

Volatile Compounds in Actinomycete Communities: A New Tool for Biosynthetic Gene Cluster Activation, Cooperative Growth Promotion, and Drug Discovery
 , and
, and 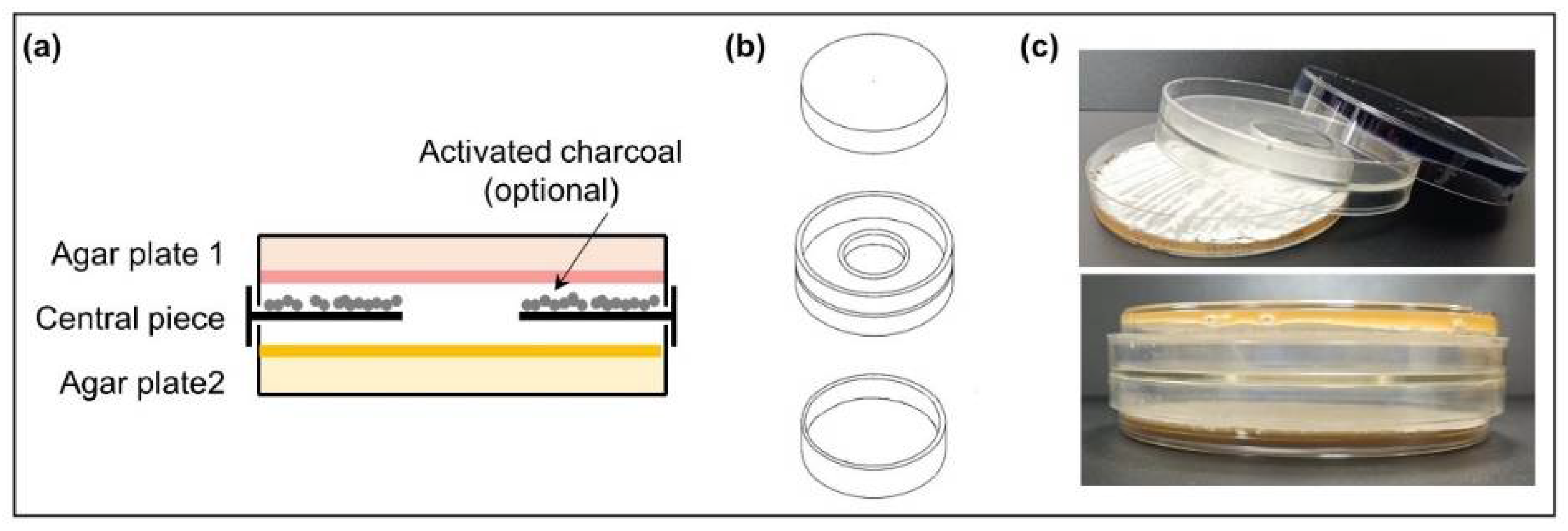{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Strains and Culture Conditions
2.2. Dual-Culture Actinomycetes in VOC Chambers
2.3. Multiple Coculture of Actinomycetes
2.4. Antibiotic Production in Co-Culture
2.5. Extraction of Secondary Metabolites Produced in VOC Chambers, Analysis with UPLC, and Dereplication
2.6. Purification of Rumycins
2.7. Biosynthetic Gene Cluster Prediction and Sequence Analysis
3. Results and Discussion
3.1. Morphological and Developmental State of Actinomycete strains in VOC Chambers
3.2. New or Incremental Increases in Antibiotic Production in VOC Chambers
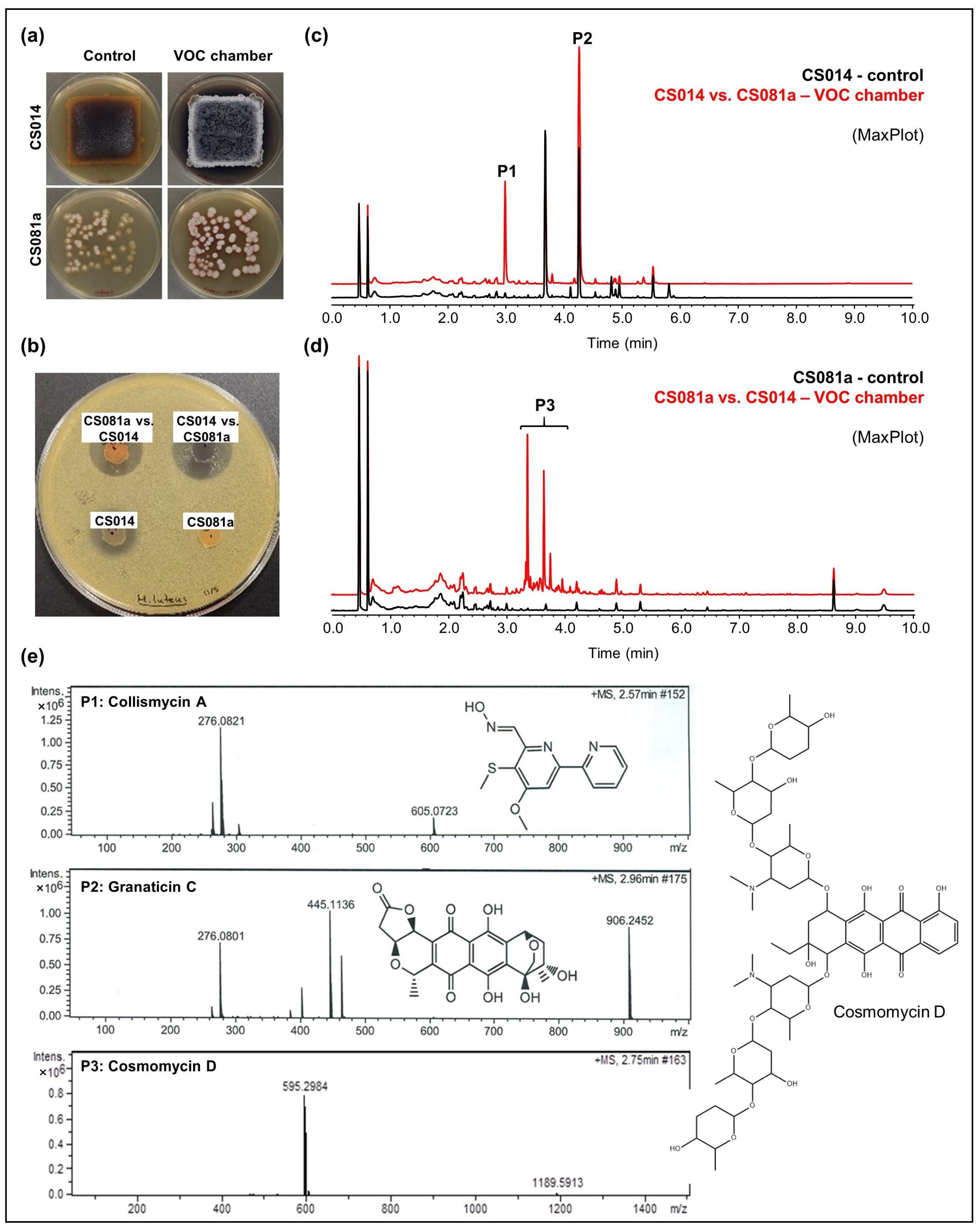
- CS014: An increased antibiotic activity against M. luteus was detected in coculture with CS057, CS081a, CS090a, CS131, and CS149 strains grown on YMA plates (Figure 3a,b). Comparative UPLC analysis revealed the activation of collismycin production in the presence of VOCs from the strains mentioned above. Furthermore, increased production of granaticin C was observed (Figure 3c).
