

The Potential for Placental Activation of PPARγ to Improve the Angiogenic Profile in Preeclampsia


Abstract
:1. Introduction
2. Materials and Methods
2.1. Tissue Collection
2.2. Explant Culture
2.3. Luminex Assay
2.4. Human Umbilical Vein Endothelial Cell Culture and Tube Formation Assay
2.5. Statistical Analysis
3. Results
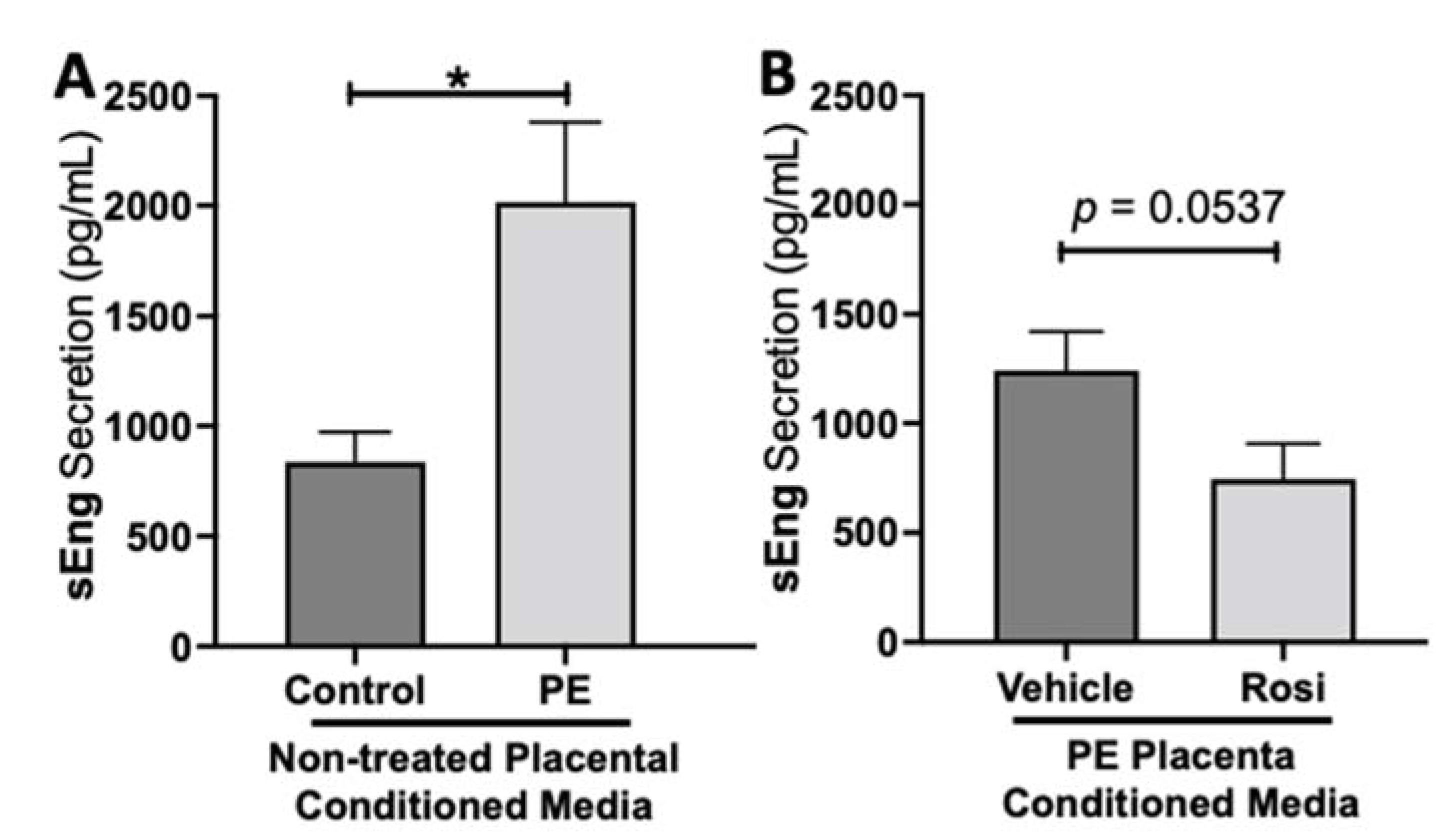
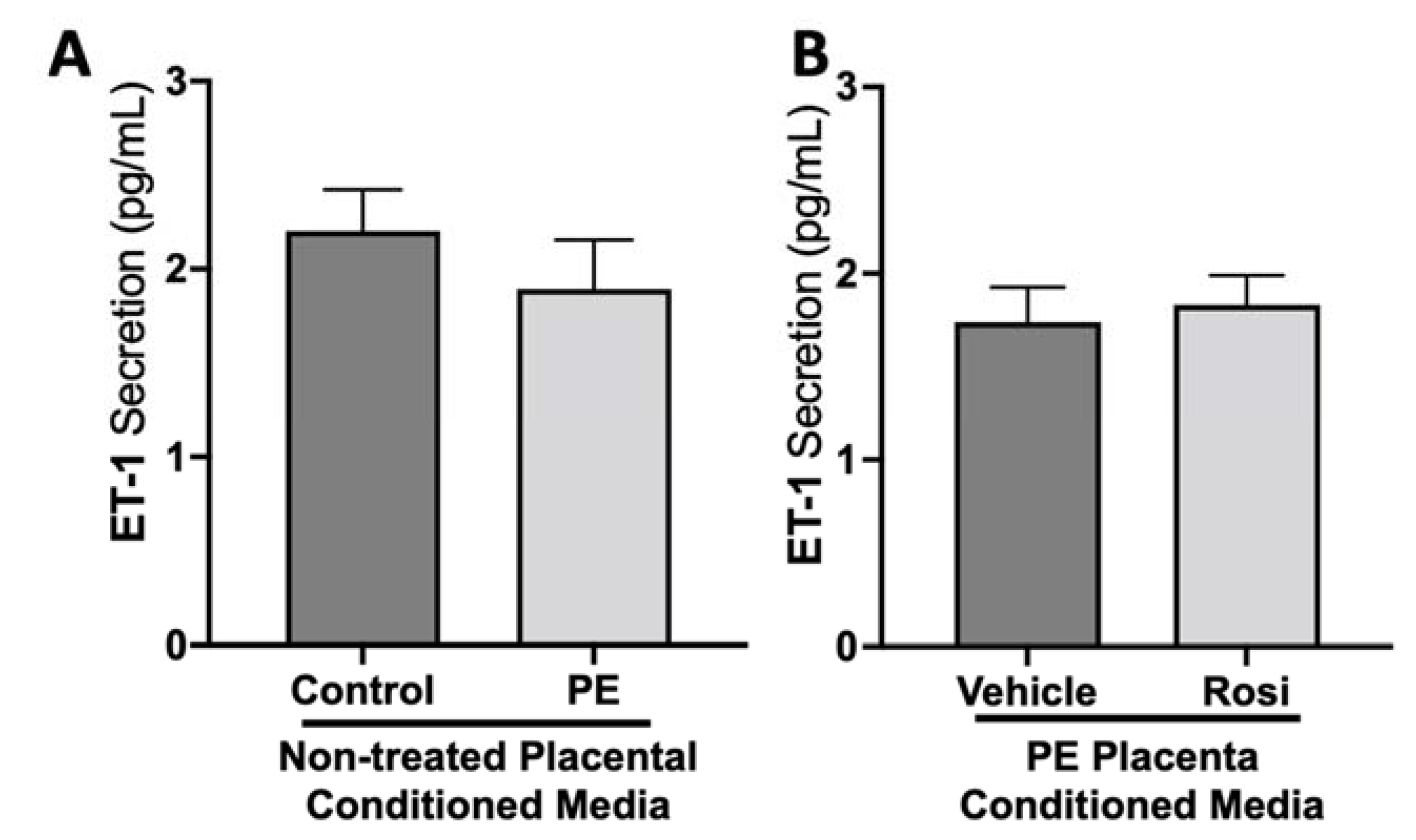
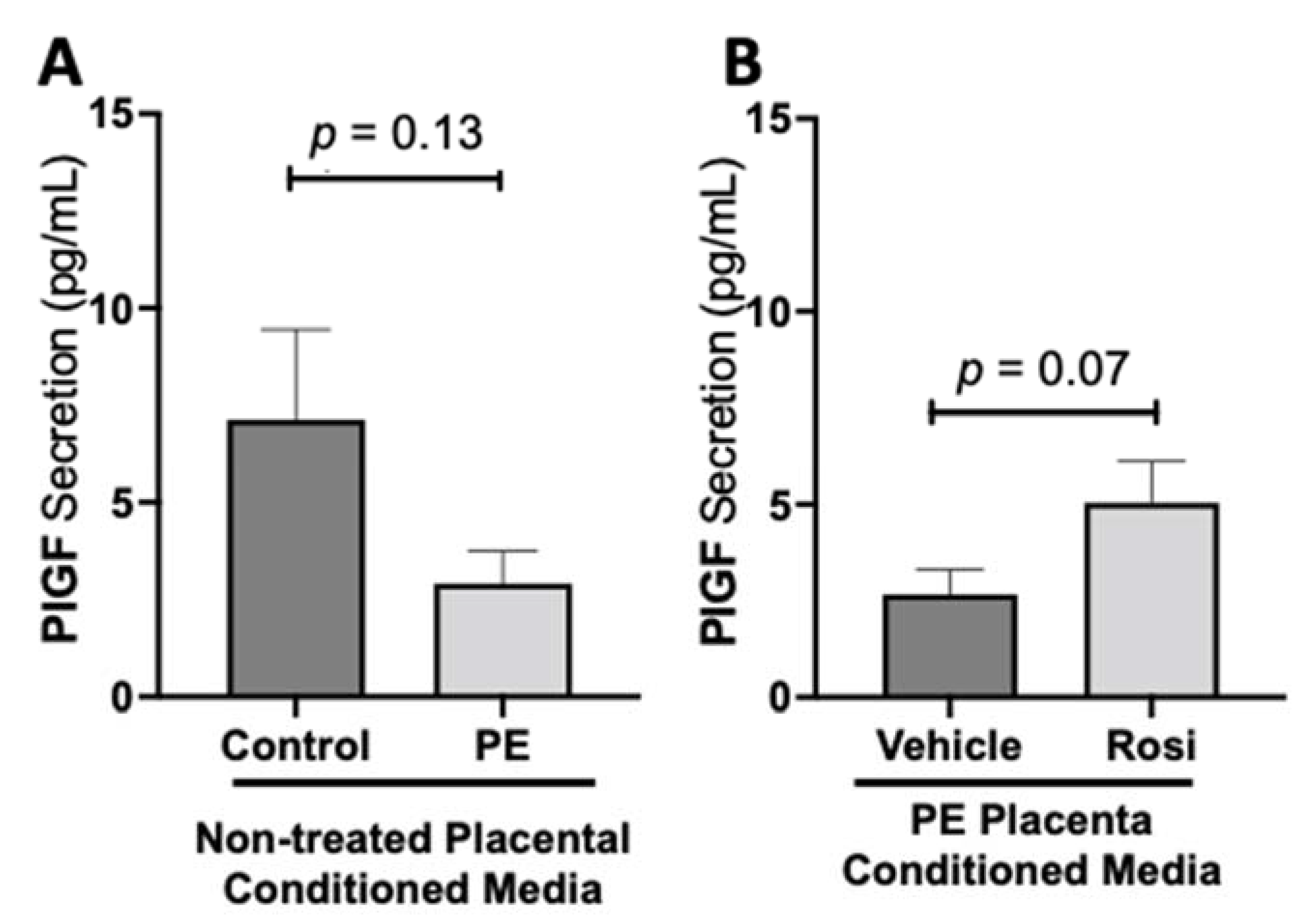
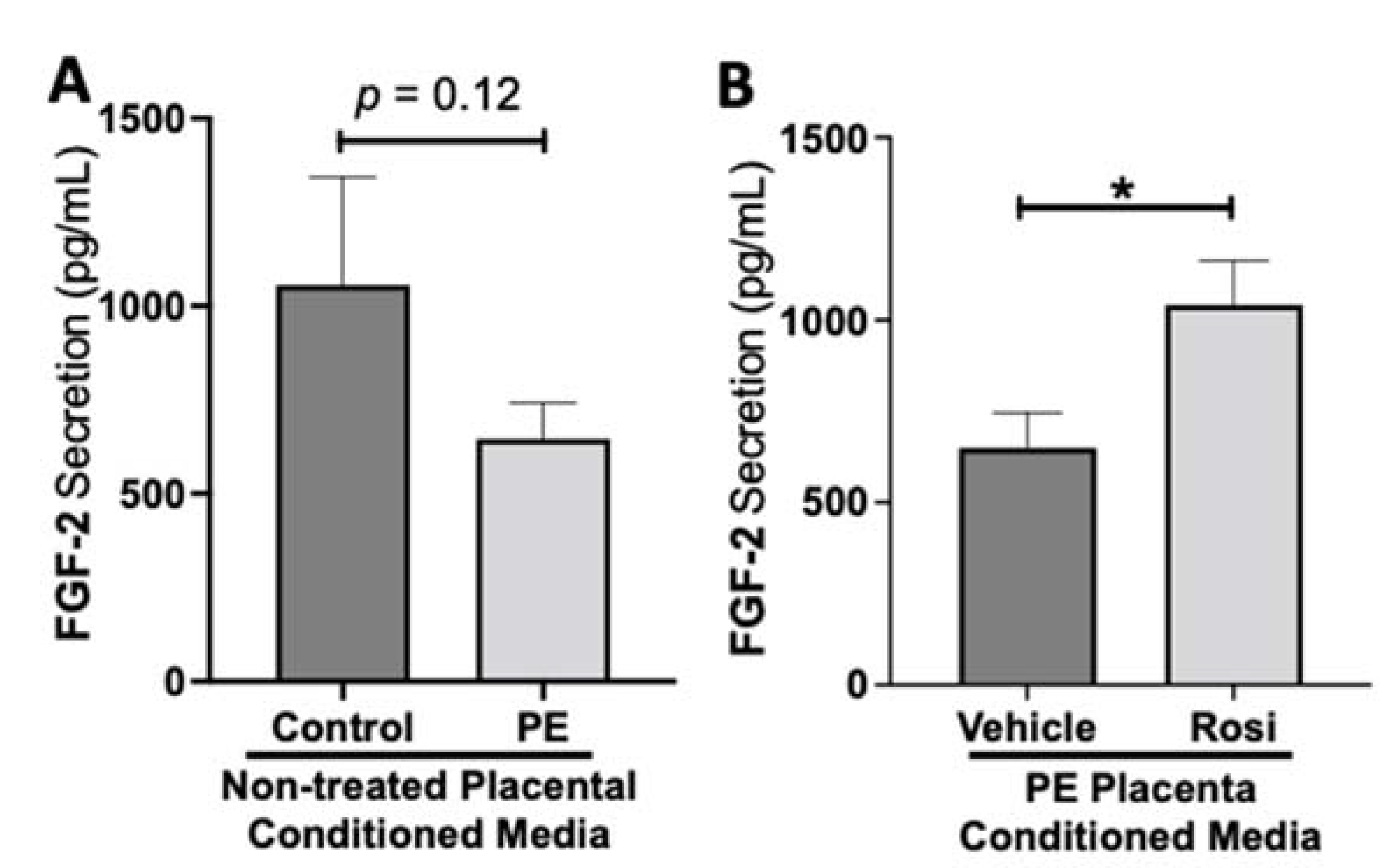
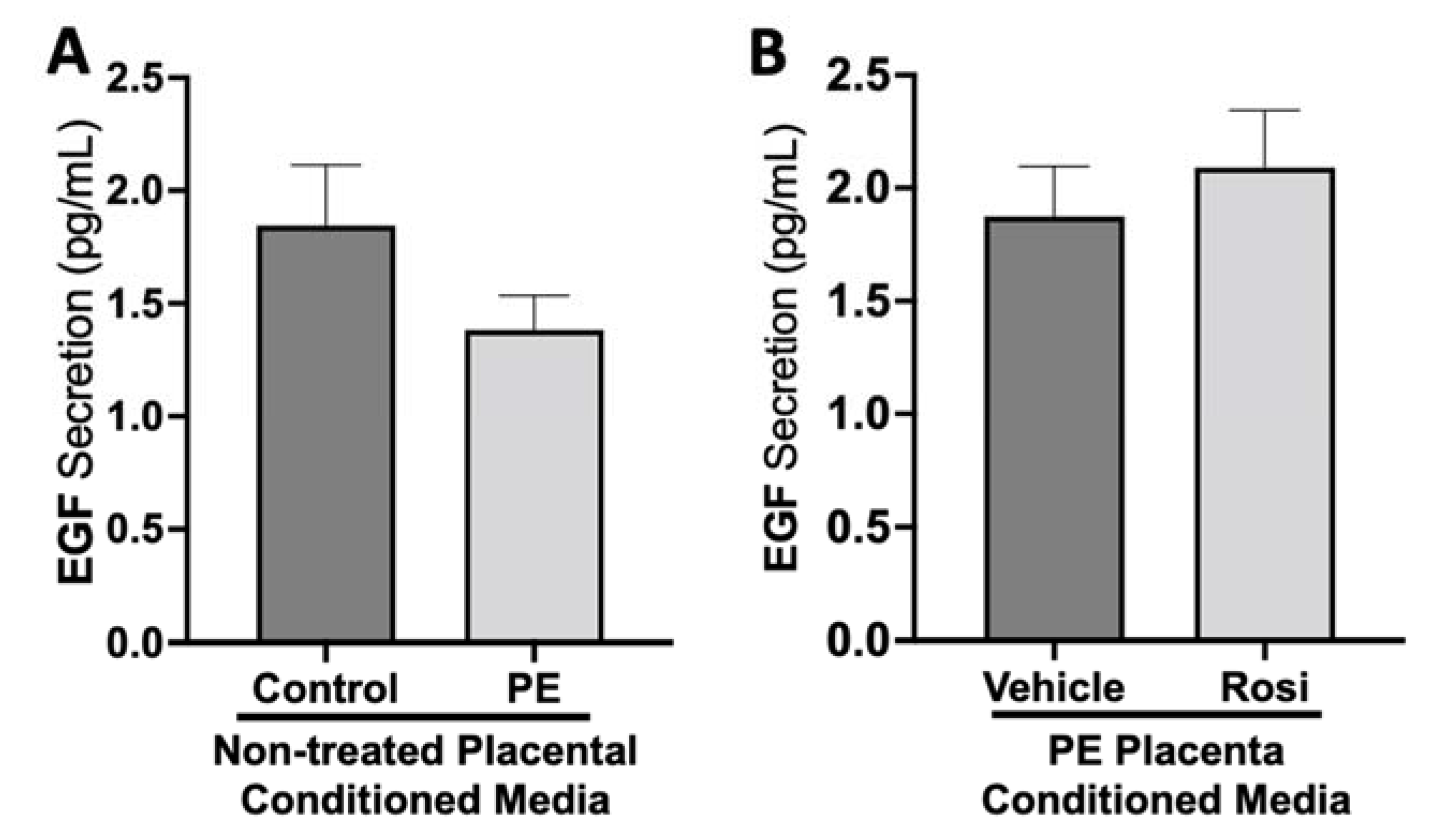
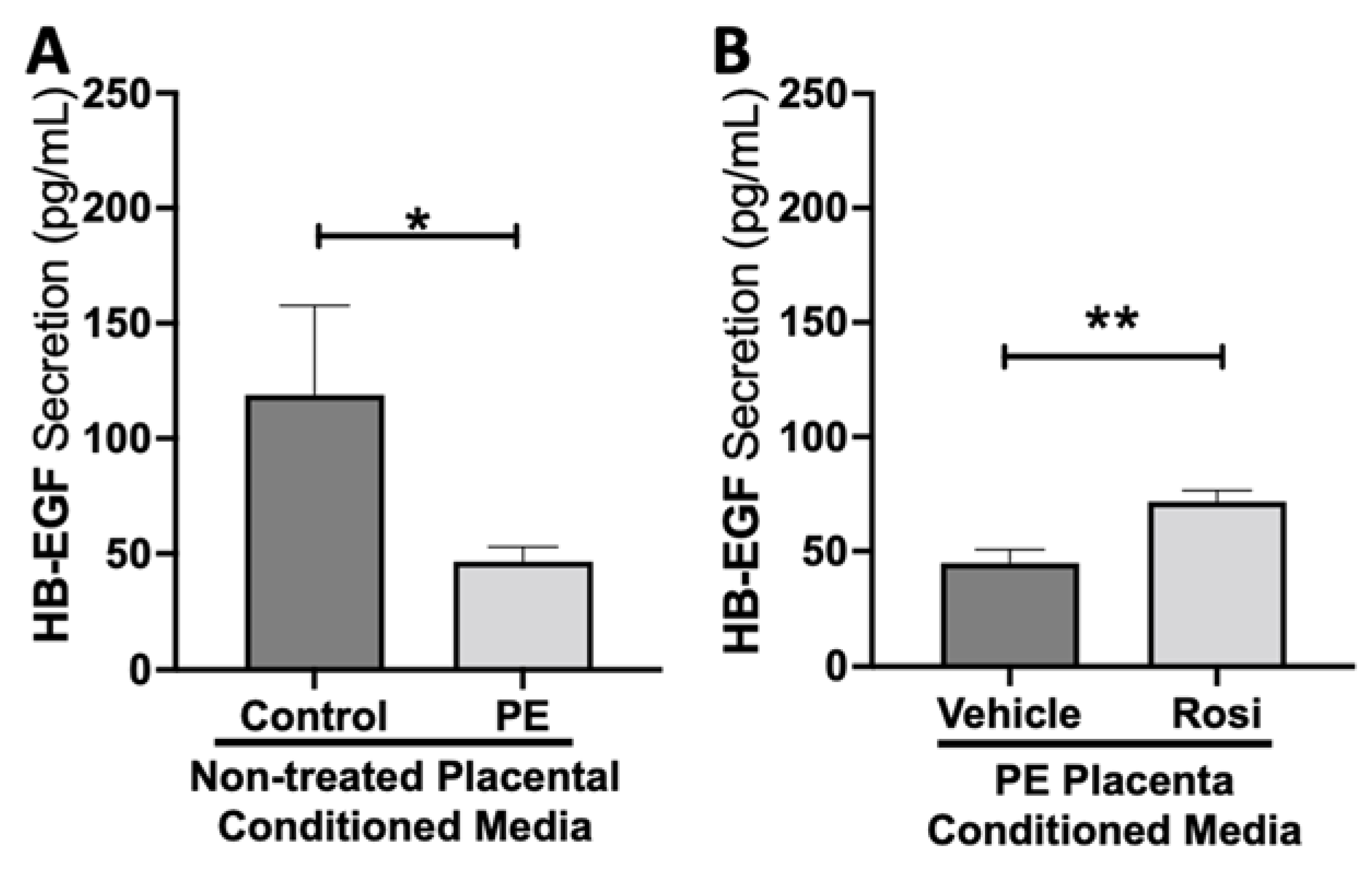
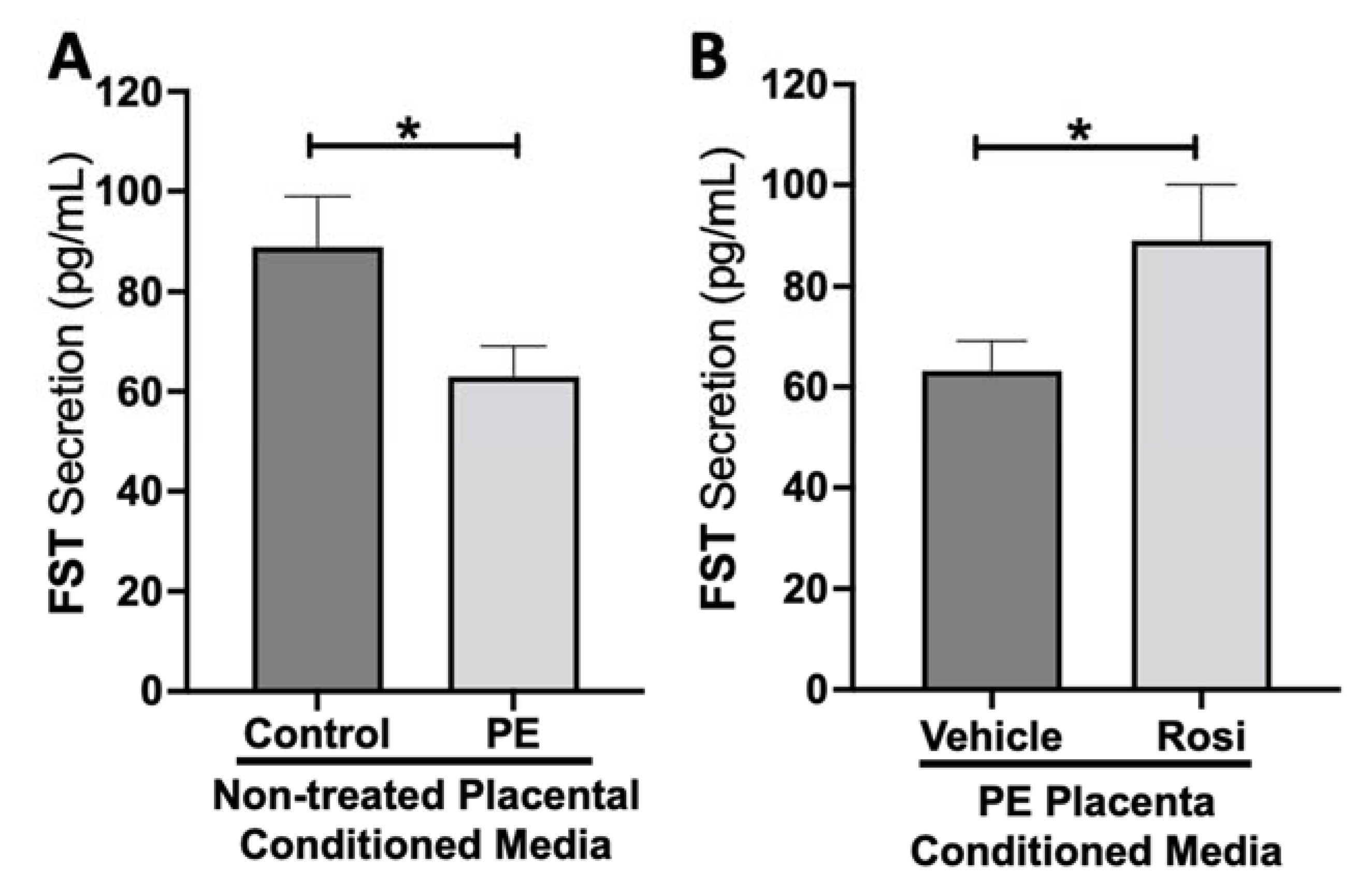
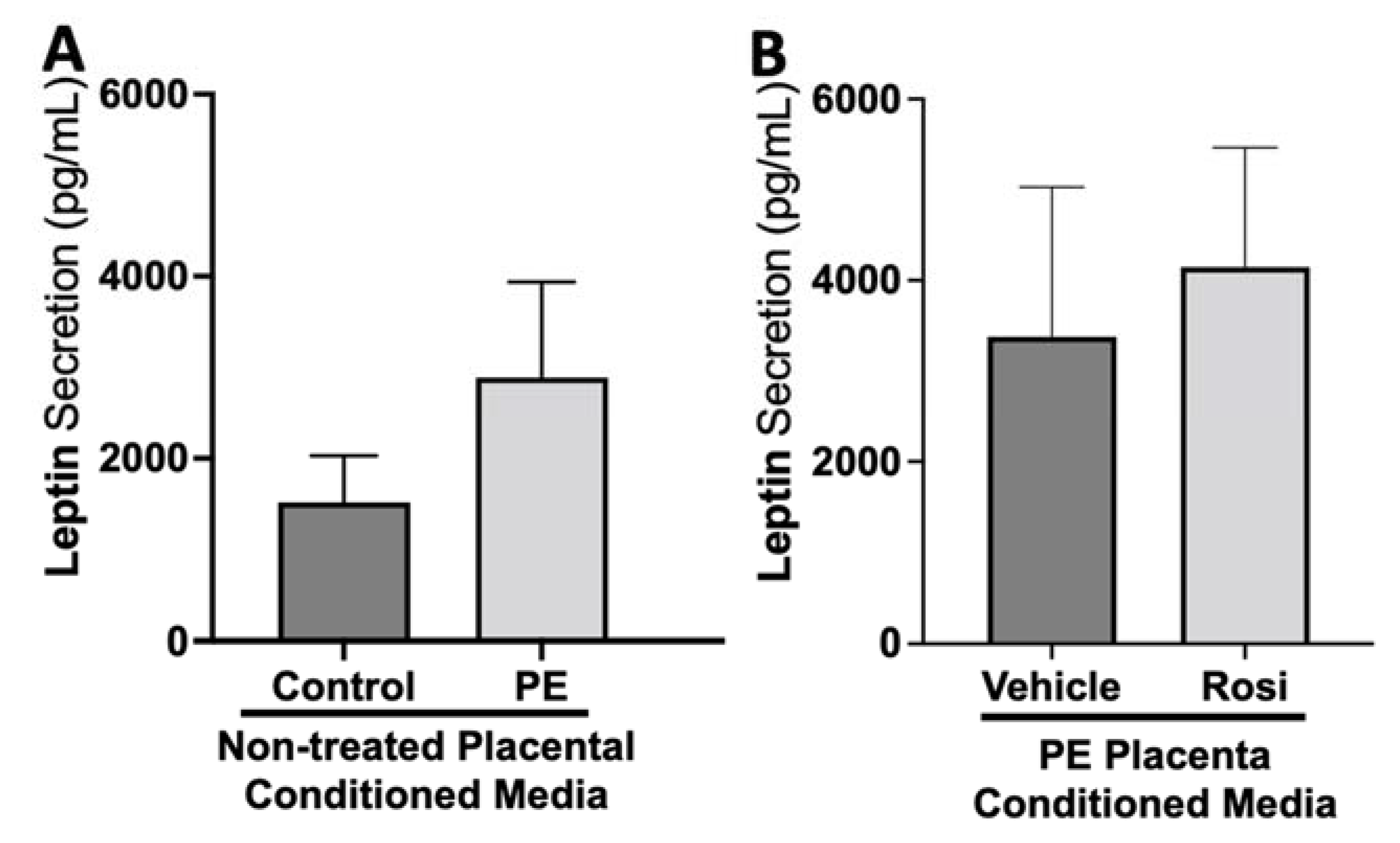
3.1. Rosiglitazone Has a Significant Impact on Angiogenic and Growth Factor Protein Secretion from the Preeclamptic Placenta
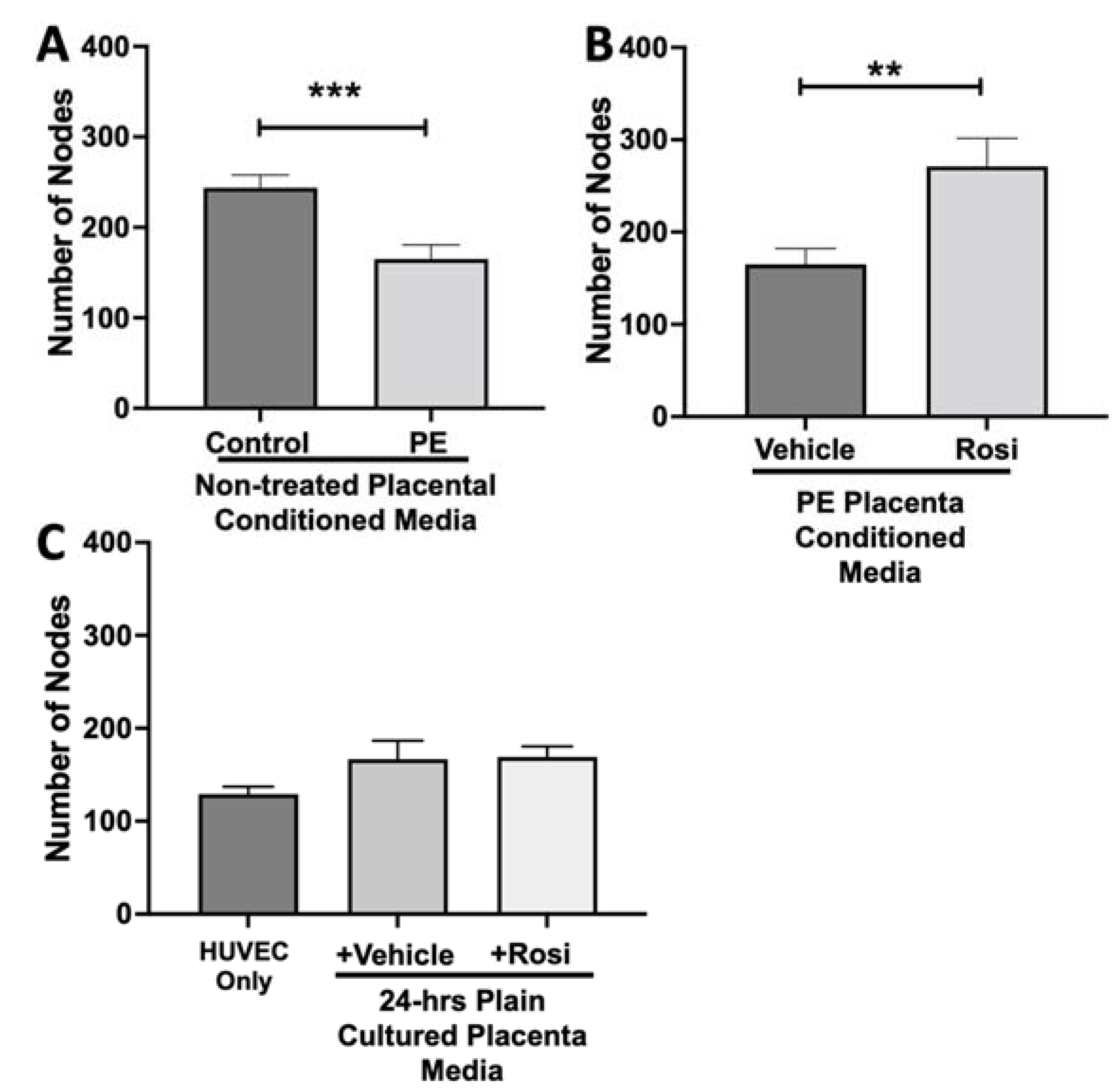
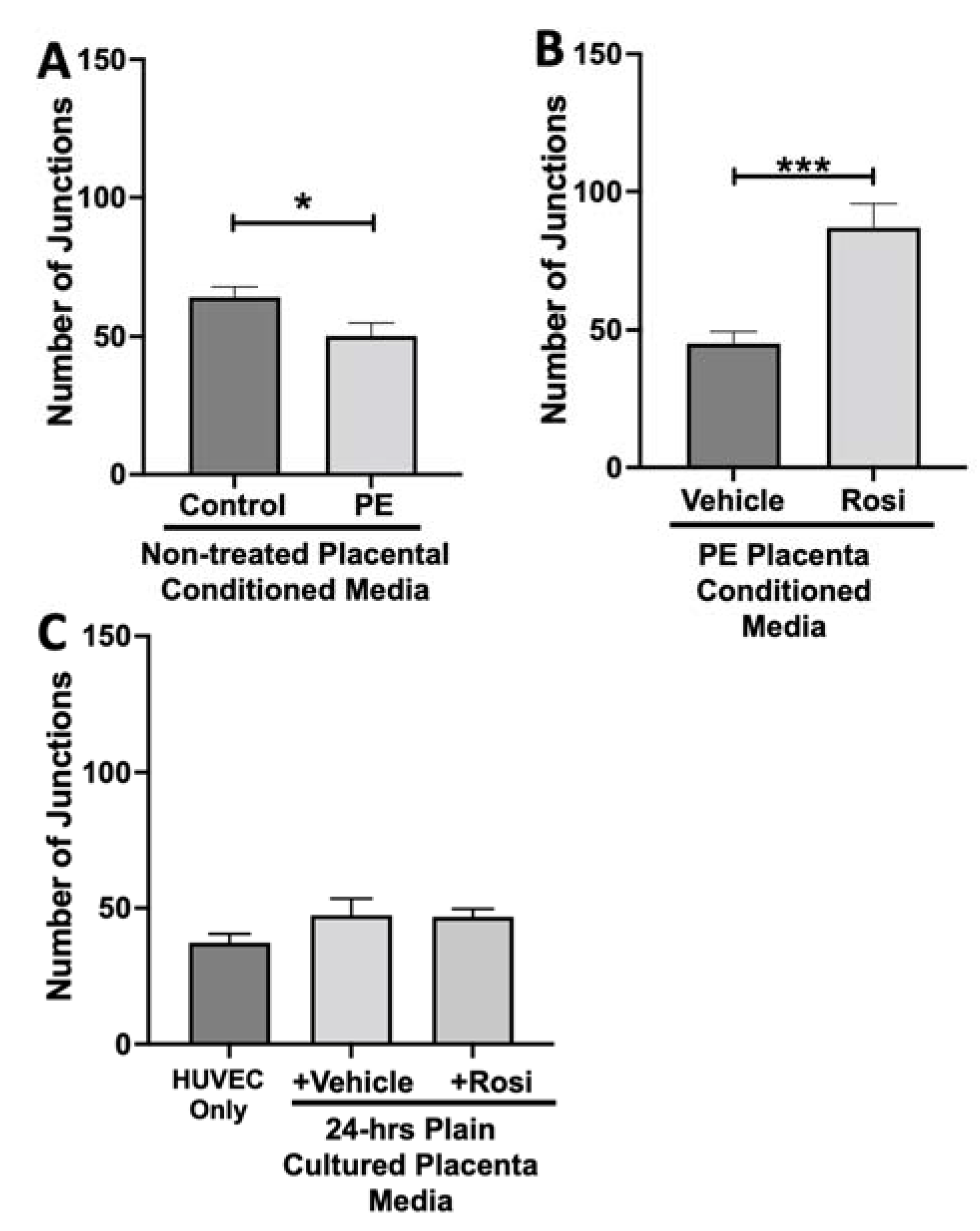
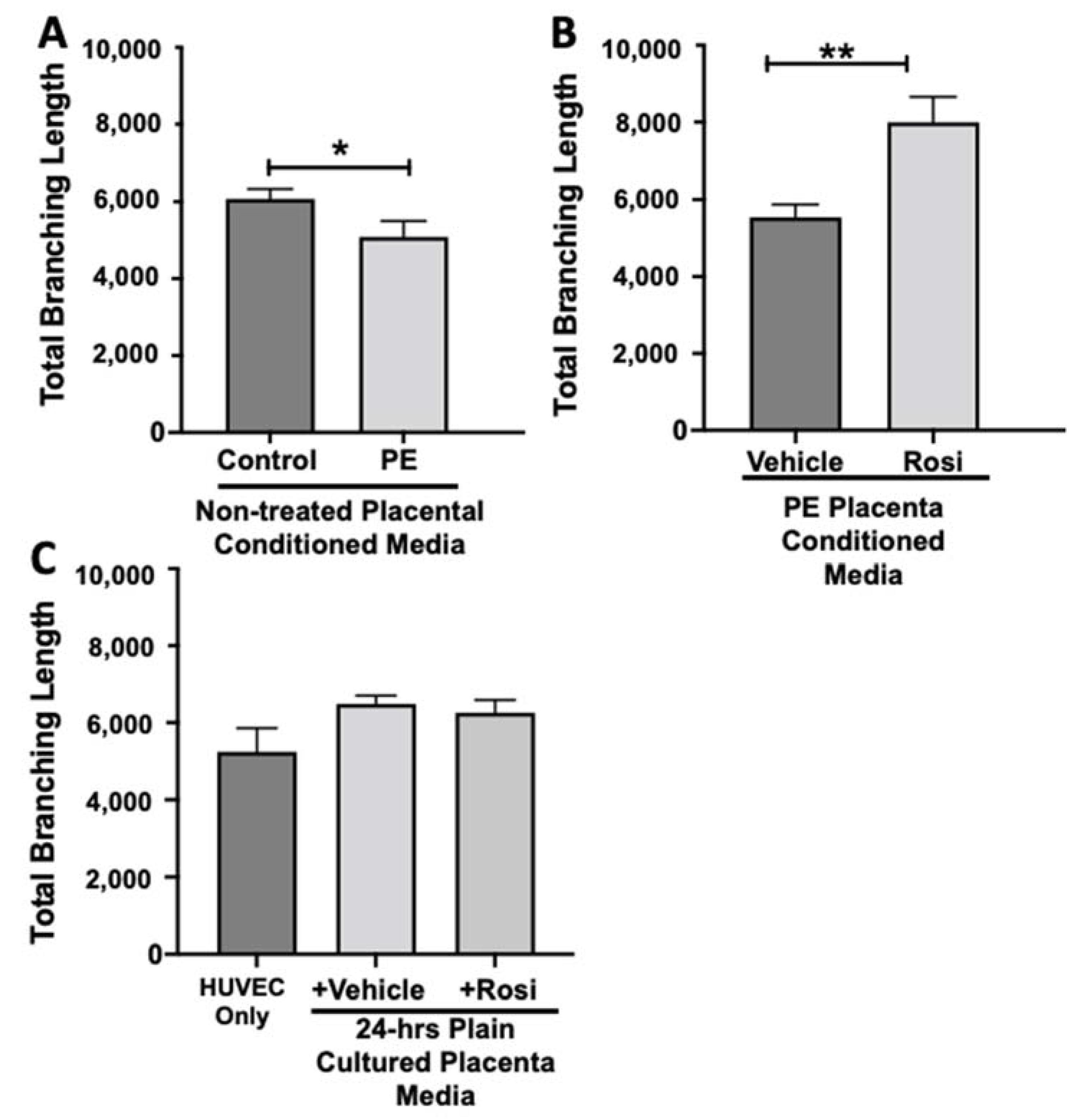
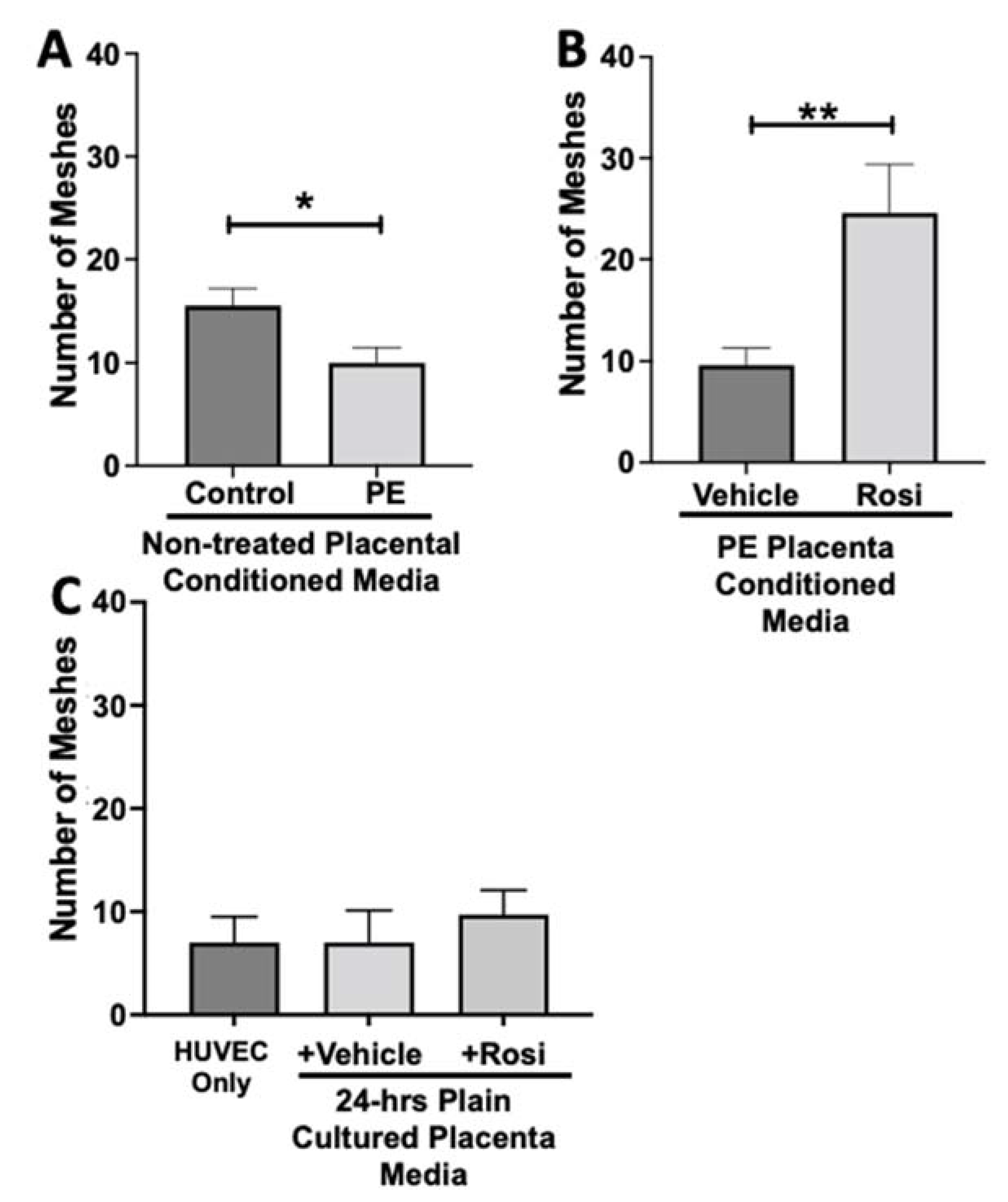
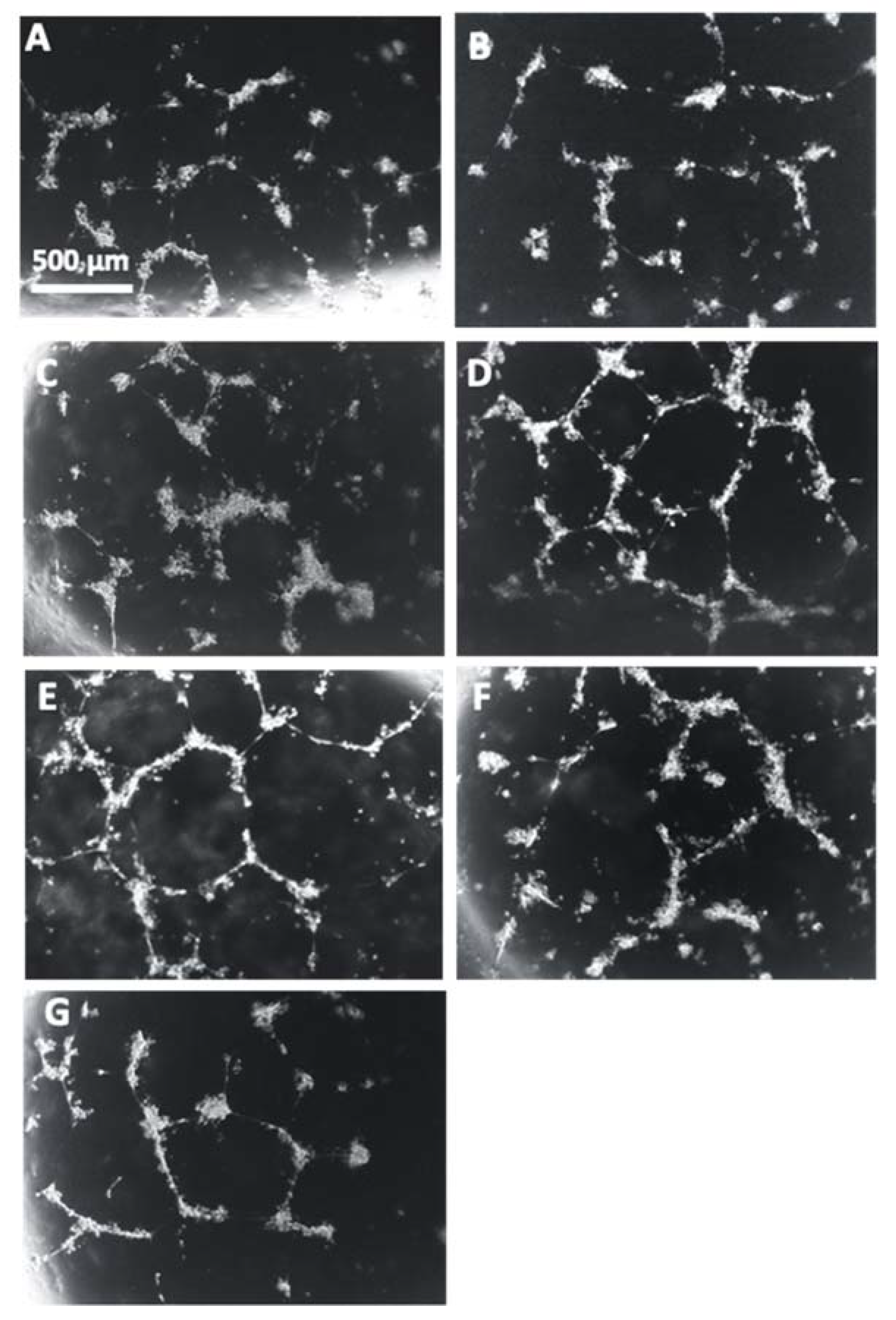
3.2. Tube Formation Assays Reveals a Pro-Angiogenic Effect from Rosiglitazone-Treated Preeclamptic Placentas
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Jeyabalan, A. Epidemiology of preeclampsia: Impact of obesity. Nutr. Rev. 2013, 71, S18–S25. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- ACOG Practice Bulletin No. 202: Gestational Hypertension and Preeclampsia. Obstet. Gynecol. 2019, 133, e1–e25. [CrossRef]
- Toal, M.; Chan, C.; Fallah, S.; Alkazaleh, F.; Chaddha, V.; Windrim, R.C.; Kingdom, J.C. Usefulness of a placental profile in high-risk pregnancies. Am. J. Obstet. Gynecol. 2007, 196, 363.e1–363.e7. [Google Scholar] [CrossRef] [PubMed]
- Sibai, B.M. Evaluation and management of severe preeclampsia before 34 weeks’ gestation. Am. J. Obstet. Gynecol. 2011, 205, 191–198. [Google Scholar] [CrossRef]
- Staff, A.C. The two-stage placental model of preeclampsia: An update. J. Reprod. Immunol. 2019, 134-135, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Fantone, S.; Mazzucchelli, R.; Giannubilo, S.R.; Ciavattini, A.; Marzioni, D.; Tossetta, G. AT-rich interactive domain 1A protein expression in normal and pathological pregnancies complicated by preeclampsia. Histochem. Cell Biol. 2020, 154, 339–346. [Google Scholar] [CrossRef]
- Regnault, T.; Galan, H.; Parker, T.; Anthony, R. Placental Development in Normal and Compromised Pregnancies—A Review. Placenta 2002, 23, S119–S129. [Google Scholar] [CrossRef]
- Phipps, E.A.; Thadhani, R.; Benzing, T.; Karumanchi, S.A. Pre-eclampsia: Pathogenesis, novel diagnostics and therapies. Nat. Rev. Nephrol. 2019, 15, 275–289. [Google Scholar] [CrossRef]
- Pereira, R.D.; De Long, N.E.; Wang, R.C.; Yazdi, F.T.; Holloway, A.C.; Raha, S. Angiogenesis in the Placenta: The Role of Reactive Oxygen Species Signaling. BioMed Res. Int. 2015, 2015, 814543. [Google Scholar] [CrossRef] [Green Version]
- O’Brien, M.; Baczyk, D.; Kingdom, J.C. Endothelial Dysfunction in Severe Preeclampsia is Mediated by Soluble Factors, Rather than Extracellular Vesicles. Sci. Rep. 2017, 7, 5887. [Google Scholar] [CrossRef]
