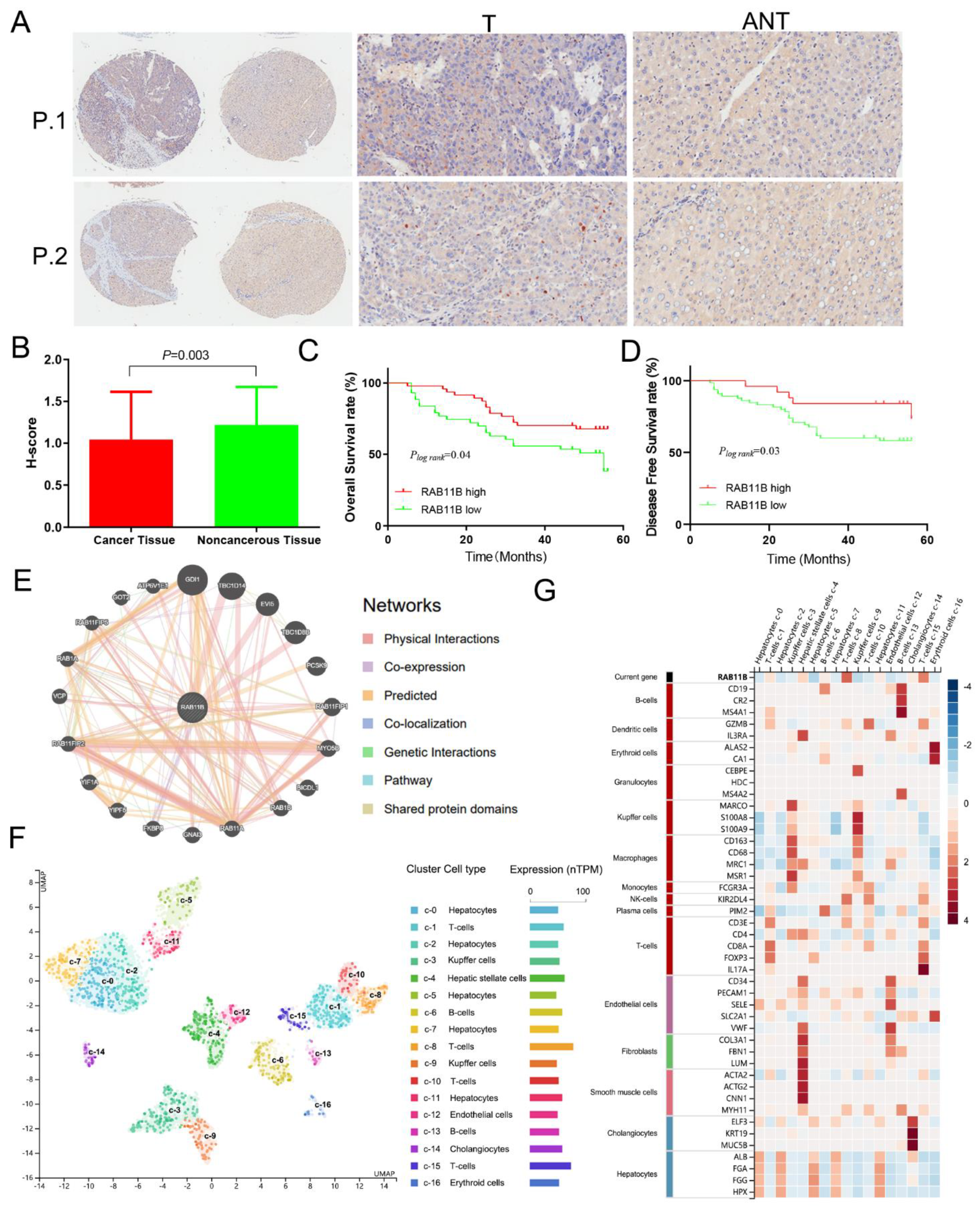
2.2. Immunohistochemistry (IHC) Assay of RAB11B Protein in HCC Tissues
2.3. Protein-Protein Interactions of RAB11B
2.4. Human Protein Atlas (HPA)
2.5. Screening of lnc-RAB11B-AS1 Co-Expressed Genes in HCC and Functional Enrichment Analysis
2.6. GSEA Analysis
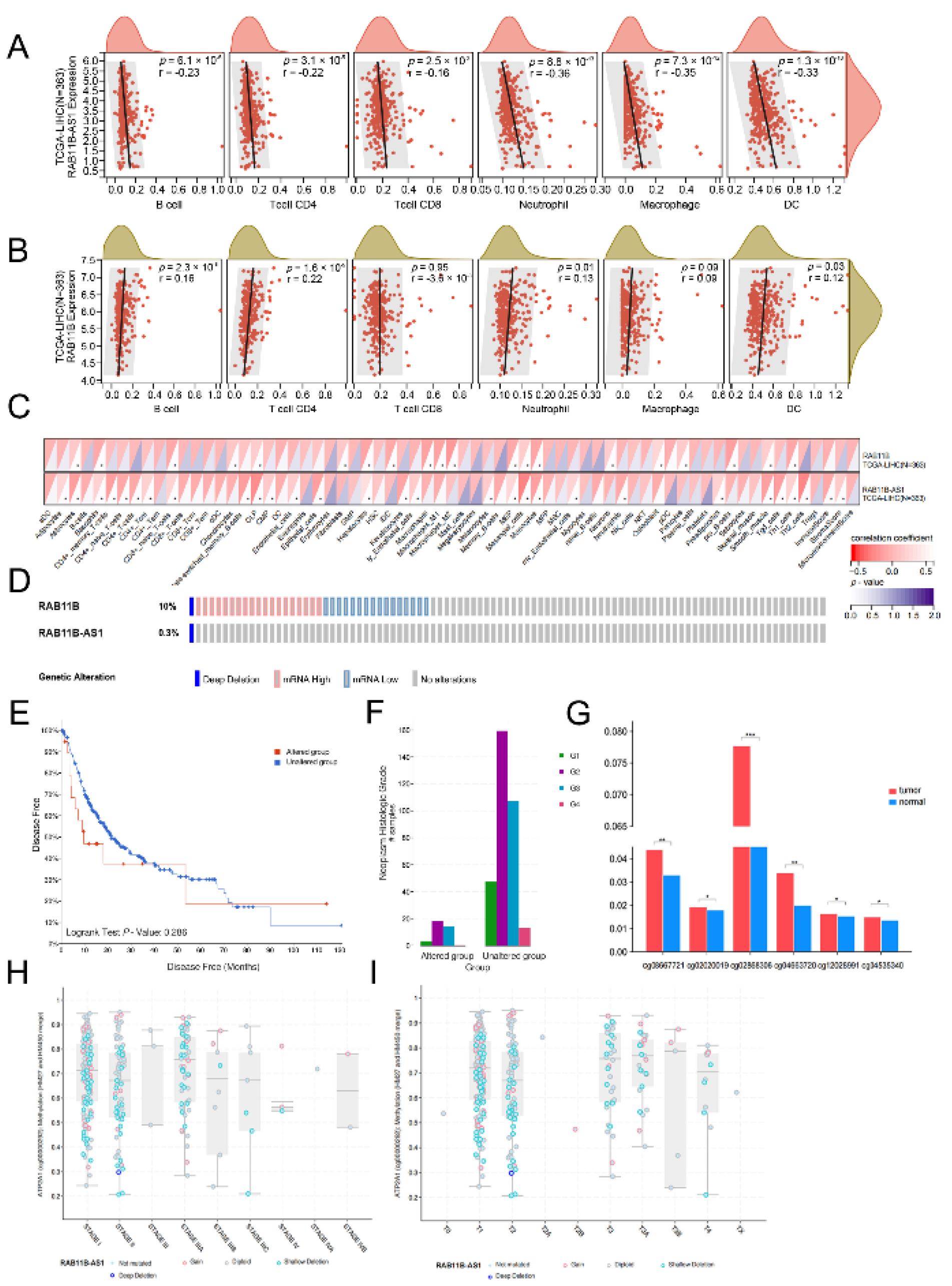
2.7. An Association between lnc-RAB11B-AS1, RAB11B, and Tumor Immune Cell Infiltration
2.8. Alternation of lnc-RAB11B-AS1 and RAB11B in HCC
2.9. Correlation between Methylation and lnc-RAB11B-AS1 Expression
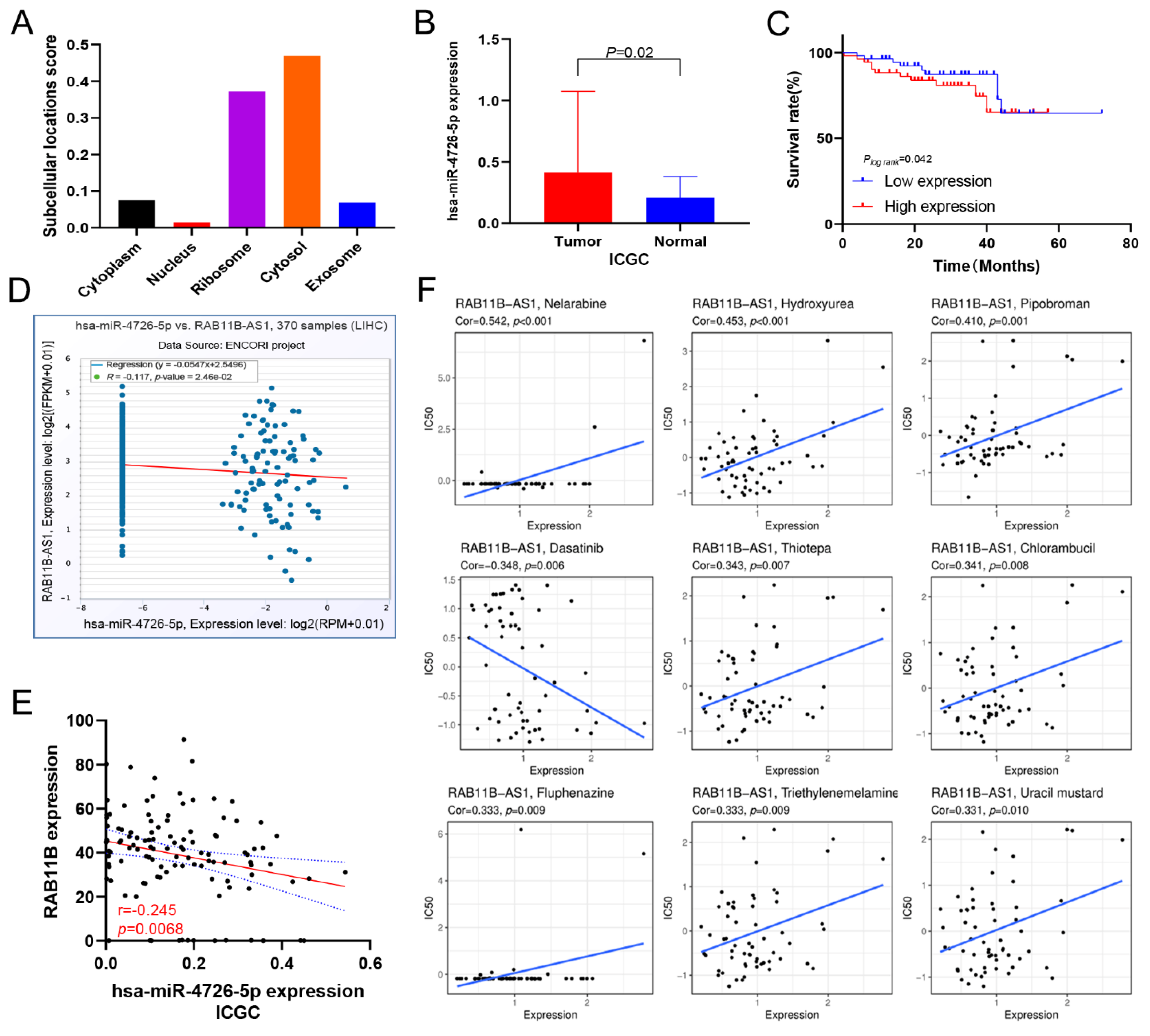
2.10. Drug Susceptibility Analysis
2.11. Prediction of Potential Candidate miRNAs
2.12. Statistical Analysis
3. Results
3.1. Lnc-RAB11B-AS1 Was Markedly Low Expressed and Positively Correlated with RAB11B in HCC
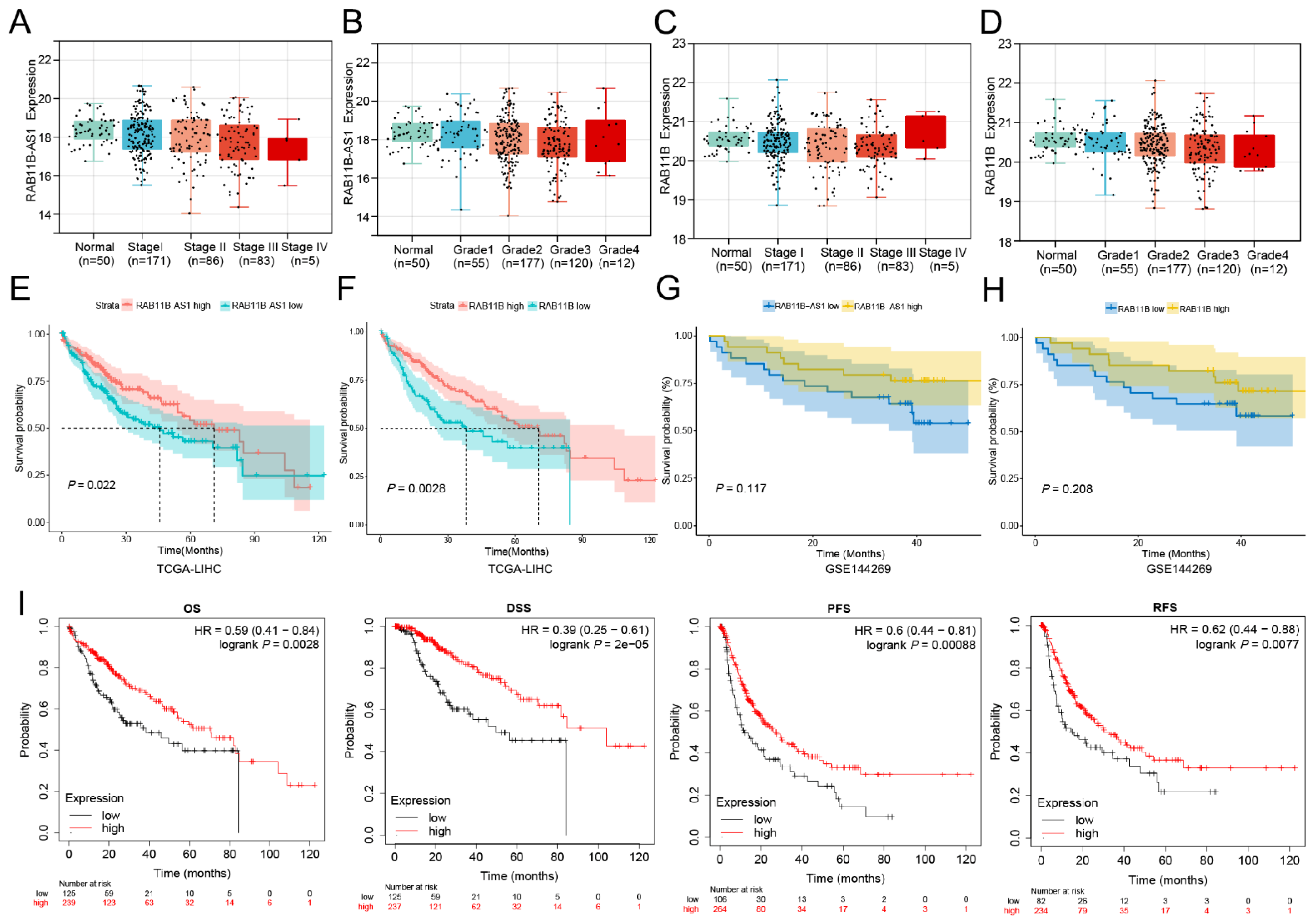
3.2. The Effects of Overexpressed lnc-RAB11B-AS1 and RAB11B on Clinicopathological Characteristics
3.3. The Relationship between lnc-RAB11B-AS1 Expression and Prognosis of HCC Patients
3.4. Cox Regression Model Analysis
3.5. External Validation of RAB11B Protein Expression and Clinicopathologic Features of HCC Patients
3.6. RAB11B-Associated PPI Network
3.7. RAB11B Expression in Different Cells of HCC
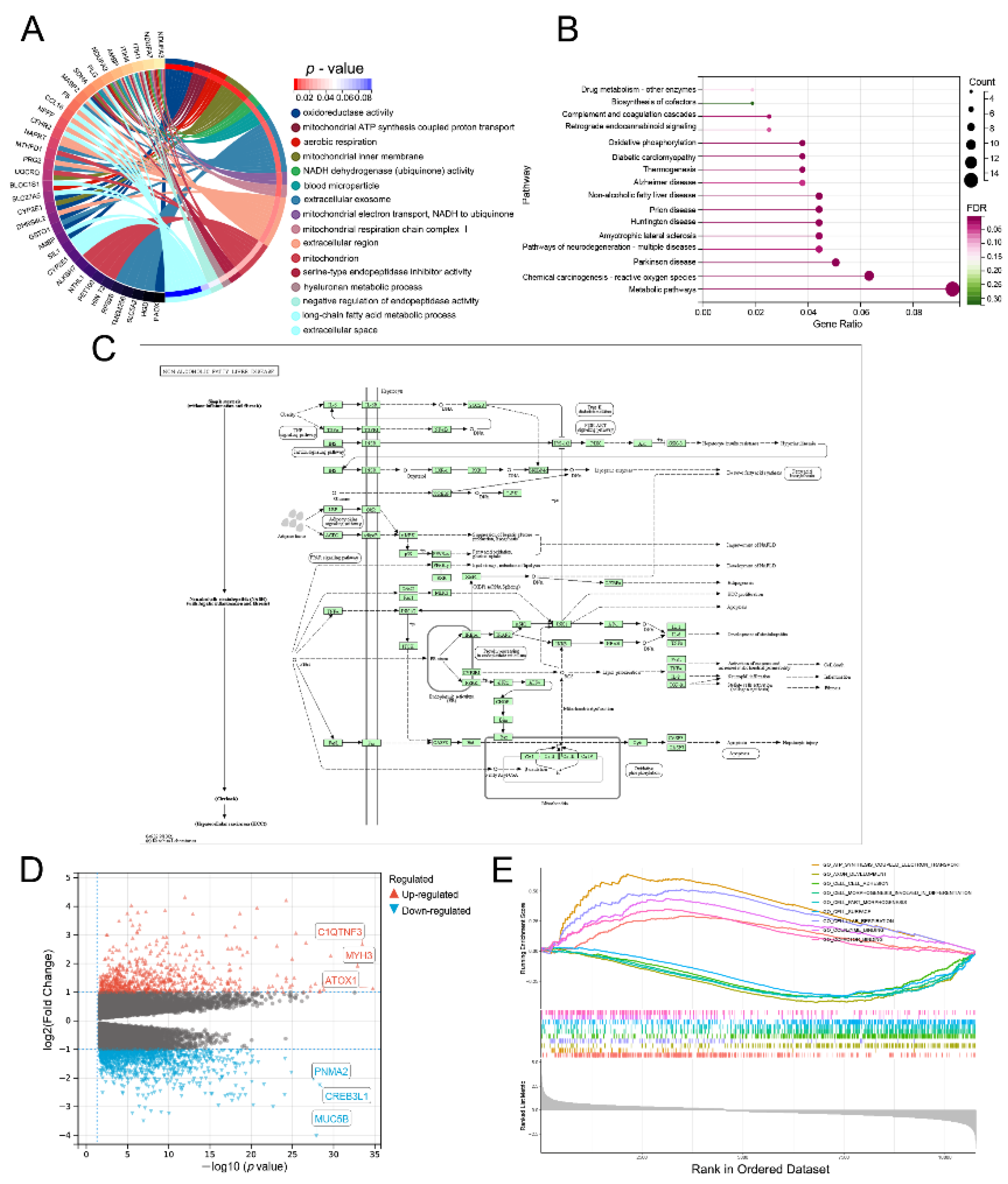
3.8. Lnc-RAB11B-AS1-Related Genes and Functional Enrichment Analysis
3.9. Lnc-RAB11B-AS1-Related Signaling Pathways Obtained by GSEA
3.10. Correlation Analysis between lnc-RAB11B-AS1 and RAB11B Expression, and Tumor-Infiltrating Immune Cells in HCC
3.11. Alternation of lnc-RAB11B-AS1 and RAB11B in HCC
3.12. Correlation between mRNA Expression and Methylation of lnc-RAB11B-AS1
3.13. Prediction of lnc-RAB11B-AS1 and RAB11B Targeted miRNAs
3.14. Correlation between lnc-RAB11B-AS1 Expression and Drug Sensitivity
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Cheng, J.; Wei, D.; Ji, Y.; Chen, L.; Yang, L.; Li, G.; Wu, L.; Hou, T.; Xie, L.; Ding, G.; et al. Integrative analysis of DNA methylation and gene expression reveals hepatocellular carcinoma-specific diagnostic biomarkers. Genome Med. 2018, 10, 42. [Google Scholar] [CrossRef] [PubMed]
- Sung, H.; Ferlay, J.; Siegel, R.L.; Laversanne, M.; Soerjomataram, I.; Jemal, A.; Bray, F. Global cancer statistics 2020: Globocan estimates of incidence and mortality worldwide for 36 cancers in 185 countries. CA Cancer J. Clin. 2021, 71, 209–249. [Google Scholar] [CrossRef] [PubMed]
- Xie, D.Y.; Ren, Z.G.; Zhou, J.; Fan, J.; Gao, Q. 2019 chinese clinical guidelines for the management of hepatocellular carcinoma: Updates and insights. Hepatobiliary Surg. Nutr. 2020, 9, 452–463. [Google Scholar] [CrossRef] [PubMed]
- Helmberger, T. Guideline on hepatocellular carcinoma. Radiologe 2022, 62, 197–199. [Google Scholar] [CrossRef] [PubMed]
- Park, I.Y.; Sohn, B.H.; Yu, E.; Suh, D.J.; Chung, Y.H.; Lee, J.H.; Surzycki, S.J.; Lee, Y.I. Aberrant epigenetic modifications in hepatocarcinogenesis induced by hepatitis b virus X protein. Gastroenterology 2007, 132, 1476–1494. [Google Scholar] [CrossRef]
- Chen, Z.; Xie, H.; Hu, M.; Huang, T.; Hu, Y.; Sang, N.; Zhao, Y. Recent progress in treatment of hepatocellular carcinoma. Am. J. Cancer Res. 2020, 10, 2993–3036. [Google Scholar]
