
Hyaluronan Regulates Neuronal and Immune Function in the Rat Small Intestine and Colonic Microbiota after Ischemic/Reperfusion Injury

 , , , , , ,
, , , , , ,  ,
,  and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Animals and In Vivo Treatment
2.2. Histology
2.3. Immunohistochemistry
2.3.1. Cross-Sections
2.3.2. Whole-Mount Immunohistochemistry
2.4. Microscopy, Image Acquisition and Analysis
2.4.1. Histology
2.4.2. Immunohistochemistry
2.5. HA ELISA Assay
2.6. RNA Isolation and Quantitative RT PCR
2.7. Western Immunoblot Analysis
2.8. Next Generation Sequencing
2.9. Excitatory and Inhibitory In Vitro Motor Responses
2.10. Chemicals
2.11. Statistical Analysis
3. Results
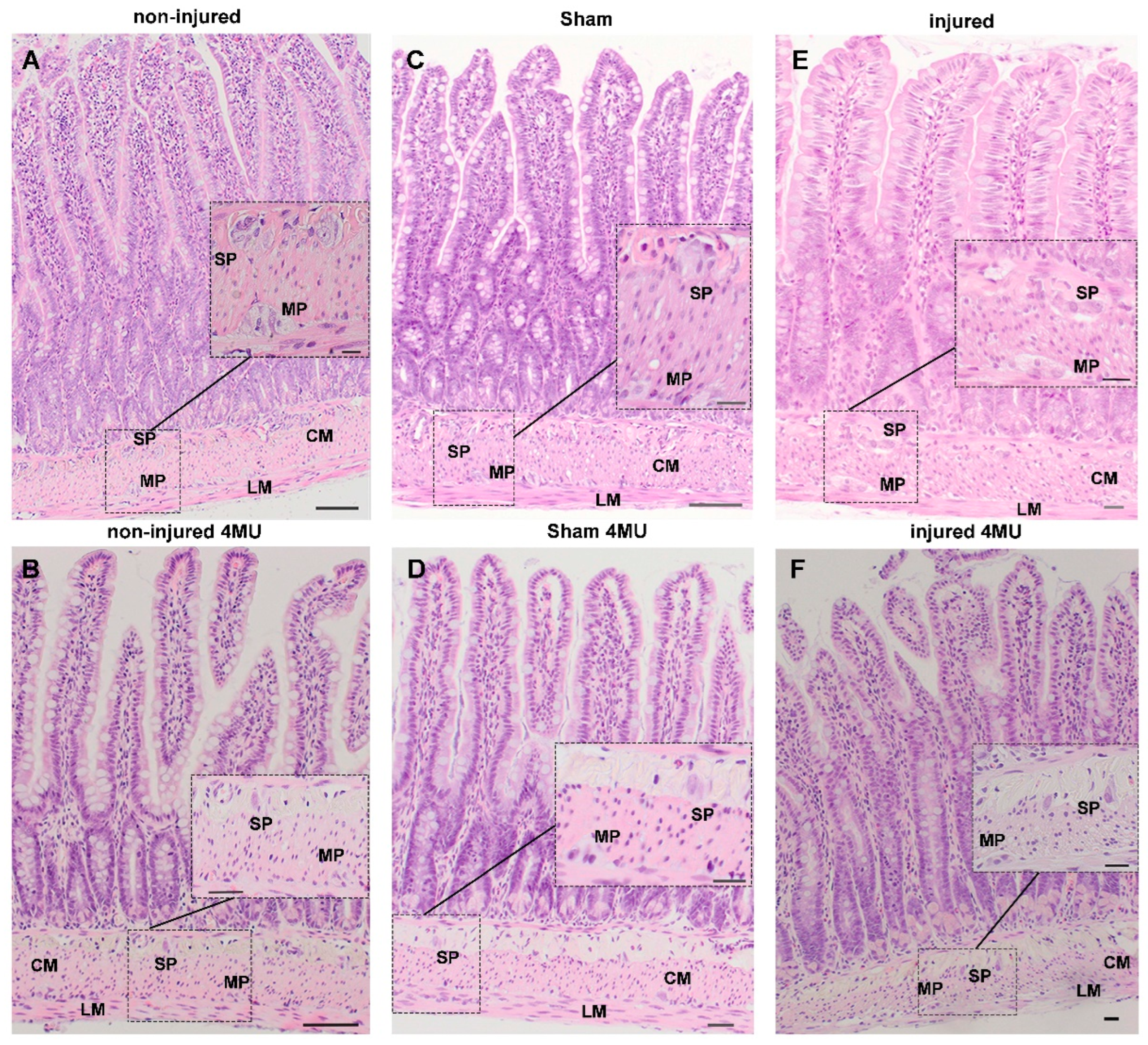
3.1. Effect of IRI Injury and HA Synthesis Blockade on Rat Small Intestine: General Observations and Histological Assessment
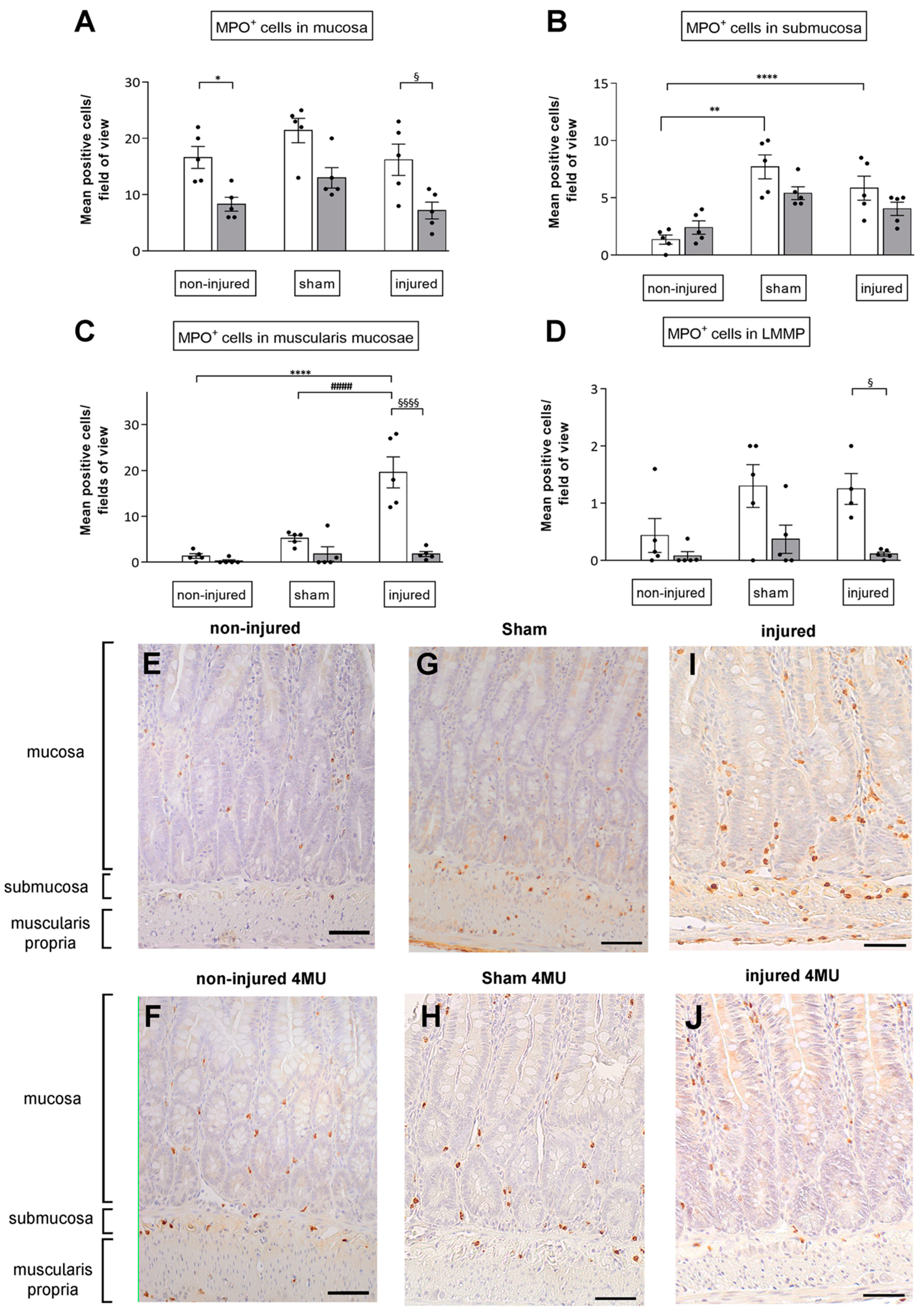
3.2. Degree of Neutrophil Infiltration in the Different Intestinal Layers
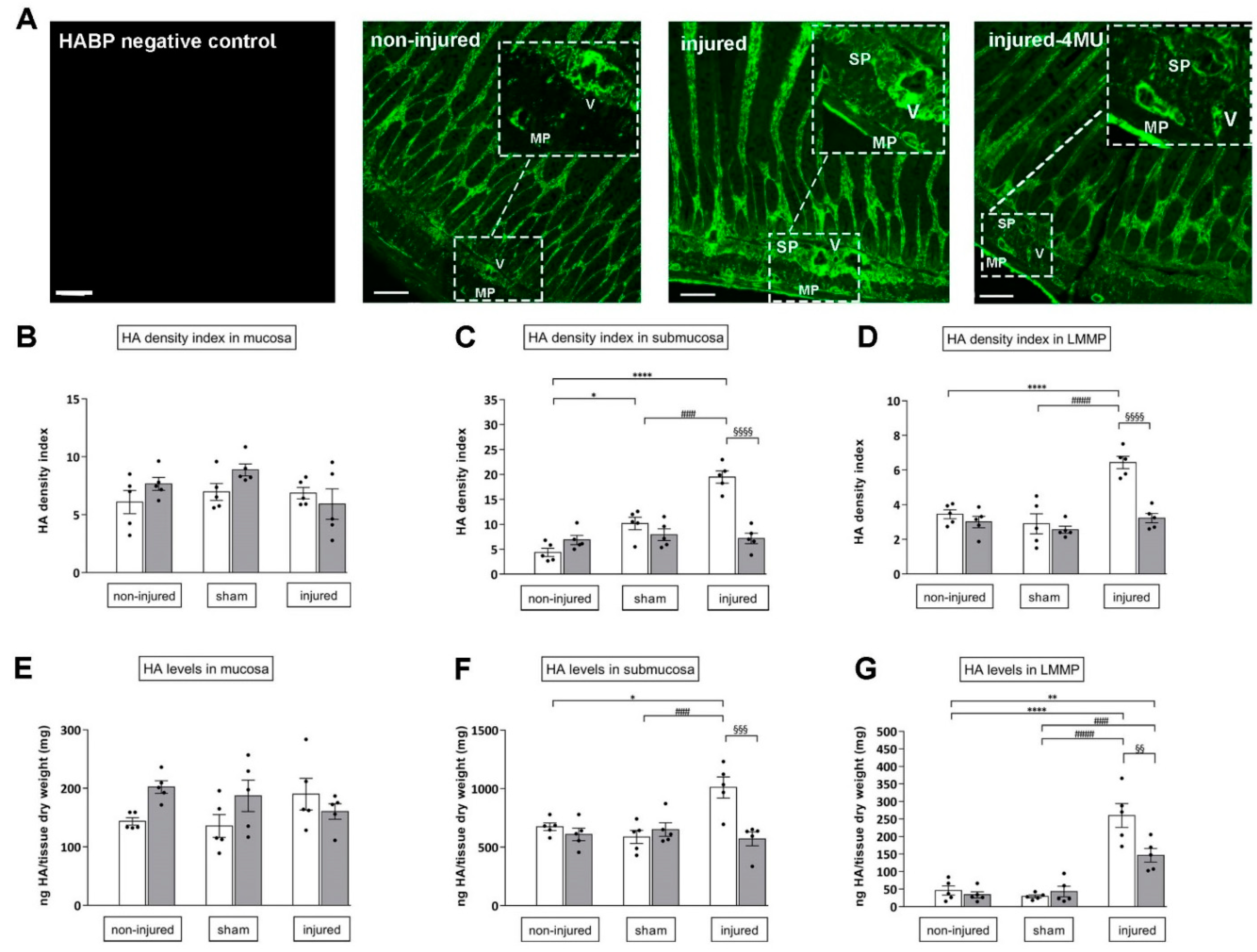
3.3. Effect of IRI Injury and HA Synthesis Blockade in Rat Small Intestine
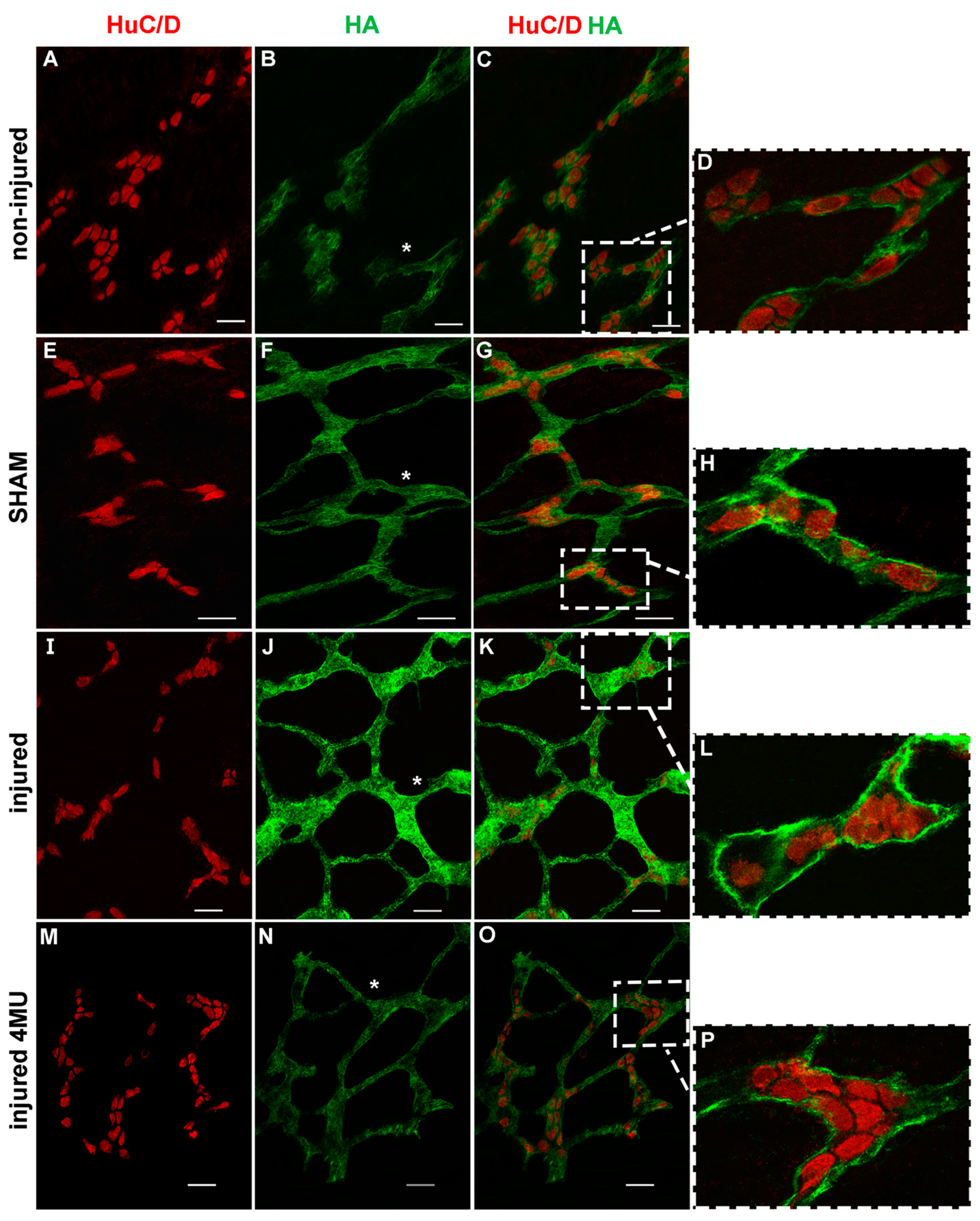
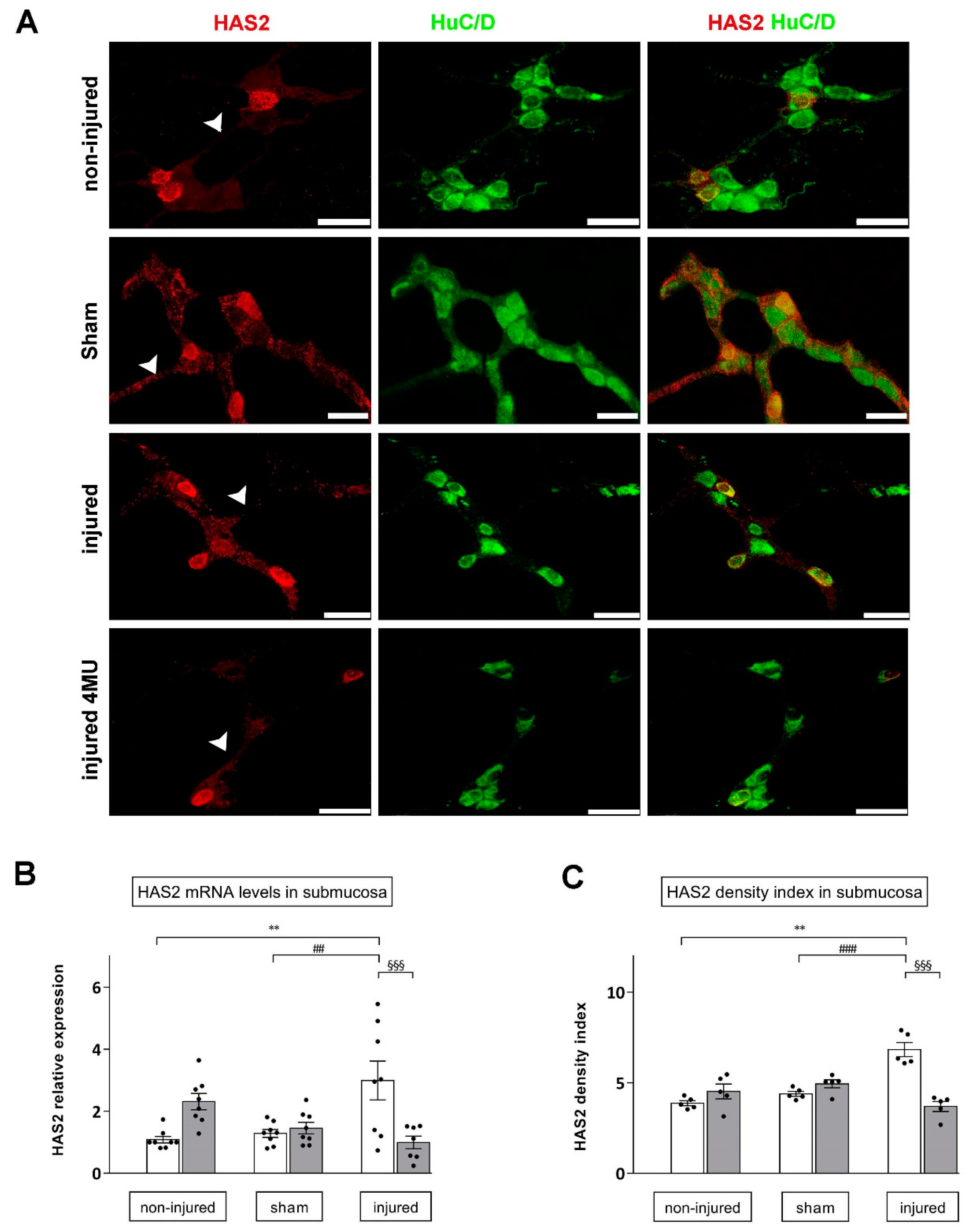
3.4. Effect of IRI Injury and HA Synthesis Blockade in Rat Small Intestine Submucosal Ganglia
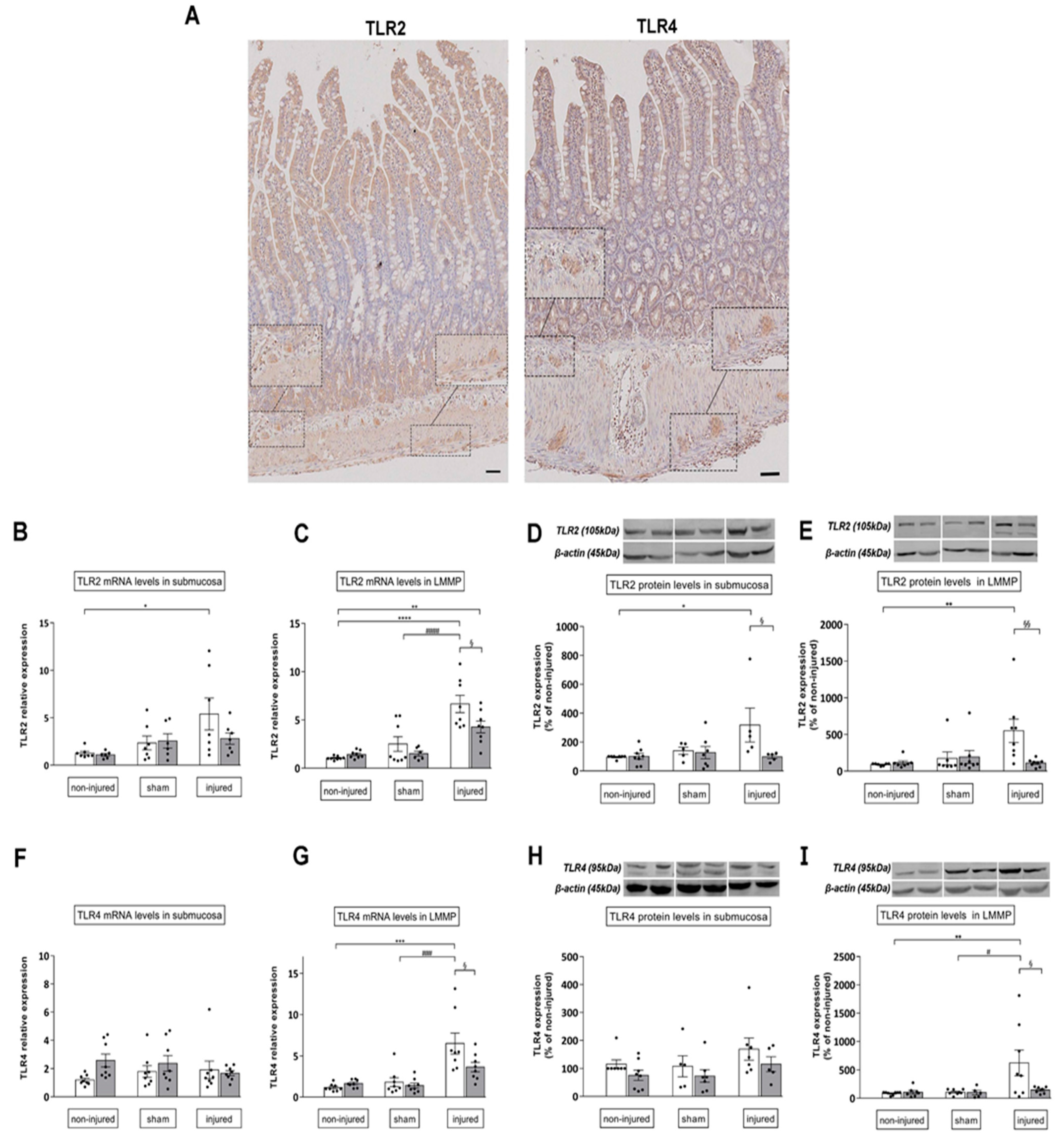
3.5. 4-MU Influences TLR2, TLR4 Expression of in the Rat Small Intestine after Iri Injury
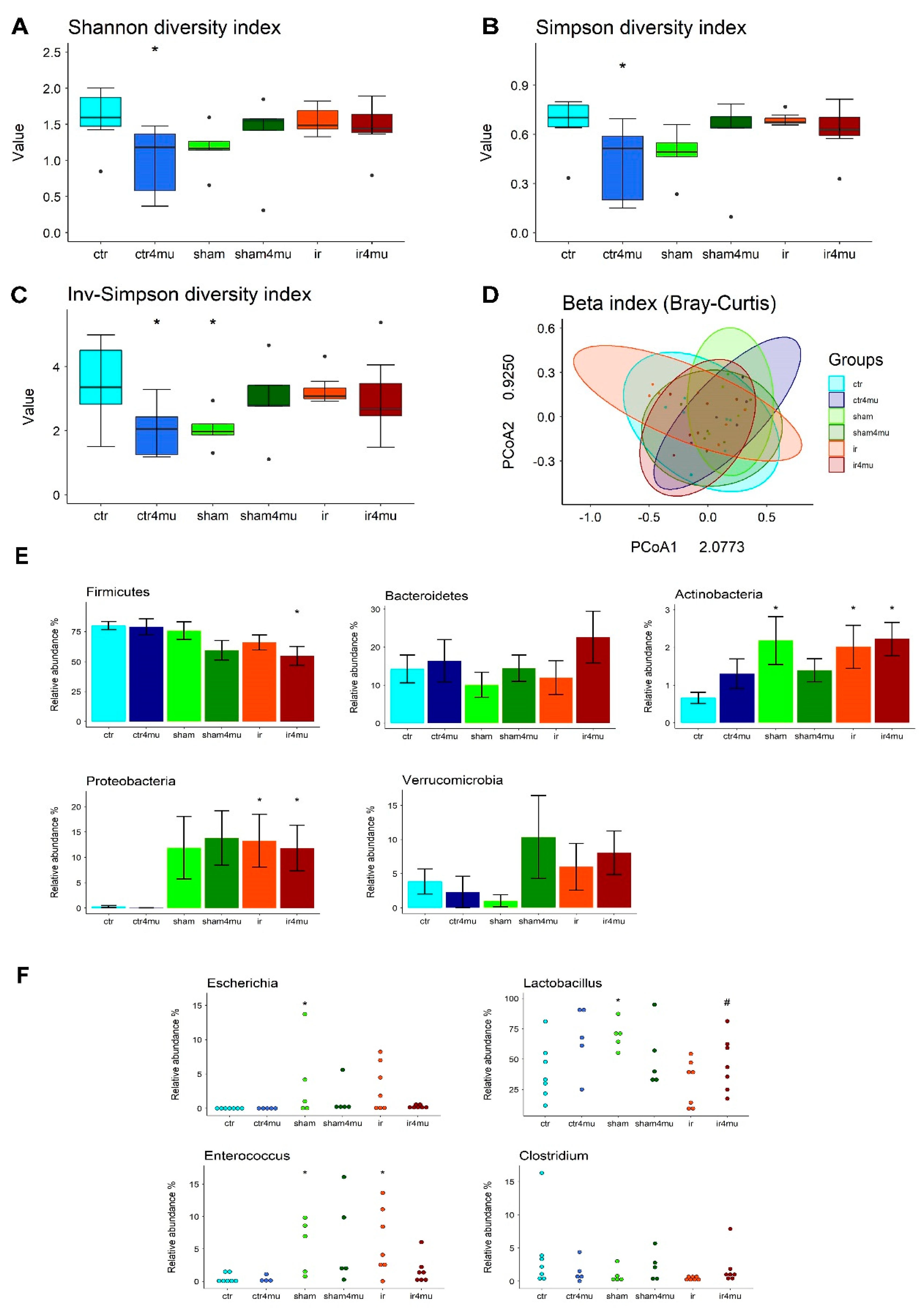
3.6. Gut Microbiota Composition
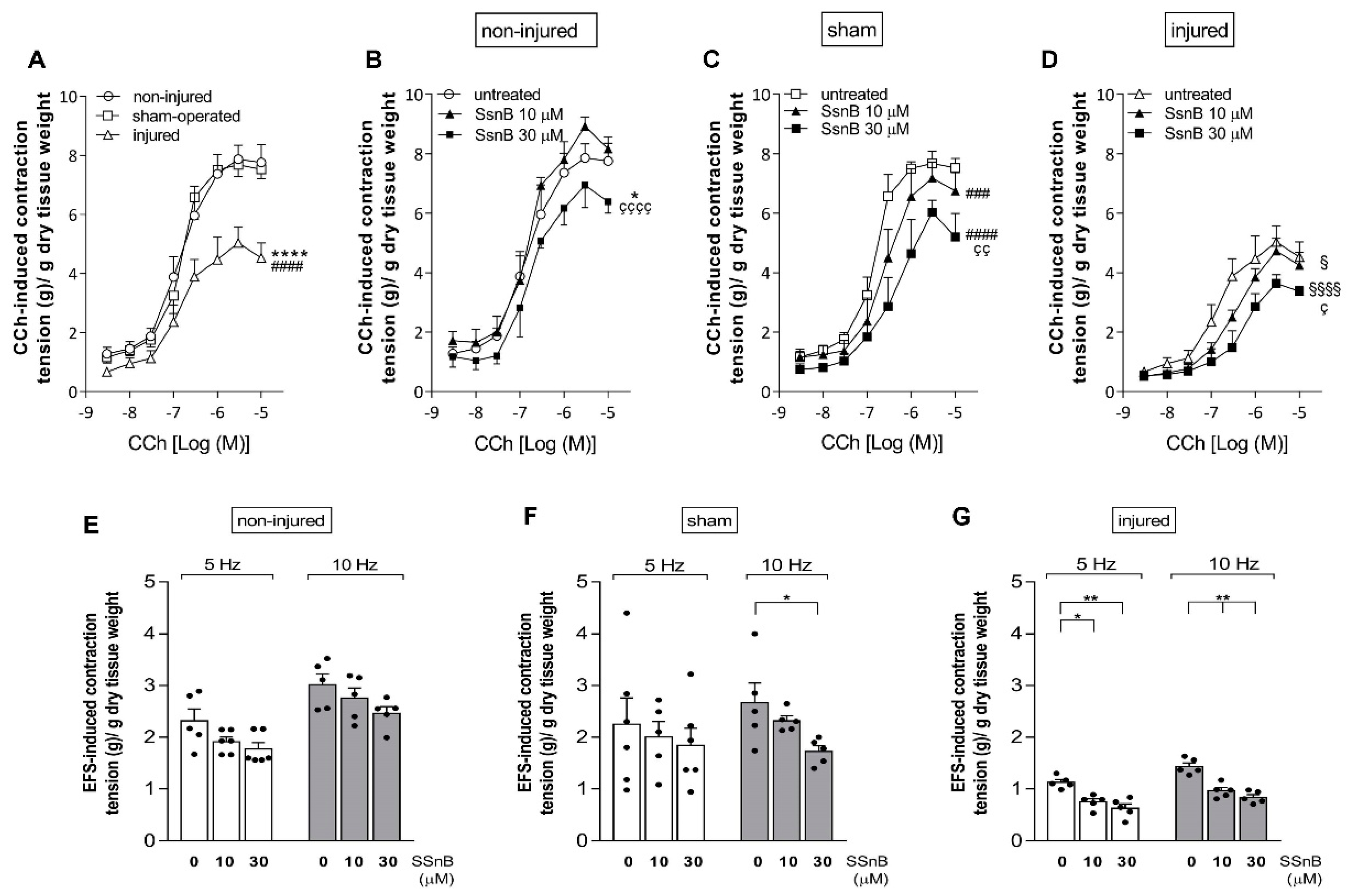
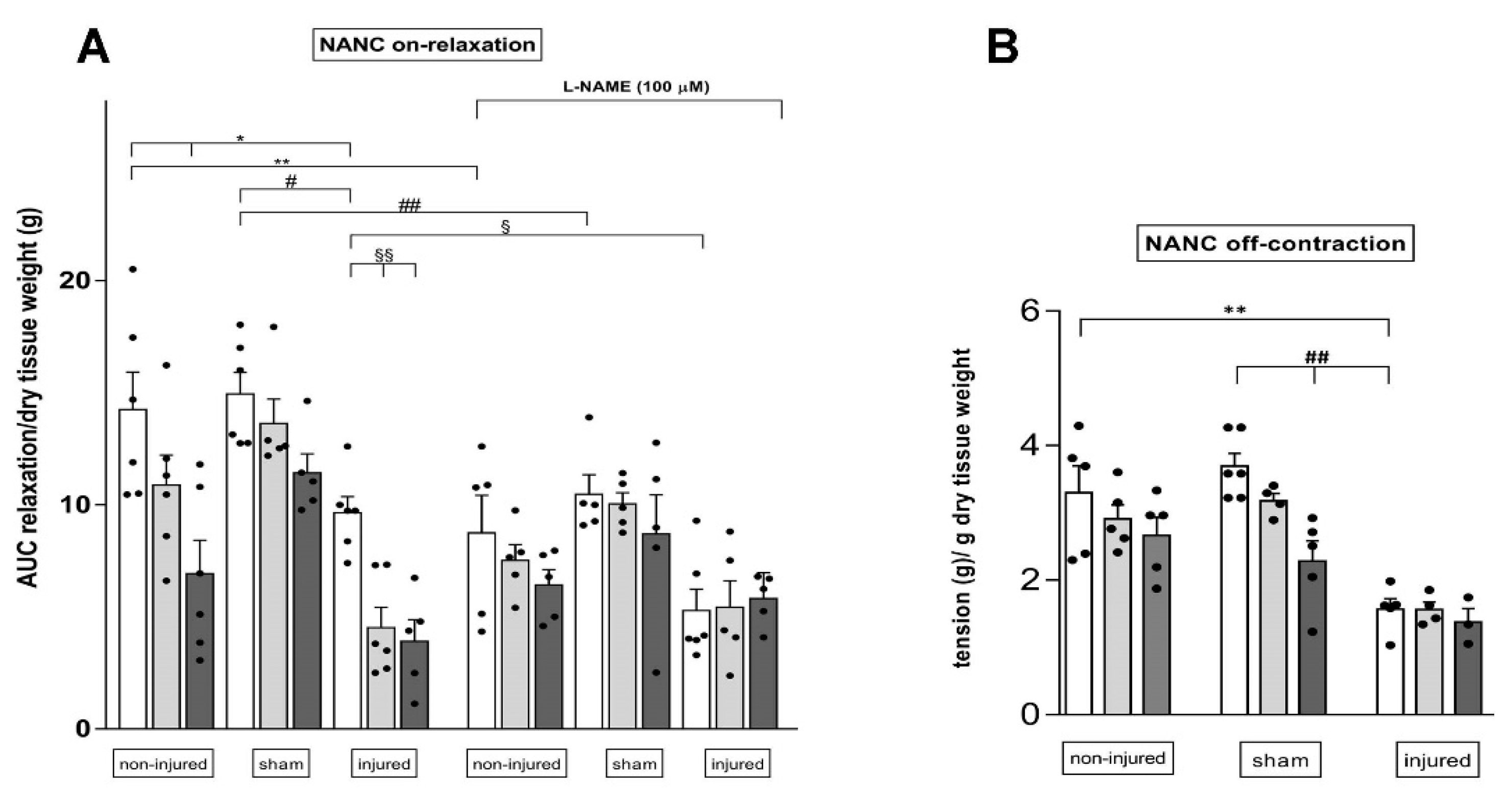
3.7. Effect of the TLR2 and TLR4 Inhibitor Sparstolonin B on the Rat Small Intestine Neuromuscular Function after IRI Injury
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Haglund, U.; Bergqvist, D. Intestinal ischemia—The basics. Langenbeck’s Arch. Surg. 1999, 384, 233–238. [Google Scholar] [CrossRef] [PubMed]
- Roussel, A.; Castier, Y.; Nuzzo, A.; Pellenc, Q.; Sibert, A.; Panis, Y.; Bouhnik, Y.; Corcos, O. Revascularization of acute mesenteric ischemia after creation of a dedicated multidisciplinary center. J. Vasc. Surg. 2015, 62, 1251–1256. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mallick, I.H.; Yang, W.; Winslet, M.C.; Seifalian, A.M. Ischemia-reperfusion injury of the intestine and protective strategies against injury. Dig. Dis. Sci. 2004, 49, 1359–1377. [Google Scholar] [CrossRef] [PubMed]
