
Simultaneous Identification of Functional Antigen-Specific CD8+ and CD4+ Cells after In Vitro Expansion Using Elongated Peptides

 , , , and
, , , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. PBMC Isolation, Cell Freezing, and Thawing
2.2. Synthetic Peptides
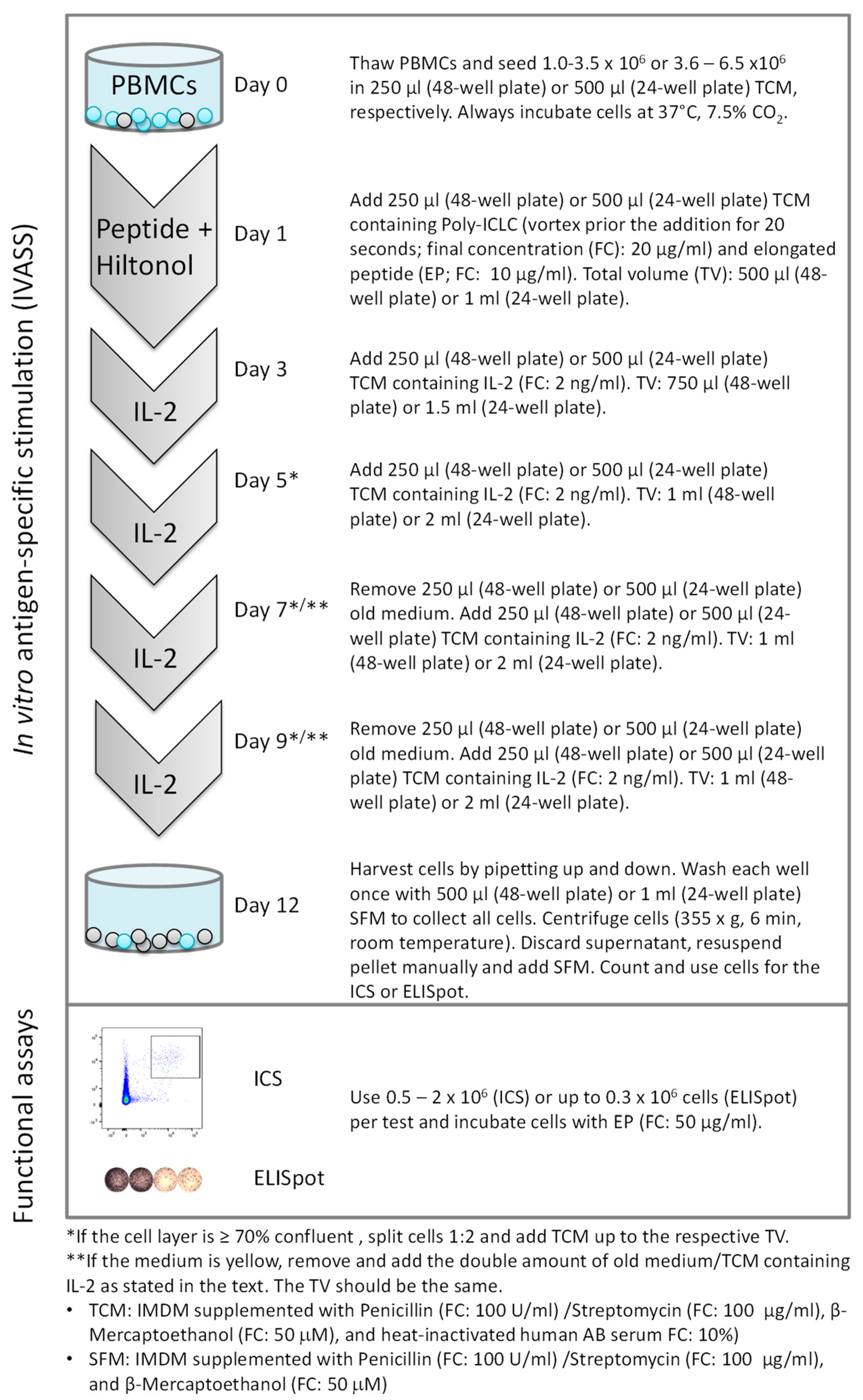
2.3. Twelve-Day In Vitro Stimulation of PBMCs
2.4. Intracellular Cytokine Staining Assay for T Cell Detection
2.5. Flow Cytometry Staining of Immune Cell Subsets
2.6. Enzyme-Linked Immunospot Assay
2.7. Statistics
3. Results
3.1. Ex Vivo Detection of Viral-Specific T Cells Using EPs Alone Can Be Suboptimal
3.2. The Addition of Poly-ICLC and GM-CSF Improves the Detection of Viral-Specific T Cells with EPs after 12 Days of In Vitro Expansion
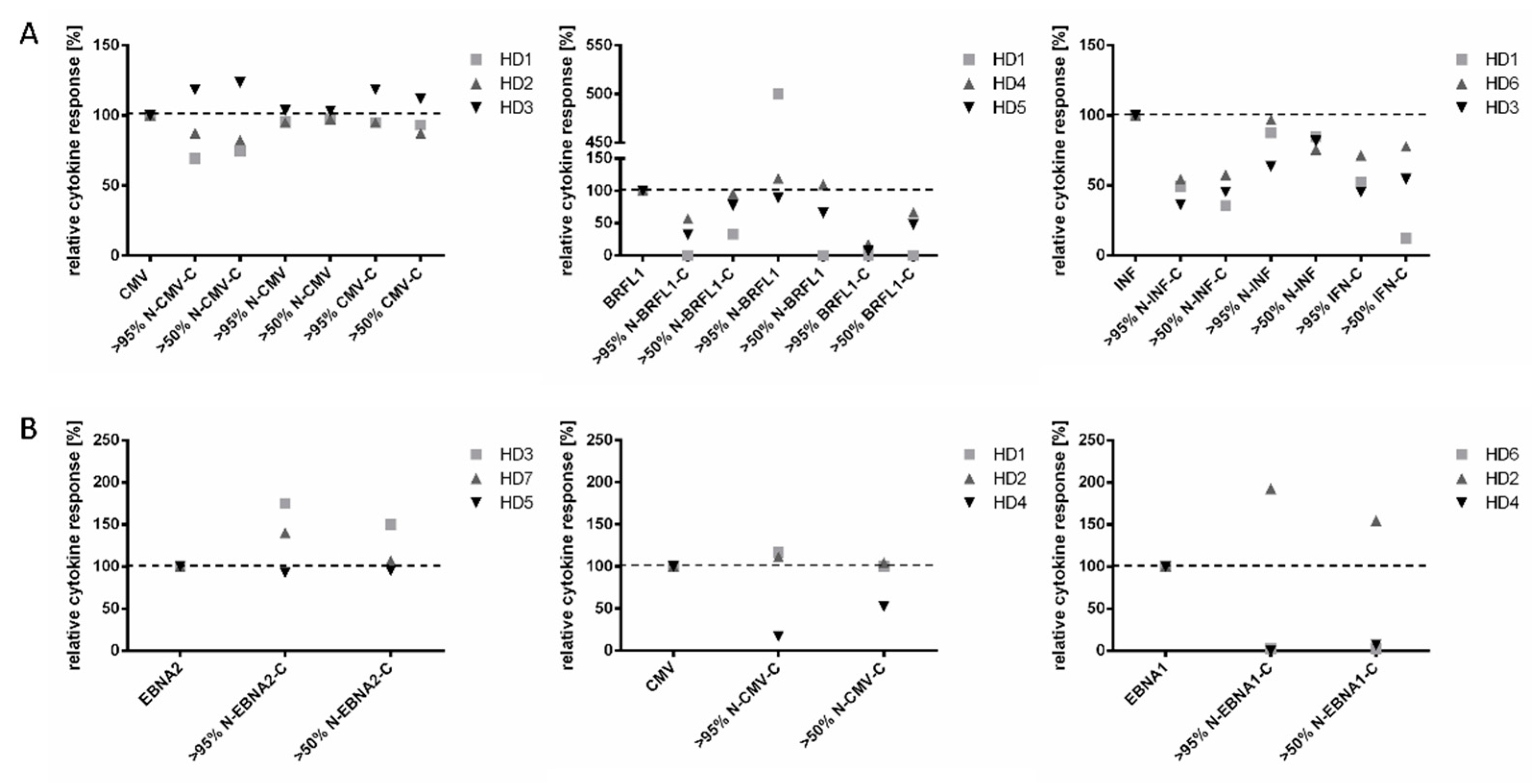
3.3. The Single Addition of Poly-ICLC Is Superior to the Combination of Poly-ICLC and GM-CSF for T Cell Amplification with EPs
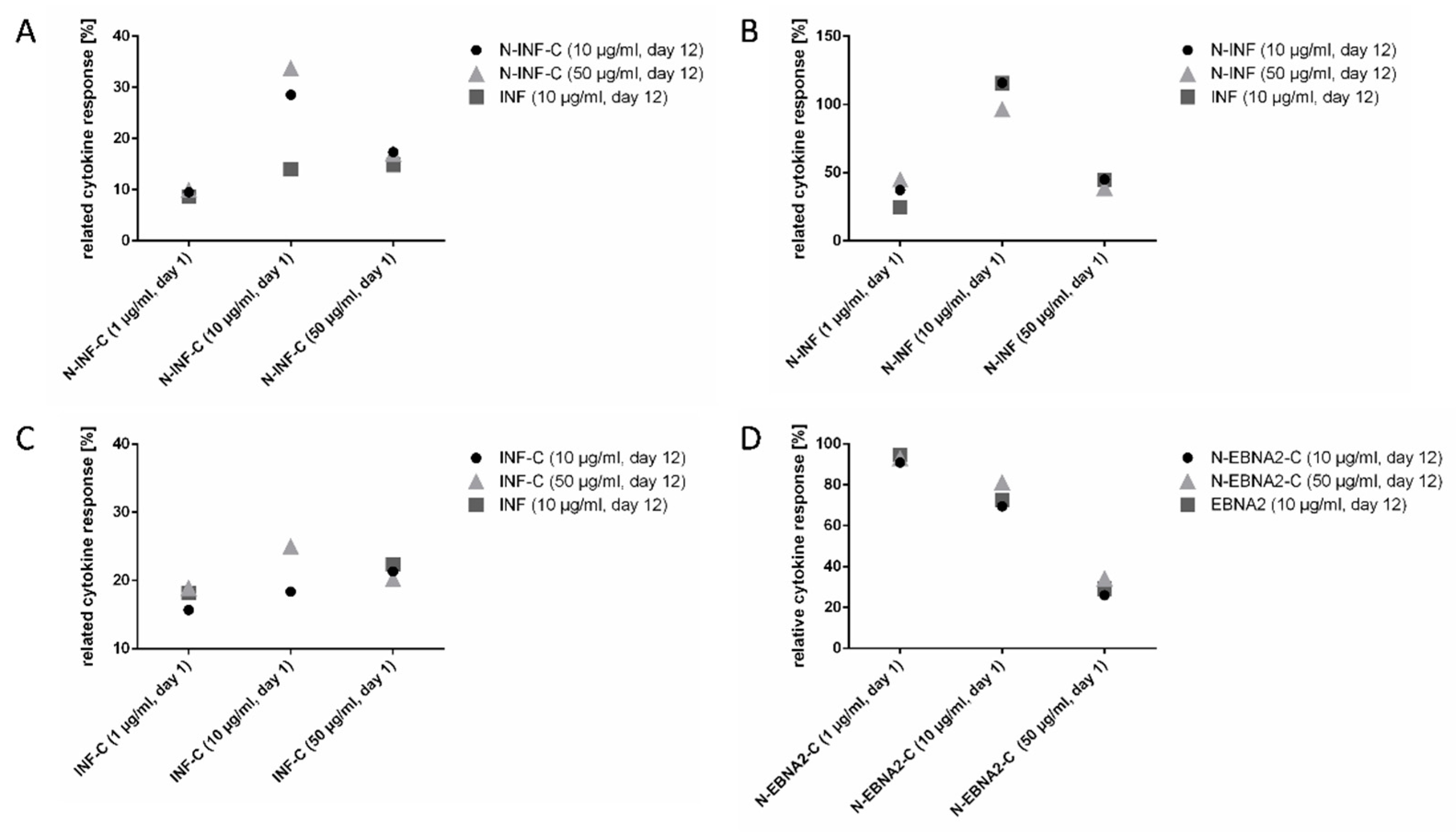
3.4. Optimal Peptide Concentration Finding for Simultaneous Read-Out of CD8+ and CD4+ T Cells with EPs
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Chudley, L.; McCann, K.J.; Coleman, A.; Cazaly, A.M.; Bidmon, N.; Britten, C.M.; van der Burg, S.H.; Gouttefangeas, C.; Jandus, C.; Laske, K.; et al. Harmonisation of Short-Term in Vitro Culture for the Expansion of Antigen-Specific CD8+ T Cells with Detection by ELISPOT and HLA-Multimer Staining. Cancer Immunol. Immunother. 2014, 63, 1199–1211. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Löffler, M.W.; Chandran, P.A.; Laske, K.; Schroeder, C.; Bonzheim, I.; Walzer, M.; Hilke, F.J.; Trautwein, N.; Kowalewski, D.J.; Schuster, H.; et al. Personalized Peptide Vaccine-Induced Immune Response Associated with Long-Term Survival of a Metastatic Cholangiocarcinoma Patient. J. Hepatol. 