- Chowdhary, A.; Satagopam, V.; Schneider, R. Long non-coding RNAs: Mechanisms, experimental, and computational approaches in identification, characterization, and their biomarker potential in cancer. Front. Genet. 2021, 12, 649619. [Google Scholar] [CrossRef]
- Yao, W.; Du, X.; Zhang, J.; Wang, Y.; Wang, M.; Pan, Z.; Li, Q. SMAD4-induced knockdown of the antisense long noncoding Rna BRE-AS contributes to granulosa cell apoptosis. Mol. Ther. Nucleic Acids 2021, 25, 251–263. [Google Scholar] [CrossRef]
- Yang, T.; Chen, W.C.; Shi, P.C.; Liu, M.R.; Jiang, T.; Song, H.; Wang, J.Q.; Fan, R.Z.; Pei, D.S.; Song, J. Long noncoding RNA MAPKAPK5-as1 promotes colorectal cancer progression by cis-regulating the nearby gene MK5 and acting as a let-7f-1-3p sponge. J. Exp. Clin. Cancer Res. CR 2020, 39, 139. [Google Scholar] [CrossRef]
- Pfeffer, S.R. Rab gtpases: Master regulators that establish the secretory and endocytic pathways. Mol. Biol. Cell 2017, 28, 712–715. [Google Scholar] [CrossRef]
- Homma, Y.; Hiragi, S.; Fukuda, M. Rab family of small gtpases: An updated view on their regulation and functions. FEBS J. 2021, 288, 36–55. [Google Scholar] [CrossRef] [PubMed]
- Bhuin, T.; Roy, J.K. Rab11 in disease progression. Int. J. Mol. Cell. Med. 2015, 4, 1–8. [Google Scholar] [PubMed]
- Li, T.; Wu, D.; Liu, Q.; Chen, J.; Zhao, H.; Zhang, L.; Xie, C.; Zhu, W.; Chen, Z.; Zhou, Y. Upregulation of long noncoding RNA RAB11B-AS1 promotes tumor metastasis and predicts poor prognosis in lung cancer. Ann. Transl. Med. 2020, 8, 582. [Google Scholar] [CrossRef] [PubMed]
- Dong, Q.; Fu, L.; Zhao, Y.; Du, Y.; Li, Q.; Qiu, X.; Wang, E. RAB11A promotes proliferation and invasion through regulation of yap in non-small cell lung cancer. Oncotarget 2017, 8, 27800–27811. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dong, W.; Qin, G.; Shen, R. RAB11-FIP2 promotes the metastasis of gastric cancer cells. Int. J. Cancer 2016, 138, 1680–1688. [Google Scholar] [CrossRef] [Green Version]
- Li, Y.; Jia, Q.; Zhang, Q.; Wan, Y. Rab25 upregulation correlates with the proliferation, migration, and invasion of renal cell carcinoma. Biochem. Biophys. Res. Commun. 2015, 458, 745–750. [Google Scholar] [CrossRef]
- Chen, Z.; Liu, Z.; Yang, Y.; Zhu, Z.; Liang, R.; Huang, B.; Wu, D.; Yang, L.; Lu, H.; Jin, D.; et al. Long non-coding RNA RAB11B-AS1 prevents osteosarcoma development and progression via its natural antisense transcript rab11b. Oncotarget 2018, 9, 26770–26786. [Google Scholar] [CrossRef] [Green Version]
- Gao, G.F.; Parker, J.S.; Reynolds, S.M.; Silva, T.C.; Wang, L.B.; Zhou, W.; Akbani, R.; Bailey, M.; Balu, S.; Berman, B.P.; et al. Before and after: Comparison of legacy and harmonized tcga genomic data commons’ data. Cell Syst. 2019, 9, 24–34.e10. [Google Scholar] [CrossRef] [Green Version]
- Barrett, T.; Wilhite, S.E.; Ledoux, P.; Evangelista, C.; Kim, I.F.; Tomashevsky, M.; Marshall, K.A.; Phillippy, K.H.; Sherman, P.M.; Holko, M.; et al. Ncbi geo: Archive for functional genomics data sets--update. Nucleic Acids Res. 2013, 41, D991–D995. [Google Scholar] [CrossRef] [Green Version]
- Carrieri, C.; Cimatti, L.; Biagioli, M.; Beugnet, A.; Zucchelli, S.; Fedele, S.; Pesce, E.; Ferrer, I.; Collavin, L.; Santoro, C.; et al. Long non-coding antisense rna controls uchl1 translation through an embedded SINEB2 repeat. Nature 2012, 491, 454–457. [Google Scholar] [CrossRef] [Green Version]
