
The Atypical Antipsychotic Lurasidone Affects Brain but Not Liver Cytochrome P450 2D (CYP2D) Activity. A Comparison with Other Novel Neuroleptics and Significance for Drug Treatment of Schizophrenia



Abstract
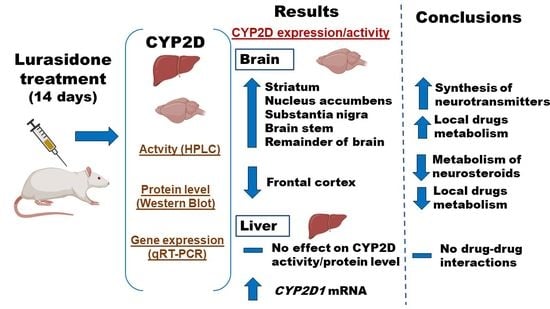
:
1. Introduction
2. Materials and Methods
2.1. Animals
2.2. Drugs and Chemicals
2.3. Animal Treatment and Preparation of Brain and Liver Microsomes
2.4. Measurement of the CYP2D Enzyme Activity in Brain and Liver Microsomes
2.5. Evaluation of CYP2D Protein in Brain and Liver Microsomes
2.6. Examination of the Expression of Genes Coding for CYP2D Enzymes in the Liver
2.7. Data Analysis
3. Results
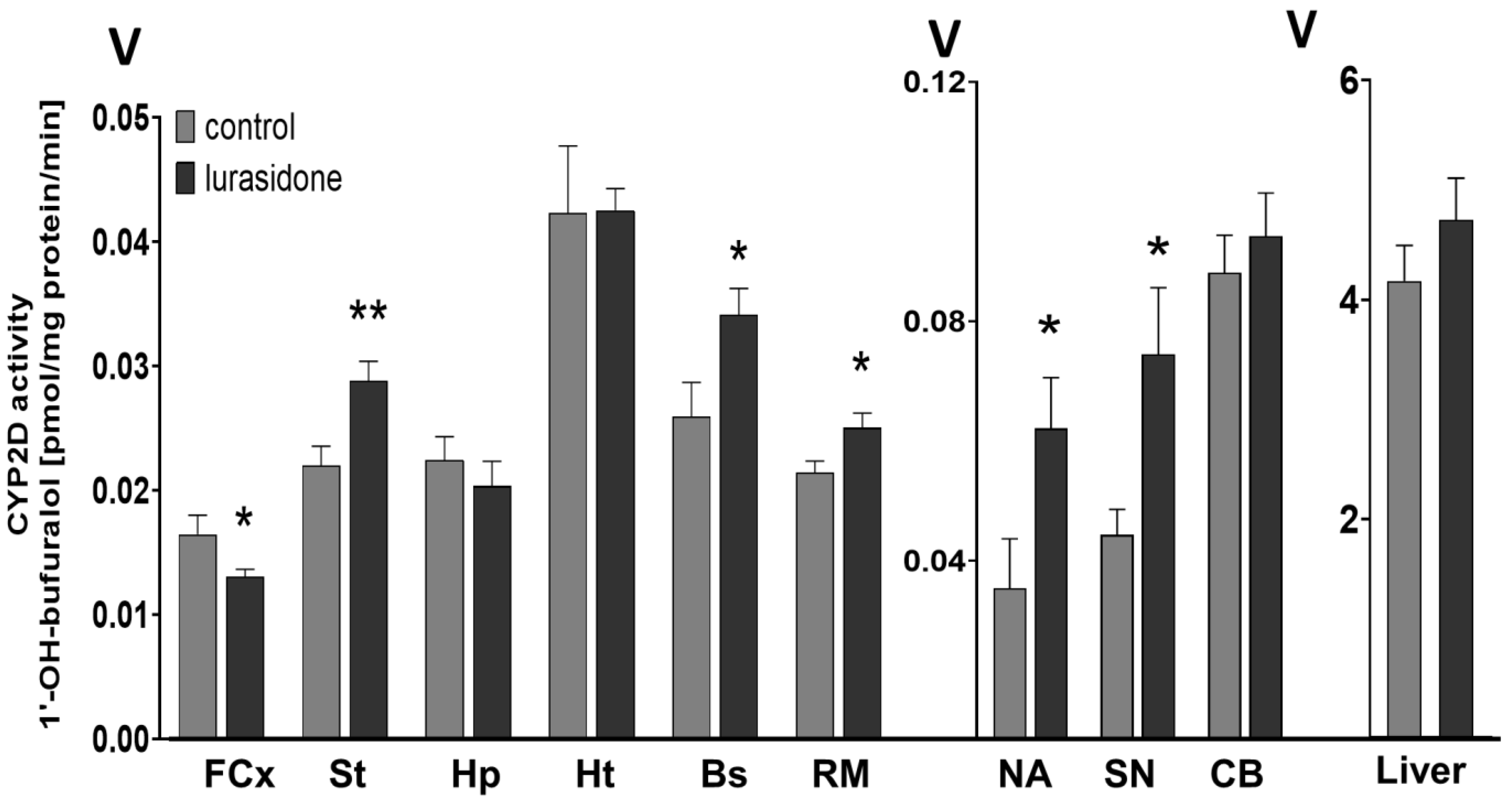
3.1. The Effect of Chronic Lurasidone Treatment on the CYP2D Activity in the Brain and Liver Microsomes of Rats
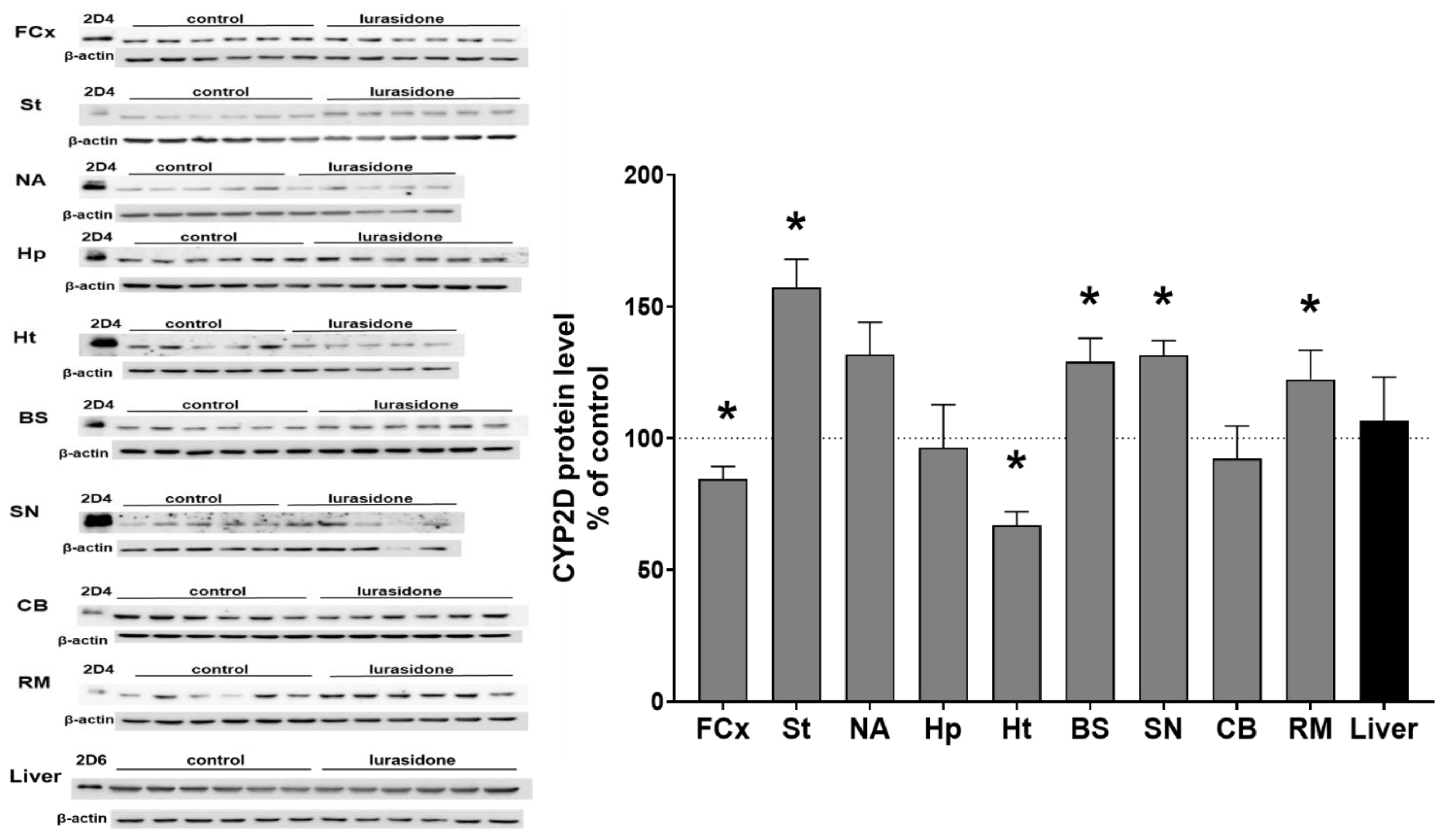
3.2. The Effect of Chronic Lurasidone Treatment on the CYP2D Protein Level in Microsomes from the Brain and Liver
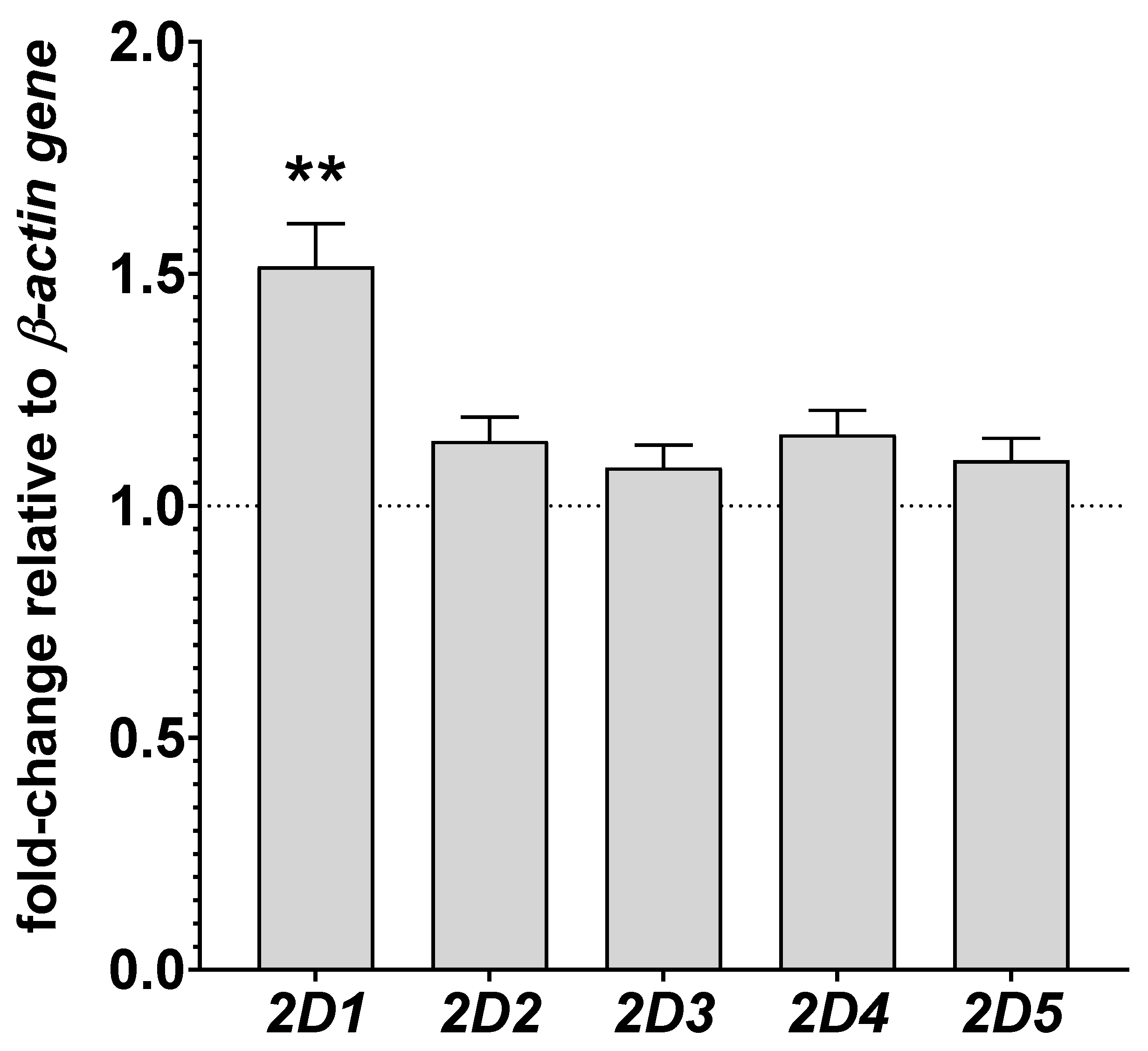
3.3. The Effect of Chronic Lurasidone Treatment on the CYP2D Gene Expression in the Rat Liver
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Zanger, U.M.; Schwab, M. Cytochrome P450 Enzymes in Drug Metabolism: Regulation of Gene Expression, Enzyme Activities, and Impact of Genetic Variation. Pharmacol. Ther. 2013, 138, 103–141. [Google Scholar] [CrossRef]
- Daniel, W.A.; Bromek, E.; Danek, P.J.; Haduch, A. The Mechanisms of Interactions of Psychotropic Drugs with Liver and Brain Cytochrome P450 and Their Significance for Drug Effect and Drug-Drug Interactions. Biochem. Pharmacol. 2022, 199, 115006. [Google Scholar] [CrossRef]
- Hiroi, T.; Imaoka, S.; Chow, T.; Funae, Y. Tissue Distributions of CYP2D1, 2D2, 2D3 and 2D4 MRNA in Rats Detected by RT-PCR. Biochim. Biophys. Acta 1998, 1380, 305–312. [Google Scholar] [CrossRef]