2016, 65, 849–855. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hilf, N.; Kuttruff-Coqui, S.; Frenzel, K.; Bukur, V.; Stevanović, S.; Gouttefangeas, C.; Platten, M.; Tabatabai, G.; Dutoit, V.; van der Burg, S.H.; et al. Actively Personalized Vaccination Trial for Newly Diagnosed Glioblastoma. Nature 2019, 565, 240–245. [Google Scholar] [CrossRef] [PubMed]
- Nelde, A.; Bilich, T.; Heitmann, J.S.; Maringer, Y.; Salih, H.R.; Roerden, M.; Lübke, M.; Bauer, J.; Rieth, J.; Wacker, M.; et al. SARS-CoV-2-Derived Peptides Define Heterologous and COVID-19-Induced T Cell Recognition. Nat. Immunol. 2021, 22, 74–85. [Google Scholar] [CrossRef]
- Rosalia, R.A.; Quakkelaar, E.D.; Redeker, A.; Khan, S.; Camps, M.; Drijfhout, J.W.; Silva, A.L.; Jiskoot, W.; van Hall, T.; van Veelen, P.A.; et al. Dendritic Cells Process Synthetic Long Peptides Better than Whole Protein, Improving Antigen Presentation and T-Cell Activation. Eur. J. Immunol. 2013, 43, 2554–2565. [Google Scholar] [CrossRef] [PubMed]
- Firbas, C.; Jilma, B.; Tauber, E.; Buerger, V.; Jelovcan, S.; Lingnau, K.; Buschle, M.; Frisch, J.; Klade, C.S. Immunogenicity and Safety of a Novel Therapeutic Hepatitis C Virus (HCV) Peptide Vaccine: A Randomized, Placebo Controlled Trial for Dose Optimization in 128 Healthy Subjects. Vaccine 2006, 24, 4343–4353. [Google Scholar] [CrossRef]