- Gupta, R.A.; Shah, N.; Wang, K.C.; Kim, J.; Horlings, H.M.; Wong, D.J.; Tsai, M.C.; Hung, T.; Argani, P.; Rinn, J.L.; et al. Long non-coding RNA hotair reprograms chromatin state to promote cancer metastasis. Nature 2010, 464, 1071–1076. [Google Scholar] [CrossRef]
- Warde-Farley, D.; Donaldson, S.L.; Comes, O.; Zuberi, K.; Badrawi, R.; Chao, P.; Franz, M.; Grouios, C.; Kazi, F.; Lopes, C.T.; et al. The genemania prediction server: Biological network integration for gene prioritization and predicting gene function. Nucleic Acids Res. 2010, 38, W214–W220. [Google Scholar] [CrossRef] [Green Version]
- Asplund, A.; Edqvist, P.H.; Schwenk, J.M.; Pontén, F. Antibodies for profiling the human proteome-the human protein atlas as a resource for cancer research. Proteomics 2012, 12, 2067–2077. [Google Scholar] [CrossRef]
- Sherman, B.T.; Hao, M.; Qiu, J.; Jiao, X.; Baseler, M.W.; Lane, H.C.; Imamichi, T.; Chang, W. David: A web server for functional enrichment analysis and functional annotation of gene lists (2021 update). Nucleic Acids Res. 2022, 50, W216–W221. [Google Scholar] [CrossRef]
- Tang, Z.; Li, C.; Kang, B.; Gao, G.; Li, C.; Zhang, Z. Gepia: A web server for cancer and normal gene expression profiling and interactive analyses. Nucleic Acids Res. 2017, 45, W98–W102. [Google Scholar] [CrossRef] [Green Version]
- Li, T.; Fan, J.; Wang, B.; Traugh, N.; Chen, Q.; Liu, J.S.; Li, B.; Liu, X.S. Timer: A web server for comprehensive analysis of tumor-infiltrating immune cells. Cancer Res. 2017, 77, e108–e110. [Google Scholar] [CrossRef] [Green Version]
- Gao, J.; Aksoy, B.A.; Dogrusoz, U.; Dresdner, G.; Gross, B.; Sumer, S.O.; Sun, Y.; Jacobsen, A.; Sinha, R.; Larsson, E. Integrative analysis of complex cancer genomics and clinical profiles using the cbioportal. Sci. Signal. 2013, 6, l1. [Google Scholar] [CrossRef] [Green Version]
- Goldman, M.J.; Craft, B.; Hastie, M.; Repečka, K.; McDade, F.; Kamath, A.; Banerjee, A.; Luo, Y.; Rogers, D.; Brooks, A.N. Visualizing and interpreting cancer genomics data via the xena platform. Nat. Biotechnol. 2020, 38, 675–678. [Google Scholar] [CrossRef]
- Reinhold, W.C.; Sunshine, M.; Liu, H.; Varma, S.; Kohn, K.W.; Morris, J.; Doroshow, J.; Pommier, Y. Cellminer: A web-based suite of genomic and pharmacologic tools to explore transcript and drug patterns in the NCI-60 cell line set. Cancer Res. 2012, 72, 3499–3511. [Google Scholar] [CrossRef] [Green Version]
- Sticht, C.; De La Torre, C.; Parveen, A.; Gretz, N. Mirwalk: An online resource for prediction of microRNA binding sites. PLoS ONE 2018, 13, e0206239. [Google Scholar] [CrossRef] [PubMed]
- Li, J.H.; Liu, S.; Zhou, H.; Qu, L.H.; Yang, J.H. Starbase v2.0: Decoding miRNA-ceRNA, miRNA-ncRNA and protein-RNA interaction networks from large-scale clip-seq data. Nucleic Acids Res. 2014, 42, D92–D97. [Google Scholar] [CrossRef] [PubMed]
- Bertucci, F.; Chaffanet, M.; Birnbaum, D. An ICGC major achievement in breast cancer: A comprehensive catalog of mutations and mutational signatures. Chin. Clin. Oncol. 2017, 6, 4. [Google Scholar] [CrossRef] [PubMed]
- Győrffy, B.; Benke, Z.; Lánczky, A.; Balázs, B.; Szállási, Z.; Timár, J.; Schäfer, R. Recurrenceonline: An online analysis tool to determine breast cancer recurrence and hormone receptor status using microarray data. Breast Cancer Res. Treat. 2012, 132, 1025–1034. [Google Scholar] [CrossRef] [Green Version]