- Hiroi, T.; Chow, T.; Imaoka, S.; Funae, Y. Catalytic Specificity of CYP2D Isoforms in Rat and Human. Drug Metab. Dispos. 2002, 30, 970–976. [Google Scholar] [CrossRef] [Green Version]
- Wyss, A.; Gustafsson, J.A.; Warner, M. Cytochromes P450 of the 2D Subfamily in Rat Brain. Mol. Pharmacol. 1995, 47, 1148–1155. [Google Scholar]
- Martignoni, M.; Groothuis, G.M.M.; de Kanter, R. Species Differences between Mouse, Rat, Dog, Monkey and Human CYP-Mediated Drug Metabolism, Inhibition and Induction. Expert Opin. Drug Metab. Toxicol. 2006, 2, 875–894. [Google Scholar] [CrossRef]
- Meyer, R.P.; Gehlhaus, M.; Knoth, R.; Volk, B. Expression and Function of Cytochrome P450 in Brain Drug Metabolism. Curr. Drug Metab. 2007, 8, 297–306. [Google Scholar] [CrossRef]
- Miksys, S.; Tyndale, R.F. Cytochrome P450-Mediated Drug Metabolism in the Brain. J. Psychiatry Neurosci. 2013, 38, 152–163. [Google Scholar] [CrossRef] [Green Version]
- Hedlund, E.; Wyss, A.; Kainu, T.; Backlund, M.; Köhler, C.; Pelto-Huikko, M.; Gustafsson, J.A.; Warner, M. Cytochrome P4502D4 in the Brain: Specific Neuronal Regulation by Clozapine and Toluene. Mol. Pharmacol. 1996, 50, 342–350. [Google Scholar]
- Greenberg, W.M.; Citrome, L. Pharmacokinetics and Pharmacodynamics of Lurasidone Hydrochloride, a Second-Generation Antipsychotic: A Systematic Review of the Published Literature. Clin. Pharmacokinet. 2017, 56, 493–503. [Google Scholar] [CrossRef]
- Tarazi, F.I.; Stahl, S.M. Iloperidone, Asenapine and Lurasidone: A Primer on Their Current Status. Expert Opin. Pharmacother. 2012, 13, 1911–1922. [Google Scholar] [CrossRef]
- Caccia, S.; Pasina, L.; Nobili, A. Critical Appraisal of Lurasidone in the Management of Schizophrenia. Neuropsychiatr. Dis. Treat. 2012, 8, 155–168. [Google Scholar] [CrossRef]
