
Precision Phenotyping of Nectar-Related Traits Using X-ray Micro Computed Tomography

 , ,
, , 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
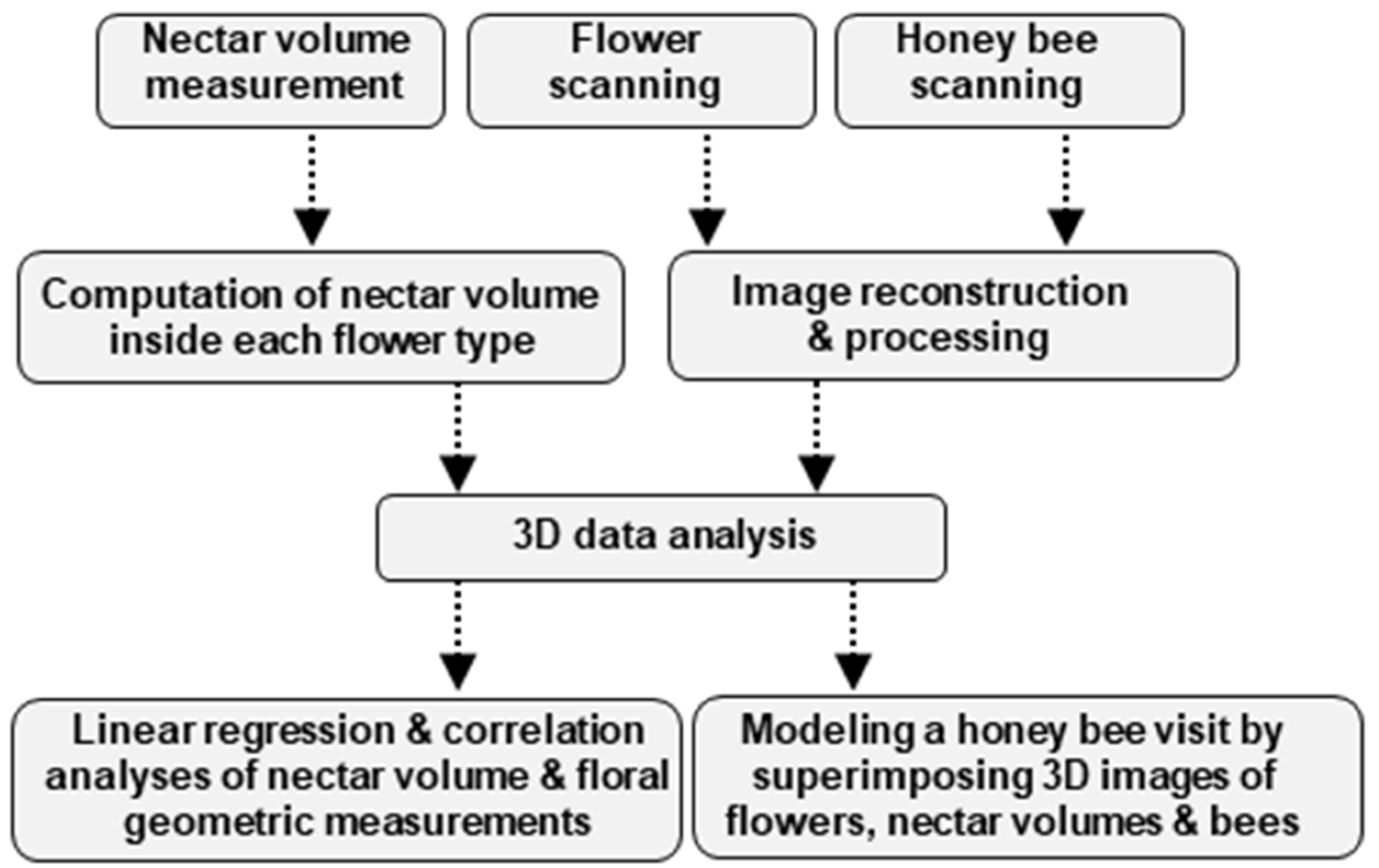
2. Materials and Methods
2.1. Plant Material and Sampling
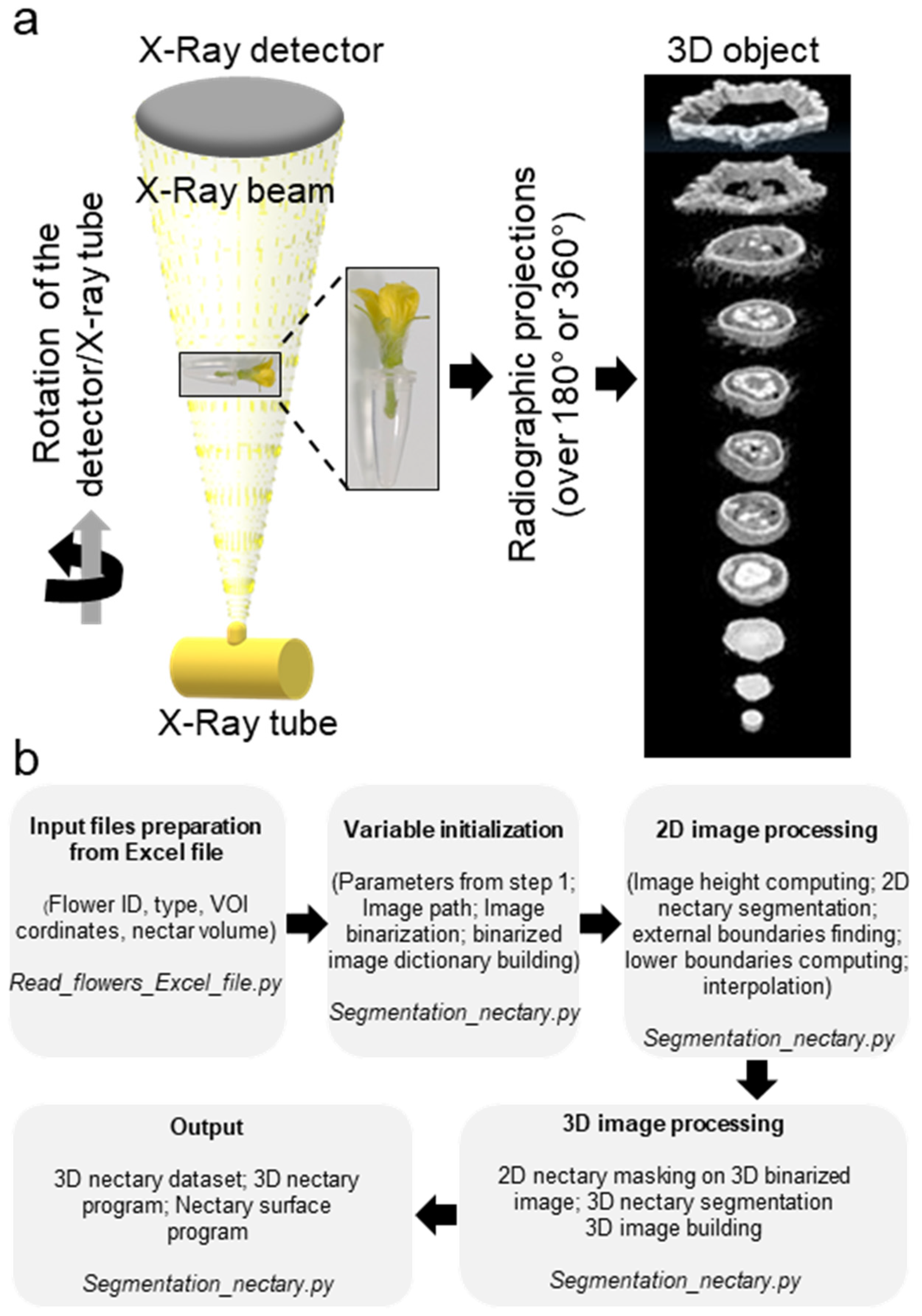
2.2. Micro-CT Scanning Conditions
2.3. Reconstruction of 3D Images
2.4. Honey Bee Preparation
2.5. Nectary Cross-Sectional Area Measurements
2.6. Computation of Nectary Surface and Volume
2.7. Nectar Volume Measurements
2.8. Statistical Analysis
3. Results
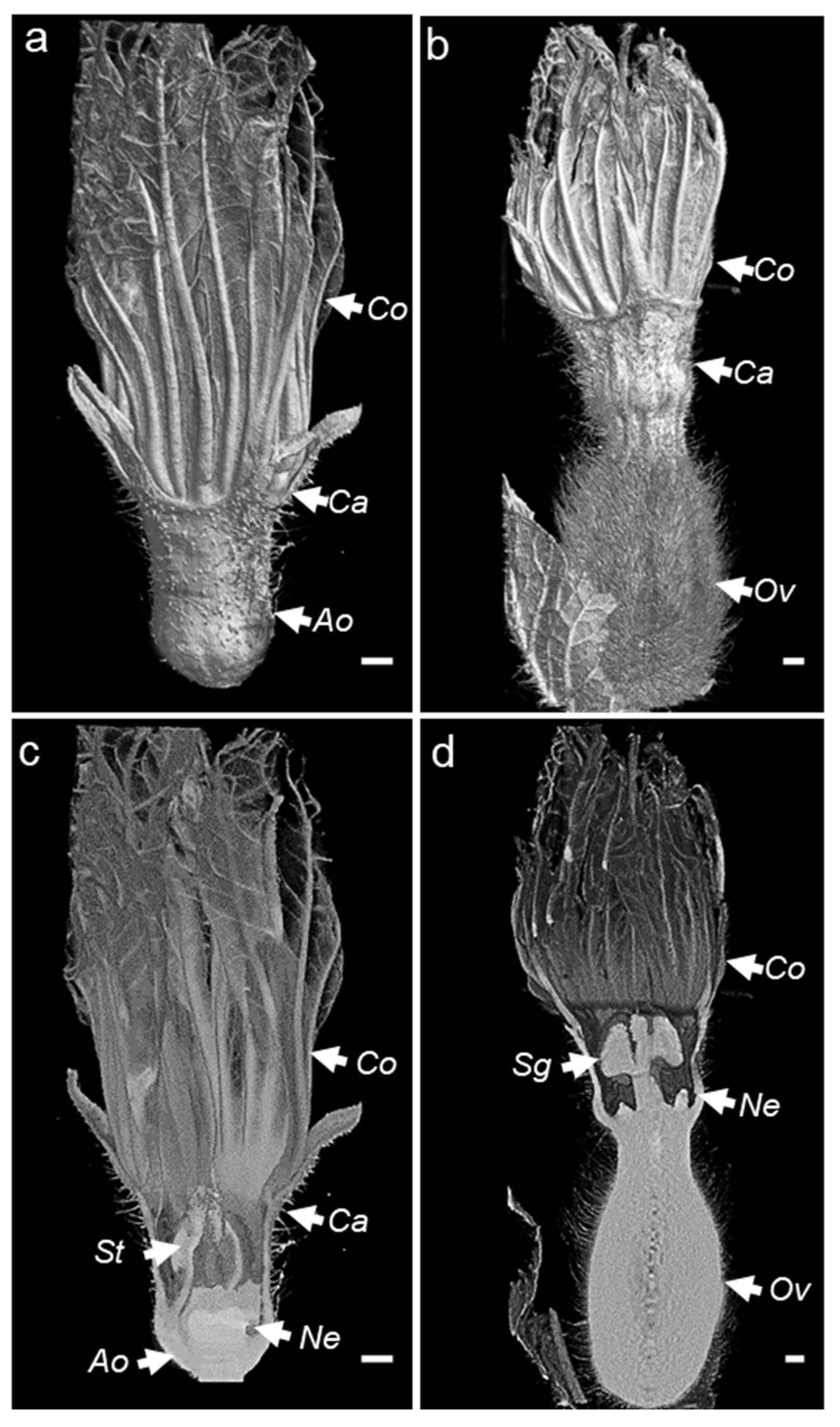
3.1. Imager and Imaging Melon Flowers
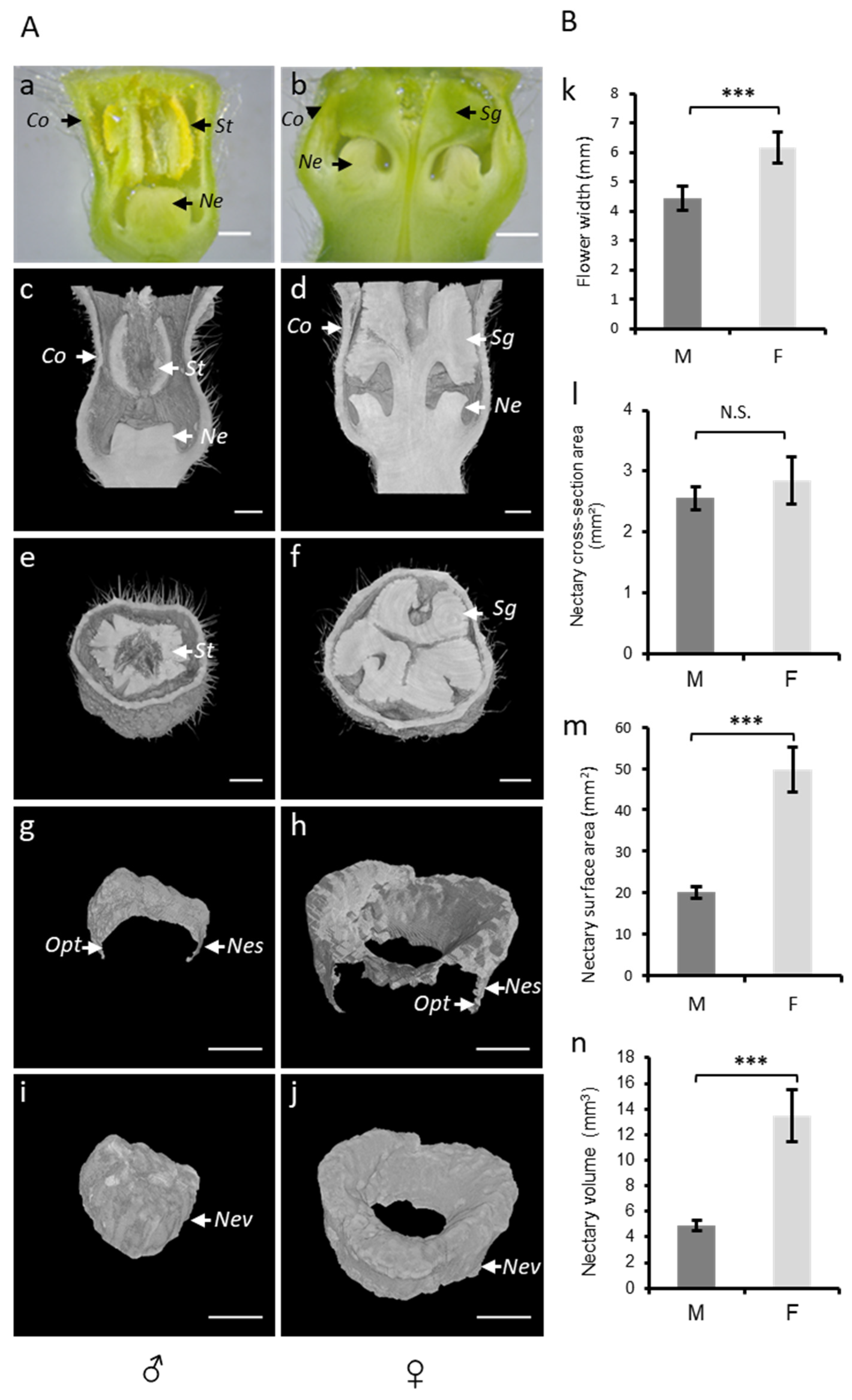
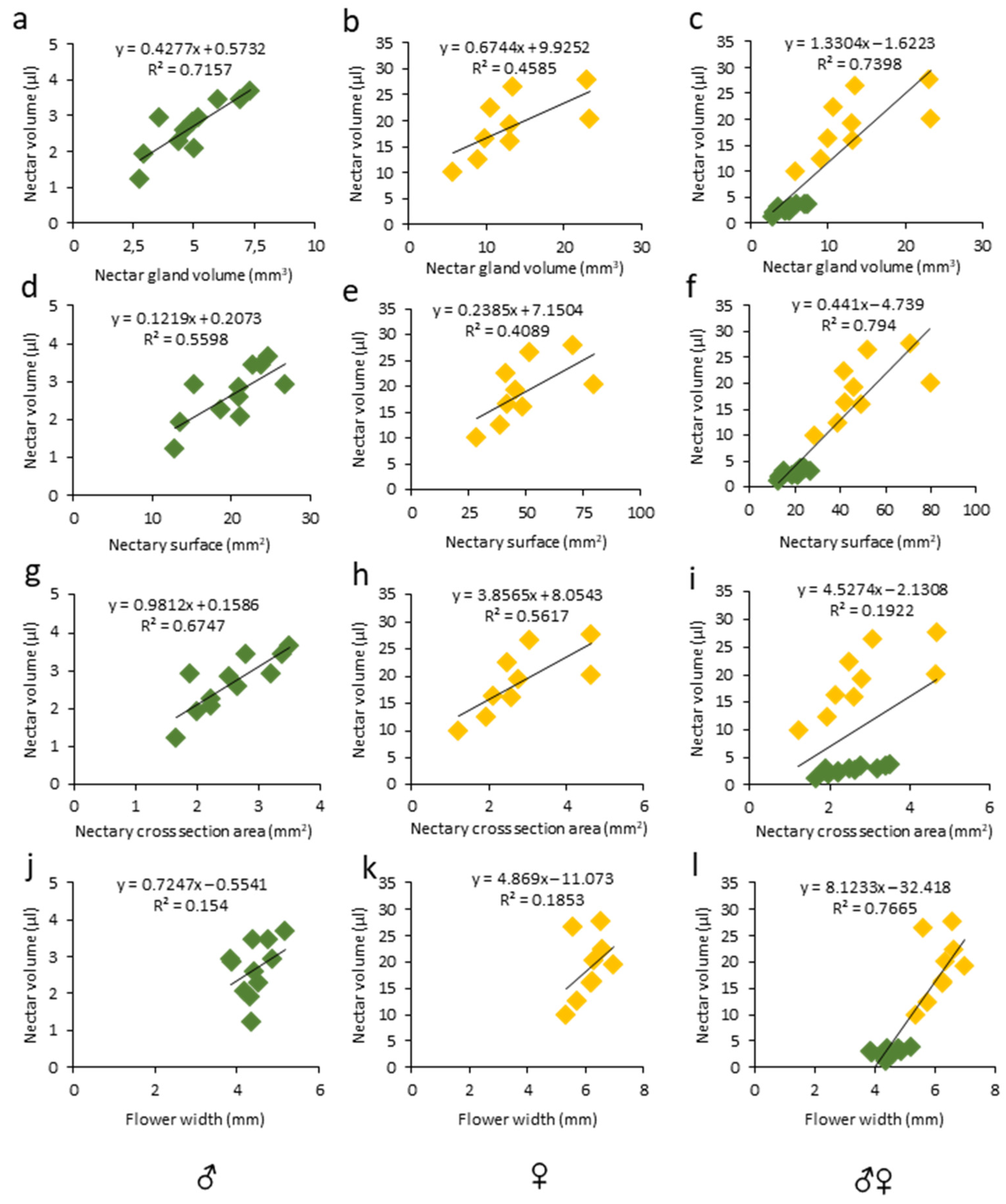
3.2. Phenotyping Nectar-Related Traits Using Micro-CT
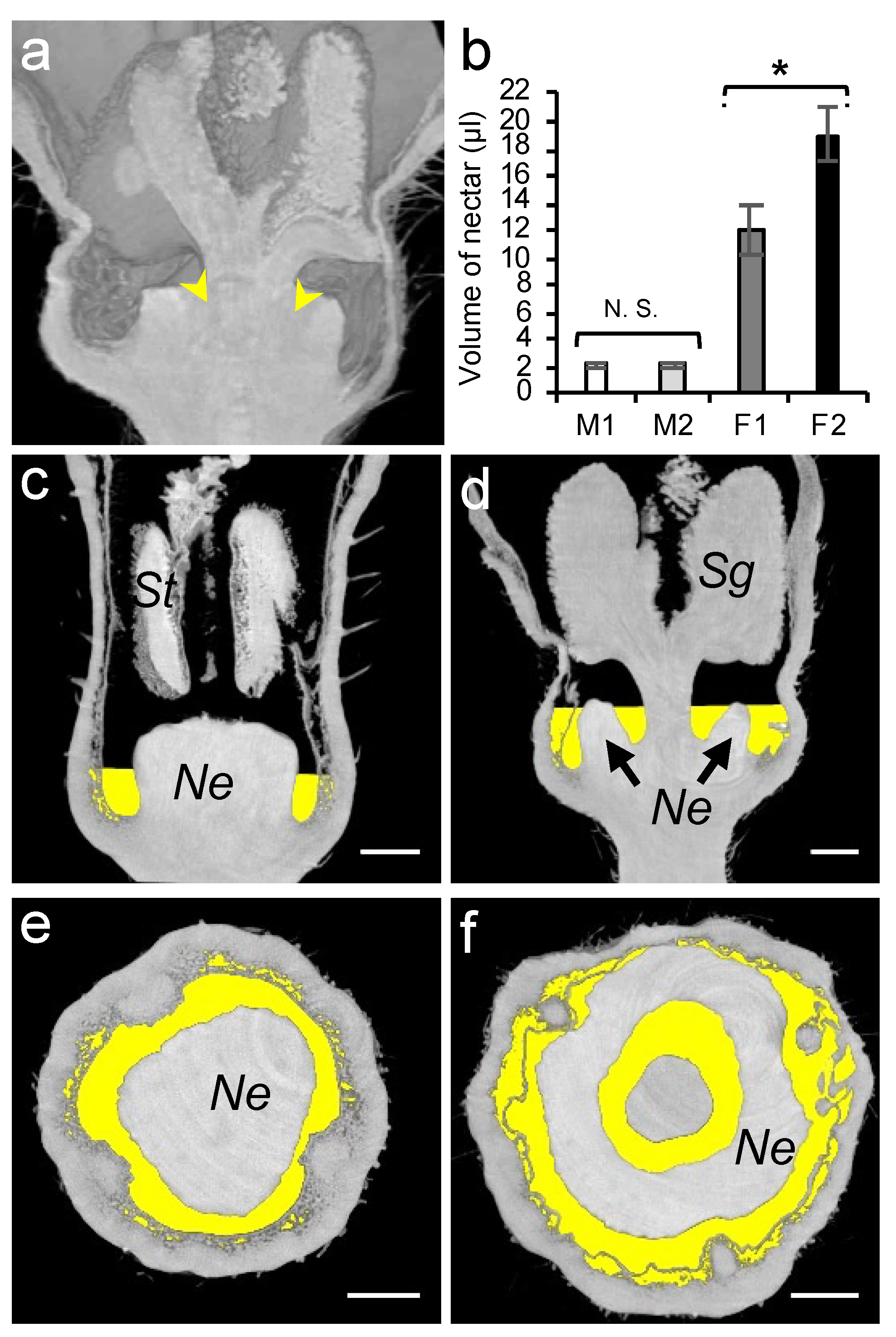
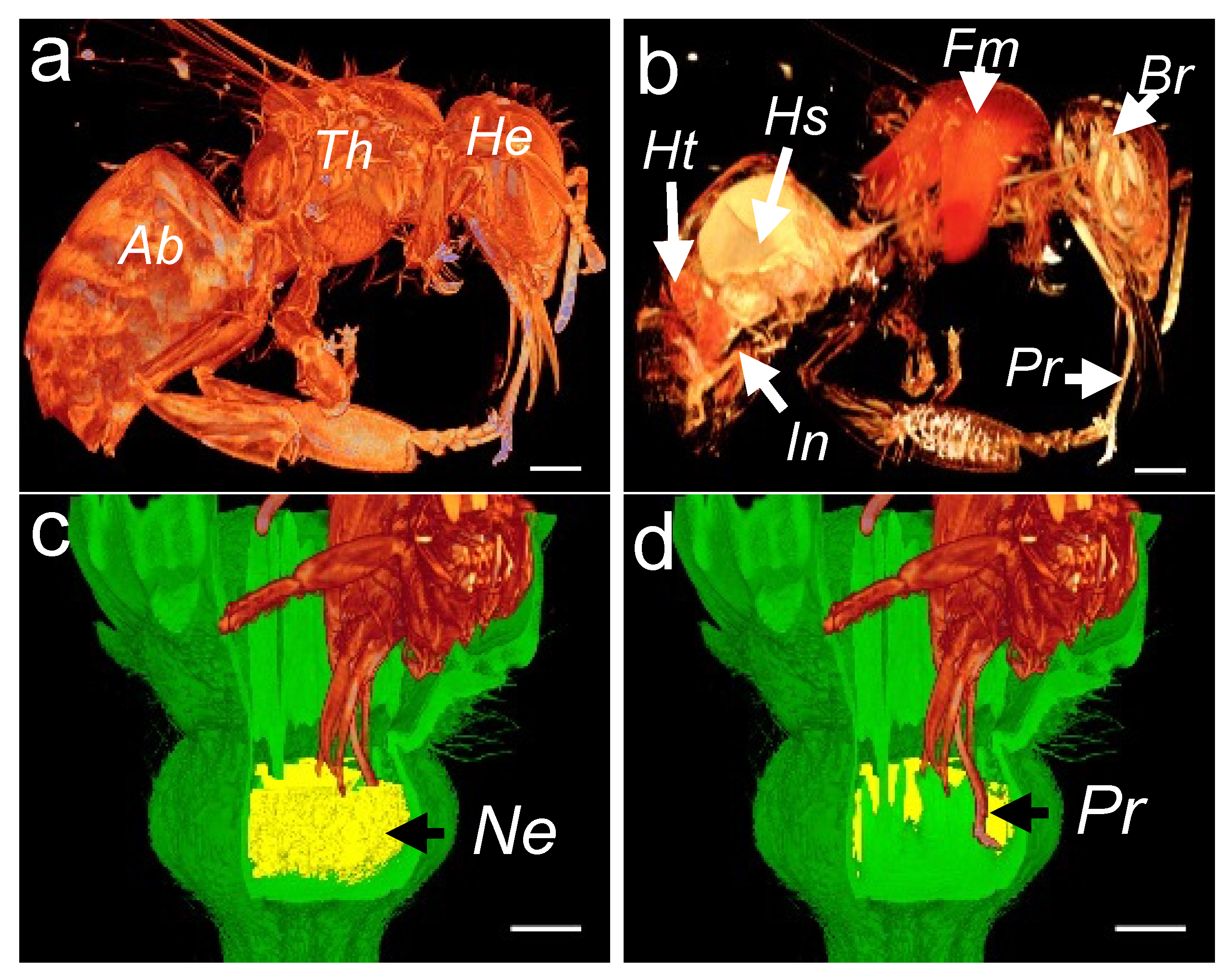
3.3. Assessment of Nectar Accessibility to Honey Bees
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Borghi, M.; Fernie, A.R.; Schiestl, F.P.; Bouwmeester, H.J. The Sexual Advantage of Looking, Smelling, and Tasting Good: The Metabolic Network That Produces Signals for Pollinators. Trends Plant Sci. 2017, 22, 338–350. [Google Scholar] [CrossRef] [PubMed]
- Muhlemann, J.K.; Klempien, A.; Dudareva, N. Floral volatiles: From biosynthesis to function: Floral volatiles. Plant Cell Environ. 2014, 37, 1936–1949. [Google Scholar] [CrossRef] [PubMed]
- Hermann, K.; Kuhlemeier, C. The genetic architecture of natural variation in flower morphology. Curr. Opin. Plant Biol. 2011, 14, 60–65. [Google Scholar] [CrossRef] [PubMed]
- Dorado, J.; Vázquez, D.P. The diversity-stability relationship in floral production. Oikos 2014, 123, 1137–1143. [Google Scholar] [CrossRef]