- Lindeström, L.-M.; Ekblad, E. Structural and neuronal changes in rat ileum after ischemia with reperfusion. Dig. Dis. Sci. 2004, 49, 1212–1222. [Google Scholar] [CrossRef]
- Hierholzer, C.; Kalff, J.C.; Audolfsson, G.; Billiar, T.R.; Tweardy, D.J.; Bauer, A.J. Molecular and functional contractile sequelae of rat intestinal ischemia/reperfusion injury. Transplantation 1999, 68, 1244–1254. [Google Scholar] [CrossRef]
- Carpanese, E.; Moretto, P.; Filpa, V.; Marchet, S.; Moro, E.; Crema, F.; Frigo, G.; Giaroni, C. Antagonism of ionotropic glutamate receptors attenuates chemical ischemia-induced injury in rat primary cultured myenteric ganglia. PLoS ONE 2014, 9, e113613. [Google Scholar] [CrossRef]
- Bistoletti, M.; Bosi, A.; Caon, I.; Chiaravalli, A.M.; Moretto, P.; Genoni, A.; Moro, E.; Karousou, E.; Viola, M.; Crema, F.; et al. Involvement of hyaluronan in the adaptive changes of the rat small intestine neuromuscular function after ischemia/reperfusion injury. Sci. Rep. 2020, 10, 11521. [Google Scholar] [CrossRef]
- Bistoletti, M.; Bosi, A.; Banfi, D.; Cristina, G.; Baj, A. The microbiota-gut-brain axis: Focus on the fundamental communication pathways. In Progress in Molecular Biology and Translational Science; Elsevier B.V.: Amsterdam, The Netherlands, 2020; Volume 176, pp. 43–110. [Google Scholar]
- Anitha, M.; Vijay-Kumar, M.; Sitaraman, S.V.; Gewirtz, A.T.; Srinivasan, S. Gut microbial products regulate murine gastrointestinal motility via Toll-like receptor 4 signaling. Gastroenterology 2012, 143, 1006–1016. [Google Scholar] [CrossRef] [Green Version]
- Grasa, L.; Abecia, L.; Forcén, R.; Castro, M.; de Jalón, J.A.G.; Latorre, E.; Alcalde, A.I.; Murillo, M.D. Antibiotic-Induced Depletion of Murine Microbiota Induces Mild Inflammation and Changes in Toll-Like Receptor Patterns and Intestinal Motility. Microb. Ecol. 2015, 70, 835–848. [Google Scholar] [CrossRef]
- Yoshiya, K.; Lapchak, P.H.; Thai, T.H.; Kannan, L.; Rani, P.; Lucca, J.J.D.; Tsokos, G.C. Depletion of gut commensal bacteria attenuates intestinal ischemia/reperfusion injury. Am. J. Physiol. Gastrointest. Liver Physiol. 2011, 301, G1020–G1030. [Google Scholar] [CrossRef]
- Caon, I.; Parnigoni, A.; Viola, M.; Karousou, E.; Passi, A.; Vigetti, D. Cell Energy Metabolism and Hyaluronan Synthesis. J. Histochem. Cytochem. 2020, 69, 35–47. [Google Scholar] [CrossRef] [PubMed]