- Jaeschke, R.R.; Sowa-Kućma, M.; Pańczyszyn-Trzewik, P.; Misztak, P.; Styczeń, K.; Datka, W. Lurasidone: The 2016 Update on the Pharmacology, Efficacy and Safety Profile. Pharmacol. Rep. 2016, 68, 748–755. [Google Scholar] [CrossRef]
- Mauri, M.C.; Paletta, S.; Maffini, M.; Colasanti, A.; Dragogna, F.; Di Pace, C.; Altamura, A.C. Clinical Pharmacology of Atypical Antipsychotics: An Update. EXCLI J. 2014, 13, 1163–1191. [Google Scholar]
- Danek, P.J.; Bromek, E.; Haduch, A.; Daniel, W.A. Chronic Treatment with Asenapine Affects Cytochrome P450 2D (CYP2D) in Rat Brain and Liver. Pharmacological Aspects. Neurochem. Int. 2021, 151, 105209. [Google Scholar] [CrossRef]
- Danek, P.J.; Daniel, W.A. Long-Term Treatment with Atypical Antipsychotic Iloperidone Modulates Cytochrome P450 2D (CYP2D) Expression and Activity in the Liver and Brain via Different Mechanisms. Cells 2021, 10, 3472. [Google Scholar] [CrossRef]
- Danek, P.J.; Wójcikowski, J.; Daniel, W.A. The Atypical Neuroleptics Iloperidone and Lurasidone Inhibit Human Cytochrome P450 Enzymes in Vitro. Evaluation of Potential Metabolic Interactions. Pharmacol. Rep. 2020, 72, 1685–1694. [Google Scholar] [CrossRef] [Green Version]
- Horiguchi, M.; Hannaway, K.E.; Adelekun, A.E.; Jayathilake, K.; Meltzer, H.Y. Prevention of the Phencyclidine-Induced Impairment in Novel Object Recognition in Female Rats by Co-Administration of Lurasidone or Tandospirone, a 5-HT(1A) Partial Agonist. Neuropsychopharmacology 2012, 37, 2175–2183. [Google Scholar] [CrossRef] [Green Version]
- Huang, M.; Panos, J.J.; Kwon, S.; Oyamada, Y.; Rajagopal, L.; Meltzer, H.Y. Comparative Effect of Lurasidone and Blonanserin on Cortical Glutamate, Dopamine, and Acetylcholine Efflux: Role of Relative Serotonin (5-HT)2A and DA D2 Antagonism and 5-HT1A Partial Agonism. J. Neurochem. 2014, 128, 938–949. [Google Scholar] [CrossRef]
