
Recent Advancements in Molecular Therapeutics for Corneal Scar Treatment

Abstract
:
1. Introduction
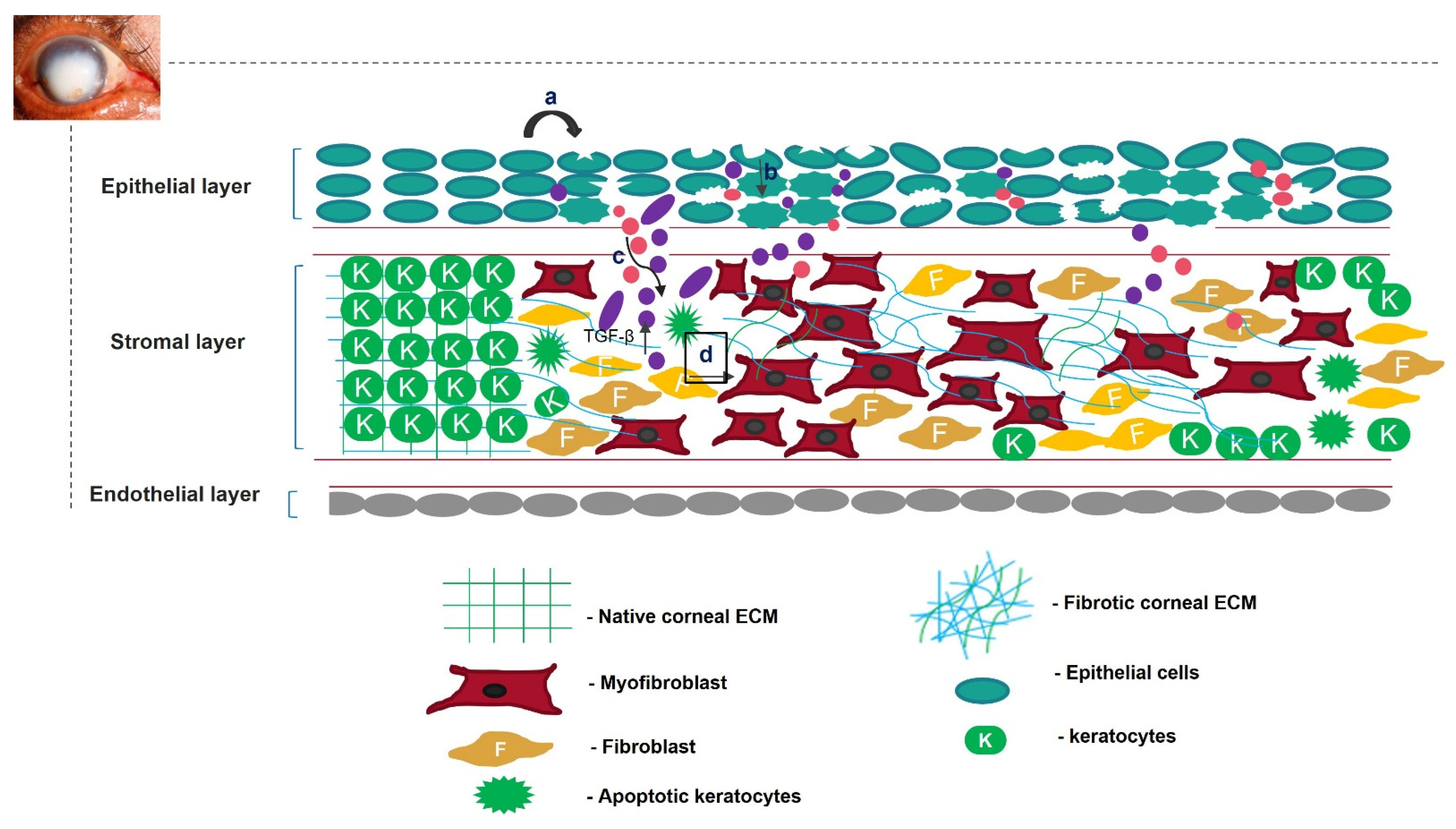
2. Scarring of Cornea
3. Regeneration of Scarless Cornea
3.1. Exosomes as A Therapeutic Tool
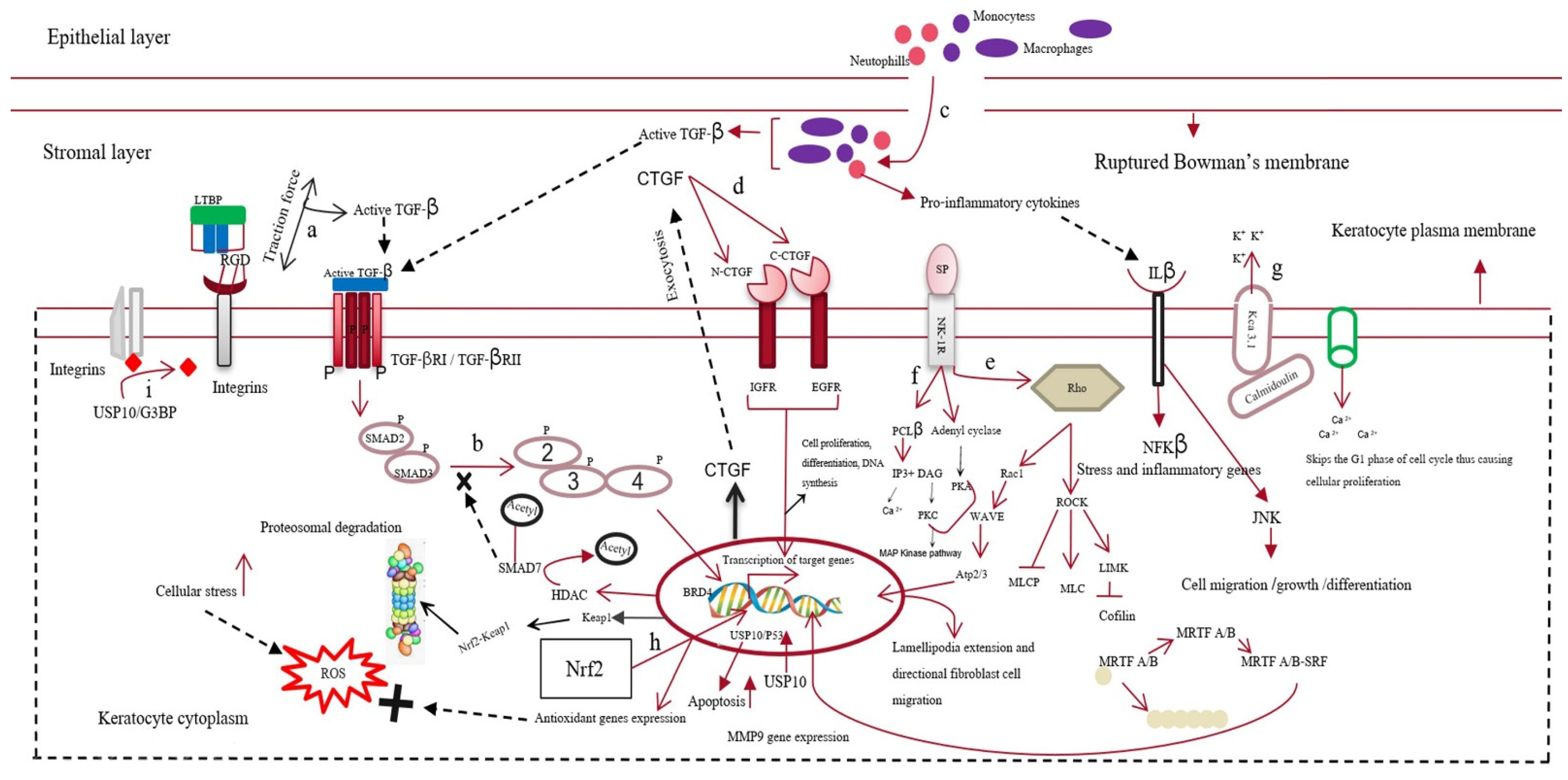
3.2. Targeted Gene Silencing for the Generation of A Scarless Cornea
3.2.1. Targeting Semaphorin 3A in Scarless Cornea Regeneration
3.2.2. Silencing USP10, A Deubiquitinase, Can Prevent Corneal Scarring
3.2.3. Knockout of Kca3.1 Ion Channel for Preventing Corneal Scarring
3.3. Protein Overexpression in Preventing Corneal Scarring
3.3.1. Overexpression of KLF4 in Preventing Scar Formation
3.3.2. Overexpression of Id3 for Reviving Cornea without Scars
3.3.3. Overexpression of SMAD7 for Regenerating Cornea without Scars
3.4. MicroRNA Therapies in Regenerating Cornea without Scars
3.5. Bioactive Molecules as HDAC Inhibitors in the Regeneration of Scarless Cornea
3.6. Guided Wound Healing to Prevent Scarring
3.7. Clinical Therapy for Scar Prevention
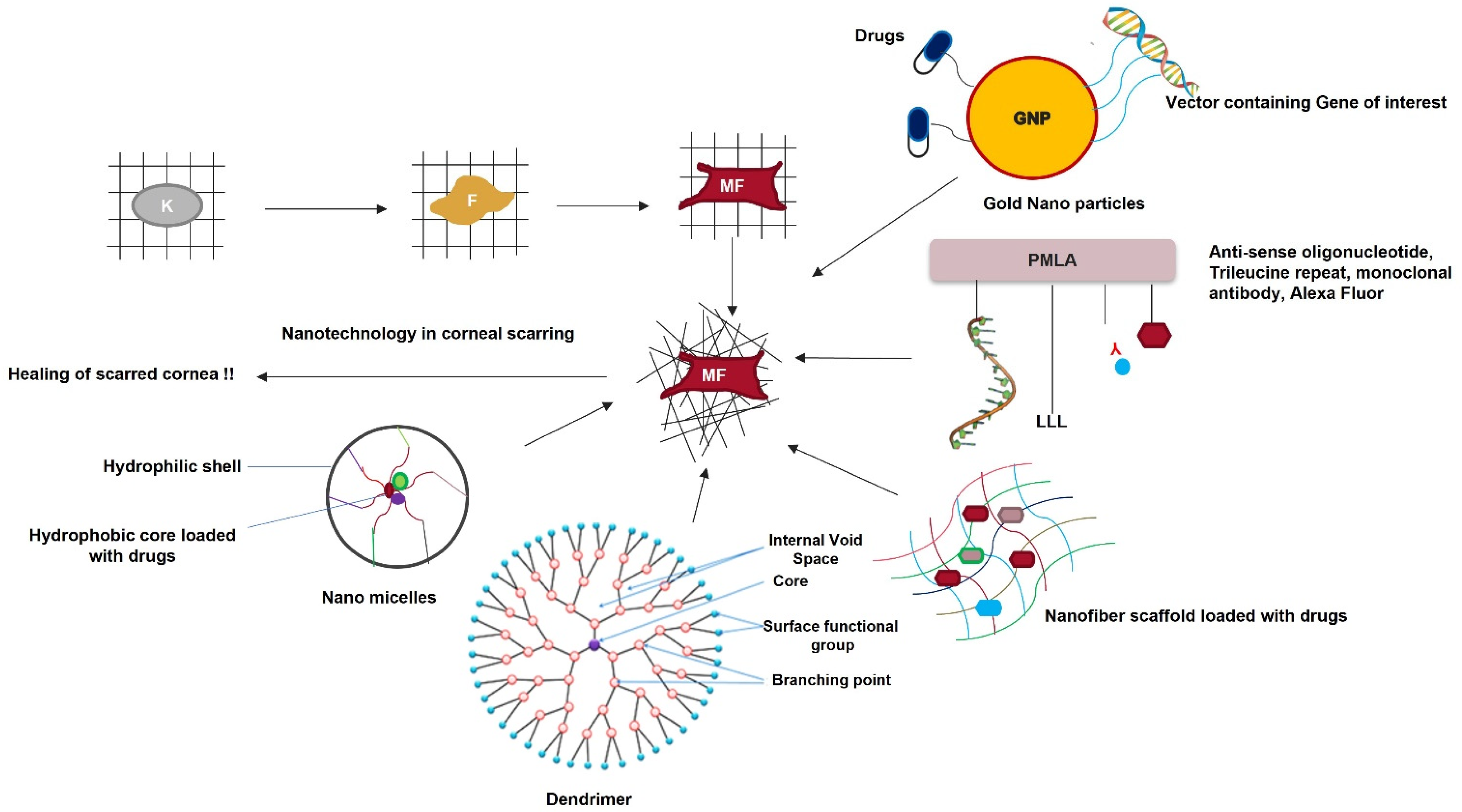
3.8. Nanomedicine in Corneal Scarring Treatment
4. Conclusions and Future Perspectives
Funding
Acknowledgments
Conflicts of Interest
References
- Song, P.; Wang, S.; Zhang, P.; Sui, W.; Zhang, Y.; Liu, T.; Gao, H. The Superficial Stromal Scar Formation Mechanism in Keratoconus: A Study Using Laser Scanning In Vivo Confocal Microscopy. Biomed Res. Int. 2016, 2016, 7092938. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wilson, S.E.; Netto, M.; Ambrósio, R. Corneal Cells: Chatty in Development, Homeostasis, Wound Healing, and Disease. Am. J. Ophthalmol. 2003, 136, 530–536. [Google Scholar] [CrossRef]
- West-Mays, J.A.; Dwivedi, D.J. The Keratocyte: Corneal Stromal Cell with Variable Repair Phenotypes. Int. J. Biochem. Cell Biol. 2006, 38, 1625–1631. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Whitcher, J.P.; Srinivasan, M.; Upadhyay, M.P. Corneal Blindness: A Global Perspective. Bull. World Health Organ. 2001, 79, 214–221. [Google Scholar]
- Agrawal, P.K. The pathology of cornea (a histopathological study). Indian J. Ophthalmol. 1983, 31, 662. [Google Scholar] [PubMed]
- McClintic, S.M.; Srinivasan, M.; Mascarenhas, J.; Greninger, D.A.; Acharya, N.R.; Lietman, T.M.; Keenan, J.D. Improvement in Corneal Scarring Following Bacterial Keratitis. Eye 2013, 27, 443–446. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gain, P.; Jullienne, R.; He, Z.; Aldossary, M.; Acquart, S.; Cognasse, F.; Thuret, G. Global Survey of Corneal Transplantation and Eye Banking. JAMA Ophthalmol. 2016, 134, 167–173. [Google Scholar] [CrossRef] [Green Version]
- Tseng, S.C.G.; Prabhasawat, P.; Lee, S.-H. Amniotic Membrane Transplantation for Conjunctival Surface Reconstruction. Am. J. Ophthalmol. 1997, 124, 765–774. [Google Scholar] [CrossRef]
- Salminen, L. Review: Systemic Absorption of Topically Applied Ocular Drugs in Humans. J. Ocul. Pharmacol. Ther. 1990, 6, 243–249. [Google Scholar] [CrossRef] [PubMed]
- Eghrari, A.O.; Riazuddin, S.A.; Gottsch, J.D. Overview of the Cornea: Structure, Function, and Development. In Progress in Molecular Biology and Translational Science; Academic Press: Waltham, MA, USA, 2015; Volume 134, pp. 7–23. [Google Scholar] [CrossRef]
- Mobaraki, M.; Abbasi, R.; Omidian Vandchali, S.; Ghaffari, M.; Moztarzadeh, F.; Mozafari, M. Corneal Repair and Regeneration: Current Concepts and Future Directions. Front. Bioeng. Biotechnol. 2019, 7, 135. [Google Scholar] [CrossRef] [PubMed]
- Wilson, S.E. Bowman’s Layer in the Cornea– Structure and Function and Regeneration. Exp. Eye Res. 2020, 195, 108033. [Google Scholar] [CrossRef] [PubMed]
- Ljubimov, A.V.; Saghizadeh, M. Progress in Corneal Wound Healing. Prog. Retin. Eye Res. 2015, 49, 17–45. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Imanishi, J.; Kamiyama, K.; Iguchi, I.; Kita, M.; Sotozono, C.; Kinoshita, S. Growth Factors: Importance in Wound Healing and Maintenance of Transparency of the Cornea. Prog. Retin. Eye Res. 2000, 19, 113–129. [Google Scholar] [CrossRef]
- Haber, M.; Cao, Z.; Panjwani, N.; Bedenice, D.; Li, W.W.; Provost, P.J. Effects of Growth Factors (EGF, PDGF-BB and TGF-Beta1) on Cultured Equine Epithelial Cells and Keratocytes: Implications for Wound Healing. Vet. Ophthalmol. 2003, 6, 211–217. [Google Scholar] [CrossRef] [PubMed]
- Hong, J.W.; Liu, J.J.; Lee, J.S.; Mohan, R.R.; Mohan, R.R.; Woods, D.J.; He, Y.G.; Wilson, S.E. Proinflammatory chemokine induction in keratocytes and inflammatory cell infiltration into the cornea. Investig. Ophthalmol. Vis. Sci. 2001, 42, 2795–2803. [Google Scholar]
- Stepp, M.A. Corneal Integrins and Their Functions. Exp. Eye Res. 2006, 83, 3–15. [Google Scholar] [CrossRef]
- Imai, K.; Hiramatsu, A.; Fukushima, D.; Pierschbacher, M.D.; Okada, Y. Degradation of Decorin by Matrix Metalloproteinases: Identification of the Cleavage Sites, Kinetic Analyses and Transforming Growth Factor-Β1 Release. Biochem. J. 1997, 322, 809–814. [Google Scholar] [CrossRef] [PubMed]
- Tandon, A.; Tovey, J.C.K.; Sharma, A.; Gupta, R.; Mohan, R.R. Role of Transforming Growth Factor Beta in Corneal Function, Biology and Pathology. Curr. Mol. Med. 2010, 10, 565–578. [Google Scholar] [CrossRef]
- Blanco-Mezquita, J.T.; Hutcheon, A.E.K.; Stepp, M.A.; Zieske, J.D. AVβ6 Integrin Promotes Corneal Wound Healing. Investig. Opthalmol. Vis. Sci. 2011, 52, 8505–8513. [Google Scholar] [CrossRef] [PubMed]
- Martin, P.; Leibovich, S.J. Inflammatory Cells during Wound Repair: The Good, the Bad and the Ugly. Trends Cell Biol. 2005, 15, 599–607. [Google Scholar] [CrossRef] [PubMed]
- Marrazzo, G.; Bellner, L.; Halilovic, A.; Li Volti, G.; Drago, F.; Dunn, M.W.; Schwartzman, M.L. The Role of Neutrophils in Corneal Wound Healing in HO-2 Null Mice. PLoS ONE 2011, 6, e21180. [Google Scholar] [CrossRef] [PubMed]
- Prame Kumar, K.; Nicholls, A.J.; Wong, C.H.Y. Partners in Crime: Neutrophils and Monocytes/Macrophages in Inflammation and Disease. Cell Tissue Res. 2018, 371, 551–565. [Google Scholar] [CrossRef] [Green Version]