- McCarthy, F.P.; Drewlo, S.; English, F.A.; Kingdom, J.; Johns, E.J.; Kenny, L.C.; Walsh, S.K. Evidence Implicating Peroxisome Proliferator-Activated Receptor-γ in the Pathogenesis of Preeclampsia. Hypertension 2011, 58, 882–887. [Google Scholar] [CrossRef] [Green Version]
- Maynard, S.E.; Karumanchi, S.A. Angiogenic Factors and Preeclampsia. Semin. Nephrol. 2011, 31, 33–46. [Google Scholar] [CrossRef] [Green Version]
- Armistead, B.; Kadam, L.; Drewlo, S.; Kohan-Ghadr, H.-R. The Role of NFκB in Healthy and Preeclamptic Placenta: Trophoblasts in the Spotlight. Int. J. Mol. Sci. 2020, 21, 1775. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Opichka, M.A.; Rappelt, M.W.; Gutterman, D.D.; Grobe, J.L.; McIntosh, J.J. Vascular Dysfunction in Preeclampsia. Cells 2021, 10, 3055. [Google Scholar] [CrossRef]
- Ditisheim, A.; Wuerzner, G.; Ponte, B.; Vial, Y.; Irion, O.; Burnier, M.; Boulvain, M.; Pechère-Bertschi, A. Prevalence of Hypertensive Phenotypes After Preeclampsia: A Prospective Cohort Study. Hypertension 2018, 71, 103–109. [Google Scholar] [CrossRef] [PubMed]
- Goel, A.; Maski, M.R.; Bajracharya, S.; Wenger, J.B.; Zhang, D.; Salahuddin, S.; Shahul, S.S.; Thadhani, R.; Seely, E.W.; Karumanchi, S.A.; et al. Epidemiology and Mechanisms of De Novo and Persistent Hypertension in the Postpartum Period. Circulation 2015, 132, 1726–1733. [Google Scholar] [CrossRef] [Green Version]
- Lindheimer, M.D. Preface. In Chesley’s Hypertensive Disorders in Pregnancy, 4th ed.; Ix-X; Academic Press: Cambridge, MA, USA, 2015. [Google Scholar]
- Wang, S.; Dougherty, E.J.; Danner, R.L. PPARγ signaling and emerging opportunities for improved therapeutics. Pharmacol. Res. 2016, 111, 76–85. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Janani, C.; Ranjitha Kumari, B.D. PPAR gamma gene—A review. Diabetes Metab. Syndr. 2015, 9, 46–50. [Google Scholar] [CrossRef]
- Ahmadian, M.; Suh, J.M.; Hah, N.; Liddle, C.; Atkins, A.R.; Downes, M.; Evans, R.M. PPARγ signaling and metabolism: The good, the bad and the future. Nat. Med. 2013, 19, 557–566. [Google Scholar] [CrossRef] [Green Version]
- Abdelrahman, M.; Sivarajah, A.; Thiemermann, C. Beneficial effects of PPAR-gamma ligands in ischemia-reperfusion injury, inflammation and shock. Cardiovasc. Res. 2005, 65, 772–781. [Google Scholar] [CrossRef] [Green Version]
- Kadam, L.; Kohan-Ghadr, H.R.; Drewlo, S. The balancing act - PPAR-gamma’s roles at the maternal-fetal interface. Syst. Biol. Reprod. Med. 2015, 61, 65–71. [Google Scholar] [CrossRef] [PubMed]
- McCarthy, F.P.; Drewlo, S.; Kingdom, J.; Johns, E.J.; Walsh, S.K.; Kenny, L.C. Peroxisome Proliferator-Activated Receptor-gamma as a Potential Therapeutic Target in the Treatment of Preeclampsia. Hypertension 2011, 58, 280–286. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Levytska, K.; Drewlo, S.; Baczyk, D.; Kingdom, J. PPAR-γ Regulates Trophoblast Differentiation in the BeWo Cell Model. PPAR Res. 2014, 2014, 637251. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Armistead, B.; Kadam, L.; Siegwald, E.; McCarthy, F.P.; Kingdom, J.C.; Kohan-Ghadr, H.-R.; Drewlo, S. Induction of the PPARγ (Peroxisome Proliferator-Activated Receptor γ)-GCM1 (Glial Cell Missing 1) Syncytialization Axis Reduces sFLT1 (Soluble fms-Like Tyrosine Kinase 1) in the Preeclamptic Placenta. Hypertension 2021, 78, 230–240. [Google Scholar] [CrossRef]