- Passi, A.; Vigetti, D. Hyaluronan as tunable drug delivery system. Adv. Drug Deliv. Rev. 2019, 146, 83–96. [Google Scholar] [CrossRef] [PubMed]
- Filpa, V.; Bistoletti, M.; Caon, I.; Moro, E.; Grimaldi, A.; Moretto, P.; Baj, A.; Giron, M.C.; Karousou, E.; Viola, M.; et al. Changes in hyaluronan deposition in the rat myenteric plexus after experimentally-induced colitis. Sci. Rep. 2017, 7, 17644. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tavianatou, A.G.; Caon, I.; Franchi, M.; Piperigkou, Z.; Galesso, D.; Karamanos, N.K. Hyaluronan: Molecular size-dependent signaling and biological functions in inflammation and cancer. FEBS J. 2019, 286, 2883–2908. [Google Scholar] [CrossRef] [Green Version]
- Cyphert, J.M.; Trempus, C.S.; Garantziotis, S. Size Matters: Molecular Weight Specificity of Hyaluronan Effects in Cell Biology. Int. J. Cell Biol. 2015, 2015, 563818. [Google Scholar] [CrossRef] [Green Version]
- De La Motte, C.A.; Hascall, V.C.; Calabro, A.; Yen-Lieberman, B.; Strong, S.A. Mononuclear leukocytes preferentially bind via CD44 to hyaluronan on human intestinal mucosal smooth muscle cells after virus infection or treatment with poly(I.C). J. Biol. Chem. 1999, 274, 30747–30755. [Google Scholar] [CrossRef] [Green Version]
- Kessler, S.; Rho, H.; West, G.; Fiocchi, C.; Drazba, J.; de la Motte, C. Hyaluronan (HA) deposition precedes and promotes leukocyte recruitment in intestinal inflammation. Clin. Transl. Sci. 2008, 1, 57–61. [Google Scholar] [CrossRef] [Green Version]
- Jiang, D.; Liang, J.; Fan, J.; Yu, S.; Chen, S.; Luo, Y.; Prestwich, G.D.; Mascarenhas, M.M.; Garg, H.G.; Quinn, D.A.; et al. Regulation of lung injury and repair by Toll-like receptors and hyaluronan. Nat. Med. 2005, 11, 1173–1179. [Google Scholar] [CrossRef]
- Kultti, A.; Pasonen-Seppänen, S.; Jauhiainen, M.; Rilla, K.J.; Kärnä, R.; Pyöriä, E.; Tammi, R.H.; Tammi, M.I. 4-Methylumbelliferone inhibits hyaluronan synthesis by depletion of cellular UDP-glucuronic acid and downregulation of hyaluronan synthase 2 and 3. Exp. Cell Res. 2009, 315, 1914–1923. [Google Scholar] [CrossRef]
- Nagy, N.; Kuipers, H.F.; Frymoyer, A.R.; Ishak, H.D.; Bollyky, J.B.; Wight, T.N.; Bollyky, P.L. 4-Methylumbelliferone tretament and hyaluronan inhbition as a therapeutic strategy in inflammation, autoimmunity, and cancer. Front. Immunol. 2015, 6, 123. [Google Scholar] [CrossRef]