- Chrysanthopoulou, A.; Mitroulis, I.; Apostolidou, E.; Arelaki, S.; Mikroulis, D.; Konstantinidis, T.; Sivridis, E.; Koffa, M.; Giatromanolaki, A.; Boumpas, D.T.; et al. Neutrophil Extracellular Traps Promote Differentiation and Function of Fibroblasts. J. Pathol. 2014, 233, 294–307. [Google Scholar] [CrossRef]
- Phan, S.H. Biology of Fibroblasts and Myofibroblasts. Proc. Am. Thorac. Soc. 2008, 5, 334–337. [Google Scholar] [CrossRef]
- Wilson, S.E. Corneal Myofibroblast Biology and Pathobiology: Generation, Persistence, and Transparency. Exp. Eye Res. 2012, 99, 78–88. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huang, T.; David, L.; Mendoza, V.; Yang, Y.; Villarreal, M.; De, K.; Sun, L.; Fang, X.; López-Casillas, F.; Wrana, J.L.; et al. TGF-β Signalling Is Mediated by Two Autonomously Functioning TβRI:TβRII Pairs. EMBO J. 2011, 30, 1263–1276. [Google Scholar] [CrossRef] [Green Version]
- Walton, K.L.; Johnson, K.E.; Harrison, C.A. Targeting TGF-β Mediated SMAD Signaling for the Prevention of Fibrosis. Front. Pharmacol. 2017, 8, 461. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Słoniecka, M.; Danielson, P. Substance P Induces Fibrotic Changes through Activation of the RhoA/ROCK Pathway in an in Vitro Human Corneal Fibrosis Model. J. Mol. Med. 2019, 97, 1477–1489. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Prudnikova, T.Y.; Rawat, S.J.; Chernoff, J. Molecular Pathways: Targeting the Kinase Effectors of RHO-Family GTPases. Clin. Cancer Res. 2015, 21, 24–29. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yu-Wai-Man, C.; Treisman, R.; Bailly, M.; Khaw, P.T. The Role of the MRTF-A/SRF Pathway in Ocular Fibrosis. Investig. Opthalmol. Vis. Sci. 2014, 55, 4560–4567. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Harvey, S.A.K.; Anderson, S.C.; SundarRaj, N. Downstream Effects of ROCK Signaling in Cultured Human Corneal Stromal Cells: Microarray Analysis of Gene Expression. Investig. Opthalmol. Vis. Sci. 2004, 45, 2168–2176. [Google Scholar] [CrossRef] [PubMed]
- Snijdelaar, D.G.; Dirksen, R.; Slappendel, R.; Crul, B.J.P. Substance P. Eur. J. Pain 2000, 4, 121–135. [Google Scholar] [CrossRef] [PubMed]
- Suvas, S. Role of Substance P Neuropeptide in Inflammation, Wound Healing, and Tissue Homeostasis. J. Immunol. 2017, 199, 1543–1552. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Grgic, I.; Kiss, E.; Kaistha, B.P.; Busch, C.; Kloss, M.; Sautter, J.; Muller, A.; Kaistha, A.; Schmidt, C.; Raman, G.; et al. Renal Fibrosis Is Attenuated by Targeted Disruption of KCa3.1 Potassium Channels. Proc. Natl. Acad. Sci. USA 2009, 106, 14518–14523. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Krstić, J.; Trivanović, D.; Mojsilović, S.; Santibanez, J.F. Transforming Growth Factor-Beta and Oxidative Stress Interplay: Implications in Tumorigenesis and Cancer Progression. Oxid. Med. Cell. Longev. 2015, 2015, 654594. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, L.; Mongan, M.; Meng, Q.; Wang, Q.; Kao, W.; Xia, Y. Corneal Wound Healing Requires IKB Kinase β Signaling in Keratocytes. PLoS ONE 2016, 11, e0151869. [Google Scholar] [CrossRef] [Green Version]
- Stratton, M.S.; Haldar, S.M.; McKinsey, T.A. BRD4 Inhibition for the Treatment of Pathological Organ Fibrosis. F1000Research 2017, 6, 1015. [Google Scholar] [CrossRef] [Green Version]
- Liu, X.-F.; Zhou, D.-D.; Xie, T.; Malik, T.H.; Lu, C.-B.; Li, H.-J.; Wang, F.; Shu, C.; Liu, C.; Lu, C.-W.; et al. Nrf2, a Potential Therapeutic Target against Oxidative Stress in Corneal Diseases. Oxid. Med. Cell. Longev. 2017, 2017, 2326178. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yuan, J.; Luo, K.; Zhang, L.; Cheville, J.C.; Lou, Z. USP10 Regulates P53 Localization and Stability by Deubiquitinating P53. Cell 2010, 140, 384–396. [Google Scholar] [CrossRef] [Green Version]
- Steen, E.H.; Wang, X.; Balaji, S.; Butte, M.J.; Bollyky, P.L.; Keswani, S.G. The Role of the Anti-Inflammatory Cytokine Interleukin-10 in Tissue Fibrosis. Adv. Wound Care 2020, 9, 184–198. [Google Scholar] [CrossRef] [Green Version]
- Ramaesh, T.; Ramaesh, K.; Leask, R.; Springbett, A.; Riley, S.C.; Dhillon, B.; West, J.D. Increased Apoptosis and Abnormal Wound-Healing Responses in the Heterozygous Pax6+/− Mouse Cornea. Investig. Opthalmol. Vis. Sci. 2006, 47, 1911–2171. [Google Scholar] [CrossRef] [PubMed]
- Zheng, K.; Huang, H.; Peng, K.; Cai, J.; Jhanji, V.; Chen, H. Change of Optical Intensity during Healing Process of Corneal Wound on Anterior Segment Optical Coherence Tomography. Sci. Rep. 2016, 6, 32352. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ghatak, S.; Maytin, E.V.; Mack, J.A.; Hascall, V.C.; Atanelishvili, I.; Moreno Rodriguez, R.; Markwald, R.R.; Misra, S. Roles of Proteoglycans and Glycosaminoglycans in Wound Healing and Fibrosis. Int. J. Cell Biol. 2015, 2015, 1–20. [Google Scholar] [CrossRef] [Green Version]
- Shu, D.Y.; Lovicu, F.J. Myofibroblast Transdifferentiation: The Dark Force in Ocular Wound Healing and Fibrosis. Prog. Retin. Eye Res. 2017, 60, 44–65. [Google Scholar] [CrossRef] [PubMed]
- Cintron, C.; Covington, H.I.; Kublin, C.L. Morphologic Analyses of Proteoglycans in Rabbit Corneal Scars. Investig. Ophthalmol. Vis. Sci. 1990, 31, 1789–1798. [Google Scholar]
- Raposo, G.; Stoorvogel, W. Extracellular Vesicles: Exosomes, Microvesicles, and Friends. J. Cell Biol. 2013, 200, 373–383. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Turturici, G.; Tinnirello, R.; Sconzo, G.; Geraci, F. Extracellular Membrane Vesicles as a Mechanism of Cell-to-Cell Communication: Advantages and Disadvantages. Am. J. Physiol. Physiol. 2014, 306, C621–C633. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Das, C.K.; Jena, B.C.; Banerjee, I.; Das, S.; Parekh, A.; Bhutia, S.K.; Mandal, M. Exosome as a Novel Shuttle for Delivery of Therapeutics across Biological Barriers. Mol. Pharm. 2019, 16, 24–40. [Google Scholar] [CrossRef] [PubMed]
- Kuschert, G.S.V.; Hoogewerf, A.J.; Proudfoot, A.E.I.; Chung, C.; Cooke, R.M.; Hubbard, R.E.; Wells, T.N.C.; Sanderson, P.N. Identification of a Glycosaminoglycan Binding Surface on Human Interleukin-8. Biochemistry 1998, 37, 11193–11201. [Google Scholar] [CrossRef]