- Drewlo, S.; Czikk, M.; Baczyk, D.; Lye, S.; Kingdom, J. Glial cell missing-1 mediates over-expression of tissue inhibitor of metalloproteinase-4 in severe pre-eclamptic placental villi. Hum. Reprod. 2011, 26, 1025–1034. [Google Scholar] [CrossRef] [Green Version]
- Baczyk, D.; Drewlo, S.; Proctor, L.M.; Dunk, C.; Lye, S.; Kingdom, J. Glial cell missing-1 transcription factor is required for the differentiation of the human trophoblast. Cell Death Differ. 2009, 16, 719–727. [Google Scholar] [CrossRef]
- Baczyk, D.; Dunk, C.; Huppertz, B.; Maxwell, C.; Reister, F.; Giannoulias, D.; Kingdom, J. Bi-potential Behaviour of Cytotrophoblasts in First Trimester Chorionic Villi. Placenta 2006, 27, 367–374. [Google Scholar] [CrossRef]
- Baczyk, D.; Satkunaratnam, A.; Nait-Oumesmar, B.; Huppertz, B.; Cross, J.; Kingdom, J. Complex Patterns of GCM1 mRNA and Protein in Villous and Extravillous Trophoblast Cells of the Human Placenta. Placenta 2004, 25, 553–559. [Google Scholar] [CrossRef]
- Carpentier, G.; Berndt, S.; Ferratge, S.; Rasband, W.; Cuendet, M.; Uzan, G.; Albanese, P. Angiogenesis Analyzer for ImageJ—A comparative morphometric analysis of “Endothelial Tube Formation Assay” and “Fibrin Bead Assay”. Sci. Rep. 2020, 10, 11568. [Google Scholar] [CrossRef]
- Wu, C.C.; Chen, Y.C.; Chang, Y.C.; Wang, L.W.; Lin, Y.C.; Chiang, Y.L.; Ho, C.J.; Huang, C.C. Human umbilical vein endothelial cells protect against hypoxic-ischemic damage in neonatal brain via stromal cell-derived factor 1/C-X-C chemokine receptor type 4. Stroke 2013, 44, 1402–1409. [Google Scholar] [CrossRef]
- Wada, Y.; Otu, H.; Wu, S.; Abid, R.; Okada, H.; Libermann, T.; Kodama, T.; Shih, S.; Minami, T.; Aird, W.C. Preconditioning of primary human endothelial cells with inflammatory mediators alters the “set point” of the cell. FASEB J. 2005, 19, 1914–1916. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Labitzke, R.; Friedl, P. A serum-free medium formulation supporting growth of human umbilical cord vein endothelial cells in long-term cultivation. Cytotechnology 2001, 35, 87–92. [Google Scholar] [CrossRef] [PubMed]
- Potapova, I.A.; Gaudette, G.R.; Brink, P.R.; Robinson, R.B.; Rosen, M.R.; Cohen, I.S.; Doronin, S.V. Mesenchymal Stem Cells Support Migration, Extracellular Matrix Invasion, Proliferation, and Survival of Endothelial Cells In Vitro. Stem Cells 2007, 25, 1761–1768. [Google Scholar] [CrossRef] [PubMed]
- Shi, H.; Zuo, Y.; Navaz, S.; Harbaugh, A.; Hoy, C.K.; Gandhi, A.A.; Sule, G.; Yalavarthi, S.; Gockman, K.; Madison, J.A.; et al. Endothelial cell-activating antibodies in COVID-19. medRxiv 2021. [Google Scholar] [CrossRef] [PubMed]