- Kiill, L.H.P.; Feitoza, E.D.A.; Siqueira, K.M.M.D.; Ribeiro, M.D.F.; Silva, E.M.S.D. Evaluation of Floral Characteristics of Melon Hybrids (Cucumis Melo L.) in Pollinator Attractiveness. Rev. Bras. Frutic. 2016, 38, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Alexandersson, R.; Johnson, S.D. Pollinator–mediated selection on flower–tube length in a hawkmoth–pollinated Gladiolus (Iridaceae). Proc. R. Soc. Lond. B 2002, 269, 631–636. [Google Scholar] [CrossRef] [Green Version]
- Slavković, F.; Dogimont, C.; Morin, H.; Boualem, A.; Bendahmane, A. The Genetic Control of Nectary Development. Trends Plant Sci. 2021, 26, 260–271. [Google Scholar] [CrossRef]
- Ren, G.; Healy, R.A.; Klyne, A.M.; Horner, H.T.; James, M.G.; Thornburg, R.W. Transient starch metabolism in ornamental tobacco floral nectaries regulates nectar composition and release. Plant Sci. 2007, 173, 277–290. [Google Scholar] [CrossRef]
- Davis, A.R. Searching and Breeding for Structural Features of Flowers Correlated with High Nectar-Carbohydrate Production. Acta Hortic. 2001, 561, 107–122. [Google Scholar] [CrossRef]
- Shuel, R.W. Improving Honey Production through Plant Breeding. Bee World 1989, 70, 36–45. [Google Scholar] [CrossRef]
- Teuber, L.R.; Albertsen, M.C.; Barnes, D.K.; Heichel, G.H. Structure of Floral Nectaries of Alfalfa (Medicago Sativa L.) in Relation to Nectar Production. Am. J. Bot. 1980, 67, 433–439. [Google Scholar] [CrossRef]
- Davis, A.R.; Sawhney, V.K.; Fowke, L.C.; Low, N.H. Floral nectar secretion and ploidy in Brassica rapa and B napus (Brassicaceae). I. Nectary size and nectar carbohydrate production and composition. Apidologie 1994, 25, 602–614. [Google Scholar] [CrossRef] [Green Version]
- Wessinger, C.A.; Hileman, L.C.; Rausher, M.D. Identification of major quantitative trait loci underlying floral pollination syndrome divergence in Penstemon. Phil. Trans. R. Soc. B 2014, 369, 20130349. [Google Scholar] [CrossRef] [Green Version]