- Hertsenberg, A.J.; Shojaati, G.; Funderburgh, M.L.; Mann, M.M.; Du, Y.; Funderburgh, J.L. Corneal Stromal Stem Cells Reduce Corneal Scarring by Mediating Neutrophil Infiltration after Wounding. PLoS ONE 2017, 12, e0171712. [Google Scholar] [CrossRef] [Green Version]
- Dyer, D.P.; Thomson, J.M.; Hermant, A.; Jowitt, T.A.; Handel, T.M.; Proudfoot, A.E.I.; Day, A.J.; Milner, C.M. TSG-6 Inhibits Neutrophil Migration via Direct Interaction with the Chemokine CXCL8. J. Immunol. 2014, 192, 2177–2185. [Google Scholar] [CrossRef] [Green Version]
- Shojaati, G.; Khandaker, I.; Funderburgh, M.L.; Mann, M.M.; Basu, R.; Stolz, D.B.; Geary, M.L.; Dos Santos, A.; Deng, S.X.; Funderburgh, J.L. Mesenchymal Stem Cells Reduce Corneal Fibrosis and Inflammation via Extracellular Vesicle-Mediated Delivery of MiRNA. Stem Cells Transl. Med. 2019, 8, 1192–1201. [Google Scholar] [CrossRef]
- Shen, T.; Zheng, Q.-Q.; Shen, J.; Li, Q.-S.; Song, X.-H.; Luo, H.-B.; Hong, C.-Y.; Yao, K. Effects of Adipose-Derived Mesenchymal Stem Cell Exosomes on Corneal Stromal Fibroblast Viability and Extracellular Matrix Synthesis. Chin. Med. J. (Engl). 2018, 131, 704–712. [Google Scholar] [CrossRef]
- Hofmann, T.G.; Stollberg, N.; Schmitz, M.L.; Will, H. HIPK2 Regulates Transforming Growth Factor-Beta-Induced c-Jun NH(2)-Terminal Kinase Activation and Apoptosis in Human Hepatoma Cells. Cancer Res. 2003, 63, 8271–8277. [Google Scholar]
- Shen, T.; Zheng, Q.; Luo, H.; Li, X.; Chen, Z.; Song, Z.; Zhou, G.; Hong, C. Exosomal MiR-19a from Adipose-Derived Stem Cells Suppresses Differentiation of Corneal Keratocytes into Myofibroblasts. Aging (Albany NY) 2020, 12, 4093–4110. [Google Scholar] [CrossRef]
- Lai, Y.; Lee, P.; Lu, C.; Liu, Y.; Wang, S.; Liu, C.; Chang, Y.; Chen, Y.; Su, C.; Li, C.; et al. Thrombospondin 1-induced Exosomal Proteins Attenuate Hypoxia-induced Paraptosis in Corneal Epithelial Cells and Promote Wound Healing. FASEB J. 2021, 35, e21200. [Google Scholar] [CrossRef]
- Tao, H.; Chen, X.; Cao, H.; Zheng, L.; Li, Q.; Zhang, K.; Han, Z.; Han, Z.-C.; Guo, Z.; Li, Z.; et al. Mesenchymal Stem Cell-Derived Extracellular Vesicles for Corneal Wound Repair. Stem Cells Int. 2019, 2019, 5738510. [Google Scholar] [CrossRef] [Green Version]
- Jablonska, E.; Garley, M.; Surazynski, A.; Grubczak, K.; Iwaniuk, A.; Borys, J.; Moniuszko, M.; Ratajczak-Wrona, W. Neutrophil Extracellular Traps (NETs) Formation Induced by TGF-β in Oral Lichen Planus—Possible Implications for the Development of Oral Cancer. Immunobiology 2020, 225, 151901. [Google Scholar] [CrossRef] [PubMed]
- Mun, Y.; Hwang, J.S.; Shin, Y.J. Role of Neutrophils on the Ocular Surface. Int. J. Mol. Sci. 2021, 22, 10386. [Google Scholar] [CrossRef] [PubMed]
- Yin, H.; Lu, Q.; Wang, X.; Majumdar, S.; Jun, A.S.; Stark, W.J.; Grant, M.P.; Elisseeff, J.H. Tissue-Derived Microparticles Reduce Inflammation and Fibrosis in Cornea Wounds. Acta Biomater. 2019, 85, 192–202. [Google Scholar] [CrossRef] [PubMed]
- Yamaguchi, T. Inflammatory Response in Dry Eye. Investig. Opthalmol. Vis. Sci. 2018, 59, DES192–DES199. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Peterson, J.L.; Ceresa, B.P. Epidermal Growth Factor Receptor Expression in the Corneal Epithelium. Cells 2021, 10, 2409. [Google Scholar] [CrossRef] [PubMed]
- Alto, L.T.; Terman, J.R. Semaphorins and Their Signaling Mechanisms. In Semaphorin Signaling, Methods in Molecular Biology; Terman, J.R., Ed.; Humana Press: New York, NY, USA, 2017; Volume 1493. [Google Scholar] [CrossRef]
- Jeon, K.-I.; Nehrke, K.; Huxlin, K.R. Semaphorin 3A Potentiates the Profibrotic Effects of Transforming Growth Factor-Β1 in the Cornea. Biochem. Biophys. Res. Commun. 2020, 521, 333–339. [Google Scholar] [CrossRef] [PubMed]
- Morishige, N.; Ko, J.-A.; Morita, Y.; Nishida, T. Expression of Semaphorin 3A in the Rat Corneal Epithelium during Wound Healing. Biochem. Biophys. Res. Commun. 2010, 395, 451–457. [Google Scholar] [CrossRef] [PubMed]
- Yamazaki, R.; Yamazoe, K.; Yoshida, S.; Hatou, S.; Inagaki, E.; Okano, H.; Tsubota, K.; Shimmura, S. The Semaphorin 3A Inhibitor SM-345431 Preserves Corneal Nerve and Epithelial Integrity in a Murine Dry Eye Model. Sci. Rep. 2017, 7, 15584. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Takayama, K.; Suzuki, T.; Fujimura, T.; Takahashi, S.; Inoue, S. Association of USP10 with G3BP2 Inhibits P53 Signaling and Contributes to Poor Outcome in Prostate Cancer. Mol. Cancer Res. 2018, 16, 846–856. [Google Scholar] [CrossRef] [Green Version]
- Zhang, H.; Zhang, S.; He, H.; Zhang, C.; Yu, D.; Shao, R. Downregulation of G3BPs Inhibits the Growth, Migration and Invasion of Human Lung Carcinoma H1299 Cells by Suppressing the Src/FAK-Associated Signaling Pathway. Cancer Gene Ther. 2013, 20, 622–629. [Google Scholar] [CrossRef]
- Gillespie, S.R.; Tedesco, L.J.; Wang, L.; Bernstein, A.M. The Deubiquitinase USP10 Regulates Integrin Beta1 and Beta5 and Fibrotic Wound Healing. J. Cell Sci. 2017, 130, 3481–3495. [Google Scholar] [CrossRef] [Green Version]
- Boumil, E.F.; Castro, N.; Phillips, A.T.; Chatterton, J.E.; McCauley, S.M.; Wolfson, A.D.; Shmushkovich, T.; Ridilla, M.; Bernstein, A.M. USP10 Targeted Self-Deliverable SiRNA to Prevent Scarring in the Cornea. Mol. Ther. Nucleic Acids 2020, 21, 1029–1043. [Google Scholar] [CrossRef]
- Morales, P.; Garneau, L.; Klein, H.; Lavoie, M.-F.; Parent, L.; Sauvé, R. Contribution of the KCa3.1 Channel–Calmodulin Interactions to the Regulation of the KCa3.1 Gating Process. J. Gen. Physiol. 2013, 142, 37–60. [Google Scholar] [CrossRef] [Green Version]
- Pérez-García, M.T.; Cidad, P.; López-López, J.R. The Secret Life of Ion Channels: Kv1.3 Potassium Channels and Proliferation. Am. J. Physiol. Physiol. 2018, 314, C27–C42. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Girault, A.; Chebli, J.; Privé, A.; Trinh, N.T.N.; Maillé, E.; Grygorczyk, R.; Brochiero, E. Complementary Roles of KCa3.1 Channels and Β1-Integrin during Alveolar Epithelial Repair. Respir. Res. 2015, 16, 100. [Google Scholar] [CrossRef] [PubMed]
- Arcangeli, A.; Becchetti, A. Complex Functional Interaction between Integrin Receptors and Ion Channels. Trends Cell Biol. 2006, 16, 631–639. [Google Scholar] [CrossRef] [PubMed]