- Miyauchi, M.; Neugebauer, N.M.; Oyamada, Y.; Meltzer, H.Y. Nicotinic Receptors and Lurasidone-Mediated Reversal of Phencyclidine-Induced Deficit in Novel Object Recognition. Behav. Brain Res. 2016, 301, 204–212. [Google Scholar] [CrossRef]
- Paxinos, G.; Watson, C. The Rat Brain in Stereotaxic Coordinates, 6th ed.; Academic Press: London, UK, 2007. [Google Scholar]
- Hiroi, T.; Kishimoto, W.; Chow, T.; Imaoka, S.; Igarashi, T.; Funae, Y. Progesterone Oxidation by Cytochrome P450 2D Isoforms in the Brain. Endocrinology 2001, 142, 3901–3908. [Google Scholar] [CrossRef]
- Daniel, W.A.; Haduch, A.; Wójcikowski, J. Inhibition and Possible Induction of Rat CYP2D after Short- and Long-Term Treatment with Antidepressants. J. Pharm. Pharmacol. 2002, 54, 1545–1552. [Google Scholar] [CrossRef]
- Haduch, A.; Rysz, M.; Papp, M.; Daniel, W.A. The Activity of Brain and Liver Cytochrome P450 2D (CYP2D) Is Differently Affected by Antidepressants in the Chronic Mild Stress (CMS) Model of Depression in the Rat. Biochem. Pharmacol. 2018, 156, 398–405. [Google Scholar] [CrossRef]
- Bromek, E.; Haduch, A.; Gołembiowska, K.; Daniel, W.A. Cytochrome P450 Mediates Dopamine Formation in the Brain in Vivo. J. Neurochem. 2011, 118, 806–815. [Google Scholar] [CrossRef]
- Lowry, O.H.; Rosebrough, N.J.; Farr, A.L.; Randall, R.J. Protein Measurement with the Folin Phenol Reagent. J. Biol. Chem. 1951, 193, 265–275. [Google Scholar] [CrossRef]
- Rysz, M.; Bromek, E.; Haduch, A.; Liskova, B.; Wójcikowski, J.; Daniel, W.A. The Reverse Role of the Hypothalamic Paraventricular (PVN) and Arcuate (ARC) Nuclei in the Central Serotonergic Regulation of the Liver Cytochrome P450 Isoform CYP2C11. Biochem. Pharmacol. 2016, 112, 82–89. [Google Scholar] [CrossRef]