- Nepi, M.; Guarnieri, M.; Pacini, E. Nectar Secretion, Reabsorption, and Sugar Composition in Male and Female Flowers of Cucurbita pepo. Int. J. Plant Sci. 2001, 162, 353–358. [Google Scholar] [CrossRef]
- Nepi, M.; Cresti, L.; Guarnieri, M.; Pacini, E. Dynamics of Nectar Production and Nectar Homeostasis in Male Flowers of Cucurbita pepo L. Int. J. Plant Sci. 2011, 172, 183–190. [Google Scholar] [CrossRef]
- Bailes, E.J.; Ollerton, J.; Pattrick, J.G.; Glover, B.J. How can an understanding of plant–pollinator interactions contribute to global food security? Curr. Opin. Plant Biol. 2015, 26, 72–79. [Google Scholar] [CrossRef] [Green Version]
- Borghi, M.; Fernie, A.R. Outstanding questions in flower metabolism. Plant J. 2020, 103, 1275–1288. [Google Scholar] [CrossRef]
- Miller, J.S.; Venable, D.L. Floral Morphometrics and the Evolution of Sexual Dimorphism in Lycium (Solanaceae). Evolution 2003, 57, 74–86. [Google Scholar] [CrossRef]
- Pérez, R.; Vargas, P.; Arroyo, J. Convergent evolution of flower polymorphism in Narcissus (Amaryllidaceae). New Phytol. 2004, 161, 235–252. [Google Scholar] [CrossRef] [Green Version]
- Kobayashi, K.; Horisaki, A.; Niikura, S.; Ohsawa, R. Diallel analysis of floral morphology in radish (Raphanus sativus L.). Euphytica 2007, 158, 153–165. [Google Scholar] [CrossRef]
- Fernández-Mazuecos, M.; Blanco-Pastor, J.L.; Gómez, J.M.; Vargas, P. Corolla morphology influences diversification rates in bifid toadflaxes (Linaria sect. Versicolores). Ann. Bot. 2013, 112, 1705–1722. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Berger, B.A.; Ricigliano, V.A.; Savriama, Y.; Lim, A.; Thompson, V.; Howarth, D.G. Geometric morphometrics reveals shifts in flower shape symmetry and size following gene knockdown of CYCLOIDEA and ANTHOCYANIDIN SYNTHASE. BMC Plant Biol. 2017, 17, 205. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, C.-N.; Hsu, H.-C.; Wang, C.-C.; Lee, T.-K.; Kuo, Y.-F. Quantifying floral shape variation in 3D using microcomputed tomography: A case study of a hybrid line between actinomorphic and zygomorphic flowers. Front. Plant Sci. 2015, 6, 724. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Deyneko, I.V.; Kel, A.; Bloecker, H.; Kauer, G. Signal-theoretical DNA similarity measure revealing unexpected similarities of E. coli promoters. Silico Biol. 2005, 51, 547–555. [Google Scholar]
- Kuhl, F.P.; Giardina, C.R. Elliptic Fourier features of a closed contour. Comput. Graph. Image Process. 1982, 18, 236–258. [Google Scholar] [CrossRef]
- Parekh, H.; Thakore, D.; Jaliya, U. A survey on object detection and tracking methods. Int. J. Innov. Res. Comput. Commun. Eng. 2014, 2, 2970–2979. [Google Scholar]
- Adams, D.C.; Rohlf, F.J.; Slice, D.E. Geometric morphometrics: Ten years of progress following the “revolution”. Ital. J. Zool. 2004, 71, 5–16. [Google Scholar] [CrossRef] [Green Version]
- Zelditch, M.; Swiderski, D.; Sheets, H. Geometric Morphometrics for Biologists: A Primer; Academic Press Cambridge: Cambridge, UK, 2012. [Google Scholar]
- Rozov, S.M.; Deineko, E.V.; Deyneko, I.V. FlowerMorphology: Fully automatic flower morphometry software. Planta 2018, 247, 1163–1173. [Google Scholar] [CrossRef]
- Russ, J. The Image Processing Handbook, 5th ed.; CRC Press: Boca Raton, FL, USA, 2007. [Google Scholar]
- Dhondt, S.; Vanhaeren, H.; van Loo, D.; Cnudde, V.; Inzé, D. Plant structure visualization by high-resolution X-ray computed tomography. Trends Plant Sci. 2010, 15, 419–422. [Google Scholar] [CrossRef]