- Cao, Z.; Pan, X.; Yang, Y.; Huang, Y.; Shen, H.B. The lnclocator: A subcellular localization predictor for long non-coding RNAs based on a stacked ensemble classifier. Bioinformatics 2018, 34, 2185–2194. [Google Scholar] [CrossRef] [PubMed]
- Morris, K.V.; Vogt, P.K. Long antisense non-coding rnas and their role in transcription and oncogenesis. Cell Cycle 2010, 9, 2544–2547. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Y.T.; Li, B.P.; Zhang, B.; Ma, P.; Wu, Q.L.; Ming, L.; Xie, L.M. Lncrna sbf2-as1 promotes hepatocellular carcinoma metastasis by regulating emt and predicts unfavorable prognosis. Eur. Rev. Med. Pharmacol. Sci. 2018, 22, 6333–6341. [Google Scholar]
- Takahashi, K.; Yan, I.K.; Kogure, T.; Haga, H.; Patel, T. Extracellular vesicle-mediated transfer of long non-coding RNA ROR modulates chemosensitivity in human hepatocellular cancer. FEBS Open Bio. 2014, 4, 458–467. [Google Scholar] [CrossRef] [Green Version]
- Rothzerg, E.; Ho, X.D.; Xu, J.; Wood, D.; Märtson, A.; Kõks, S. Upregulation of 15 antisense long non-coding RNAs in osteosarcoma. Genes 2021, 12, 1132. [Google Scholar] [CrossRef]
- Niu, Y.; Bao, L.; Chen, Y.; Wang, C.; Luo, M.; Zhang, B.; Zhou, M.; Wang, J.E.; Fang, Y.V.; Kumar, A. HIF2-induced long noncoding RNA RAB11B-AS1 promotes hypoxia-mediated angiogenesis and breast cancer metastasis. Cancer Res. 2020, 80, 964–975. [Google Scholar] [CrossRef]
- Ningbo, Y.; Xiaodong, J.; Yao, K. LncRNA RAB11B-AS1 up-regulating miR-628-3p inhibits proliferation and migration of gastric cancer cell line. Basic Clin. Med. 2021, 41, 1762–1766. (In Chinese) [Google Scholar]
- Jiang, Y.; Chen, J.; Ling, J.; Zhu, X.; Jiang, P.; Tang, X.; Zhou, H.; Li, R. Construction of a glycolysis-related long noncoding RNA signature for predicting survival in endometrial cancer. J. Cancer 2021, 12, 1431–1444. [Google Scholar] [CrossRef]
- Gilbert, A.; Williams, C.; Azuero, A.; Burkard, M.E.; Kenzik, K.; Garrett, M.E.; Meersman, S.; Rocque, G. Utilizing data visualization to identify survival and treatment differences between women with de novo and recurrent metastatic breast cancer. Clin. Breast Cancer 2021, 21, 292–301. [Google Scholar] [CrossRef]
- Kariyama, K.; Nouso, K.; Hiraoka, A.; Wakuta, A.; Oonishi, A.; Kuzuya, T.; Toyoda, H.; Tada, T.; Tsuji, K.; Itobayashi, E. Ez-albi score for predicting hepatocellular carcinoma prognosis. Liver Cancer 2020, 9, 734–743. [Google Scholar] [CrossRef]
- Zhao, Z.; Zhu, Y.; Ni, X.; Lin, J.; Li, H.; Zheng, L.; Zhang, C.; Qi, X.; Huo, H.; Lou, X. Serum GGT/ALT ratio predicts vascular invasion in HBV-related HCC. Cancer Cell Int. 2021, 21, 517. [Google Scholar] [CrossRef]
- Núñez Abad, M.; Calabuig-Fariñas, S.; Lobo de Mena, M.; Torres, M.S.; García, G.C.; García, G.J.; Iranzo, G.C.V.; Camps, H.C. Programmed death-ligand 1 (PD-l1) as immunotherapy biomarker in breast cancer. Cancers 2022, 14, 307. [Google Scholar] [CrossRef]
- Peng, L.; Chen, Y.; Ou, Q.; Wang, X.; Tang, N. LncRNA MIAT correlates with immune infiltrates and drug reactions in hepatocellular carcinoma. Int. Immunopharmacol. 2020, 89, 107071. [Google Scholar] [CrossRef]
- Park, J.G.; Kim, G.; Jang, S.Y.; Lee, Y.R.; Lee, E.; Lee, H.W.; Han, M.H.; Chun, J.M.; Han, Y.S.; Yoon, J.S. Plasma long noncoding RNA LEXIS is a potential diagnostic marker for non-alcoholic steatohepatitis. Life 2020, 10, 230. [Google Scholar] [CrossRef]