- Marx, C.E.; Lee, J.; Subramaniam, M.; Rapisarda, A.; Bautista, D.C.T.; Chan, E.; Kilts, J.D.; Buchanan, R.W.; Wai, E.P.; Verma, S.; et al. Proof-of-Concept Randomized Controlled Trial of Pregnenolone in Schizophrenia. Psychopharmacology 2014, 231, 3647–3662. [Google Scholar] [CrossRef]
- Kreinin, A.; Bawakny, N.; Ritsner, M.S. Adjunctive Pregnenolone Ameliorates the Cognitive Deficits in Recent-Onset Schizophrenia: An 8-Week, Randomized, Double-Blind, Placebo-Controlled Trial. Clin. Schizophr. Relat. Psychoses 2017, 10, 201–210. [Google Scholar] [CrossRef]
- Cai, H.; Cao, T.; Zhou, X.; Yao, J.K. Neurosteroids in Schizophrenia: Pathogenic and Therapeutic Implications. Front. Psychiatry 2018, 9, 73. [Google Scholar] [CrossRef] [Green Version]
- Ratner, M.H.; Kumaresan, V.; Farb, D.H. Neurosteroid Actions in Memory and Neurologic/Neuropsychiatric Disorders. Front. Endocrinol. 2019, 10, 169. [Google Scholar] [CrossRef]
- Haduch, A.; Bromek, E.; Kot, M.; Kamińska, K.; Gołembiowska, K.; Daniel, W.A. The Cytochrome P450 2D-Mediated Formation of Serotonin from 5-Methoxytryptamine in the Brain in Vivo: A Microdialysis Study. J. Neurochem. 2015, 133, 83–92. [Google Scholar] [CrossRef]
- Stahl, S.M. Stahl’s Essential Psychopharmacology: Neuroscientific Basis and Practical Applications; Cambridge University Press: Cambridge, UK, 2013; ISBN 978-1-139-83259-5. [Google Scholar]
- Haduch, A.; Daniel, W.A. The Engagement of Brain Cytochrome P450 in the Metabolism of Endogenous Neuroactive Substrates: A Possible Role in Mental Disorders. Drug Metab. Rev. 2018, 50, 415–429. [Google Scholar] [CrossRef]