- Betts, M.R.; Ambrozak, D.R.; Douek, D.C.; Bonhoeffer, S.; Brenchley, J.M.; Casazza, J.P.; Koup, R.A.; Picker, L.J. Analysis of Total Human Immunodeficiency Virus (HIV)-Specific CD4(+) and CD8(+) T-Cell Responses: Relationship to Viral Load in Untreated HIV Infection. J. Virol. 2001, 75, 11983–11991. [Google Scholar] [CrossRef] [Green Version]
- Draenert, R.; Altfeld, M.; Brander, C.; Basgoz, N.; Corcoran, C.; Wurcel, A.G.; Stone, D.R.; Kalams, S.A.; Trocha, A.; Addo, M.M.; et al. Comparison of Overlapping Peptide Sets for Detection of Antiviral CD8 and CD4 T Cell Responses. J. Immunol. Methods 2003, 275, 19–29. [Google Scholar] [CrossRef]
- Maecker, H.T.; Dunn, H.S.; Suni, M.A.; Khatamzas, E.; Pitcher, C.J.; Bunde, T.; Persaud, N.; Trigona, W.; Fu, T.-M.; Sinclair, E.; et al. Use of Overlapping Peptide Mixtures as Antigens for Cytokine Flow Cytometry. J. Immunol. Methods 2001, 255, 27–40. [Google Scholar] [CrossRef]
- Grifoni, A.; Weiskopf, D.; Ramirez, S.I.; Mateus, J.; Dan, J.M.; Moderbacher, C.R.; Rawlings, S.A.; Sutherland, A.; Premkumar, L.; Jadi, R.S.; et al. Targets of T Cell Responses to SARS-CoV-2 Coronavirus in Humans with COVID-19 Disease and Unexposed Individuals. Cell 2020, 181, 1489–1501.e15. [Google Scholar] [CrossRef]
- Ott, P.A.; Hu, Z.; Keskin, D.B.; Shukla, S.A.; Sun, J.; Bozym, D.J.; Zhang, W.; Luoma, A.; Giobbie-Hurder, A.; Peter, L.; et al. An Immunogenic Personal Neoantigen Vaccine for Patients with Melanoma. Nature 2017, 547, 217–221. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kenter, G.G.; Welters, M.J.P.; Valentijn, A.R.P.M.; Lowik, M.J.G.; Berends-van der Meer, D.M.A.; Vloon, A.P.G.; Essahsah, F.; Fathers, L.M.; Offringa, R.; Drijfhout, J.W.; et al. Vaccination against HPV-16 Oncoproteins for Vulvar Intraepithelial Neoplasia. N. Engl. J. Med. 2009, 361, 1838–1847. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Melief, C.J.M.; van der Burg, S.H. Immunotherapy of Established (Pre)Malignant Disease by Synthetic Long Peptide Vaccines. Nat. Rev. Cancer 2008, 8, 351–360. [Google Scholar] [CrossRef] [PubMed]
- Knutson, K.L.; Schiffman, K.; Disis, M.L. Immunization with a HER-2/neu helper peptide vaccine generates HER-2/neu CD8 T-cell immunity in cancer patients. J. Clin. Investig. 2001, 107, 477–484. [Google Scholar] [CrossRef] [Green Version]
- Odunsi, K.; Qian, F.; Matsuzaki, J.; Mhawech-Fauceglia, P.; Andrews, C.; Hoffman, E.W.; Pan, L.; Ritter, G.; Villella, J.; Thomas, B.; et al. Vaccination with an NY-ESO-1 peptide of HLA class I/II specificities induces integrated humoral and T cell responses in ovarian cancer. Proc. Natl. Acad. Sci. USA 2007, 104, 12837–12842. [Google Scholar] [CrossRef] [Green Version]
- Gjertsen, M.K.; Bjørheim, J.; Saeterdal, I.; Myklebust, J.; Gaudernack, G. Cytotoxic CD4+ and CD8+ T Lymphocytes, Generated by Mutant P21-Ras (12Val) Peptide Vaccination of a Patient, Recognize 12Val-dependent nested epitopes present within the vaccine peptide and kill autologous tumor cells carrying this mutation. Int. J. Cancer 1997, 72, 784–790. [Google Scholar] [CrossRef]
- Kiecker, F.; Streitz, M.; Ay, B.; Cherepnev, G.; Volk, H.D.; Volkmer-Engert, R.; Kern, F. Analysis of Antigen-Specific T-Cell Responses with Synthetic Peptides—What Kind of Peptide for Which Purpose? Hum. Immunol. 2004, 65, 523–536. [Google Scholar] [CrossRef]
- Hoffmeister, B.; Kiecker, F.; Tesfa, L.; Volk, H.D.; Picker, L.J.; Kern, F. Mapping T Cell Epitopes by Flow Cytometry. Methods 2003, 29, 270–281. [Google Scholar] [CrossRef]