- CS081a: This strain only demonstrated anti-M. luteus activity when grown on YMA medium in VOC chambers cultured against CS014 strain (Figure 3b). The chromatographic analysis of samples extracted with ethyl acetate exposed the biosynthetic induction of the cosmomycin anthracycline antibiotic family by CS081a under the effect of CS014 VOCs (Figure 3d). In this case, the signaling caused by volatiles emitted by the strains inside the chamber was bidirectional as VOCs from CS014 impacted the secondary metabolism of the CS081a strain and vice versa (see the previous paragraph).
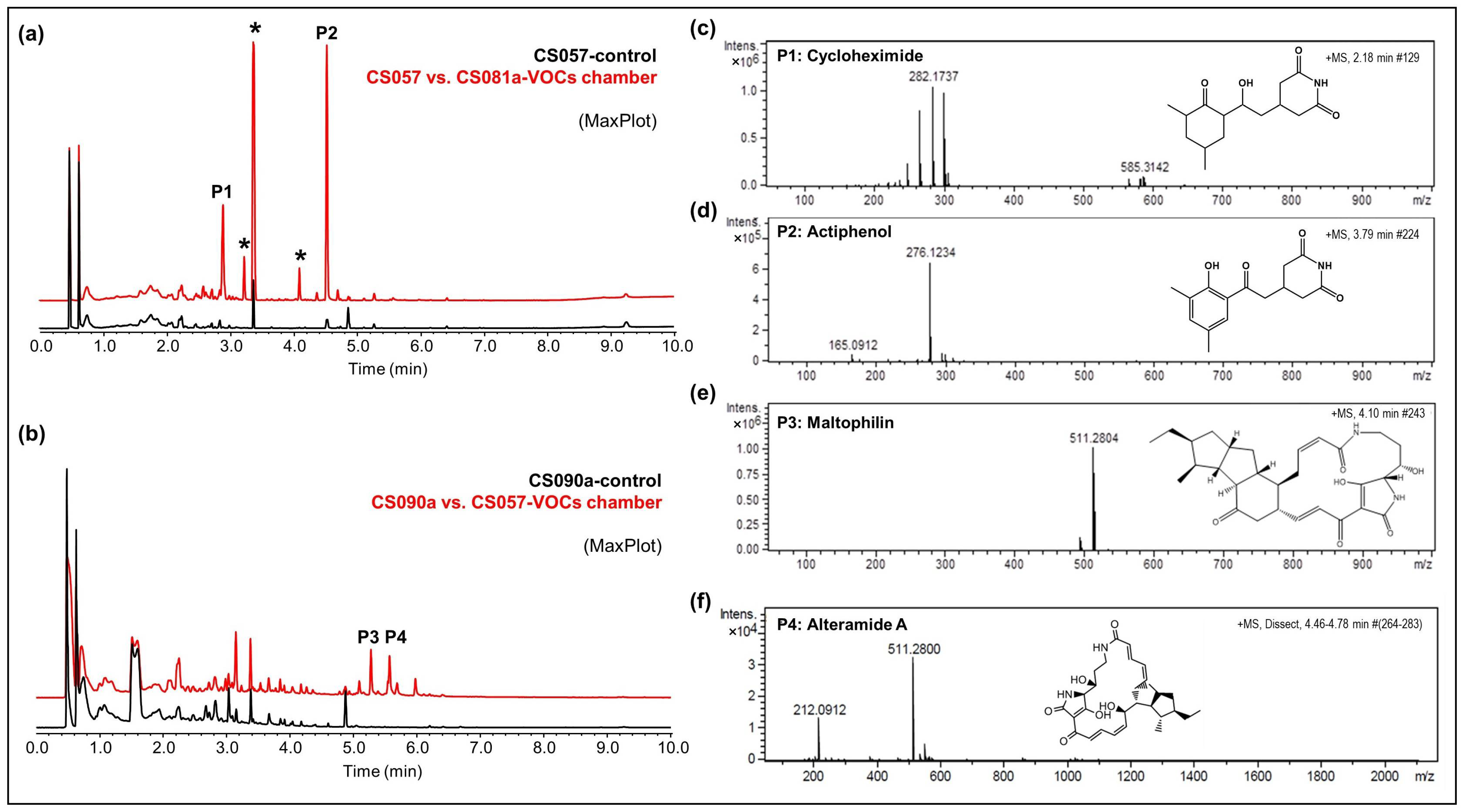
- CS057: The strain growing on YMA agar plates in a VOC chamber together with CS081a demonstrated stronger bioactivity against M. luteus and E. weberi. UPLC analysis revealed an increased production of the related compounds cycloheximide and actiphenol by CS057 exposed to VOCs from CS081a (Figure 4a,c,d).