- Angiogenesis Protocols. Third Edition. Anticancer Res. 2016, 36, 4370. [Google Scholar]
- Schneider, C.A.; Rasband, W.S.; Eliceiri, K.W. NIH Image to ImageJ: 25 Years of image analysis. Nat. Methods 2012, 9, 671–675. [Google Scholar] [CrossRef]
- Soto, E.; Romero, R.; Kusanovic, J.P.; Ogge, G.; Hussein, Y.; Yeo, L.; Hassan, S.S.; Kim, C.J.; Chaiworapongsa, T. Late-onset preeclampsia is associated with an imbalance of angiogenic and anti-angiogenic factors in patients with and without placental lesions consistent with maternal underperfusion. J. Matern. Neonatal Med. 2011, 25, 498–507. [Google Scholar] [CrossRef] [Green Version]
- Owaki, Y.; Watanabe, K.; Iwasaki, A.; Saitou, T.; Matsushita, H.; Wakatsuki, A. Placental hypoplasia and maternal organic vascular disorder in pregnant women with gestational hypertension and preeclampsia. J. Matern. Neonatal Med. 2019, 34, 353–359. [Google Scholar] [CrossRef]
- Rana, S.; Lemoine, E.; Granger, J.P.; Karumanchi, S.A. Preeclampsia: Pathophysiology, Challenges, and Perspectives. Circ. Res. 2019, 124, 1094–1112. [Google Scholar] [CrossRef]
- Kohan-Ghadr, H.-R.; A Kilburn, B.; Kadam, L.; Johnson, E.; Kolb, B.L.; Rodriguez-Kovacs, J.; Hertz, M.; Armant, D.R.; Drewlo, S. Rosiglitazone augments antioxidant response in the human trophoblast and prevents apoptosis†. Biol. Reprod. 2018, 100, 479–494. [Google Scholar] [CrossRef]
- Kohan-Ghadr, H.-R.; Kadam, L.; Jain, C.; Armant, D.R.; Drewlo, S. Potential role of epigenetic mechanisms in regulation of trophoblast differentiation, migration, and invasion in the human placenta. Cell Adhes. Migr. 2016, 10, 126–135. [Google Scholar] [CrossRef] [PubMed]
- Kadam, L.; Kilburn, B.; Baczyk, D.; Kohan-Ghadr, H.R.; Kingdom, J.; Drewlo, S. Rosiglitazone blocks first trimester in-vitro placental injury caused by NF-κB-mediated inflammation. Sci. Rep. 2019, 9, 2018. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McCarthy, F.P.; Walsh, S.K.; Drewlo, S.; Kingdom, J.C.; Johns, E.J.; Kenny, L.C. Peroxisome Proliferator Activated Receptor Gamma Critically Regulates the Risk of Preeclampsia in Rodent Gestation. Reprod. Sci. 2011, 18, 285. [Google Scholar]
- Kappou, D.; Sifakis, S.; Konstantinidou, A.; Papantoniou, N.; Spandidos, D. Role of the angiopoietin/Tie system in pregnancy (Review). Exp. Ther. Med. 2015, 9, 1091–1096. [Google Scholar] [CrossRef] [Green Version]
- Akwii, R.G.; Sajib, M.S.; Zahra, F.T.; Mikelis, C.M. Role of Angiopoietin-2 in Vascular Physiology and Pathophysiology. Cells 2019, 8, 471. [Google Scholar] [CrossRef] [Green Version]
- David, S.; Kümpers, P.; Hellpap, J.; Horn, R.; Leitolf, H.; Haller, H.; Kielstein, J.T. Angiopoietin 2 and Cardiovascular Disease in Dialysis and Kidney Transplantation. Am. J. Kidney Dis. 2009, 53, 770–778. [Google Scholar] [CrossRef]
- Hirokoshi, K.; Maeshima, Y.; Kobayashi, K.; Matsuura, E.; Sugiyama, H.; Yamasaki, Y.; Masuyama, H.; Hiramatsu, Y.; Makino, H. Increase of Serum Angiopoietin-2 During Pregnancy Is Suppressed in Women with Preeclampsia. Am. J. Hypertens. 2005, 18, 1181–1188. [Google Scholar] [CrossRef] [Green Version]