- Dhondt, S.; Wuyts, N.; Inzé, D. Cell to whole-plant phenotyping: The best is yet to come. Trends Plant Sci. 2013, 18, 428–439. [Google Scholar] [CrossRef]
- Van der Niet, T.; Zollikofer, C.P.E.; de León, M.S.P.; Johnson, S.D.; Linder, H.P. Three-dimensional geometric morphometrics for studying floral shape variation. Trends Plant Sci. 2010, 15, 423–426. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ijiri, T.; Yoshizawa, S.; Yokota, H.; Igarashi, T. Flower modeling via X-ray computed tomography. ACM Trans. Graph. 2014, 33, 1–10. [Google Scholar] [CrossRef]
- Tracy, S.R.; Gómez, J.F.; Sturrock, C.J.; Wilson, Z.A.; Ferguson, A.C. Non-destructive determination of floral staging in cereals using X-ray micro computed tomography (µCT). Plant Methods 2017, 13, 9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Staedler, Y.M.; Kreisberger, T.; Manafzadeh, S.; Chartier, M.; Handschuh, S.; Pamperl, S.; Sontag, S.; Paun, O.; Schönenberger, J. Novel computed tomography-based tools reliably quantify plant reproductive investment. J. Exp. Bot. 2018, 69, 525–535. [Google Scholar] [CrossRef] [PubMed]
- Bellaire, A.; Ischebeck, T.; Staedler, Y.; Weinhaeuser, I.; Mair, A.; Parameswaran, S.; Ito, T.; Schönenberger, J.; Weckwerth, W. Metabolism and development—Integration of micro computed tomography data and metabolite profiling reveals metabolic reprogramming from floral initiation to silique development. New Phytol. 2014, 202, 322–335. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Steppe, K.; Cnudde, V.; Girard, C.; Lemeur, R.; Cnudde, J.-P.; Jacobs, P. Use of X-ray computed microtomography for non-invasive determination of wood anatomical characteristics. J. Struct. Biol. 2004, 148, 11–21. [Google Scholar] [CrossRef]