- Kakizaki, I.; Kojima, K.; Takagaki, K.; Endo, M.; Kannagi, R.; Ito, M.; Maruo, Y.; Sato, H.; Yasuda, T.; Mita, S.; et al. A novel mechanism for the inhibition of hyaluronan biosynthesis by 4-methylumbelliferone. J. Biol. Chem. 2004, 279, 33281–33289. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vigetti, D.; Rizzi, M.; Viola, M.; Karousou, E.; Genasetti, A.; Clerici, M.; Bartolini, B.; Hascall, V.C.; De Luca, G.; Passi, A. The effects of 4-methylumbelliferone on hyaluronan synthesis, MMP2 activity, proliferation, and motility of human aortic smooth muscle cells. Glycobiology 2009, 19, 537–546. [Google Scholar] [CrossRef] [Green Version]
- Qiaol, L.; Wu, Q.; Jiang, J.; Duan, J.; Wang, C.; Smith, M.D.; Lu, H.; Wang, Q.; Nagarkatti, P.; Daping, F. Characterization of spartstolonin B, a chinese herb-derived compound, as a selective Toll-like receptor antagonist with potent anti-inflammatory properties. J. Biol. Chem. 2011, 286, 26470–26479. [Google Scholar]
- Olivares, C.N.; Alaniz, L.D.; Menger, M.D.; Barañao, R.I.; Laschke, M.W.; Meresman, G.F. Inhibition of Hyaluronic Acid Synthesis Suppresses Angiogenesis in Developing Endometriotic Lesions. PLoS ONE 2016, 11, e0152302. [Google Scholar] [CrossRef]
- Filpa, V.; Carpanese, E.; Marchet, S.; Pirrone, C.; Conti, A.; Rainero, A.; Moro, E.; Chiaravalli, A.M.; Zucchi, I.; Moriondo, A.; et al. Nitric oxide regulates homeoprotein OTX1 and OTX2 expression in the rat myenteric plexus after intestinal ischemia-reperfusion injury. Am. J. Physiol.-Gastrointest. Liver Physiol. 2017, 312, G374–G389. [Google Scholar] [CrossRef] [PubMed]
- Desmet, A.; Cirillo, C.; Vanden Berghe, P. Distinct subcellular localization of the neuronal marker HuC/D reveals hypoxia-induced damage in enteric neurons. Neurogastroenterol. Motil. 2014, 26, 1131–1143. [Google Scholar] [CrossRef] [Green Version]
- Yoshida, H.; Nagaoka, A.; Kusaka-Kikushima, A.; Tobiishi, M.; Kawabata, K.; Sayo, T.; Sakai, S.; Sugiyama, Y.; Enomoto, H.; Okada, Y.; et al. KIAA1199, a deafness gene of unknown function, is a new hyaluronan binding protein involved in hyaluronan depolymerization. Proc. Natl. Acad. Sci. USA 2013, 110, 5612–5617. [Google Scholar] [CrossRef] [Green Version]
- Caon, I.; Bartolini, B.; Moretto, P.; Parnigoni, A.; Caravà, E.; Vitale, D.L.; Alaniz, L.; Viola, M.; Karousou, E.; De Luca, G.; et al. Sirtuin 1 reduces hyaluronan synthase 2 expression by inhibiting nuclear translocation of NF-κB and expression of the long-noncoding RNA HAS2-AS1. J. Biol. Chem. 2020, 295, 3485–3496. [Google Scholar] [CrossRef] [Green Version]