- Han, S.Y.; Jun, J.K.; Lee, C.-H.; Park, J.S.; Syn, H.C. Angiopoietin-2: A Promising Indicator for the Occurrence of Severe Preeclampsia. Hypertens. Pregnancy 2010, 31, 189–199. [Google Scholar] [CrossRef]
- Bolin, M.; Wiberg-Itzel, E.; Wikström, A.K.; Goop, M.; Larsson, A.; Olovsson, M.; Akerud, H. Angiopoietin-1/angiopoietin-2 ratio for prediction of preeclampsia. Am. J. Hypertens. 2009, 22, 891–895. [Google Scholar] [CrossRef] [Green Version]
- Myatt, L.; Webster, R.P. Vascular biology of preeclampsia. J. Thromb. Haemost. 2009, 7, 375–384. [Google Scholar] [CrossRef]
- Ahmed, A. New insights into the etiology of preeclampsia: Identification of key elusive factors for the vascular complications. Thromb. Res. 2011, 127, S72–S75. [Google Scholar] [CrossRef]
- Huynh, D.T.N.; Heo, K.-S. Therapeutic targets for endothelial dysfunction in vascular diseases. Arch. Pharm. Res. 2019, 42, 848–861. [Google Scholar] [CrossRef] [PubMed]
- Saleh, L.; Verdonk, K.; Visser, W.; van den Meiracker, A.H.; Danser, A.H.J. The emerging role of endothelin-1 in the pathogenesis of pre-eclampsia. Ther. Adv. Cardiovasc. Dis. 2016, 10, 282–293. [Google Scholar] [CrossRef] [PubMed]
- Marasciulo, M.M.A.M.A.P.F.L.; Montagnani, M.; Potenza, M.A. Endothelin-1: The Yin and Yang on Vascular Function. Curr. Med. Chem. 2006, 13, 1655–1665. [Google Scholar] [CrossRef] [PubMed]
- Lankhorst, S.; Danser, A.H.; van den Meiracker, A.H. Endothelin-1 and antiangiogenesis. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2016, 310, R230–R234. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kang, B.-Y.; Park, K.K.; Kleinhenz, J.M.; Murphy, T.C.; Green, D.E.; Bijli, K.M.; Yeligar, S.M.; Carthan, K.A.; Searles, C.D.; Sutliff, R.L.; et al. Peroxisome Proliferator–Activated Receptor γ and microRNA 98 in Hypoxia-Induced Endothelin-1 Signaling. Am. J. Respir. Cell Mol. Biol. 2016, 54, 136–146. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Delerive, P.; Martin-Nizard, F.; Chinetti, G.; Trottein, F.; Fruchart, J.-C.; Najib, J.; Duriez, P.; Staels, B. Peroxisome proliferator-activated receptor activators inhibit thrombin-induced endothelin-1 production in human vascular endothelial cells by inhibiting the activator protein-1 signaling pathway. Circ. Res. 1999, 85, 394–402. [Google Scholar] [CrossRef] [Green Version]
- Kliche, S.; Waltenberger, J. VEGF Receptor Signaling and Endothelial Function. IUBMB Life 2001, 52, 61–66. [Google Scholar] [CrossRef]
- McLaughlin, K.; Snelgrove, J.W.; Audette, M.C.; Syed, A.; Hobson, S.R.; Windrim, R.C.; Melamed, N.; Carmona, S.; Kingdom, J.C. PlGF (Placental Growth Factor) Testing in Clinical Practice: Evidence from a Canadian Tertiary Maternity Referral Center. Hypertension 2021, 77, 2057–2065. [Google Scholar] [CrossRef]
- Chang, M.; Mukherjea, D.; Gobble, R.M.; Groesch, K.A.; Torry, R.J.; Torry, D.S. Glial Cell Missing 1 Regulates Placental Growth Factor (PGF) Gene Transcription in Human Trophoblast1. Biol. Reprod. 2008, 78, 841–851. [Google Scholar] [CrossRef] [Green Version]