- Welters, M.J.P.; Kenter, G.G.; Piersma, S.J.; Vloon, A.P.G.; Löwik, M.J.G.; Berends-van Der Meer, D.M.A.; Drijfhout, J.W.; Valentijn, A.R.P.M.; Wafelman, A.R.; Oostendorp, J.; et al. Induction of Tumor-Specific CD4+ and CD8+ T-Cell Immunity in Cervical Cancer Patients by a Human Papillomavirus Type 16 E6 and E7 Long Peptides Vaccine. Clin. Cancer Res. 2008, 14, 178–187. [Google Scholar] [CrossRef] [Green Version]
- Singh, S.K.; Meyering, M.; Ramwadhdoebe, T.H.; Stynenbosch, L.F.M.; Redeker, A.; Kuppen, P.J.K.; Melief, C.J.M.; Welters, M.J.P.; van der Burg, S.H. The Simultaneous Ex Vivo Detection of Low-Frequency Antigen-Specific CD4+ and CD8+ T-Cell Responses Using Overlapping Peptide Pools. Cancer Immunol. Immunother. 2012, 61, 1953–1963. [Google Scholar] [CrossRef]
- Sabbatini, P.; Tsuji, T.; Ferran, L.; Ritter, E.; Sedrak, C.; Tuballes, K.; Jungbluth, A.A.; Ritter, G.; Aghajanian, C.; Bell-McGuinn, K.; et al. Phase I Trial of Overlapping Long Peptides from a Tumor Self-Antigen and Poly-ICLC Shows Rapid Induction of Integrated Immune Response in Ovarian Cancer Patients. Clin. Cancer Res. 2012, 18, 6497–6508. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Widenmeyer, M.; Griesemann, H.; Stevanović, S.; Feyerabend, S.; Klein, R.; Attig, S.; Hennenlotter, J.; Wernet, D.; Kuprash, D.V.; Sazykin, A.Y.; et al. Promiscuous Survivin Peptide Induces Robust CD4 + T-Cell Responses in the Majority of Vaccinated Cancer Patients. Int. J. Cancer 2012, 131, 140–149. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Löffler, M.W.; Nussbaum, B.; Jäger, G.; Jurmeister, P.S.; Budczies, J.; Pereira, P.L.; Clasen, S.; Kowalewski, D.J.; Mühlenbruch, L.; Königsrainer, I.; et al. A Non-Interventional Clinical Trial Assessing Immune Responses After Radiofrequency Ablation of Liver Metastases from Colorectal Cancer. Front. Immunol. 2019, 10, 2526. [Google Scholar] [CrossRef] [PubMed]
- Rammensee, H.G.; Gouttefangeas, C.; Heidu, S.; Klein, R.; Preuß, B.; Walz, J.S.; Nelde, A.; Haen, S.P.; Reth, M.; Yang, J.; et al. Designing a SARS-CoV-2 T-Cell-Inducing Vaccine for High-Risk Patient Groups. Vaccines 2021, 9, 428. [Google Scholar] [CrossRef] [PubMed]
- Slingluff, C.L.; Petroni, G.R.; Chianese-Bullock, K.A.; Smolkin, M.E.; Hibbitts, S.; Murphy, C.; Johansen, N.; Grosh, W.W.; Yamshchikov, G.v.; Neese, P.Y.; et al. Immunologic and Clinical Outcomes of a Randomized Phase II Trial of Two Multipeptide Vaccines for Melanoma in the Adjuvant Setting. Clin. Cancer Res. 2007, 13, 6386–6395. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zandvliet, M.L.; Kester, M.G.D.; van Liempt, E.; de Ru, A.H.; van Veelen, P.A.; Griffioen, M.; Guchelaar, H.J.; Falkenburg, J.H.F.; Meij, P. Efficiency and Mechanism of Antigen-Specific CD8 + T-Cell Activation Using Synthetic Long Peptides. J. Immunother. 2012, 35, 142–153. [Google Scholar] [CrossRef] [Green Version]
- Bilich, T.; Nelde, A.; Bichmann, L.; Roerden, M.; Salih, H.R.; Kowalewski, D.J.; Schuster, H.; Tsou, C.C.; Marcu, A.; Neidert, M.C.; et al. The HLA Ligandome Landscape of Chronic Myeloid Leukemia Delineates Novel T-Cell Epitopes for Immunotherapy. Blood 2019, 133, 550–565. [Google Scholar] [CrossRef] [Green Version]