- Rinaldi, E.; Consonni, A.; Cordiglieri, C.; Sacco, G.; Crasà, C.; Fontana, A.; Morelli, L.; Elli, M.; Mantegazza, R.; Baggi, F. Therapeutic Effect of Bifidobacterium Administration on Experimental Autoimmune Myasthenia Gravis in Lewis Rats. Front. Immunol. 2019, 10, 2949. [Google Scholar] [CrossRef]
- Xing, F.; Zhang, W.; Wen, J.; Bai, L.; Gu, H.; Li, Z.; Zhang, J.; Tao, Y.X.; Xu, J.T. TLR4/NF-κB signaling activation in plantar tissue and dorsal root ganglion involves in the development of postoperative pain. Mol. Pain 2018, 14, 1744806918807050. [Google Scholar] [CrossRef]
- Cha, M.; Lee, K.H.; Lee, B.H. Astroglial changes in the zona incerta in response to motor cortex stimulation in a rat model of chronic neuropathy. Sci. Rep. 2020, 10, 943. [Google Scholar] [CrossRef] [PubMed]
- Ceccotti, C.; Giaroni, C.; Bistoletti, M.; Viola, M.; Crema, F.; Terova, G. Neurochemical characterization of myenteric neurons in the juvenile gilthead sea bream (Sparus aurata) intestine. PLoS ONE 2018, 13, e0201760. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bistoletti, M.; Micheloni, G.; Baranzini, N.; Bosi, A.; Conti, A.; Filpa, V.; Pirrone, C.; Millefanti, G.; Moro, E.; Grimaldi, A.; et al. Homeoprotein OTX1 and OTX2 involvement in rat myenteric neuron adaptation after DNBS-induced colitis. PeerJ 2020, 8, e8442. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Takahashi, S.; Tomita, J.; Nishioka, K.; Hisada, T.; Nishijima, M. Development of a Prokaryotic Universal Primer for Simultaneous Analysis of Bacteria and Archaea Using Next-Generation Sequencing. PLoS ONE 2014, 9, e105592. [Google Scholar] [CrossRef] [Green Version]
- Baj, A.; Bistoletti, M.; Bosi, A.; Moro, E.; Giaroni, C.; Crema, F. Marine Toxins and Nociception: Potential Therapeutic Use in the Treatment of Visceral Pain Associated with Gastrointestinal Disorders. Toxins 2019, 11, 449. [Google Scholar] [CrossRef] [Green Version]
- Cerantola, S.; Caputi, V.; Marsilio, I.; Ridolfi, M.; Faggin, S.; Bistoletti, M.; Giaroni, C.; Giron, M.C. Involvement of Enteric Glia in Small Intestine Neuromuscular Dysfunction of Toll-Like Receptor 4-Deficient Mice. Cells 2020, 9, 838. [Google Scholar] [CrossRef] [Green Version]