- Li, S.; Roberson, M.S. Dlx3 and GCM-1 functionally coordinate the regulation of placental growth factor in human trophoblast-derived cells. J. Cell Physiol. 2017, 232, 2900–2914. [Google Scholar] [CrossRef]
- Chiu, Y.-H.; Yang, M.-R.; Wang, L.-J.; Chen, M.-H.; Chang, G.-D.; Chen, H. New insights into the regulation of placental growth factor gene expression by the transcription factors GCM1 and DLX3 in human placenta. J. Biol. Chem. 2018, 293, 9801–9811. [Google Scholar] [CrossRef] [PubMed]
- Arany, E.; Hill, D. Fibroblast growth factor-2 and fibroblast growth factor receptor-1 mRNA expression and peptide localization in placentae from normal and diabetic pregnancies. Placenta 1998, 19, 133–142. [Google Scholar] [CrossRef]
- Özkan, S.; Vural, B.; Fi˙li˙z, S.; Coştur, P.; Dalçik, H. Placental expression of insulin-like growth factor-I, fibroblast growth factor-basic, and neural cell adhesion molecule in preeclampsia. J. Matern. Neonatal Med. 2008, 21, 831–838. [Google Scholar] [CrossRef] [PubMed]
- Hohlagschwandtner, M.; Knöfler, M.; Ploner, M.; Zeisler, H.; Joura, E.A.; Husslein, P. Basic Fibroblast Growth Factor and Hypertensive Disorders in Pregnancy. Hypertens. Pregnancy 2002, 21, 235–241. [Google Scholar] [CrossRef]
- Yasuda, E.; Tokuda, H.; Ishisaki, A.; Hirade, K.; Kanno, Y.; Hanai, Y.; Nakamura, N.; Noda, T.; Katagiri, Y.; Kozawa, O. PPAR-gamma ligands up-regulate basic fibroblast growth factor-induced VEGF release through amplifying SAPK/JNK activation in osteoblasts. Biochem. Biophys. Res. Commun. 2005, 328, 137–143. [Google Scholar] [CrossRef]
- E Leach, R.; Romero, R.; Kim, Y.M.; Chaiworapongsa, T.; Kilburn, B.; Das, S.K.; Dey, S.K.; Johnson, A.; Qureshi, F.; Jacques, S.; et al. Pre-eclampsia and expression of heparin-binding EGF-like growth factor. Lancet 2002, 360, 1215–1219. [Google Scholar] [CrossRef]
- Leach, R.E.; Khalifa, R.; Ramirez, N.D.; Das, S.K.; Wang, J.; Dey, S.K.; Romero, R.; Armant, D.R. Multiple Roles for Heparin-Binding Epidermal Growth Factor-Like Growth Factor Are Suggested by Its Cell-Specific Expression during the Human Endometrial Cycle and Early Placentation1. J. Clin. Endocrinol. Metab. 1999, 84, 3355–3363. [Google Scholar] [CrossRef]
- Jessmon, P.; Leach, R.E.; Armant, D.R. Diverse functions of HBEGF during pregnancy. Mol. Reprod. Dev. 2009, 76, 1116–1127. [Google Scholar] [CrossRef] [Green Version]
- Imudia, A.; Kilburn, B.; Petkova, A.; Edwin, S.; Romero, R.; Armant, D. Expression of Heparin-binding EGF-like Growth Factor in Term Chorionic Villous Explants and Its Role in Trophoblast Survival. Placenta 2008, 29, 784–789. [Google Scholar] [CrossRef] [Green Version]