- Kaminuma, E.; Heida, N.; Tsumoto, Y.; Yamamoto, N.; Goto, N.; Okamoto, N.; Konagaya, A.; Matsui, M.; Toyoda, T. Automatic quantification of morphological traits via three-dimensional measurement of Arabidopsis. Plant J. 2004, 38, 358–365. [Google Scholar] [CrossRef]
- Cloetens, P.; Mache, R.; Schlenker, M.; Lerbs-Mache, S. Quantitative phase tomography of Arabidopsis seeds reveals intercellular void network. Proc. Natl. Acad. Sci. USA 2006, 103, 14626–14630. [Google Scholar] [CrossRef] [Green Version]
- Verboven, P.; Kerckhofs, G.; Mebatsion, H.K.; Ho, Q.T.; Temst, K.; Wevers, M.; Cloetens, P.; Nicolaï, B.M. Three-Dimensional Gas Exchange Pathways in Pome Fruit Characterized by Synchrotron X-ray Computed Tomography. Plant Physiol. 2008, 147, 518–527. [Google Scholar] [CrossRef] [Green Version]
- Staedler, Y.M.; Masson, D.; Schönenberger, J. Plant Tissues in 3D via X-ray Tomography: Simple Contrasting Methods Allow High Resolution Imaging. PLoS ONE 2013, 8, e75295. [Google Scholar] [CrossRef] [Green Version]
- Das, R.; Yadav, R.N.; Sihota, P.; Uniyal, P.; Kumar, N.; Bhushan, B. Biomechanical Evaluation of Wasp and Honeybee Stingers. Sci. Rep. 2018, 8, 14945. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Facchini, E.; Nalon, L.; Andreis, M.E.; di Giancamillo, M.; Rizzi, R.; Mortarino, M. Honeybee pupal length assessed by CT-scan technique: Effects of Varroa infestation, developmental stage and spatial position within the brood comb. Sci. Rep. 2019, 9, 10614. [Google Scholar] [CrossRef] [PubMed]
- Pichot, C.; Djari, A.; Tran, J.; Verdenaud, M.; Marande, W.; Huneau, C.; Gautier, V.; Latrasse, D.; Arribat, S.; Sommard, V.; et al. Cantaloupe melon genome reveals 3D chromatin features and structural relationship with the ancestral cucurbitaceae karyotype. iScience 2022, 25, 103696. [Google Scholar] [CrossRef] [PubMed]