- Schuster, H.; Peper, J.K.; Bösmüller, H.C.; Röhle, K.; Backert, L.; Bilich, T.; Ney, B.; Löffler, M.W.; Kowalewski, D.J.; Trautwein, N.; et al. The Immunopeptidomic Landscape of Ovarian Carcinomas. Proc. Natl. Acad. Sci. USA 2017, 114, E9942–E9951. [Google Scholar] [CrossRef] [Green Version]
- Caskey, M.; Lefebvre, F.; Filali-Mouhim, A.; Cameron, M.J.; Goulet, J.P.; Haddad, E.K.; Breton, G.; Trumpfheller, C.; Pollak, S.; Shimeliovich, I.; et al. Synthetic Double-Stranded RNA Induces Innate Immune Responses Similar to a Live Viral Vaccine in Humans. J. Exp. Med. 2011, 208, 2357–2366. [Google Scholar] [CrossRef] [Green Version]
- Gnjatic, S.; Atanackovic, D.; Matsuo, M.; Jäger, E.; Lee, S.Y.; Valmori, D.; Chen, Y.-T.; Ritter, G.; Knuth, A.; Old, L.J. Cross-Presentation of HLA Class I Epitopes from Exogenous NY-ESO-1 Polypeptides by Nonprofessional APCs. J. Immunol. 2003, 170, 1191–1196. [Google Scholar] [CrossRef]
- Dorner, M.; Brandt, S.; Tinguely, M.; Zucol, F.; Bourquin, J.P.; Zauner, L.; Berger, C.; Bernasconi, M.; Speck, R.F.; Nadal, D. Plasma Cell Toll-like Receptor (TLR) Expression Differs from That of B Cells, and Plasma Cell TLR Triggering Enhances Immunoglobulin Production. Immunology 2009, 128, 573–579. [Google Scholar] [CrossRef] [PubMed]
- Douagi, I.; Gujer, C.; Sundling, C.; Adams, W.C.; Smed-Sörensen, A.; Seder, R.A.; Karlsson Hedestam, G.B.; Loré, K. Human B Cell Responses to TLR Ligands Are Differentially Modulated by Myeloid and Plasmacytoid Dendritic Cells. J. Immunol. 2009, 182, 1991–2001. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- The Human Protein Atlas; Version 19.3; TLR3 RNA Expression Data in PBMCs; Knut and Alice Wallenberg Foundation: Stockholm, Sweden, 2009.