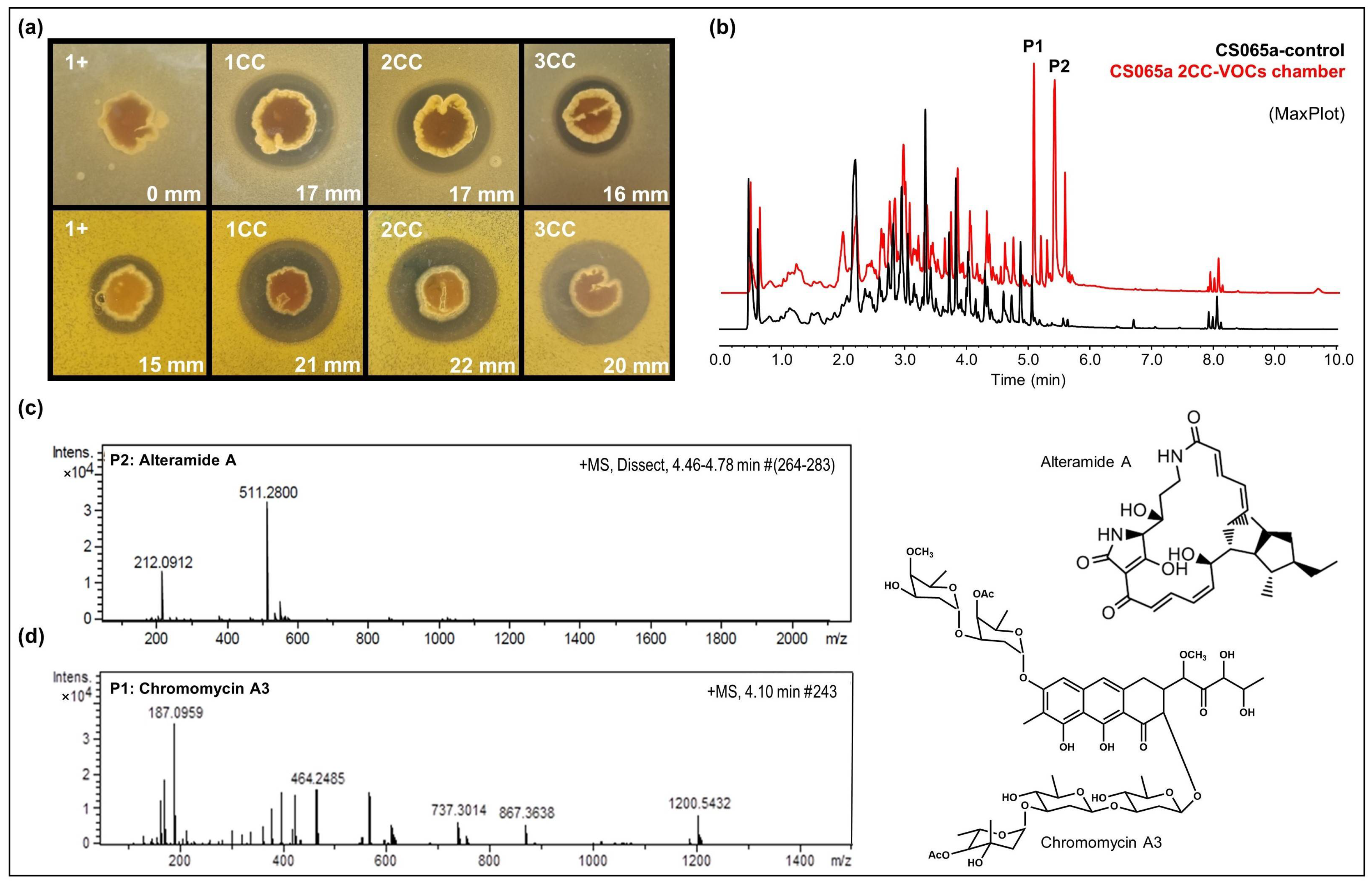
- CS090a: Antifungal compounds active against E. weberi were only produced by the CS090a strain when it was grown on YMA under the effect of VOCs originating from CS057 and CS081a. Chromatographic analysis found maltophilin and alteramide activation of the production (Figure 4b,e,f).