- Boualem, A.; Troadec, C.; Camps, C.; Lemhemdi, A.; Morin, H.; Sari, M.-A.; Fraenkel-Zagouri, R.; Kovalski, I.; Dogimont, C.; Perl-Treves, R.; et al. A cucurbit androecy gene reveals how unisexual flowers develop and dioecy emerges. Science 2015, 350, 688–691. [Google Scholar] [CrossRef]
- Klein, A.-M.; Vaissière, B.E.; Cane, J.H.; Steffan-Dewenter, I.; Cunningham, S.A.; Kremen, C.; Tscharntke, T. Importance of pollinators in changing landscapes for world crops. Proc. R. Soc. B Biol. Sci. 2007, 274, 303–313. [Google Scholar] [CrossRef] [Green Version]
- Ondruš, J.; Hubatka, F.; Kulich, P.; Odehnalová, N.; Harabiš, V.; Hesko, B.; Sychra, O.; Široký, P.; Turánek, J.; Novobilský, A. A novel approach to imaging engorged ticks: Micro-CT scanning of Ixodes ricinus fed on blood enriched with gold nanoparticles. Ticks Tick-Borne Dis. 2021, 12, 101559. [Google Scholar] [CrossRef]
- Liu, C.-N.; Morin, J.; Dokmanovich, M.; Bluette, C.T.; Goldstein, R.; Manickam, B.; Bagi, C.M. Nanoparticle contrast-enhanced micro-CT: A preclinical tool for the 3D imaging of liver and spleen in longitudinal mouse studies. J. Pharmacol. Toxicol. Methods 2019, 96, 67–77. [Google Scholar] [CrossRef]
- Korte, N.; Porembski, S. Anatomical analysis of turgescent and semi-dry resurrection plants: The effect of sample preparation on the sample, resolution, and image quality of X-ray micro-computed tomography (μCT). Microsc. Res. Tech. 2011, 74, 364–369. [Google Scholar] [CrossRef]
- Tercedor, J.; Alba-Alejandre, I. Comparing micro-CT results insects with classical anatomical studies: The europan honey bee (Apis mellifera Linnaeus, 1758) as a benchmark (Insecta: Hymenoptera, Apidae). Microscopy 2017. Available online: https://analyticalscience.wiley.com/do/10.1002/micro.2789 (accessed on 22 September 2022).
