

Key Genes Associated with Pyroptosis in Gout and Construction of a miRNA-mRNA Regulatory Network

Abstract
:1. Introduction
2. Materials and Methods
2.1. Microarray Data Acquisition
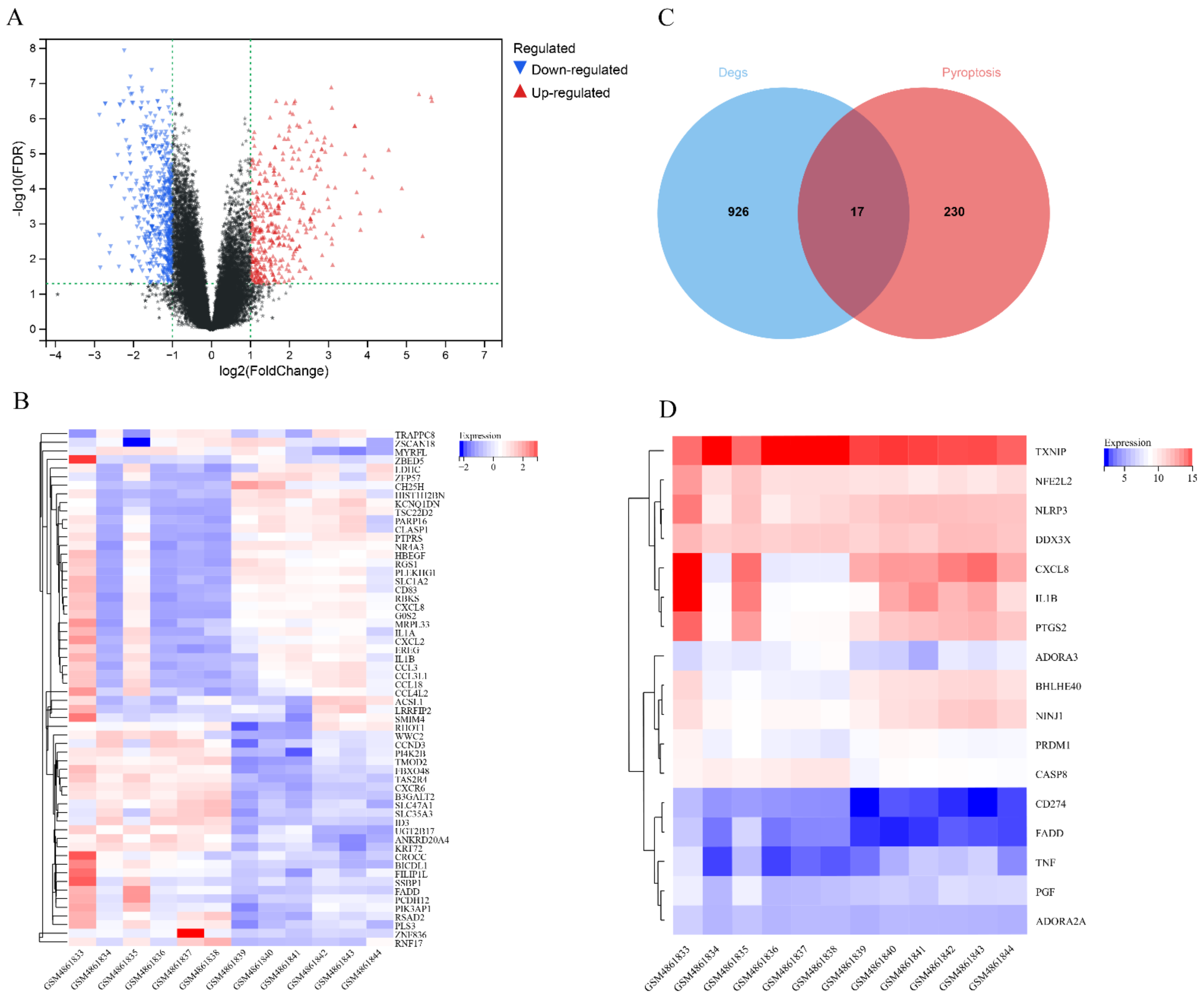
2.2. Differentially Expressed Genes Associated with Pyroptosis in Gout
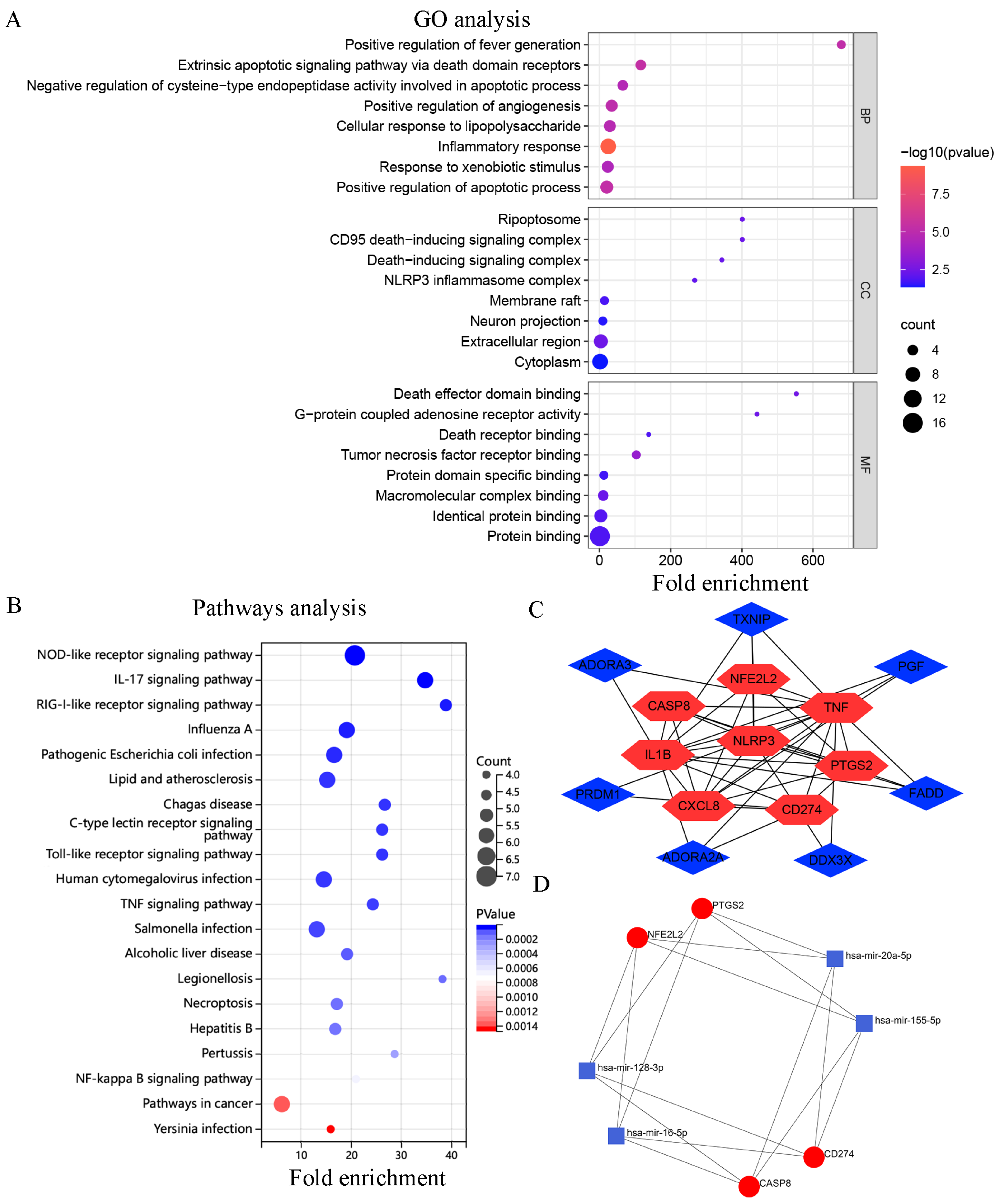
2.3. Functional Enrichment Analysis