- Kong, F.; You, H.; Tang, R.; Zheng, K. The regulation of proteins associated with the cytoskeleton by hepatitis B virus X protein during hepatocarcinogenesis. Oncol. Lett. 2017, 13, 2514–2520. [Google Scholar] [CrossRef] [Green Version]
- Liu, X.; Wu, S.; Yang, Y.; Zhao, M.; Zhu, G.; Hou, Z. The prognostic landscape of tumor-infiltrating immune cell and immunomodulators in lung cancer. Biomed. Pharmacother. 2017, 95, 55–61. [Google Scholar] [CrossRef]
- Hong, W.; Liang, L.; Gu, Y.; Qi, Z.; Qiu, H.; Yang, X.; Zeng, W.; Ma, L.; Xie, J. Immune-related lncrna to construct novel signature and predict the immune landscape of human hepatocellular carcinoma. Mol. Ther. Nucleic Acids 2020, 22, 937–947. [Google Scholar] [CrossRef]
- Luzón-Toro, B.; Fernández, R.M.; Martos-Martínez, J.M.; Rubio, M.M.; Antiñolo, G.; Borrego, S. LncRNA LUCAT1 as a novel prognostic biomarker for patients with papillary thyroid cancer. Sci. Rep. 2019, 9, 14374. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, Z.; Yang, B.; Zhang, M.; Guo, W.; Wu, Z.; Wang, Y.; Jia, L.; Li, S.; Xie, W.; Yang, D. LncRNA epigenetic landscape analysis identifies EPIC1 as an oncogenic lncRNA that interacts with MYC and promotes cell-cycle progression in cancer. Cancer Cell 2018, 33, 706–720.e709. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, J.; Zhao, X.; Ma, X.; Yuan, Z.; Hu, M. KCNQ1OT1 contributes to sorafenib resistance and programmed death-ligand-1-mediated immune escape via sponging miR-506 in hepatocellular carcinoma cells. Int. J. Mol. Med. 2020, 46, 1794–1804. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Yang, L.; Chen, T.; Liu, X.; Guo, Y.; Zhu, Q.; Tong, X.; Yang, W.; Xu, Q.; Huang, D. A novel lncRNA MCM3AP-AS1 promotes the growth of hepatocellular carcinoma by targeting miR-194-5p/FOXA1 axis. Mol. Cancer 2019, 18, 28. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Characteristics | n | Lnc-RAB11B-AS1 Expression | χ2 | p-Value | |
|---|---|---|---|---|---|
| High (n = 184) | Low (n = 184) | ||||
| Sex | 9.908 | 0.002 | |||
| Male | 248 | 139 (56.1) | 109 (43.9) | ||
| Female | 120 | 46 (38.3) | 74 (61.7) | ||
| Age | 2.297 | 0.130 | |||
| ≤60 | 179 | 82 (45.8) | 97 (54.2) | ||
| >60 | 188 | 101 (53.7) | 87 (46.3) | ||
| Missing | 1 | 1 (100.0) | 0 (0) | ||
| Race | 0.603 | 0.740 | |||
| Asian | 158 | 75 (47.5) | 83 (52.5) | ||
| White | 182 | 89 (48.9) | 93 (51.1) | ||
| Others | 28 | 14 (50.0) | 14 (50.0) | ||
| Historical risk factors | 4.225 | 0.238 | |||
| History of hepatitis B or hepatitis C | 101 | 53 (52.5) | 48 (47.5) | ||
| Alcohol consumption | 65 | 25 (38.5) | 40 (61.5) | ||
| Smoking | 8 | 4 (50.0) | 4 (50.0) | ||
| Others | 194 | 102 (52.6) | 92 (47.4) | ||
| Clinical stage | 4.054 | 0.044 | |||
| I, II | 257 | 140 (54.5) | 117 (45.5) | ||
| III, IV | 88 | 37 (42.0) | 51 (58.0) | ||
| Missing | 23 | 7 (30.4) | 16 (69.6) | ||
| T | 2.472 | 0.116 | |||
| T1, T2 | 275 | 144 (52.4) | 131 (47.6) | ||
| T3, T4 | 91 | 39 (42.9) | 52 (57.1) | ||
| Missing | 2 | 1 (50.0) | 1 (50.0) | ||
| N | 4.605 | 0.032 | |||
| N0 | 250 | 121 (48.2) | 129 (51.8) | ||
| N1 | 5 | 0 (0) | 5 (100.0) | ||
| Missing | 113 | 63 (55.8) | 50 (44.2) | ||
| M | 1.03 | 0.310 | |||