- Takada, Y.; Ye, X.; Simon, S. The integrins. Genome. Biol. 2007, 8, 215. [Google Scholar] [CrossRef] [Green Version]
- Brown, B.M.; Pressley, B.; Wulff, H. KCa3.1 Channel Modulators as Potential Therapeutic Compounds for Glioblastoma. Curr. Neuropharmacol. 2018, 16, 618–626. [Google Scholar] [CrossRef]
- Anumanthan, G.; Gupta, S.; Fink, M.K.; Hesemann, N.P.; Bowles, D.K.; McDaniel, L.M.; Muhammad, M.; Mohan, R.R. KCa3.1 Ion Channel: A Novel Therapeutic Target for Corneal Fibrosis. PLoS ONE 2018, 13, e0192145. [Google Scholar] [CrossRef] [Green Version]
- Fujimoto, S.; Hayashi, R.; Hara, S.; Sasamoto, Y.; Harrington, J.; Tsujikawa, M.; Nishida, K. KLF4 Prevents Epithelial to Mesenchymal Transition in Human Corneal Epithelial Cells via Endogenous TGF-Β2 Suppression. Regen. Ther. 2019, 11, 249–257. [Google Scholar] [CrossRef]
- Gupta, S.; Martin, L.M.; Sinha, N.R.; Smith, K.E.; Sinha, P.R.; Dailey, E.M.; Hesemann, N.P.; Mohan, R.R. Role of Inhibitor of Differentiation 3 Gene in Cellular Differentiation of Human Corneal Stromal Fibroblasts. Mol. Vis. 2020, 26, 742. [Google Scholar]
- Saitoh, M.; Miyazawa, K. Transcriptional and Post-Transcriptional Regulation in TGF—Mediated Epithelial-Mesenchymal Transition. J. Biochem. 2012, 151, 563–571. [Google Scholar] [CrossRef] [Green Version]
- Xu, J.; Lamouille, S.; Derynck, R. TGF-β-Induced Epithelial to Mesenchymal Transition. Cell Res. 2009, 19, 156–172. [Google Scholar] [CrossRef]
- Chaudhary, J.; Sadler-Riggleman, I.; Ague, J.M.; Skinner, M.K. The Helix-Loop-Helix Inhibitor of Differentiation (ID) Proteins Induce Post-Mitotic Terminally Differentiated Sertoli Cells to Re-Enter the Cell Cycle and Proliferate. Biol. Reprod. 2005, 72, 1205–1217. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yan, X.; Liao, H.; Cheng, M.; Shi, X.; Lin, X.; Feng, X.-H.; Chen, Y.-G. Smad7 Protein Interacts with Receptor-Regulated Smads (R-Smads) to Inhibit Transforming Growth Factor-β (TGF-β)/Smad Signaling. J. Biol. Chem. 2016, 291, 382–392. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kavsak, P.; Rasmussen, R.K.; Causing, C.G.; Bonni, S.; Zhu, H.; Thomsen, G.H.; Wrana, J.L. Smad7 Binds to Smurf2 to Form an E3 Ubiquitin Ligase That Targets the TGFβ Receptor for Degradation. Mol. Cell 2000, 6, 1365–1375. [Google Scholar] [CrossRef]
- Gupta, S.; Rodier, J.T.; Sharma, A.; Giuliano, E.A.; Sinha, P.R.; Hesemann, N.P.; Ghosh, A.; Mohan, R.R. Targeted AAV5-Smad7 Gene Therapy Inhibits Corneal Scarring In Vivo. PLoS ONE 2017, 12, e0172928. [Google Scholar] [CrossRef] [Green Version]
- O’Brien, J.; Hayder, H.; Zayed, Y.; Peng, C. Overview of MicroRNA Biogenesis, Mechanisms of Actions, and Circulation. Front. Endocrinol. (Lausanne) 2018, 9, 402. [Google Scholar] [CrossRef] [Green Version]
- An, J.; Chen, X.; Chen, W.; Liang, R.; Reinach, P.S.; Yan, D.; Tu, L. MicroRNA Expression Profile and the Role of MiR-204 in Corneal Wound Healing. Investig. Opthalmol. Vis. Sci. 2015, 56, 3673–3683. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.; Li, W.; Zang, X.; Chen, N.; Liu, T.; Tsonis, P.A.; Huang, Y. MicroRNA-204-5p Regulates Epithelial-to-Mesenchymal Transition during Human Posterior Capsule Opacification by Targeting SMAD4. Investig. Opthalmol. Vis. Sci. 2013, 54, 323–332. [Google Scholar] [CrossRef] [Green Version]
- Li, Y.; Zhao, Z.; Xu, C.; Zhou, Z.; Zhu, Z.; You, T. HMGA2 Induces Transcription Factor Slug Expression to Promote Epithelial-to-Mesenchymal Transition and Contributes to Colon Cancer Progression. Cancer Lett. 2014, 355, 130–140. [Google Scholar] [CrossRef]
- Medici, D.; Hay, E.D.; Olsen, B.R. Snail and Slug Promote Epithelial-Mesenchymal Transition through β-Catenin–T-Cell Factor-4-Dependent Expression of Transforming Growth Factor-Β3. Mol. Biol. Cell 2008, 19, 4875–4887. [Google Scholar] [CrossRef] [Green Version]
- Lu, Y.; Tai, P.W.L.; Ai, J.; Gessler, D.J.; Su, Q.; Yao, X.; Zheng, Q.; Zamore, P.D.; Xu, X.; Gao, G. Transcriptome Profiling of Neovascularized Corneas Reveals MiR-204 as a Multi-Target Biotherapy Deliverable by RAAVs. Mol. Ther. Nucleic Acids 2018, 10, 349–360. [Google Scholar] [CrossRef] [Green Version]
- Ratuszny, D.; Gras, C.; Bajor, A.; Börger, A.-K.; Pielen, A.; Börgel, M.; Framme, C.; Blasczyk, R.; Figueiredo, C. MiR-145 Is a Promising Therapeutic Target to Prevent Cornea Scarring. Hum. Gene Ther. 2015, 26, 698–707. [Google Scholar] [CrossRef] [PubMed]
- Xu, N.; Papagiannakopoulos, T.; Pan, G.; Thomson, J.A.; Kosik, K.S. MicroRNA-145 Regulates OCT4, SOX2, and KLF4 and Represses Pluripotency in Human Embryonic Stem Cells. Cell 2009, 137, 647–658. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sun, H.; Peng, Z.; Tang, H.; Xie, D.; Jia, Z.; Zhong, L.; Zhao, S.; Ma, Z.; Gao, Y.; Zeng, L.; et al. Loss of KLF4 and Consequential Downregulation of Smad7 Exacerbate Oncogenic TGF-β Signaling in and Promote Progression of Hepatocellular Carcinoma. Oncogene 2017, 36, 2957–2968. [Google Scholar] [CrossRef]
- Li, X.M.; Kim, S.J.; Hong, D.-K.; Jung, K.E.; Choi, C.W.; Seo, Y.-J.; Lee, J.-H.; Lee, Y.; Kim, C.-D. KLF4 Suppresses the Tumor Activity of Cutaneous Squamous Cell Carcinoma (SCC) Cells via the Regulation of SMAD Signaling and SOX2 Expression. Biochem. Biophys. Res. Commun. 2019, 516, 1110–1115. [Google Scholar] [CrossRef]
- Gjymishka, A.; Pi, L.; Oh, S.-H.; Jorgensen, M.; Liu, C.; Protopapadakis, Y.; Patel, A.; Petersen, B.E. MiR-133b Regulation of Connective Tissue Growth Factor. Am. J. Pathol. 2016, 186, 1092–1102. [Google Scholar] [CrossRef] [Green Version]
- Grotendorst, G.R.; Duncan, M.R. Individual Domains of Connective Tissue Growth Factor Regulate Fibroblast Proliferation and Myofibroblast Differentiation. FASEB J. 2005, 19, 729–738. [Google Scholar] [CrossRef] [PubMed]
- Zhao, X.; Song, W.; Chen, Y.; Liu, S.; Ren, L. Collagen-Based Materials Combined with MicroRNA for Repairing Cornea Wounds and Inhibiting Scar Formation. Biomater. Sci. 2019, 7, 51–62. [Google Scholar] [CrossRef]
- Ashby, B.D.; Garrett, Q.; Willcox, M.D.P. Corneal Injuries and Wound Healing—Review of Processes and Therapies. Austin J. Clin. Ophthalmol. 2014, 1, 1017. [Google Scholar]