- Liang, Q.; Dong, S.; Lei, L.; Liu, J.; Zhang, J.; Li, J.; Duan, J.; Fan, D. Protective effects of Sparstolonin B, a selective TLR2 and TLR4 antagonist, on mouse endotoxin shock. Cytokine 2015, 75, 302–309. [Google Scholar] [CrossRef] [Green Version]
- Sekirov, I.; Russell, S.L.; Antunes, L.C.; Finlay, B.B. Gut microbiota in health and disease. Physiol. Rev. 2010, 90, 859–904. [Google Scholar] [CrossRef] [Green Version]
- Pontell, L.; Sharma, P.; Rivera, L.R.; Thacker, M.; Tan, Y.H.; Brock, J.A.; Furness, J.B. Damaging effects of ischemia/reperfusion on intestinal muscle. Cell Tissue Res. 2011, 343, 411–419. [Google Scholar] [CrossRef]
- Parnigoni, A.; Viola, M.; Karousou, E.; Rovera, S.; Giaroni, C.; Passi, A.; Vigetti, D. Hyaluronan in pathophysiology of vascular diseases: Specific roles in smooth muscle cells, endothelial cells, and macrophages. Am. J. Physiol. Cell Physiol. 2022, 323, C505–C519. [Google Scholar] [CrossRef] [PubMed]
- Blikslager, A.T.; Moeser, A.J.; Gookin, J.L.; Jones, S.L.; Odle, J. Restoration of barrier function in injured intestinal mucosa. Physiol. Rev. 2007, 87, 545–564. [Google Scholar] [CrossRef] [PubMed]
- Petrey, A.C.; de la Motte, C.A. Hyaluronan, a crucial regulator of inflammation. Front. Immunol. 2014, 5, 101. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Matthijsen, R.A.; Derikx, J.P.; Kuipers, D.; van Dam, R.M.; Dejong, C.H.; Buurman, W.A. Enterocyte shedding and epithelial lining repair following ischemia of the human small intestine attenuate inflammation. PLoS ONE 2009, 4, e7045. [Google Scholar] [CrossRef] [PubMed]
- Banfi, D.; Moro, E.; Bosi, A.; Bistoletti, M.; Cerantola, S.; Crema, F.; Maggi, F.; Giron, M.C.; Giaroni, C.; Baj, A. Impact of Microbial Metabolites on Microbiota–Gut–Brain Axis in Inflammatory Bowel Disease. Int. J. Mol. Sci. 2021, 22, 1623. [Google Scholar] [CrossRef]
- Al’Qteishat, A.; Gaffney, J.; Krupinski, J.; Rubio, F.; West, D.; Kumar, S.; Kumar, P.; Mitsios, N.; Slevin, M. Changes in hyaluronan production and metabolism following ischaemic stroke in man. Brain 2006, 129, 2158–2176. [Google Scholar] [CrossRef]
- Colombaro, V.; Declèves, A.-E.; Jadot, I.; Voisin, V.; Giordano, L.; Habsch, I.; Nonclercq, D.; Flamion, B.; Caron, N. Inhibition of hyaluronan is protective against renal ischaemia-reperfusion injury. Nephrol. Dial. Transplant. 2013, 28, 2484–2493. [Google Scholar] [CrossRef] [Green Version]