| M0 | 265 | 134 (50.6) | 131 (49.4) | ||
| M1 | 4 | 1 (25.0) | 3 (75.0) | ||
| Missing | 99 | 49 (49.5) | 50 (50.5) | ||
| Histologic grade | 4.416 | 0.036 | |||
| G1, G2 | 232 | 125 (53.9) | 107 (46.1) | ||
| G3, G4 | 132 | 56 (42.4) | 76 (57.6) | ||
| Missing | 4 | 3 (75.0) | 1 (25.0) | ||
| AFP (μg/L) | 8.066 | 0.018 | |||
| ≤20 | 147 | 86 (58.5) | 61 (41.5) | ||
| 20 < AFP ≤ 400 | 65 | 28 (43.1) | 37 (56.9) | ||
| >400 | 65 | 26 (40.0) | 39 (60.0) | ||
| Missing | 91 | 44 (48.4) | 47 (51.6) | ||
| T-Bil (μmol/L) | |||||
| Medical reference value | 4 | 4 (100.0) | 0 (0) | 4.500 | 0.034 |
| Abnormal value | 289 | 151 (52.2) | 138 (47.8) | ||
| Missing | 75 | 29 (38.7) | 46 (61.3) | ||
| ALB (g/L) | 3.713 | 0.054 | |||
| Medical reference value | 4 | 4 (100.0) | 0 (0) | ||
| Abnormal value | 291 | 150 (51.5) | 141 (48.5) | ||
| Missing | 73 | 30 (41.1) | 43 (58.9) | ||
| Creatinine (μmol/L) | 0.165 | 0.685 | |||
| Medical reference value | 182 | 97 (53.3) | 85 (46.7) | ||
| Abnormal value | 114 | 58 (50.9) | 56 (49.1) | ||
| Missing | 72 | 29 (40.3) | 43 (59.7) | ||
| Child pugh grade | 0.018 | 0.895 | |||
| A | 216 | 121 (56.0) | 95 (44.0) | ||
| B,C | 22 | 12 (54.5) | 10 (45.5) | ||
| Missing | 130 | 51 (39.2) | 79 (60.8) | ||
| Treatment or therapy | 1.703 | 0.192 | |||
| Yes | 39 | 16 (41.0) | 23 (59.0) | ||
| No | 307 | 160 (52.1) | 147 (47.9) | ||
| Missing | 22 | 8 (36.4) | 14 (63.6) | ||
| Treatment type | 0.003 | 0.959 | |||
| Pharmaceutical Therapy | 185 | 93 (50.1) | 92 (49.9) | ||
| Radiation Therapy | 183 | 92 (50.3) | 91 (49.7) | ||
| Cancer first-degree relatives | 2.247 | 0.134 | |||
| ≤1 | 65 | 42 (64.6) | 23 (35.4) | ||
| >1 | 53 | 27 (50.9) | 26 (49.1) | ||
| Missing | 250 | 115 (46.0) | 135 (54.0) | ||
| BMI | 335 | 168 (50.1) | 167 (49.9) | 0.884 | 0.377 |
| Missing | 33 | 16 (48.5) | 17 (51.5) | ||
| Characteristics | n | RAB11B Expression | χ2 | p-Value | |
|---|---|---|---|---|---|
| High (n = 184) | Low (n = 184) | ||||
| Sex | 0.012 | 0.911 | |||
| Male | 249 | 124 (49.8) | 125 (50.2) | ||
| Female | 119 | 60 (50.4) | 59 (49.6) | ||
| Age | 0.067 | 0.796 | |||
| ≤60 | 176 | 89 (50.6) | 87 (49.4) | ||
| >60 | 191 | 94 (49.2) | 97 (50.8) | ||
| Missing | 1 | 1 (100.0) | 0 (0) | ||
| Race | 0.854 | 0.653 | |||
| Asian | 158 | 75 (47.5) | 83 (52.5) | ||
| White | 181 | 93 (51.4) | 88 (48.6) | ||
| Others | 29 | 16 (55.2) | 13 (44.8) | ||
| Historical risk factors | 2.200 | 0.532 | |||
| History of hepatitis B or hepatitis C | 101 | 51 (50.5) | 50 (49.5) | ||
| Alcohol consumption | 65 | 28 (43.1) | 37 (56.9) | ||
| Smoking | 9 | 4 (44.4) | 5 (55.6) | ||
| Others | 193 | 103 (53.4) | 90 (46.6) | ||
| Clinical stage | 3.426 | 0.064 | |||
| I, II | 256 | 134 (52.3) | 122 (47.7) | ||
| III, IV | 89 | 36 (40.4) | 53 (59.6) | ||
| Missing | 23 | 14 (60.9) | 9 (39.1) | ||
| T | 2.472 | 0.116 | |||
| T1, T2 | 275 | 144 (52.4) | 131 (47.6) | ||
| T3, T4 | 91 | 39 (42.9) | 52 (57.1) | ||
| Missing | 2 | 1 (50.0) | 1 (50.0) | ||
| N | 0.850 | 0.357 | |||
| N0 | 251 | 121 (48.2) | 130 (51.8) | ||
| N1 | 4 | 1 (25.0) | 3 (75.0) | ||
| Missing | 113 | 62 (54.9) | 51 (45.1) | ||
| M | 0.025 | 0.875 | |||
| M0 | 265 | 122 (46.1) | 143 (53.9) | ||
| M1 | 4 | 2 (50.0) | 2 (50.0) | ||
| Missing | 99 | 60 (60.6) | 39 (39.4) | ||
| Histologic grade | 5.920 | 0.015 | |||