- Landén, N.X.; Li, D.; Ståhle, M. Transition from Inflammation to Proliferation: A Critical Step during Wound Healing. Cell. Mol. Life Sci. 2016, 73, 3861–3885. [Google Scholar] [CrossRef] [Green Version]
- Zhou, Q.; Wang, Y.; Yang, L.; Wang, Y.; Chen, P.; Wang, Y.; Dong, X.; Xie, L. Histone Deacetylase Inhibitors Blocked Activation and Caused Senescence of Corneal Stromal cells. Mol. Vis. 2008, 14, 2556–2565. [Google Scholar]
- Sun, G.; Reddy, M.A.; Yuan, H.; Lanting, L.; Kato, M.; Natarajan, R. Epigenetic Histone Methylation Modulates Fibrotic Gene Expression. J. Am. Soc. Nephrol. 2010, 21, 2069–2080. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lu, C.; Sidoli, S.; Kulej, K.; Ross, K.; Wu, C.H.; Garcia, B.A. Coordination between TGF-β Cellular Signaling and Epigenetic Regulation during Epithelial to Mesenchymal Transition. Epigenetics Chromatin 2019, 12, 11. [Google Scholar] [CrossRef] [PubMed]
- Tang, J.; Yan, H.; Zhuang, S. Histone Deacetylases as Targets for Treatment of Multiple Diseases. Clin. Sci. 2013, 124, 651–662. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhou, Q.; Yang, L.; Wang, Y.; Qu, M.; Chen, P.; Wang, Y.; Xie, L.; Zhao, J.; Wang, Y. TGFβ Mediated Transition of Corneal Fibroblasts from a Proinflammatory State to a Profibrotic State through Modulation of Histone Acetylation. J. Cell. Physiol. 2010, 224, 135–143. [Google Scholar] [CrossRef] [PubMed]
- Xiao, W.; Chen, X.; Liu, X.; Luo, L.; Ye, S.; Liu, Y. Trichostatin A, a Histone Deacetylase Inhibitor, Suppresses Proliferation and Epithelial–Mesenchymal Transition in Retinal Pigment Epithelium Cells. J. Cell. Mol. Med. 2014, 18, 646–655. [Google Scholar] [CrossRef] [PubMed]
- Sharma, A.; Mehan, M.M.; Sinha, S.; Cowden, J.W.; Mohan, R.R. Trichostatin A Inhibits Corneal Haze In Vitro and In Vivo. Investig. Opthalmol. Vis. Sci. 2009, 50, 2695–2701. [Google Scholar] [CrossRef] [Green Version]
- Donnelly, K.S.; Giuliano, E.A.; Sharma, A.; Mohan, R.R. Suberoylanilide Hydroxamic Acid (Vorinostat): Its Role on Equine Corneal Fibrosis and Matrix Metalloproteinase Activity. Vet. Ophthalmol. 2014, 17, 61–68. [Google Scholar] [CrossRef]
- Etheredge, L.; Kane, B.P.; Hassell, J.R. The Effect of Growth Factor Signaling on Keratocytes In Vitro and Its Relationship to the Phases of Stromal Wound Repair. Investig. Opthalmol. Vis. Sci. 2009, 50, 3128–3136. [Google Scholar] [CrossRef]
- Sarenac, T.; Trapecar, M.; Gradisnik, L.; Rupnik, M.S.; Pahor, D. Single-Cell Analysis Reveals IGF-1 Potentiation of Inhibition of the TGF-β/Smad Pathway of Fibrosis in Human Keratocytes In Vitro. Sci. Rep. 2016, 6, 34373. [Google Scholar] [CrossRef] [Green Version]
- Ghiasi, Z.; Gray, T.; Tran, P.; Dubielzig, R.; Murphy, C.; McCartney, D.L.; Reid, T.W. The Effect of Topical Substance-P Plus Insulin-like Growth Factor-1 (IGF-1) on Epithelial Healing After Photorefractive Keratectomy in Rabbits. Transl. Vis. Sci. Technol. 2018, 7, 12. [Google Scholar] [CrossRef]
- Chen, Y.-J.; Huang, Y.-S.; Chen, J.-T.; Chen, Y.-H.; Tai, M.-C.; Chen, C.-L.; Liang, C.-M. Protective Effects of Glucosamine on Oxidative-Stress and Ischemia/Reperfusion-Induced Retinal Injury. Investig. Ophthalmol. Vis. Sci. 2015, 56, 1506–1516. [Google Scholar] [CrossRef] [PubMed]
- Pham, T.; Cornea, A.; Jenkins, A.; Blick, K.E.; Scofield, R.H. Oral Glucosamine in Doses Used to Treat Osteoarthritis Worsens Insulin Resistance. Am. J. Med. Sci. 2007, 333, 333–339. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Esfandiari, H.; Loewen, N.A. Effect of Glucosamine on Intraocular Pressure. In Handbook of Nutrition, Diet, and the Eye, 2nd ed.; Elsevier: Amsterdam, The Netherlands, 2019. [Google Scholar] [CrossRef]
- Park, J.; Lee, S.-Y.; Ooshima, A.; Yang, K.-M.; Kang, J.M.; Kim, Y.-W.; Kim, S.-J. Glucosamine Hydrochloride Exerts a Protective Effect against Unilateral Ureteral Obstruction-Induced Renal Fibrosis by Attenuating TGF-β Signaling. J. Mol. Med. 2013, 91, 1273–1284. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, D.-F.; Yang, H.-J.; GU, J.-Q.; Cao, Y.-L.; Meng, X.; Wang, X.-L.; Lin, Y.-C.; Gao, M. Suppression of Phosphatase and Tensin Homolog Protects Insulin-Resistant Cells from Apoptosis. Mol. Med. Rep. 2015, 12, 2695–2700. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Han, M.; Bernier, M.; Zheng, B.; Sun, S.; Su, M.; Zhang, R.; Fu, J.; Wen, J. Krüppel-like Factor 4 Promotes Differentiation by Transforming Growth Factor-β Receptor-Mediated Smad and P38 MAPK Signaling in Vascular Smooth Muscle Cells. J. Biol. Chem. 2010, 285, 17846–17856. [Google Scholar] [CrossRef] [Green Version]
- He, M.; Zheng, B.; Zhang, Y.; Zhang, X.-H.; Wang, C.; Yang, Z.; Sun, Y.; Wu, X.-L.; Wen, J.-K. KLF4 Mediates the Link between TGF-Β1-Induced Gene Transcription and H3 Acetylation in Vascular Smooth Muscle Cells. FASEB J. 2015, 29, 4059–4070. [Google Scholar] [CrossRef]
- Zhang, F.; Su, K.; Yang, X.; Bowe, D.B.; Paterson, A.J.; Kudlow, J.E. O-GlcNAc Modification Is an Endogenous Inhibitor of the Proteasome. Cell 2003, 115, 715–725. [Google Scholar] [CrossRef] [Green Version]
- Swamynathan, S.; Buela, K.-A.; Kinchington, P.; Lathrop, K.L.; Misawa, H.; Hendricks, R.L.; Swamynathan, S.K. Klf4 Regulates the Expression of Slurp1, Which Functions as an Immunomodulatory Peptide in the Mouse Cornea. Investig. Opthalmol. Vis. Sci. 2012, 53, 8433–8446. [Google Scholar] [CrossRef]
- Azad, A.K.; Sermsintham, N.; Chandrkrachang, S.; Stevens, W.F. Chitosan Membrane as a Wound-Healing Dressing: Characterization and Clinical Application. J. Biomed. Mater. Res. 2004, 69, 216–222. [Google Scholar] [CrossRef]
- Alsarra, I.A. Chitosan Topical Gel Formulation in the Management of Burn Wounds. Int. J. Biol. Macromol. 2009, 45, 16–21. [Google Scholar] [CrossRef]
- Fischak, C.; Klaus, R.; Werkmeister, R.M.; Hohenadl, C.; Prinz, M.; Schmetterer, L.; Garhöfer, G. Effect of Topically Administered Chitosan N Acetylcysteine on Corneal Wound Healing in a Rabbit Model. J. Ophthalmol. 2017, 2017, 5192924. [Google Scholar] [CrossRef] [Green Version]