- Schoborg, T.A.; Smith, S.L.; Smith, L.N.; Morris, H.D.; Rusan, N.M. Micro-computed tomography as a platform for exploring Drosophila development. Development 2019, 146, dev.176685. [Google Scholar] [CrossRef]
- Beutler, R.; Schöntag, A. Über die Nektarabscheidung einiger Nutzpflanzen. Z. Vergl. Physiol. 1940, 28, 254–285. [Google Scholar] [CrossRef]
- Jablonski, B. Notes on the method to investigate nectar secretion rate in flowers. J. Apic. Sci. 2002, 46, 117–125. [Google Scholar]
- Dafni, H.; Lensky, Y.; Fahn, A. Flower and Nectar Characteristics of Nine Species of Labiatae and their influence on Honeybee Visits. J. Apic. Res. 1988, 27, 103–114. [Google Scholar] [CrossRef]
- Teuber, L.R.; Barnes, D.K.; Rincker, C.M. Effectiveness of selection for nectar volume, receptacle diameter, and seed yield characteristics in alfalfa. Crop Sci. 1983, 23, 283–289. [Google Scholar] [CrossRef]
- Silva, E.M.; Dean, B.B. Effect of Nectar Composition and Nectar Concentration on Honey Bee (Hymenoptera: Apidae) Visitations to Hybrid Onion Flowers. J. Econ. Entomol. 2000, 93, 1216–1221. [Google Scholar] [CrossRef]
- Gamisch, A.; Staedler, Y.M.; Schönenberger, J.; Fischer, G.A.; Comes, H.P. Histological and Micro-CT Evidence of Stigmatic Rostellum Receptivity Promoting Auto-Pollination in the Madagascan Orchid Bulbophyllum bicoloratum. PLoS ONE 2013, 8, e72688. [Google Scholar] [CrossRef]
- Heiduk, A.; Pramanik, D.; Spaans, M.; Gast, L.; Dorst, N.; van Heuven, B.J.; Gravendeel, B. Pitfall Flower Development and Organ Identity of Ceropegia sandersonii (Apocynaceae-Asclepiadoideae). Plants 2020, 9, 1767. [Google Scholar] [CrossRef]
- Tominaga, R.; Iwata, M.; Sano, R.; Inoue, K.; Okada, K.; Wada, T. Arabidopsis CAPRICE-LIKE MYB 3 (CPL3) controls endoreduplication and flowering development in addition to trichome and root hair formation. Development 2008, 135, 1335–1345. [Google Scholar] [CrossRef] [Green Version]
- Wada, T.; Tominaga-Wada, R. Caprice family genes control flowering time through both promoting and repressing constans and flowering locus T expression. Plant Sci. 2015, 241, 260–265. [Google Scholar] [CrossRef]
- Zhang, Y.; Shen, J.; Bartholomew, E.S.; Dong, M.; Chen, S.; Yin, S.; Zhai, X.; Feng, Z.; Ren, H.; Liu, X. Tiny branched hair functions in multicellular trichome development through an ethylene pathway in Cucumis sativus L. Plant J. 2021, 106, 753–765. [Google Scholar] [CrossRef]
- Wang, X.; Shen, C.; Meng, P.; Tan, G.; Lv, L. Analysis and review of trichomes in plants. BMC Plant Biol. 2021, 21, 70. [Google Scholar] [CrossRef] [PubMed]
- Fenster, C.B.; Armbruster, W.S.; Wilson, P.; Dudash, M.R.; Thomson, J.D. Pollination Syndromes and Floral Specialization. Annu. Rev. Ecol. ESyst. 2004, 35, 375–403. [Google Scholar] [CrossRef]
- Campos, E.O.; Bradshaw, H.D.; Daniel, T.L. Shape matters: Corolla curvature improves nectar discovery in the hawkmoth Manduca sexta. Funct. Ecol. 2015, 29, 462–468. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bao, T.; Melenka, G.W.; Ljubotina, M.K.; Carey, J.P.; Cahill, J.F. A new method for the rapid characterization of root growth and distribution using digital image correlation. New Phytol. 2018, 218, 835–846. [Google Scholar] [CrossRef] [PubMed]
- Tafforeau, P.; Boistel, R.; Boller, E.; Bravin, A.; Brunet, M.; Chaimanee, Y.; Cloetens, P.; Feist, M.; Hoszowska, J.; Jaeger, J.-J.; et al. Applications of X-ray synchrotron microtomography for non-destructive 3D studies of paleontological specimens. Appl. Phys. A 2006, 83, 195–202. [Google Scholar] [CrossRef]
- Smith, S.Y.; Collinson, M.E.; Rudall, P.J.; Simpson, D.A.; Marone, F.; Stampanoni, M. Virtual taphonomy using synchrotron tomographic microscopy reveals cryptic features and internal structure of modern and fossil plants. Proc. Natl. Acad. Sci. USA 2009, 106, 12013–12018. [Google Scholar] [CrossRef]







Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Begot, L.; Slavkovic, F.; Oger, M.; Pichot, C.; Morin, H.; Boualem, A.; Favier, A.-L.; Bendahmane, A. Precision Phenotyping of Nectar-Related Traits Using X-ray Micro Computed Tomography. Cells 2022, 11, 3452. https://doi.org/10.3390/cells11213452
Begot L, Slavkovic F, Oger M, Pichot C, Morin H, Boualem A, Favier A-L, Bendahmane A. Precision Phenotyping of Nectar-Related Traits Using X-ray Micro Computed Tomography. Cells. 2022; 11(21):3452. https://doi.org/10.3390/cells11213452
Chicago/Turabian StyleBegot, Laurent, Filip Slavkovic, Myriam Oger, Clement Pichot, Halima Morin, Adnane Boualem, Anne-Laure Favier, and Abdelhafid Bendahmane. 2022. "Precision Phenotyping of Nectar-Related Traits Using X-ray Micro Computed Tomography" Cells 11, no. 21: 3452. https://doi.org/10.3390/cells11213452