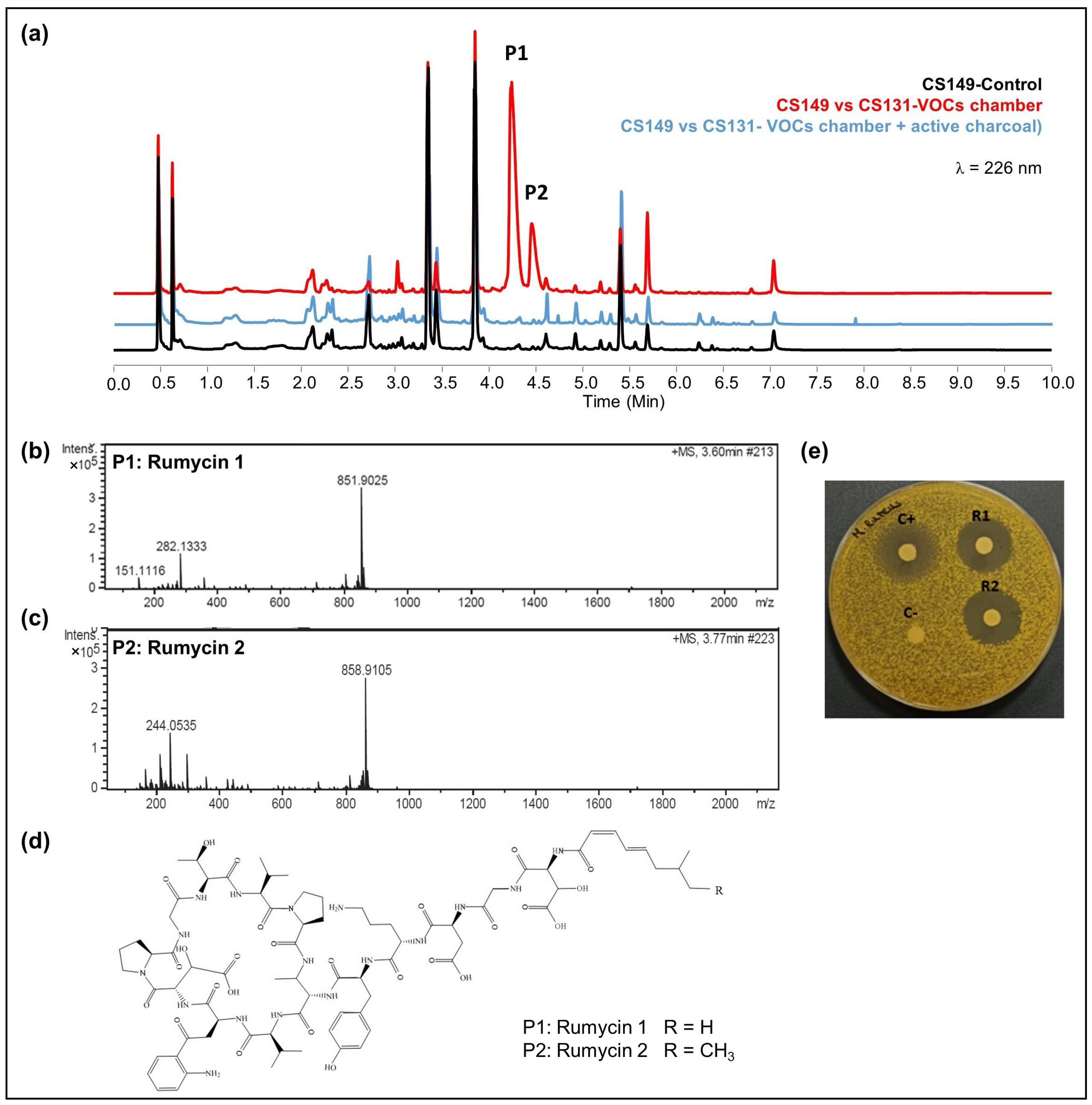
- CS149: No antibiotic activity against the tested microorganisms was detected when CS149 was grown in monoculture. However, when paired with CS081a or CS131 in VOC chambers, potent bioactivity against M. luteus was observed. Chromatographic analysis highlighted the activation of the production of two different compounds that were not present in samples extracted from monocultures of CS149 (Figure 5a). These compounds were identified by dereplication as the anti-MRSA cyclic lipopeptides rumycin 1 and rumycin 2 (Figure 5b–d). In a step-forward study to confirm that the observed alteration of the CS149 secondary metabolism was due to the VOCs emitted by CS131, a VOC chamber with 2 g of activated charcoal was assembled and then the samples extracted with ethyl acetate from SFM CS149-CS131 dual cultures were analyzed by UPLC. No rumycins could be detected (Figure 5a) indicating that VOCs from CS131 were adsorbed by the activated charcoal, and thus could not exert their inductive effect on CS149 biosynthetic machinery. On the other hand, biosynthesis of rumycins by Streptomyces sp. CS149 does not depend strictly on the growth medium of the strain; this has been verified in both SFM and R5A. Otherwise, the VOCs that induced the activation of the secondary metabolism of Streptomyces sp. CS149 were only produced by CS081a or CS131 if grown on SFM since rumycins could not be detected when the VOC-emitting strain was cultured on R5A or YMA. We purified 3.6 mg of rumycin 1 and 2.8 mg of rumycin 2 from thirty plates (total volume of 600 mL) of SFM (dual cultures between CS149 and CS131 strains in VOC chambers). Pure compounds were used for the testing of bioactivity via agar diffusion bioassay. They demonstrated potent antibacterial activity against M. luteus, even stronger than the commonly used antibiotic apramycin, but no activity against E. coli, C. albicans, and E. weberi (Figure 5e).