- Zahir-Jouzdani, F.; Mahbod, M.; Soleimani, M.; Vakhshiteh, F.; Arefian, E.; Shahosseini, S.; Dinarvand, R.; Atyabi, F. Chitosan and Thiolated Chitosan: Novel Therapeutic Approach for Preventing Corneal Haze after Chemical Injuries. Carbohydr. Polym. 2018, 179, 42–49. [Google Scholar] [CrossRef] [PubMed]
- Chen, Q.; Lu, H.; Yang, H. Chitosan Inhibits Fibroblasts Growth in Achilles Tendon via TGF-Β1/Smad3 Pathway by MiR-29b. Int. J. Clin. Exp. Pathol. 2014, 7, 8462–8470. [Google Scholar] [PubMed]
- Gan, F.; Liu, Q.; Liu, Y.; Huang, D.; Pan, C.; Song, S.; Huang, K. Lycium Barbarum Polysaccharides Improve CCl4-Induced Liver Fibrosis, Inflammatory Response and TLRs/NF-KB Signaling Pathway Expression in Wistar Rats. Life Sci. 2018, 192, 205–212. [Google Scholar] [CrossRef] [PubMed]
- Du, S.; Han, B.; Li, K.; Zhang, X.; Sha, X.; Gao, L. Lycium Barbarum Polysaccharides Protect Rat Corneal Epithelial Cells against Ultraviolet B-Induced Apoptosis by Attenuating the Mitochondrial Pathway and Inhibiting JNK Phosphorylation. Biomed Res. Int. 2017, 2017, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Kwok, S.S.; Wong, F.S.-Y.; Shih, K.C.; Chan, Y.-K.; Bu, Y.; Chan, T.C.-Y.; Ng, A.L.-K.; Lo, A.C.-Y.; Tong, L.; Yam, G.H.-F.; et al. Lycium Barbarum Polysaccharide Suppresses Expression of Fibrotic Proteins in Primary Human Corneal Fibroblasts. J. Clin. Med. 2020, 9, 3572. [Google Scholar] [CrossRef]
- Richter, K.; Konzack, A.; Pihlajaniemi, T.; Heljasvaara, R.; Kietzmann, T. Redox-Fibrosis: Impact of TGFβ1 on ROS Generators, Mediators and Functional Consequences. Redox Biol. 2015, 6, 344–352. [Google Scholar] [CrossRef]
- Cucoranu, I.; Clempus, R.; Dikalova, A.; Phelan, P.J.; Ariyan, S.; Dikalov, S.; Sorescu, D. NAD(P)H Oxidase 4 Mediates Transforming Growth Factor-Β1–Induced Differentiation of Cardiac Fibroblasts Into Myofibroblasts. Circ. Res. 2005, 97, 900–907. [Google Scholar] [CrossRef] [Green Version]
- Liu, R.-M.; Desai, L.P. Reciprocal Regulation of TGF-β and Reactive Oxygen Species: A Perverse Cycle for Fibrosis. Redox Biol. 2015, 6, 565–577. [Google Scholar] [CrossRef] [Green Version]
- Barnes, J.L.; Gorin, Y. Myofibroblast Differentiation during Fibrosis: Role of NAD(P)H Oxidases. Kidney Int. 2011, 79, 944–956. [Google Scholar] [CrossRef] [Green Version]
- Bourji, K.; Meyer, A.; Chatelus, E.; Pincemail, J.; Pigatto, E.; Defraigne, J.-O.; Singh, F.; Charlier, C.; Geny, B.; Gottenberg, J.-E.; et al. High Reactive Oxygen Species in Fibrotic and Nonfibrotic Skin of Patients with Diffuse Cutaneous Systemic Sclerosis. Free Radic. Biol. Med. 2015, 87, 282–289. [Google Scholar] [CrossRef]
- Serafini, M.M.; Catanzaro, M.; Fagiani, F.; Simoni, E.; Caporaso, R.; Dacrema, M.; Romanoni, I.; Govoni, S.; Racchi, M.; Daglia, M.; et al. Modulation of Keap1/Nrf2/ARE Signaling Pathway by Curcuma- and Garlic-Derived Hybrids. Front. Pharmacol. 2020, 10, 1597. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Qu, M.; Zhang, X.; Hu, X.; Dong, M.; Pan, X.; Bian, J.; Zhou, Q. BRD4 Inhibitor JQ1 Inhibits and Reverses Mechanical Injury-Induced Corneal Scarring. Cell Death Discov. 2018, 4, 64. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Balser, C.; Wolf, A.; Herb, M.; Langmann, T. Co-Inhibition of PGF and VEGF Blocks Their Expression in Mononuclear Phagocytes and Limits Neovascularization and Leakage in the Murine Retina. J. Neuroinflamm. 2019, 16, 1–12. [Google Scholar] [CrossRef]
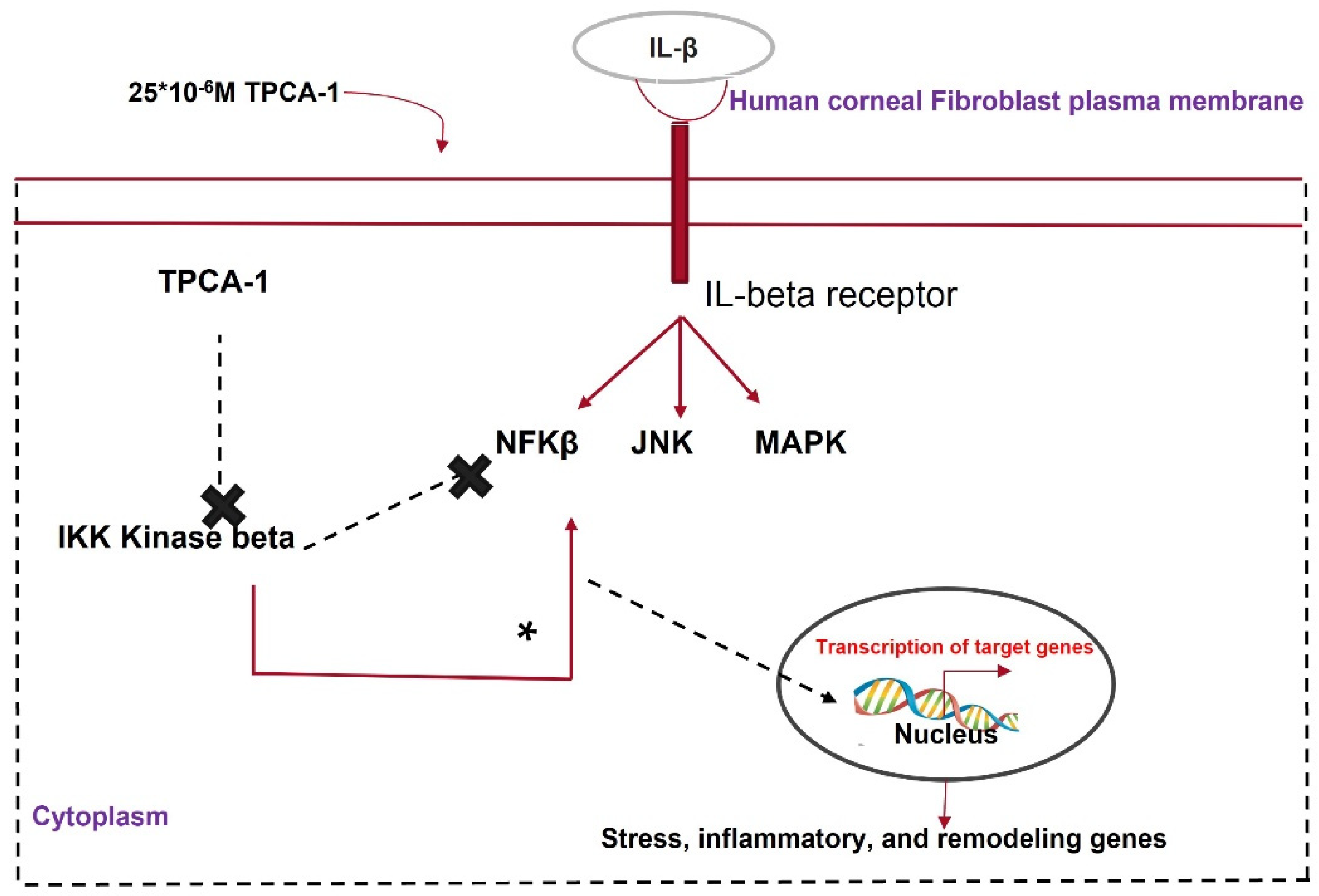
- Zhang, W.; Chen, J.; Qu, M.; Backman, L.J.; Zhang, A.; Liu, H.; Zhang, X.; Zhou, Q.; Danielson, P. Sustained Release of TPCA-1 from Silk Fibroin Hydrogels Preserves Keratocyte Phenotype and Promotes Corneal Regeneration by Inhibiting Interleukin-1 β Signaling. Adv. Healthc. Mater. 2020, 9, e2000591. [Google Scholar] [CrossRef] [PubMed]
- Kondo, Y.; Fukuda, K.; Adachi, T.; Nishida, T. Inhibition by a Selective IκB Kinase-2 Inhibitor of Interleukin-1–Induced Collagen Degradation by Corneal Fibroblasts in Three-Dimensional Culture. Investig. Opthalmol. Vis. Sci. 2008, 49, 4850–4857. [Google Scholar] [CrossRef]
- Mulholland, B.; Tuft, S.J.; Khaw, P.T. Matrix Metalloproteinase Distribution during Early Corneal Wound Healing. Eye 2005, 19, 584–588. [Google Scholar] [CrossRef]
- Pardo, A.; Cabrera, S.; Maldonado, M.; Selman, M. Role of Matrix Metalloproteinases in the Pathogenesis of Idiopathic Pulmonary Fibrosis. Respir. Res. 2016, 17, 23. [Google Scholar] [CrossRef] [Green Version]
- Stevenson, R.W.; Wilson, W.S. Drug-Induced Depletion of Acetylcholine in the Rabbit Corneal Epithelium. Biochem. Pharmacol. 1974, 23, 3449–3457. [Google Scholar] [CrossRef]
- Chernyavsky, A.I.; Galitovskiy, V.; Shchepotin, I.B.; Jester, J.V.; Grando, S.A. The Acetylcholine Signaling Network of Corneal Epithelium and Its Role in Regulation of Random and Directional Migration of Corneal Epithelial Cells. Investig. Ophthalmol. Vis. Sci. 2014, 55, 6921–6933. [Google Scholar] [CrossRef] [Green Version]
- Öztürk, F.; Kurt, E.; Inan, Ü.Ü.; Emiroglu, L.; Ilker, S.S. The Effects of Acetylcholine and Propolis Extract on Corneal Epithelial Wound Healing in Rats. Cornea 1999, 18, 466–471. [Google Scholar] [CrossRef]
- Słoniecka, M.; Backman, L.J.; Danielson, P. Acetylcholine Enhances Keratocyte Proliferation through Muscarinic Receptor Activation. Int. Immunopharmacol. 2015, 29, 57–62. [Google Scholar] [CrossRef] [Green Version]
- Gubbiotti, M.A.; Vallet, S.D.; Ricard-Blum, S.; Iozzo, R.V. Decorin Interacting Network: A Comprehensive Analysis of Decorin-Binding Partners and Their Versatile Functions. Matrix Biol. 2016, 55, 7–21. [Google Scholar] [CrossRef] [PubMed]
- Mohan, R.R.; Gupta, R.; Mehan, M.K.; Cowden, J.W.; Sinha, S. Decorin Transfection Suppresses Profibrogenic Genes and Myofibroblast Formation in Human Corneal Fibroblasts. Exp. Eye Res. 2010, 91, 238–245. [Google Scholar] [CrossRef] [PubMed]
- ABDEL-WAHAB, N.; WICKS, S.J.; MASON, R.M.; CHANTRY, A. Decorin Suppresses Transforming Growth Factor-β-Induced Expression of Plasminogen Activator Inhibitor-1 in Human Mesangial Cells through a Mechanism That Involves Ca2+-Dependent Phosphorylation of Smad2 at Serine-240. Biochem. J. 2002, 362, 362–643. [Google Scholar] [CrossRef]