- Furness, J.B.; Callaghan, B.P.; Rivera, L.R.; Cho, H.J. The enteric nervous system and gastrointestinal innervation: Integrated local and central control. Adv. Exp. Med. Biol. 2014, 817, 39–71. [Google Scholar]
- Kanashiro, A.; Hiroki, C.H.; da Fonseca, D.M.; Birbrair, A.; Ferreira, R.G.; Bassi, G.S.; Fonseca, M.D.; Kusuda, R.; Cebinelli, G.C.M.; da Silva, K.P.; et al. The role of neutrophils in neuro-immune modulation. Pharmacol. Res. 2020, 151, 104580. [Google Scholar] [CrossRef]
- Lakhan, S.E.; Kirchgessner, A. Neuroinflammation in inflammatory bowel disease. J. Neuroinflamm. 2010, 7, 37. [Google Scholar] [CrossRef] [Green Version]
- Bosi, A.; Banfi, D.; Bistoletti, M.; Moretto, P.; Moro, E.; Crema, F.; Maggi, F.; Karousou, E.; Viola, M.; Passi, A.; et al. Hyaluronan: A Neuroimmune Modulator in the Microbiota-Gut Axis. Cells 2021, 11, 126. [Google Scholar] [CrossRef] [PubMed]
- Wang, F.; Li, Q.; Wang, C.; Tang, C.; Li, J. Dynamic alteration of the colonic microbiota in intestinal ischemia-reperfusion injury. PLoS ONE 2012, 7, e42027. [Google Scholar] [CrossRef]
- Wang, Y.; Jiang, S.; Xiao, J.; Liang, Q.; Tang, M. Sparstolonin B improves neurological outcomes following intracerebral hemorrhage in mice. Exp. Ther. Med. 2018, 15, 5436–5442. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ballabeni, V.; Barocelli, E.; Bertoni, S.; Impicciatore, M. Alterations of intestinal motor responsiveness in a model of mild mesenteric ischemia/reperfusion in rats. Life Sci. 2002, 71, 2025–2035. [Google Scholar] [CrossRef]
- Marsilio, I.; Caputi, V.; Latorre, E.; Cerantola, S.; Paquola, A.; Alcalde, A.I.; Mesonero, J.E.; O’Mahony, S.M.; Bertazzo, A.; Giaroni, C.; et al. Oxidized phospholipids affect small intestine neuromuscular transmission and serotonergic pathways in juvenile mice. Neurogastroenterol. Motil. 2020, 33, e14036. [Google Scholar] [CrossRef] [PubMed]
- Corbett, A.D.; Lees, G.M. Depressant effects of hypoxia and hypoglycaemia on neuro-effector transmission of guinea-pig intestine studied in vitro with a pharmacological model. Br. J. Pharmacol. 1997, 120, 107–115. [Google Scholar] [CrossRef] [Green Version]
- Giaroni, C. Purinergic signalling and development of the autonomic nervous system. Auton. Neurosci. Basic Clin. 2015, 191, 67–77. [Google Scholar] [CrossRef]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Primary Antibody | Host Species | Dilution (HC) | Dilution (WB) | Specificity | Source |
| HuC/D biotin | Mouse | 1:100 | _ | Neuronal cells [27] | Invitrogen (A-21272) |