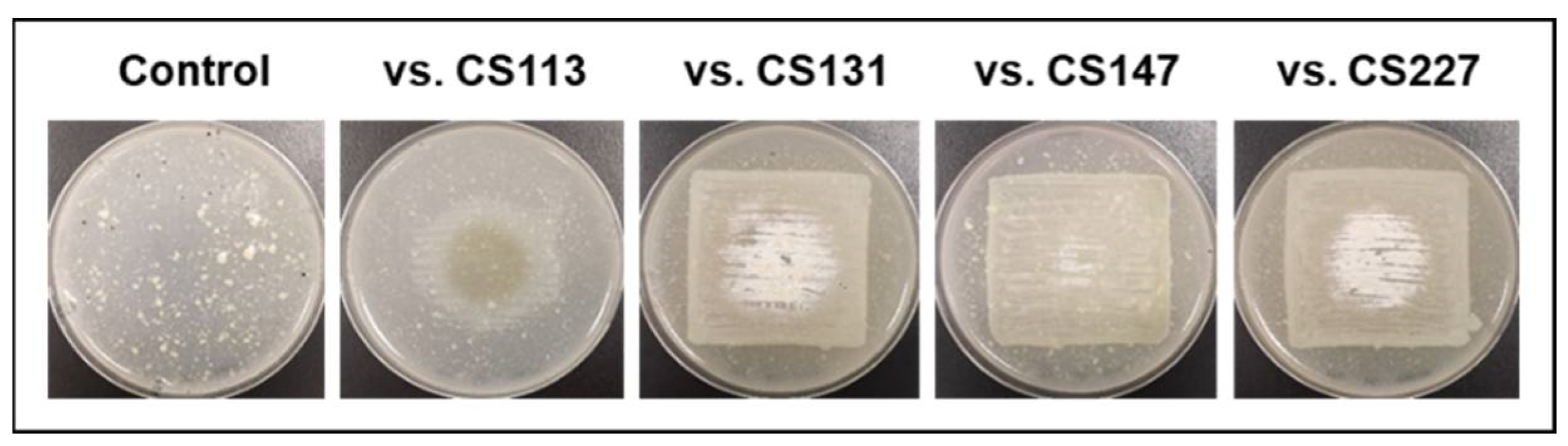
- CS194: When cultivated on SFM, the strain CS194 exposed to VOCs emitted by strains CS014, CS057, CS081a, or CS227, presented antibiotic activity versus M. luteus. As mentioned previously, this strain was not able to grow on SFM in monoculture, thus we could not state if the production of the antibiotic/s was due to a signaling effect of VOCs on growth promotion or by the induction of the CS194 secondary metabolism (or both). In any case, the antibacterial activity of CS194 could be explained by the production of bioactive piperazinediones, detected by dereplication in samples of CS194 (Figure S2, Supplementary data).
- CS207: The antibacterial (against M. luteus and E. coli) and antifungal activities of the strain CS207 were higher when cultivated on YMA in VOC chambers together with strains CS014, CS057, CS081a, or CS227. Unfortunately, the comparative UPLC analysis did not retrieve any differential peak that could explain the observed antibiotic activity, probably due to the limitation of the analytics based on UV absorbance measurements.
3.3. Effect of VOCs in Multiple Co-Culture
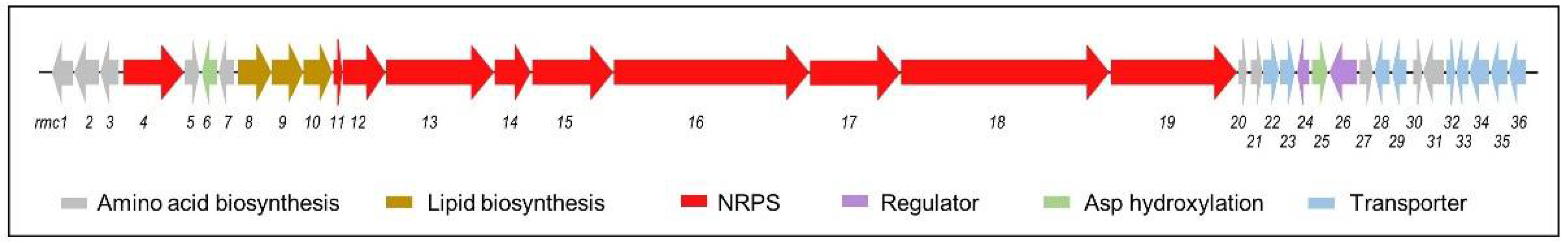
3.4. Identification of the Rumycin Biosynthetic Gene Cluster
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- WHO. Thirteenth General Programme of Work, 2019–2023. Geneva: World Health Organization. 2019. Available online: https://apps.who.int/iris/bitstream/handle/10665/324775/WHO-PRP-18.1-eng.pdf (accessed on 24 March 2022).