- Chouhan, G.; Moakes, R.J.A.; Esmaeili, M.; Hill, L.J.; deCogan, F.; Hardwicke, J.; Rauz, S.; Logan, A.; Grover, L.M. A Self-Healing Hydrogel Eye Drop for the Sustained Delivery of Decorin to Prevent Corneal Scarring. Biomaterials 2019, 210, 41–50. [Google Scholar] [CrossRef] [PubMed]
- Hill, L.J.; Moakes, R.J.A.; Vareechon, C.; Butt, G.; Ng, A.; Brock, K.; Chouhan, G.; Vincent, R.C.; Abbondante, S.; Williams, R.; et al. Sustained release of decorin to the surface of the eye enables scarless corneal regeneration. npj Regen. Med. 2018, 3, 23. [Google Scholar] [CrossRef] [Green Version]
- Słoniecka, M.; Danielson, P. Acetylcholine Decreases Formation of Myofibroblasts and Excessive Extracellular Matrix Production in an in Vitro Human Corneal Fibrosis Model. J. Cell. Mol. Med. 2020, 24, 4850–4862. [Google Scholar] [CrossRef] [Green Version]
- Chaurasia, S.; Lim, R.; Lakshminarayanan, R.; Mohan, R. Nanomedicine Approaches for Corneal Diseases. J. Funct. Biomater. 2015, 6, 277–298. [Google Scholar] [CrossRef] [Green Version]
- Pissuwan, D.; Niidome, T.; Cortie, M.B. The Forthcoming Applications of Gold Nanoparticles in Drug and Gene Delivery Systems. J. Control. Release 2011, 149, 65–71. [Google Scholar] [CrossRef] [PubMed]
- Tandon, A.; Sharma, A.; Rodier, J.T.; Klibanov, A.M.; Rieger, F.G.; Mohan, R.R. BMP7 Gene Transfer via Gold Nanoparticles into Stroma Inhibits Corneal Fibrosis In Vivo. PLoS ONE 2013, 8, e66434. [Google Scholar] [CrossRef] [Green Version]
- Bhatta, R.S.; Chandasana, H.; Chhonker, Y.S.; Rathi, C.; Kumar, D.; Mitra, K.; Shukla, P.K. Mucoadhesive Nanoparticles for Prolonged Ocular Delivery of Natamycin: In Vitro and Pharmacokinetics Studies. Int. J. Pharm. 2012, 432, 105–112. [Google Scholar] [CrossRef] [PubMed]
- Sharma, A.; Tandon, A.; Tovey, J.C.K.; Gupta, R.; Robertson, J.D.; Fortune, J.A.; Klibanov, A.M.; Cowden, J.W.; Rieger, F.G.; Mohan, R.R. Polyethylenimine-Conjugated Gold Nanoparticles: Gene Transfer Potential and Low Toxicity in the Cornea. Nanomed. Nanotechnol. Biol. Med. 2011, 7, 505–513. [Google Scholar] [CrossRef] [Green Version]
- Sriram, S.; Robinson, P.; Pi, L.; Lewin, A.S.; Schultz, G. Triple Combination of SiRNAs Targeting TGFβ1, TGFβR2, and CTGF Enhances Reduction of Collagen I and Smooth Muscle Actin in Corneal Fibroblasts. Investig. Opthalmol. Vis. Sci. 2013, 54, 8214–8223. [Google Scholar] [CrossRef]
- Zahir-Jouzdani, F.; Soleimani, M.; Mahbod, M.; Mottaghitalab, F.; Vakhshite, F.; Arefian, E.; Shahhoseini, S.; Dinarvand, R.; Atyabi, F. Corneal Chemical Burn Treatment through a Delivery System Consisting of TGF-Β1 SiRNA: In Vitro and In Vivo. Drug Deliv. Transl. Res. 2018, 8, 1127–1138. [Google Scholar] [CrossRef]
- Silva, R.O.; da Costa, B.L.; da Silva, F.R.; da Silva, C.N.; de Paiva, M.B.; Dourado, L.F.N.; Malachias, A.; de Souza Araújo, A.A.; Nunes, P.S.; Silva-Cunha, A. Treatment for Chemical Burning Using Liquid Crystalline Nanoparticles as an Ophthalmic Delivery System for Pirfenidone. Int. J. Pharm. 2019, 568, 118466. [Google Scholar] [CrossRef]
- Kramerov, A.A.; Shah, R.; Ding, H.; Holler, E.; Turjman, S.; Rabinowitz, Y.S.; Ghiam, S.; Maguen, E.; Svendsen, C.N.; Saghizadeh, M.; et al. Novel Nanopolymer RNA Therapeutics Normalize Human Diabetic Corneal Wound Healing and Epithelial Stem Cells. Nanomed. Nanotechnol. Biol. Med. 2021, 32, 102332. [Google Scholar] [CrossRef]
- Ma, X.-Y.; Bao, H.-J.; Cui, L.; Zou, J. The Graft of Autologous Adipose-Derived Stem Cells in the Corneal Stromal after Mechanic Damage. PLoS ONE 2013, 8, e76103. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cejkova, J.; Cejka, C.; Trosan, P.; Zajicova, A.; Sykova, E.; Holan, V. Treatment of Alkali-Injured Cornea by Cyclosporine A-Loaded Electrospun Nanofibers—An Alternative Mode of Therapy. Exp. Eye Res. 2016, 147, 128–137. [Google Scholar] [CrossRef]
- Duan, X.; Sheardown, H. Dendrimer Crosslinked Collagen as a Corneal Tissue Engineering Scaffold: Mechanical Properties and Corneal Epithelial Cell Interactions. Biomaterials 2006, 27, 4608–4617. [Google Scholar] [CrossRef] [PubMed]
- Ibrahim Al-Mashahedah, A.M.; Kanwar, R.K.; Kanwar, J.R. Utility of Nanomedicine Targeting Scar-Forming Myofibroblasts to Attenuate Corneal Scarring and Haze. Nanomedicine 2019, 14, 1049–1072. [Google Scholar] [CrossRef] [PubMed]
- Swaminathan, S.; Vavia, P.R.; Trotta, F.; Cavalli, R. Nanosponges Encapsulating Dexamethasone for Ocular Delivery: Formulation Design, Physicochemical Characterization, Safety and Corneal Permeability Assessment. J. Biomed. Nanotechnol. 2013, 9, 998–1007. [Google Scholar] [CrossRef]
- Weng, Y.; Liu, J.; Jin, S.; Guo, W.; Liang, X.; Hu, Z. Nanotechnology-Based Strategies for Treatment of Ocular Disease. Acta Pharm. Sin. B 2017, 7, 281–291. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Akbari, A.; Jabbari, N.; Sharifi, R.; Ahmadi, M.; Vahhabi, A.; Seyedzadeh, S.J.; Nawaz, M.; Szafert, S.; Mahmoodi, M.; Jabbari, E.; et al. Free and Hydrogel Encapsulated Exosome-Based Therapies in Regenerative Medicine. Life Sci. 2020, 249, 117447. [Google Scholar] [CrossRef] [PubMed]
- Isaacson, A.; Swioklo, S.; Connon, C.J. 3D Bioprinting of a Corneal Stroma Equivalent. Exp. Eye Res. 2018, 173, 188–193. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.; Jang, J.; Park, J.; Lee, K.-P.; Lee, S.; Lee, D.-M.; Kim, K.H.; Kim, H.K.; Cho, D.-W. Shear-Induced Alignment of Collagen Fibrils Using 3D Cell Printing for Corneal Stroma Tissue Engineering. Biofabrication 2019, 11, 035017. [Google Scholar] [CrossRef] [PubMed]
- Duarte Campos, D.F.; Rohde, M.; Ross, M.; Anvari, P.; Blaeser, A.; Vogt, M.; Panfil, C.; Yam, G.H.; Mehta, J.S.; Fischer, H.; et al. Corneal Bioprinting Utilizing Collagen-based Bioinks and Primary Human Keratocytes. J. Biomed. Mater. Res. Part A 2019, 107, 1945–1953. [Google Scholar] [CrossRef] [PubMed]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Therapeutic Method | Therapeutic Strategy | Advantages | Limitation | References | ||
|---|---|---|---|---|---|---|
| Exosomes | Origin | Target Cell | Mechanism of Action |
|
| [51,52,53,54,55,56,57,58,59] |