- Fregoneze, J.B.; Ferreira, H.S.; Luz, C.P.N. Brain Serotonergic Receptors and Control of Fluid Intake and Cardiovascular Function in Rats. In Neurobiology of Body Fluid Homeostasis: Transduction and Integration; De Luca, L.A., Menani, J.V., Johnson, A.K., Eds.; Frontiers in Neuroscience; CRC Press/Taylor & Francis: Boca Raton, FL, USA, 2014; ISBN 978-1-4665-0693-0. [Google Scholar]
- Beaulieu, J.-M.; Espinoza, S.; Gainetdinov, R.R. Dopamine Receptors—IUPHAR Review 13. Br. J. Pharmacol. 2015, 172, 1–23. [Google Scholar] [CrossRef]
- McMillan, D.M.; Tyndale, R.F. CYP-Mediated Drug Metabolism in the Brain Impacts Drug Response. Pharmacol. Ther. 2018, 184, 189–200. [Google Scholar] [CrossRef]
- Kuban, W.; Daniel, W.A. Cytochrome P450 Expression and Regulation in the Brain. Drug Metab. Rev. 2021, 53, 1–29. [Google Scholar] [CrossRef]
- Findling, R.L.; Goldman, R.; Chiu, Y.-Y.; Silva, R.; Jin, F.; Pikalov, A.; Loebel, A. Pharmacokinetics and Tolerability of Lurasidone in Children and Adolescents with Psychiatric Disorders. Clin. Ther. 2015, 37, 2788–2797. [Google Scholar] [CrossRef] [Green Version]
- Fukuyama, K.; Motomura, E.; Shiroyama, T.; Okada, M. Impact of 5-HT7 Receptor Inverse Agonism of Lurasidone on Monoaminergic Tripartite Synaptic Transmission and Pathophysiology of Lower Risk of Weight Gain. Biomed. Pharmacother. 2022, 148, 112750. [Google Scholar] [CrossRef]
- Choudhury, M.; Qadri, I.; Rahman, S.M.; Schroeder-Gloeckler, J.; Janssen, R.C.; Friedman, J.E. C/EBPβ Is AMP Kinase Sensitive and up-Regulates PEPCK in Response to ER Stress in Hepatoma Cells. Mol. Cell Endocrinol. 2011, 331, 102–108. [Google Scholar] [CrossRef] [Green Version]
- Jover, R.; Bort, R.; Gómez-Lechón, M.J.; Castell, J.V. Re-Expression of C/EBP Alpha Induces CYP2B6, CYP2C9 and CYP2D6 Genes in HepG2 Cells. FEBS Lett. 1998, 431, 227–230. [Google Scholar] [CrossRef] [Green Version]