- Hutchings, M.I.; Truman, A.W.; Wilkinson, B. Antibiotics: Past, present and future. Curr. Opin. Microbiol. 2019, 51, 72–80. [Google Scholar] [CrossRef] [PubMed]
- Bernal, F.A.; Hammann, P.; Kloss, F. Natural products in antibiotic development: Is the success story over? Curr. Opin. Biotechnol. 2022, 78, 102783. [Google Scholar] [CrossRef] [PubMed]
- Hughes, C.C.; Prieto-Davo, A.; Jensen, P.R.; Fenical, W. The marinopyrroles, antibiotics of an unprecedented structure class from a marine Streptomyces sp. Org. Lett. 2008, 10, 629–631. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bister, B.; Bischoff, D.; Strobele, M.; Riedlinger, J.; Reicke, A.; Wolter, F.; Bull, A.T.; Zahner, H.; Fiedler, H.P.; Süssmuth, R.D. Abyssomicin C—A polycyclic antibiotic from a marine Verrucosispora strain as an inhibitor of the p-aminobenzoic acid/tetrahydrofolate biosynthesis pathway. Angew. Chem. Int. Edit. 2004, 43, 2574–2576. [Google Scholar] [CrossRef]
- Chevrette, M.G.; Carlson, C.M.; Ortega, H.E.; Thomas, C.; Ananiev, G.E.; Barns, K.J.; Book, A.J.; Cagnazzo, J.; Carlos, C.; Flanigan, W.; et al. The antimicrobial potential of Streptomyces from insect microbiomes. Nat. Commun. 2019, 10, 516. [Google Scholar] [CrossRef] [Green Version]
- Malmierca, M.G.; González-Montes, L.; Pérez-Victoria, I.; Sialer, C.; Braña, A.F.; García Salcedo, R.; Martín, J.; Reyes, F.; Méndez, C.; Olano, C.; et al. Searching for glycosylated natural products in Actinomycetes and identification of novel macrolactams and angucyclines. Front. Microbiol. 2018, 9, 39. [Google Scholar] [CrossRef]
- Zhang, X.; Hindra Elliot, M.A. Unlocking the trove of metabolic treasures: Activating silent biosynthetic gene clusters in bacteria and fungi. Curr. Opin. Microbiol. 2019, 51, 9–15. [Google Scholar] [CrossRef]
- Privalsky, T.M.; Soohoo, A.M.; Wang, J.; Walsh, C.T.; Wright, G.D.; Gordon, E.M.; Gray, N.S.; Khosla, C. Prospects for antibacterial discovery and development. J. Am. Chem. Soc. 2021, 143, 21127–21142. [Google Scholar] [CrossRef]
- Hemmerling, F.; Piel, J. Strategies to access biosynthetic novelty in bacterial genomes for drug discovery. Nat. Rev. Drug Discov. 2022, 21, 359–378. [Google Scholar] [CrossRef]
- Shentu, X.; Liu, N.; Tang, G.; Tanaka, Y.; Ochi, K.; Xu, J.; Yu, X. Improved antibiotic production and silent gene activation in Streptomyces. diastatochromogenes by ribosome engineering. J. Antibiot. 2016, 69, 406–410. [Google Scholar] [CrossRef]
- Pan, R.; Bai, X.; Chen, J.; Zhang, H.; Wang, H. Exploring Structural Diversity of Microbe Secondary Metabolites Using OSMAC Strategy: A Literature Review. Front. Microbiol. 2019, 26, 294. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zong, G.; Fu, J.; Zhang, P.; Zhang, W.; Xu, Y.; Cao, G.; Zhang, R. Use of elicitors to enhance or activate the antibiotic production in Streptomyces. Crit. Rev. Biotechnol. 2021, 1–24. [Google Scholar] [CrossRef] [PubMed]
- Aksenov, A.A.; Laponogov, I.; Zhang, Z.; Doran, S.L.F.; Belluomo, I.; Veselkov, D.; Bittremieux, W.; Nothias, L.F.; Nothias-Esposito, M.; Maloney, K.N.; et al. Auto-deconvolution and molecular networking of gas chromatography–mass spectrometry data. Nat. Biotechnol. 2021, 39, 169–173. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Clarke, J.A.; McCann, S.; Hillier, N.K.; Tahlan, K. Analysis of Streptomyces volatilomes using global molecular networking reveals the presence of metabolites with diverse biological activities. Microbiol. Spectr. 2022, 10, e0055222. [Google Scholar] [CrossRef]
- Rowan, D.D. Volatile metabolites. Metabolites 2011, 1, 41–63. [Google Scholar] [CrossRef] [Green Version]
- Bérdy, J. Bioactive microbial metabolites. J. Antibiot. 2005, 58, 1–26. [Google Scholar] [CrossRef] [Green Version]
- Tyc, O.; de Jager, V.C.L.; van den Berg, M.; Gerards, S.; Janssens, T.K.S.; Zaagman, N.; Kai, M.; Svatos, A.; Zweers, H.; Hordijk, C.; et al. Exploring bacterial interspecific interactions for discovery of novel antimicrobial compounds. Microb. Biotechnol. 2017, 10, 910–925. [Google Scholar] [CrossRef] [Green Version]
- Avalos, M.; van Wezel, G.P.; Raaijmakers, J.M.; Garbeva, P. Healthy scents: Microbial volatiles as new frontier in antibiotic research? Curr. Opin. Microbiol. 2018, 45, 84–91. [Google Scholar] [CrossRef] [Green Version]
- Meredith, L.K.; Tfaily, M.M. Capturing the microbial volatilome: An oft overlooked ‘ome’. Trends Microbiol. 2022, 30, 622–631. [Google Scholar] [CrossRef]
- De Fine Licht, H.H.; Boomsma, J.J.; Tunlid, A. Symbiotic adaptations in the fungal cultivar of leaf-cutting ants. Nat. Commun. 2014, 5, 5675. [Google Scholar] [CrossRef]