| Corneal stromal stem cells | Human corneal stromal cells (keratocytes) | TSG-6 protein in the exosomes prevents neutrophil infiltration, reducing the excessive secretion of TGF-β at the wounded area of the cornea | ||||
| Adipose-derived stem cells | Human corneal stromal cells (keratocytes) | miR-19a microRNA in the exosome post-transcriptionally silences HIPK2, halting the JNK fibrotic and TGF-β pathways | ||||
| Human corneal epithelial cells | Corneal epithelial cells | TSP1 protein in the exosomes attenuates paraptosis and helps in wound healing | ||||
| Tissue-derived microparticles | Lymph node ECM | Keratocytes | Increases the expression of mucin and lacrimal gland genes (maintains tear film homeostasis), reduces profibrotic gene expression, and reduces corneal haze | Tissue-derived particles can be processed in various physical forms, such as sheets, spheres, and gels, based on clinical needs. |
| [60,61,62] |
| Therapeutic Method | Therapeutic Strategy | Advantages | Limitations | References | ||
|---|---|---|---|---|---|---|
| Targeted gene silencing | Gene Targeted | Mechanism of Action on Targeted Gene | Method of Silencing |
| No proper delivery vehicle of siRNA inside the targeted cell because of its negative charge and water solubility, resulting in a poor penetration capacity. Instability of siRNA inside the cellular environment. Digestion of the siRNA by nucleases present in the cell cytoplasm. | [64,65,66,67,68,69,70,71] |
| Semaphorin 3A | Potentiates TGF-β to enhance its fibrotic activity | siRNA targeting SEMA-3A | ||||
| USP10 | Increases apoptosis by stabilizing p53 (initial stage of wound-healing process) and prevents ubiquitination of integrins by binding to its modulator (G3BP) | siRNA targeting USP10 | ||||
| KCa3.1 ion channel | Keratocyte hyperpolarization, resulting in their escape from the G1 phase of cell cycle. This leads to excessive cell proliferation. | TRAM 34, ion channel blocker | ||||
| Targeted gene overexpression | Gene Targeted | Mechanism of Action on Targeted Gene | Method of Overexpression |
| Extra stress on the cell, as cellular resources are wasted in translating and exporting the specific protein. Instability of plasmid DNA expression vectors containing the gene of interest or misincorporation of the gene of interest in the case of homologous recombination methods. | [78,79,80,81,82,83,84,85] |
| KLF4 | Suppresses EMT | Lentiviral vector | ||||
| Id3 | Sequestering bHLH transcription factors and preventing the downregulation of epithelial cell markers to hinder EMT. | pcDNA3-mCherry LIC mammalian expression vector construct | ||||
| SMAD7 | Prevents nuclear localization of SMAD2/3 and attenuates the TGF-β pathway by preventing the phosphorylation of SMAD3. | Recombinant adeno-associated viral vector | ||||
| Micro-RNA | Advantages | Limitation | Reference | |||
|---|---|---|---|---|---|---|
| Type | Level of Mirna During Healing Process | Therapeutic Regulatory Level Required | Mechanism of Action |
|
| |
| miR-204 | Downregulated | Upregulation | Targets SMAD4 | [87,88,89,90,91] | ||
| miR-145 | Upregulated | Downregulation | [92,93,94,95] | |||
| miR-133b | Downregulated | Upregulation | Targets KLF4 | [96,97,98] | ||
| Therapeutic Biomolecule | Therapeutic Strategy | Advantage | Limitation | Reference |
|---|---|---|---|---|
| Histone deacetylase inhibitors (e.g.,: SAHA) |
| Reversible epigenetic modification to create an antifibrotic environment. | HDAC inhibitors can have major side effects, such as reducing the number of viable T cells in the body and thrombocytopenia. Minor side effects include fatigue and nausea. | [100,101,102,103,104,105,106,107,108,109] |
| Growth factor (Insulin-like growth factor 1) | Converts keratocytes into collagenous keratocytes, which secrete native corneal ECM components. | Rules out myofibroblast formation and guides the cornea’s fibrotic healing process towards an antifibrotic method of wound healing. |
| [110,111,112] |
| Glucosamine |
| Cost-effective, increasing KLF4 stability without the need for costly gene therapy. |
| [116,117,118,119,120] |
| Chitosan | Anti-angiogenic and antifibrotic properties. | Well-established wound-healing properties and highly biocompatible. | Chitosan nanoparticle preparation can be cumbersome. | [122,123,124,125,126] |
| Lycium barbarum polysaccharide (LBP) | Reduces profibrotic gene expression. | It is safe with no adverse side effects. |
| [127,128,129] |
| JQ1 | Inhibitor of BRD4; therefore, it does not allow the binding of BRD4 to keap1, and Nrf2 can remain in its stable form by binding to keap1. Nrf2 then translocates into the nucleus to increase antioxidant gene expression and decrease ROS levels. |
|
| [136] |
| TPCA-1 | An IKK inhibitor that attenuates the NF-κβ pathway. | Inhibiting NF-κβ pathway decreases the cytokine storm during the healing process. |
| [137,138] |
| Acetylcholine | Promotes faster re-epithelization of the wounded corneal epithelial layer, activates protein kinase C, and decreases profibrotic gene expression. | Neurotransmitters can be structurally modified to obtain desired pharmacological activity. |
| [145,151] |
| Decorin | Activates CAM Kinase II to phosphorylate the serine 240 residue of SMAD2, forming the inhibitory SMAD2/3/4 complex, which cannot activate fibrotic genes. | A commonly used, clinically approved ocular drug to create an antifibrotic environment. | Poor retention time on the ocular surface. | [148,149,150] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ghosh, A.; Singh, V.K.; Singh, V.; Basu, S.; Pati, F. Recent Advancements in Molecular Therapeutics for Corneal Scar Treatment. Cells 2022, 11, 3310. https://doi.org/10.3390/cells11203310
Ghosh A, Singh VK, Singh V, Basu S, Pati F. Recent Advancements in Molecular Therapeutics for Corneal Scar Treatment. Cells. 2022; 11(20):3310. https://doi.org/10.3390/cells11203310
Chicago/Turabian StyleGhosh, Anwesha, Vijay K. Singh, Vivek Singh, Sayan Basu, and Falguni Pati. 2022. "Recent Advancements in Molecular Therapeutics for Corneal Scar Treatment" Cells 11, no. 20: 3310. https://doi.org/10.3390/cells11203310