| G1, G2 | 233 | 127 (54.3) | 106 (45.7) | ||
| G3, G4 | 131 | 54 (41.2) | 77 (58.8) | ||
| Missing | 4 | 3 (75.0) | 1 (25.0) | ||
| AFP (μg/L) | 0.581 | 0.748 | |||
| ≤20 | 147 | 72 (49.0) | 75 (51.0) | ||
| 20<AFP ≤ 400 | 66 | 32 (48.5) | 34 (51.5) | ||
| >400 | 64 | 34 (53.1) | 30 (46.9) | ||
| Missing | 91 | 46 (50.5) | 45 (49.5) | ||
| T-Bil (μmol/L) | 0.894 | 0.344 | |||
| Medical reference value | 4 | 3 (75.0) | 1 (25.0) | ||
| Abnormal value | 289 | 148 (51.2) | 141 (48.8) | ||
| Missing | 75 | 33 (44.0) | 42 (56.0) | ||
| ALB (g/L) | 0.920 | 0.337 | |||
| Medical reference value | 4 | 3 (75.0) | 1 (25.0) | ||
| Abnormal value | 291 | 148 (50.9) | 143 (49.1) | ||
| Missing | 73 | 33 (45.2) | 40 (54.8) | ||
| Creatinine (μmol/L) | 0.109 | 0.741 | |||
| Medical reference value | 182 | 89 (48.9) | 93 (51.1) | ||
| Abnormal value | 114 | 58 (50.9) | 56 (49.1) | ||
| Missing | 72 | 37 (51.4) | 35 (48.6) | ||
| Child pugh grade | 0.092 | 0.762 | |||
| A | 217 | 111 (51.2) | 106 (48.8) | ||
| B,C | 22 | 12 (54.5) | 10 (45.5) | ||
| Missing | 129 | 61 (47.3) | 68 (52.7) | ||
| Treatment or therapy | 0.205 | 0.651 | |||
| Yes | 40 | 21 (52.5) | 19 (47.5) | ||
| No | 306 | 149 (48.7) | 157 (51.3) | ||
| Missing | 22 | 14 (63.6) | 8 (36.4) | ||
| Treatment type | 0.330 | 0.566 | |||
| Pharmaceutical Therapy | 185 | 95 (51.4) | 90 (48.6) | ||
| Radiation Therapy | 183 | 88 (48.1) | 94 (51.9) | ||
| Cancer first-degree relatives | 1.238 | 0.266 | |||
| ≤1 | 66 | 36 (54.5) | 30 (45.5) | ||
| >1 | 52 | 23 (44.2) | 29 (55.8) | ||
| Missing | 250 | 125 (50.0) | 125 (50.0) | ||
| BMI | 335 | 166 (49.6) | 169 (50.4) | 1.112 | 0.267 |
| Missing | 33 | 18 (54.5) | 15 (45.5) | ||
| Variable | Univariable | Multivariable | |||||
|---|---|---|---|---|---|---|---|
| HR | 95%CI | p | HR | 95%CI | p | ||
| Gender | 1.119 | 0.771–1.624 | 0.556 | 1.192 | 0.610–2.330 | 0.725 | |
| Age | 1.013 | 0.998–1.027 | 0.083 | 1.023 | 0.998–1.051 | 0.076 | |
| BMI | 1.019 | 0.967–1.028 | 0.819 | 1.031 | 0.979–1.086 | 0.377 | |
| Race | 1 | Reference | Reference | ||||
| 2 | 1.301 | 0.879–1.925 | 0.189 | 0.539 | 0.255–1.141 | 0.106 | |
| 3 | 1.526 | 0.647–3.599 | 0.334 | 1.812 | 0.232–14.105 | 0.570 | |
| Clinical stage | 1.312 | 1.011–1.703 | 0.041 | 2.628 | 1.599–4.321 | <0.001 | |
| T | 2.562 | 1.770–3.707 | <0.001 | 20.208 | 0.869–469.5 | 0.061 | |
| N | 1.991 | 1.487–8.144 | 0.038 | 8.846 | 1.013–1.096 | 0.013 | |
| M | 3.907 | 1.225–12.47 | 0.021 | 1.233 | 0.135–11.281 | 0.853 | |
| Histologic grade | 1.060 | 0.726–1.547 | 0.762 | 1.119 | 0.631–1.983 | 0.701 | |
| AFP (μg/L) | 1 | Reference | Reference | ||||
| 2 | 1.352 | 0.621–2.941 | 0.447 | 1.102 | 0.321–2.564 | 0.854 | |
| 3 | 0.924 | 0.385–2.220 | 0.860 | 1.224 | 0.278–2.402 | 0.714 | |
| T-Bil (μmol/L) | 1.356 | 0.185–9.932 | 0.764 | 1.356 | 0.185–9.932 | 0.764 | |
| ALB (g/L) | 1.398 | 0.101–10.240 | 0.742 | 1.284 | 0.175–9.434 | 0.806 | |
| Creatinine (μmol/L) | 1.710 | 1.067–1.374 | 0.031 | 1.744 | 1.297–1.862 | 0.027 | |
| Child pugh grade | 2.205 | 0.296–16.401 | 0.365 | 1.993 | 0.236–16.846 | 0.526 | |
| Treatment type | 1.231 | 0.858–1.766 | 0.260 | 1.734 | 0.979–3.071 | 0.059 | |
| Treatment or therapy | 1.039 | 0.592–1.822 | 0.895 | 1.176 | 0.492–2.807 | 0.716 | |
| Lnc-RAB11B-AS1 | 0.814 | 0.696–0.951 | 0.009 | 0.799 | 0.656–0.972 | 0.025 | |