- Currie, C.R.; Scott, J.A.; Summerbell, R.C.; Malloch, D. Fungus-growing ants use antibiotic-producing bacteria to control garden parasites. Nature 1999, 398, 701–704. [Google Scholar] [CrossRef]
- Batey, S.F.D.; Greco, C.; Hutchings, M.I.; Wilkinson, B. Chemical warfare between fungus-growing ants and their pathogens. Curr. Opin. Chem. Biol. 2020, 59, 172–181. [Google Scholar] [CrossRef]
- Dhodary, B.; Spiteller, D. Ammonia production by Streptomyces symbionts of Acromyrmex leaf-cutting ants strongly inhibits the fungal pathogen Escovopsis. Microorganisms 2021, 9, 1622. [Google Scholar] [CrossRef] [PubMed]
- Álvarez-García, S.; Mayo-Prieto, S.; Carro-Huerga, G.; Rodríguez-González, Á.; González-López, Ó.; Gutiérrez, S.; Casquero, P.A. Volatile organic compound chamber: A novel technology for microbiological volatile interaction assays. J. Fungi 2021, 7, 248. [Google Scholar] [CrossRef]
- Prado-Alonso, L.; Pérez-Victoria, I.; Malmierca, M.G.; Montero, I.; Rioja-Blanco, E.; Martín, J.; Reyes, F.; Méndez, C.; Salas, J.A.; Olano, C. Colibrimycins, novel halogenated hybrid polyketide synthase-nonribosomal peptide synthetase (PKS-NRPS) compounds produced by Streptomyces sp. Strain CS147. Appl. Environ. Microbiol. 2022, 88, e0183921. [Google Scholar] [CrossRef] [PubMed]
- Sánchez, L.; Braña, A.F. Cell density influences antibiotic biosynthesis in Streptomyces. clavuligerus. Microbiology 1996, 142, 1209–1220. [Google Scholar] [CrossRef] [Green Version]
- Fernández, E.; Weißbach, U.; Reillo, C.S.; Braña, A.F.; Méndez, C.; Rohr, J.; Salas, J.A. Identification of two genes from Streptomyces. argillaceus encoding glycosyltransferases involved in transfer of a disaccharide during biosynthesis of the antitumor drug mithramycin. J. Bacteriol. 1998, 180, 4929–4937. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hobbs, G.; Frazer, C.M.; Gardner, D.C.J.; Cullum, J.A.; Oliver, S.G. Dispersed growth of Streptomyces in liquid culture. Appl. Microbiol. Biotechnol. 1989, 31, 272–277. [Google Scholar] [CrossRef]
- Perez-Victoria, I.; Martin, J.; Reyes, F. Combined LC/UV/MS and NMR strategies for the dereplication of marine natural products. Planta Med. 2016, 82, 857–871. [Google Scholar] [CrossRef] [Green Version]
- Buckingham, J. Dictionary of Natural Products on DVD; CRC Press: Boca Raton, FL, USA, 2017; Volume 26.2. [Google Scholar]
- Blin, K.; Shaw, S.; Kloosterman, A.M.; Charlop-Powers, Z.; van Wezel, G.P.; Medema, M.H.; Weber, T. antiSMASH 6.0: Improving cluster detection and comparison capabilities. Nucleic Acids Res. 2021, 49, W29–W35. [Google Scholar] [CrossRef]
- Kautsar, S.A.; Blin, K.; Shaw, S.; Navarro-Muñoz, J.C.; Terlouw, B.R.; van der Hooft, J.J.J.; van Santen, J.A.; Tracanna, V.; Suarez Duran, H.G.; Pascal Andreu, V.; et al. MIBiG 2.0: A repository for biosynthetic gene clusters of known function. Nucleic Acids Res. 2020, 48, D454–D458. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jones, S.E.; Ho, L.; Rees, C.A.; Hill, J.E.; Nodwell, J.R.; Elliot, M.A. Streptomyces exploration is triggered by fungal interactions and volatile signals. Elife 2017, 6, e21738. [Google Scholar] [CrossRef] [PubMed]
- Curtis, T.P.; Sloan, W.T. Prokaryotic diversity and its limits: Microbial community structure in nature and implications for microbial ecology. Curr. Opin. Microbiol. 2004, 7, 221–226. [Google Scholar] [CrossRef]
- Straight, P.D.; Kolter, R. Interspecies chemical communication in bacterial development. Annu. Rev. Microbiol. 2009, 63, 99–118. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yamanaka, K.; Oikawa, H.; Ogawa, H.O.; Hosono, K.; Shinmachi, F.; Takano, H.; Sakuda, S.; Beppu, T.; Ueda, K. Desferrioxamine E produced by Streptomyces. griseus stimulates growth and development of Streptomyces tanashiensis. Microbiology 2005, 151, 2899–2905. [Google Scholar] [CrossRef]
- Onaka, H.; Tabata, H.; Igarashi, Y.; Sato, Y.; Furumai, T. Goadsporin, a chemical substance which promotes secondary metabolism and morphogenesis in streptomycetes. I. Purification and characterization. J. Antibiot. 2001, 54, 1036–1044. [Google Scholar] [CrossRef] [Green Version]
- Kato, J.Y.; Funa, N.; Watanabe, H.; Ohnishi, Y.; Horinouchi, S. Biosynthesis of gamma-butyrolactone autoregulators that switch on secondary metabolism and morphological development in Streptomyces. Proc. Natl. Acad. Sci. USA 2007, 104, 2378–2383. [Google Scholar] [CrossRef] [Green Version]
- Xu, G.; Yang, S. Regulatory and evolutionary roles of pseudo γ-butyrolactone receptors in antibiotic biosynthesis and resistance. Appl. Microbiol. Biotechnol. 2019, 103, 9373–9378. [Google Scholar] [CrossRef]