- Sercarz, E.E.; Maverakis, E. MHC-Guided Processing: Binding of Large Antigen Fragments. Nat. Rev. Immunol. 2003, 3, 621–629. [Google Scholar] [CrossRef] [PubMed]
- Schuhmacher, J.; Heidu, S.; Balchen, T.; Richardson, J.R.; Schmeltz, C.; Sonne, J.; Schweiker, J.; Rammensee, H.-G.; Thor Straten, P.; Røder, M.A.; et al. Vaccination against RhoC Induces Long-Lasting Immune Responses in Patients with Prostate Cancer: Results from a Phase I/II Clinical Trial. J. Immunother. Cancer 2020, 8, e001157. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Restriction | Source (Virus/Protein) | Peptide Length (aa) | Sequence * | Peptide Position (aa) in Source Protein | Elongation Site | Code | Purity>50% | Purity>95% |
|---|---|---|---|---|---|---|---|---|
| HLA class I (HLA-A*0201) | CMV pp65 strain AD169 Uniprot P06725 | 9 | NLVPMVATV | 495–503 | CMV | - | >95 | |
| 20 | GILARNLVPMVATVQGQNLK | 490–509 | N + C | N-CMV-C | 89 | 98 | ||
| 20 | WPPWQAGILARNLVPMVATV | 484–503 | N | N-CMV | 70 | 96 | ||
| 20 | NLVPMVATVQGQNLKYQEFF | 503–514 | C | CMV-C | 67 | 98 | ||
| Matrix Influenza strain A/Puerto Rico/8/1934 H1N1 Uniprot P03485 | 9 | GILGFVFTL | 58–66 | - | INF | - | 100 | |
| 20 | SPLTKGILGFVFTLTVPSER | 53–72 | N + C | N-INF-C | 78 | 90 | ||
| 20 | KTRPILSPLTKGILGFVFTL | 47–66 | N | N-INF | 66 | 98 | ||
| 20 | GILGFVFTLTVPSERGLQRR | 53–77 | C | INF-C | 63 | 98 | ||
| EBV BRFL1 strain B95-8 Uniport P03209 | 9 | YVLDHLIVV | 109–117 | - | BRFL1 | 86 | - | |
| 20 | PIVMRYVLDHLIVVTDRFFI | 104–123 | N + C | N-BRFL1-C | 53 | 98 | ||
| 20 | ACSIACPIVMRYVLDHLIVV | 98–117 | N | N-BRFL1 | 83 | 95 | ||
| 20 | YVLDHLIVVTDRFFIQAPSN | 109–128 | C | BRFL1-C | 52 | 96 | ||
| HLA class II | CMV pp65 strain AD169 Uniprot P06725-1 | 15 | YQEFFWDANDIYRIF | 510–524 | - | CMV | - | 95 |
| 20 | NLKYQEFFWDANDIYRIFAE | 507–526 | N + C | N-CMV-C | 74 | 98 | ||
| EBV EBNA2 strain B95-8 Uniprot P12978 | 15 | PRSPTVFYNIPPMPL | 276–290 | - | EBNA2 | 81 | - | |
| 20 | SPEPRSPTVFYNIPPMPLPP | 273–292 | N + C | N-EBNA2-C | 60 | 99 | ||
| EBV EBNA1 strain B95-8 Uniprot P03211 | 14 | KTSLYNLRRGTALA | 514–527 | - | EBNA1 | 72 | - | |
| 20 | GGSKTSLYNLRRGTALAIPQ | 511–530 | N + C | N-EBNA1-C | 89 | 96 |
| Staining Step | Marker | Fluorochrome | Clone | Manufacturer | Cat. No. |
|---|---|---|---|---|---|
| Extracellular staining | Live/Dead | Zombie Aqua | - | BioLegend | 423102 |
| CD4 | APC-Cy7 | RPA-T4 | BD | 557871 | |
| CD8 | BV605 | RPA-T8 | BioLegend | 301040 | |
| Intracellular staining | TNF | Pacific Blue | MAB11 | BioLegend | 502920 |
| IFN-γ | FITC | B27 | BD | 554700 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Schuhmacher, J.; Kleemann, L.; Richardson, J.R.; Rusch, E.; Rammensee, H.-G.; Gouttefangeas, C. Simultaneous Identification of Functional Antigen-Specific CD8+ and CD4+ Cells after In Vitro Expansion Using Elongated Peptides. Cells 2022, 11, 3451. https://doi.org/10.3390/cells11213451
Schuhmacher J, Kleemann L, Richardson JR, Rusch E, Rammensee H-G, Gouttefangeas C. Simultaneous Identification of Functional Antigen-Specific CD8+ and CD4+ Cells after In Vitro Expansion Using Elongated Peptides. Cells. 2022; 11(21):3451. https://doi.org/10.3390/cells11213451
Chicago/Turabian StyleSchuhmacher, Juliane, Leon Kleemann, Jennifer Rebecca Richardson, Elisa Rusch, Hans-Georg Rammensee, and Cécile Gouttefangeas. 2022. "Simultaneous Identification of Functional Antigen-Specific CD8+ and CD4+ Cells after In Vitro Expansion Using Elongated Peptides" Cells 11, no. 21: 3451. https://doi.org/10.3390/cells11213451