| HABP | _ | 1:100 | _ | Hyaluronan [28] | Hokudo (BC41) |
| HAS2 | Goat | 1:100 | _ | HAS2 [29] | Santa Cruz Biotechnology (sc-34068) |
| MPO | Rabbit | 1:1 | Myeloperoxidase [25] | Dako (GA511) | |
| TLR2 | Rabbit | 1:500 | 1:1000 | TLR2 [30] | ABclonal (A11225) |
| TLR4 | Rabbit | 1:800 | 1:1000 | TLR4 [31] | ABclonal (A5258) |
| β-actin | Mouse | _ | 1:1000 | Housekeeping [32] | Cell Signaling (#3700) |
| Secondary Antibody and Streptavidin complex | |||||
| Anti-rabbit IgG HRP-linked | Donkey | _ | 1:5000 | Amersham (NA934) | |
| Anti-mouse IgG HRP-linked | Horse | _ | 1:2000 | Cell Signaling (7076S) | |
| Anti-rabbit IgG, biotin | Goat | 1:200 | _ | Abnova Corporation (PAB10824) | |
| Anti-goat IgG, biotin | Rabbit | 1:200 | _ | Abnova Corporation (PAB10578) | |
| Anti-goat IgG Cy3-conjugated | Donkey | 1:500 | _ | Jackson Immuno Research Laboratories | |
| FITC-Streptavidin conjugated | 1:200 | _ | Molecular Probes (SA1001) | ||
| Cy3-Streptavidin conjugated | 1:500 | _ | Amersham (PA43001) |
| Gene | Sequence |
|---|---|
| β-actin | F 5′-TGACAGGATGCAGAAGGAGA-3′ R 5′-TAGAGCCACCAATCCACACA-3′ |
| TLR2 | F 5′-CCGAAACCTCAGACAAAGCG-3′ R 5′-ACAGCGTTTGCTGAAGAGGA-3′ |
| TLR4 | F 5′-TGAGATTGCTCAAACATGGC-3′ R 5′-CGAGGCTTTTCCATCCAATA-3′ |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bosi, A.; Banfi, D.; Bistoletti, M.; Catizzone, L.M.; Chiaravalli, A.M.; Moretto, P.; Moro, E.; Karousou, E.; Viola, M.; Giron, M.C.; et al. Hyaluronan Regulates Neuronal and Immune Function in the Rat Small Intestine and Colonic Microbiota after Ischemic/Reperfusion Injury. Cells 2022, 11, 3370. https://doi.org/10.3390/cells11213370
Bosi A, Banfi D, Bistoletti M, Catizzone LM, Chiaravalli AM, Moretto P, Moro E, Karousou E, Viola M, Giron MC, et al. Hyaluronan Regulates Neuronal and Immune Function in the Rat Small Intestine and Colonic Microbiota after Ischemic/Reperfusion Injury. Cells. 2022; 11(21):3370. https://doi.org/10.3390/cells11213370
Chicago/Turabian StyleBosi, Annalisa, Davide Banfi, Michela Bistoletti, Lucia Martina Catizzone, Anna Maria Chiaravalli, Paola Moretto, Elisabetta Moro, Evgenia Karousou, Manuela Viola, Maria Cecilia Giron, and et al. 2022. "Hyaluronan Regulates Neuronal and Immune Function in the Rat Small Intestine and Colonic Microbiota after Ischemic/Reperfusion Injury" Cells 11, no. 21: 3370. https://doi.org/10.3390/cells11213370