- Armant, D.; Fritz, R.; Kilburn, B.; Kim, Y.; Nien, J.; Maihle, N.; Romero, R.; Leach, R. Reduced expression of the epidermal growth factor signaling system in preeclampsia. Placenta 2014, 36, 270–278. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kushwaha, R.; Mishra, J.; Gupta, A.P.; Gupta, K.; Vishwakarma, J.; Chattopadhyay, N.; Gayen, J.R.; Kamthan, M.; Bandyopadhyay, S. Rosiglitazone up-regulates glial fibrillary acidic protein via HB-EGF secreted from astrocytes and neurons through PPARγ pathway and reduces apoptosis in high-fat diet-fed mice. J. Neurochem. 2019, 149, 679–698. [Google Scholar] [CrossRef] [PubMed]
- Köninger, A.; Schmidt, B.; Damaske, D.; Birdir, C.; Enekwe, A.; Kimmig, R.; Strowitzki, T.; Gellhaus, A. Follistatin during pregnancy and its potential role as an ovarian suppressing agent. Eur. J. Obstet. Gynecol. Reprod. Biol. 2017, 212, 150–154. [Google Scholar] [CrossRef] [PubMed]
- Fullerton, P.T.; Monsivais, D.; Kommagani, R.; Matzuk, M.M. Follistatin is critical for mouse uterine receptivity and decidualization. Proc. Natl. Acad. Sci. USA 2017, 114, E4772–E4781. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Petraglia, F. Inhibin, activin and follistatin in the human placenta—A new family of regulatory proteins. Placenta 1997, 18, 3–8. [Google Scholar] [CrossRef]
- Prakash, A.; Laird, S.; Tuckerman, E.; Li, T.C.; Ledger, W.L. Inhibin A and activin A may be used to predict pregnancy outcome in women with recurrent miscarriage. Fertil. Steril. 2005, 83, 1758–1763. [Google Scholar] [CrossRef]
- Prakash, A.; Li, T.C.; Tuckerman, E.; Laird, S.; Wells, M.; Ledger, W.L. A study of luteal phase expression of inhibin, activin, and follistatin subunits in the endometrium of women with recurrent miscarriage. Fertil. Steril. 2006, 86, 1723–1730. [Google Scholar] [CrossRef]
- Prakash, A.; Tuckerman, E.; Laird, S.; Ola, B.; Li, T.C.; Ledger, W. A preliminary study comparing the endometrial expression of inhibin, activin and follistatin in women with a history of implantation failure after IVF treatment and a control group. BJOG: Int. J. Obstet. Gynaecol. 2008, 115, 532–537. [Google Scholar] [CrossRef]
- Necela, B.M.; Su, W.; Thompson, E.A. Peroxisome proliferator-activated receptor gamma down-regulates follistatin in intestinal epithelial cells through SP1. J. Biol. Chem. 2008, 283, 29784–29794. [Google Scholar] [CrossRef] [Green Version]
- Ladyman, S.R.; Augustine, R.A.; Grattan, D.R. Hormone interactions regulating energy balance during pregnancy. J. Neuroendocrinol. 2010, 22, 805–817. [Google Scholar] [CrossRef]
- Armistead, B.; Johnson, E.; Vanderkamp, R.; Kula-Eversole, E.; Kadam, L.; Drewlo, S.; Kohan-Ghadr, H.-R. Placental Regulation of Energy Homeostasis During Human Pregnancy. Endocrinology 2020, 161, bqaa076. [Google Scholar] [CrossRef] [PubMed]
- Taylor, B.D.; Ness, R.B.; Olsen, J.; Hougaard, D.M.; Skogstrand, K.; Roberts, J.M.; Haggerty, C. Serum Leptin Measured in Early Pregnancy Is Higher in Women with Preeclampsia Compared with Normotensive Pregnant Women. Hypertension 2015, 65, 594–599. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Shao, Y.Y.; Ballock, R.T. Leptin Antagonizes Peroxisome Proliferator-Activated Receptor-γ Signaling in Growth Plate Chondrocytes. PPAR Res. 2012, 2012, 756198. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ma, H.; Jiang, S.; Du, L.; Liu, J.; Xu, X.; Lu, X.; Ma, L.; Zhu, H.; Wei, J.; Yu, Y. Conditioned medium from primary cytotrophoblasts, primary placenta-derived mesenchymal stem cells, or sub-cultured placental tissue promoted HUVEC angiogenesis in vitro. Stem Cell Res. Ther. 2021, 12, 141. [Google Scholar] [CrossRef]