- Burgermeister, E.; Chuderland, D.; Hanoch, T.; Meyer, M.; Liscovitch, M.; Seger, R. Interaction with MEK Causes Nuclear Export and Downregulation of Peroxisome Proliferator-Activated Receptor γ. Mol. Cell. Biol. 2007, 27, 803–817. [Google Scholar] [CrossRef] [Green Version]
- Kaplan, J.M.; Hake, P.W.; Denenberg, A.; Nowell, M.; Piraino, G.; Zingarelli, B. Phosphorylation of Extracellular Signal-Regulated Kinase (ERK)-1/2 Is Associated with the Downregulation of Peroxisome Proliferator–Activated Receptor (PPAR)-γ during Polymicrobial Sepsis. Mol. Med. 2010, 16, 491–497. [Google Scholar] [CrossRef] [PubMed]
- Zhang, F.; Li, J.; Na, S.; Wu, J.; Yang, Z.; Xie, X.; Wan, Y.; Li, K.; Yue, J. The Involvement of PPARs in the Selective Regulation of Brain CYP2D by Growth Hormone. Neuroscience 2018, 379, 115–125. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Tissue | CYP2D Activity (% of Control) | CYP2D Protein Level (% of Control) | |
|---|---|---|---|
| Brain structures | FCx | 77 ± 6.7 ↓ * | 84.5 ± 4.8 ↓ * |
| St | 131 ± 15.3 ↑ ** | 157.2 ± 10.8 ↑ * | |
| NA | 175.3 ± 42.9 ↑ * | 131.8 ± 11.2 | |
| Hp | 88.9 ± 16.7 | 96.4 ± 16.4 | |
| Ht | 99.7 ± 10.4 | 66.9 ± 4.6 ↓ * | |
| BS | 133.9 ± 17.4 ↑ * | 128.9 ± 8.9 ↑ * | |
| SN | 154.5 ± 35.9 ↑ * | 131.5 ± 5.1 ↑ * | |
| Cb | 98.7 ± 23.3 | 92.2 ± 12.4 | |
| RM | 115.9 ± 15.8 ↑ * | 122.4 ± 11.1 ↑ * | |
| Liver | 106.1 ± 28.6 | 106.6 ± 6.7 | |
| Drug | Liver | Brain Structures | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| FCx | St | NA | Hp | Ht | BS | SN | Cb | RM | ||
| Lurasidone (D2, 5-HT2A, 5-HT7, 5-HT1A, α2C) |  |  |  | | |  | | | | |
| Iloperidone (D2, D3, 5-HT2A, α1, α2) | | | | | | | | | | |