| RAB11B | 0.651 | 0.467–0.909 | 0.012 | 0.898 | 0.978–1.000 | 0.041 | |
| Characteristics | n | RAB11B Expression | χ2 | p-Value | |
|---|---|---|---|---|---|
| High (n = 25) | Low (n = 65) | ||||
| Sex | 0.339 | 0.560 | |||
| Male | 80 | 23 (28.6) | 57 (71.4) | ||
| Female | 10 | 2 (20.0) | 8 (80.0) | ||
| Age | 0.543 | 0.461 | |||
| ≤60 | 71 | 21 (29.6) | 50 (70.4) | ||
| >60 | 19 | 4 (21.1) | 15 (78.9) | ||
| Pathology grade | 15.691 | <0.001 | |||
| I | 3 | 2 (66.7) | 1 (33.3) | ||
| II | 43 | 19 (44.2) | 24 (55.8) | ||
| III | 44 | 4 (9.1) | 40 (90.9) | ||
| Clinical stage | 2.018 | 0.365 | |||
| 1 | 63 | 20 (31.7) | 43 (68.3) | ||
| 2 | 25 | 5 (20.0) | 20 (80.0) | ||
| 3 | 2 | 0 (0) | 2 (100.0) | ||
| T | 2.225 | 0.329 | |||
| T1 | 63 | 20 (31.7) | 43 (68.3) | ||
| T2 | 24 | 5 (20.8) | 19 (79.2) | ||
| T3 | 3 | 0 (0) | 3 (100.0) | ||
| Recurrence | 1.522 | 0.217 | |||
| Yes | 49 | 11 (22.4) | 38 (77.6) | ||
| No | 41 | 14 (34.1) | 27 (65.9) | ||
| HBsAg | 2.811 | 0.094 | |||
| Positive | 70 | 16 (22.9) | 54 (77.1) | ||
| Negative | 19 | 8 (42.1) | 11(57.9) | ||
| Unknown | 1 | 1 (100.0) | 0 (0) | ||
| HBcAb | 0.018 | 0.894 | |||
| Positive | 80 | 21 (26.3) | 59 (73.7) | ||
| Negative | 7 | 2 (28.6) | 5 (71.4) | ||
| Unknown | 3 | 2 (66.7) | 1 (33.3) | ||
| AntiHCV | 2.815 | 0.093 | |||
| Positive | 1 | 1 (100.0) | 0 (0) | ||
| Negative | 86 | 22 (25.6) | 64 (74.4) | ||
| Unknown | 3 | 2 (66.7) | 1 (33.3) | ||
| T-Bil (μmol/L) | 4.107 | 0.043 | |||
| Medical reference value | 63 | 21 (33.3) | 42 (66.7) | ||
| Abnormal value | 25 | 3 (12.0) | 22 (88.0) | ||
| Unknown | 2 | 1 (50.0) | 1 (50.0) | ||
| ALT (U/L) | 0.224 | 0.636 | |||
| Medical reference value | 52 | 15 (28.8) | 37 (71.2) | ||
| Abnormal value | 37 | 9 (24.3) | 28 (75.7) | ||
| Unknown | 1 | 1 (100.0) | 0 (0) | ||
| AFP (μg/L) | 0.716 | 0.397 | |||
| ≤20 | 36 | 10 (27.8) | 26 (72.2) | ||
| >20 | 53 | 14 (26.4) | 39 (73.6) | ||
| Unknown | 1 | 1 (100.0) | 0 (0) | ||
| GGT(U/L) | 3.903 | 0.048 | |||
| ≤40 | 30 | 12 (40.0) | 18 (60.0) | ||
| >40 | 59 | 12 (20.3) | 47 (79.7) | ||
| Unknown | 1 | 1 (100.0) | 0 (0) | ||
| PD-L1 expression | 9.357 | 0.002 | |||
| Low | 39 | 18 (46.2) | 21 (53.8) | ||
| High | 45 | 7 (15.6) | 38 (84.4) | ||
| Unknown | 6 | 0 (0) | 6 (100.0) | ||
| CTLA4 expression | 0.786 | 0.375 | |||
| Low | 2 | 0 (0) | 2 (100.0) | ||
| High | 81 | 23 (28.4) | 58 (71.6) | ||
| Unknown | 7 | 2 (28.6) | 5 (71.4) | ||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, D.; Hu, X.; Chen, J.; Liang, B.; Zhang, L.; Qin, P.; Wu, D. Bioinformatics Analysis and Validation of the Role of Lnc-RAB11B-AS1 in the Development and Prognosis of Hepatocellular Carcinoma. Cells 2022, 11, 3517. https://doi.org/10.3390/cells11213517
Wang D, Hu X, Chen J, Liang B, Zhang L, Qin P, Wu D. Bioinformatics Analysis and Validation of the Role of Lnc-RAB11B-AS1 in the Development and Prognosis of Hepatocellular Carcinoma. Cells. 2022; 11(21):3517. https://doi.org/10.3390/cells11213517
Chicago/Turabian StyleWang, Dedong, Xiangzhi Hu, Jinbin Chen, Boheng Liang, Lin Zhang, Pengzhe Qin, and Di Wu. 2022. "Bioinformatics Analysis and Validation of the Role of Lnc-RAB11B-AS1 in the Development and Prognosis of Hepatocellular Carcinoma" Cells 11, no. 21: 3517. https://doi.org/10.3390/cells11213517