{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Diagnosis | Gestation Age | Mode of Delivery | Maternal Age |
|---|---|---|---|
| PE | 37wks | C-section | 32 |
| PE | 39wks | C-section | 33 |
| PE | 37wks | Vaginal | 32 |
| PE | 38wks | C-section | 22 |
| PE | 37wks | Vaginal | 26 |
| PE | 37wks | Vaginal | 33 |
| PE | 35wks | Vaginal | 29 |
| PE | 37wks | Vaginal | 37 |
| PE | 37wks | C-section | 29 |
| PE | 31wks | C-section | 19 |
| sePE | 39wks | C-section | 37 |
| sePE | 37wks | Vaginal | 20 |
| sePE | 39wks | Vaginal | 20 |
| sePE | 36wks | Vaginal | 19 |
| Healthy control | 40wks | Vaginal | 33 |
| Healthy control | 39wks | Vaginal | 25 |
| Healthy control | 39wks | Vaginal | 28 |
| Healthy control | 39wks | Vaginal | 28 |
| Healthy control | 36wks | C-section | 29 |
| Healthy control | 36wks | Vaginal | 28 |
| Healthy control | 35wks | Vaginal | 33 |
| Healthy control | 36wks | C-section | 34 |
| Healthy control | 37wks | Vaginal | 36 |
| Healthy control | 37wks | C-section | 26 |
| Experimental Treatment | Culture Conditions |
|---|---|
| HUVEC Only | Standard culture medium |
| Vehicle Control | Placental culture medium (without human tissue) supplemented with DMSO and cultured for 24 h |
| Rosiglitazone Control | Placental culture medium (without human tissue) supplemented with Rosiglitazone and cultured for 24 h |
| Conditioned medium from non-treated preeclamptic placentas | Preeclamptic tissue cultured in placental culture medium for 24 h |
| Conditioned medium from preeclamptic placentas treated with vehicle | Preeclamptic tissue cultured in placental culture medium for 24 h with DMSO |
| Conditioned medium from preeclamptic placentas treated with Rosiglitazone | Preeclamptic tissue cultured in placental culture medium for 24 h with Rosiglitazone |
| Conditioned medium from non-treated healthy control placentas | Healthy placentas cultured in placental culture medium from 24 h |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Grimaldi, B.; Kohan-Ghadr, H.-R.; Drewlo, S. The Potential for Placental Activation of PPARγ to Improve the Angiogenic Profile in Preeclampsia. Cells 2022, 11, 3514. https://doi.org/10.3390/cells11213514
Grimaldi B, Kohan-Ghadr H-R, Drewlo S. The Potential for Placental Activation of PPARγ to Improve the Angiogenic Profile in Preeclampsia. Cells. 2022; 11(21):3514. https://doi.org/10.3390/cells11213514
Chicago/Turabian StyleGrimaldi, Brooke, Hamid-Reza Kohan-Ghadr, and Sascha Drewlo. 2022. "The Potential for Placental Activation of PPARγ to Improve the Angiogenic Profile in Preeclampsia" Cells 11, no. 21: 3514. https://doi.org/10.3390/cells11213514