| Asenapine (D1, D2, D3, D4, 5-HT1A, 5-HT1B, 5-HT2A, 5-HT2B, 5-HT2C, 5-HT5A, 5-HT6, 5-HT7, α1, α2, H1) | | | | | | | | | | |
 ,
,  —increase or decrease in activity, respectively;
—increase or decrease in activity, respectively;  ,
,  —increase or decrease in protein level, respectively;
—increase or decrease in protein level, respectively;  —no effect. Receptors placed in the brackets display Ki below 20 nM [11]. FCx—the frontal cortex, St—the striatum, NA—the nucleus accumbens, Hp—the hippocampus, Ht—the hypothalamus, BS—the brain stem, SN—the substantia nigra, Cb—the cerebellum, RM—the rest of brain.
—no effect. Receptors placed in the brackets display Ki below 20 nM [11]. FCx—the frontal cortex, St—the striatum, NA—the nucleus accumbens, Hp—the hippocampus, Ht—the hypothalamus, BS—the brain stem, SN—the substantia nigra, Cb—the cerebellum, RM—the rest of brain.Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Danek, P.J.; Daniel, W.A. The Atypical Antipsychotic Lurasidone Affects Brain but Not Liver Cytochrome P450 2D (CYP2D) Activity. A Comparison with Other Novel Neuroleptics and Significance for Drug Treatment of Schizophrenia. Cells 2022, 11, 3513. https://doi.org/10.3390/cells11213513
Danek PJ, Daniel WA. The Atypical Antipsychotic Lurasidone Affects Brain but Not Liver Cytochrome P450 2D (CYP2D) Activity. A Comparison with Other Novel Neuroleptics and Significance for Drug Treatment of Schizophrenia. Cells. 2022; 11(21):3513. https://doi.org/10.3390/cells11213513
Chicago/Turabian StyleDanek, Przemysław J., and Władysława A. Daniel. 2022. "The Atypical Antipsychotic Lurasidone Affects Brain but Not Liver Cytochrome P450 2D (CYP2D) Activity. A Comparison with Other Novel Neuroleptics and Significance for Drug Treatment of Schizophrenia" Cells 11, no. 21: 3513. https://doi.org/10.3390/cells11213513