- Ceniceros, A.; Cuervo, L.; Méndez, C.; Salas, J.A.; Olano, C.; Malmierca, M.G. A multidisciplinary approach to unraveling the natural product biosynthetic potential of a Streptomyces strain collection isolated from leaf-cutting ants. Microorganisms 2021, 9, 2225. [Google Scholar] [CrossRef]
- Klassen, J.L. Microbial secondary metabolites and their impacts on insect symbioses. Curr. Opin. Insect Sci. 2014, 4, 15–22. [Google Scholar] [CrossRef]
- Cafaro, M.J.; Poulsen, M.; Little, A.E.; Price, S.L.; Gerardo, N.M.; Wong, B.; Stuart, A.E.; Larget, B.; Abbot, P.; Currie, C.R. Specificity in the symbiotic association between fungus-growing ants and protective Pseudonocardia bacteria. Proc. Biol. Sci. 2011, 278, 1814–1822. [Google Scholar] [CrossRef] [Green Version]
- Elson, A.L.; Box, S.J.; Gilpin, M.L. New quinone antibiotics of the granaticin type, isolated from Streptomyces. lateritius. I. Production, isolation and properties. J. Antibiot. 1988, 41, 570–572. [Google Scholar] [CrossRef] [PubMed]
- Ding, Y.; Li, Y.; Li, Z.; Zhang, J.; Lu, C.; Wang, H.; Shen, Y.; Du, L. Alteramide B is a microtubule antagonist of inhibiting Candida albicans. Biochim. Biophys. Acta 2016, 1860, 2097–2106. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schrey, S.D.; Erkenbrack, E.; Früh, E.; Fengler, S.; Hommel, K.; Horlacher, N.; Schulz, D.; Ecke, M.; Kulik, A.; Fiedler, H.P.; et al. Production of fungal and bacterial growth modulating secondary metabolites is widespread among mycorrhiza-associated streptomycetes. BMC Microbiol. 2012, 12, 164. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Siegel, M.R.; Sisler, H.D.; Johnson, F. Relationship of structure to fungitoxicity of cycloheximide and related glutarimide derivatives. Biochem. Pharmacol. 1966, 15, 1213–1223. [Google Scholar] [CrossRef]
- Garcia, I.; Vior, N.M.; González-Sabín, J.; Braña, A.F.; Rohr, J.; Moris, F.; Méndez, C.; Salas, J.A. Engineering the biosynthesis of the polyketide-nonribosomal peptide collismycin A for generation of analogs with neuroprotective activity. Chem. Biol. 2013, 20, 1022–1032. [Google Scholar] [CrossRef] [Green Version]
- Ando, T.; Hirayama, K.; Takahashi, R.; Horino, I.; Etoh, Y.; Morioka, H.; Shibai, H.; Murai, A. Cosmomycin D, a new anthracycline antibiotic. Agric. Biol. Chem. 1985, 49, 259–262. [Google Scholar] [CrossRef]
- Jakobi, M.; Winkelmann, G.; Kaiser, D.; Kempler, C.; Jung, G.; Berg, G.; Bahl, H. Maltophilin: A new antifungal compound produced by Stenotrophomonas maltophilia R3089. J. Antibiot. 1996, 49, 1101–1104. [Google Scholar] [CrossRef] [Green Version]
- Borg, M.; Neary, J.M.; Wrigley, S.K.; Jim, J.M.; Smith, C. Cyclic Lipopeptides. PCT WO 2007/072082 Al. 2007. Available online: https://patentimages.storage.googleapis.com/a0/5d/e9/5b9d9d4b4674dc/WO2007072082A1.pdf (accessed on 16 September 2022).
- Cho, E.; Kwon, O.S.; Chung, B.; Lee, J.; Sun, J.; Shin, J.; Oh, K.B. Antibacterial activity of chromomycins from a marine-derived Streptomyces microflavus. Mar. Drugs. 2020, 18, 522. [Google Scholar] [CrossRef]
- Bertrand, S.; Bohni, N.; Schnee, S.; Schumpp, O.; Gindro, K.; Wolfender, J.L. Metabolite induction via microorganism co-culture: A potential way to enhance chemical diversity for drug discovery. Biotechnol. Adv. 2014, 32, 1180–1204. [Google Scholar] [CrossRef]
- Scherlach, K.; Hertweck, C. Chemical mediators at the bacterial-fungal interface. Annu. Rev. Microbiol. 2020, 74, 267–290. [Google Scholar] [CrossRef] [PubMed]
- Jones, S.E.; Elliot, M.A. Streptomyces exploration: Competition, volatile communication and new bacterial behaviours. Trends Microbiol. 2017, 25, 522–531. [Google Scholar] [CrossRef] [PubMed]





Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cuervo, L.; Méndez, C.; Salas, J.A.; Olano, C.; Malmierca, M.G. Volatile Compounds in Actinomycete Communities: A New Tool for Biosynthetic Gene Cluster Activation, Cooperative Growth Promotion, and Drug Discovery. Cells 2022, 11, 3510. https://doi.org/10.3390/cells11213510
Cuervo L, Méndez C, Salas JA, Olano C, Malmierca MG. Volatile Compounds in Actinomycete Communities: A New Tool for Biosynthetic Gene Cluster Activation, Cooperative Growth Promotion, and Drug Discovery. Cells. 2022; 11(21):3510. https://doi.org/10.3390/cells11213510
Chicago/Turabian StyleCuervo, Lorena, Carmen Méndez, José A. Salas, Carlos Olano, and Mónica G. Malmierca. 2022. "Volatile Compounds in Actinomycete Communities: A New Tool for Biosynthetic Gene Cluster Activation, Cooperative Growth Promotion, and Drug Discovery" Cells 11, no. 21: 3510. https://doi.org/10.3390/cells11213510