2.4. Construction of Protein–Protein Interaction and miRNA-mRNA Networks
2.5. Patient Selection
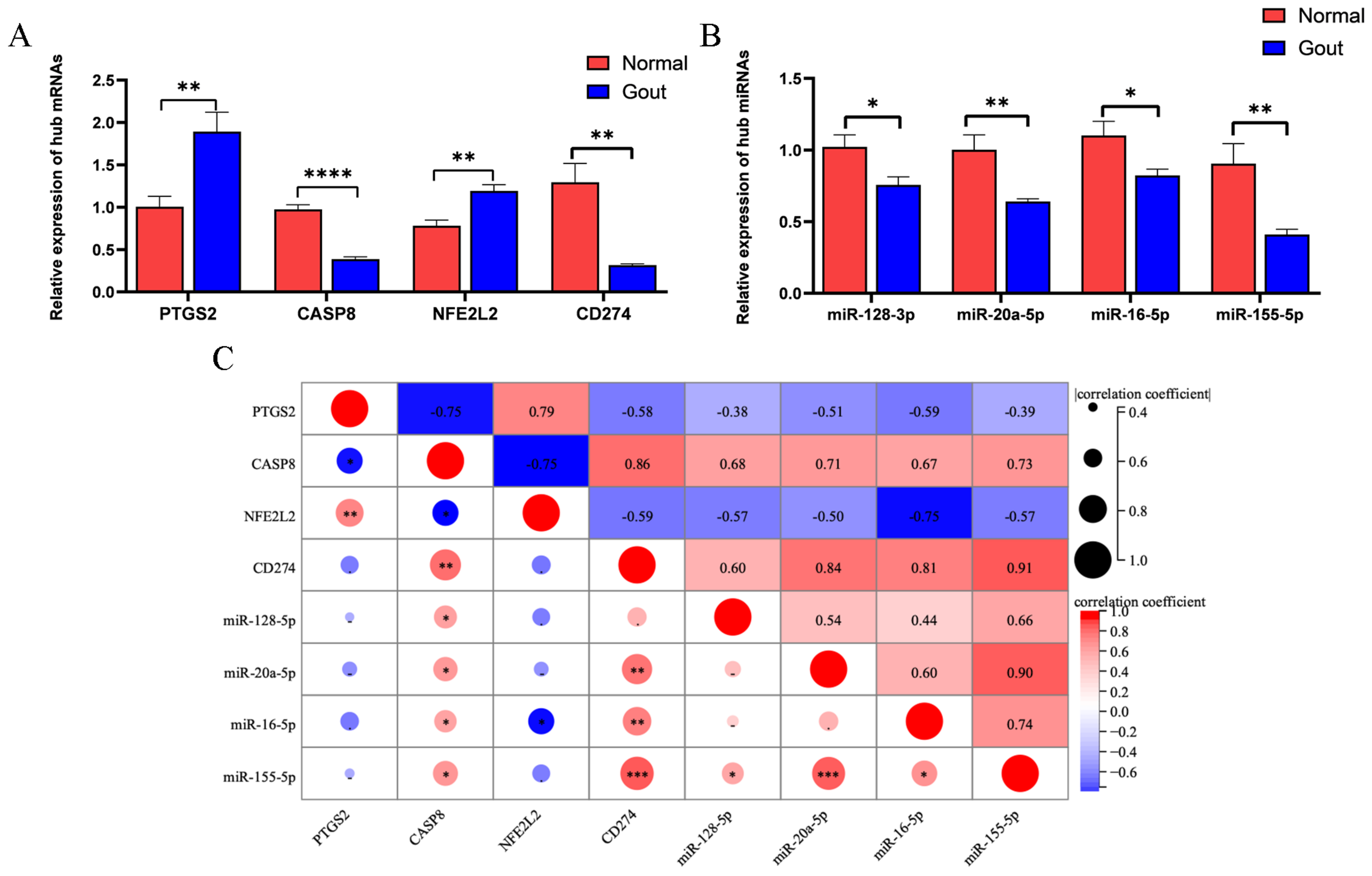
2.6. Quantitative Real-Time Polymerase Chain Reaction (qRT-PCR)
2.7. Statistical Analysis
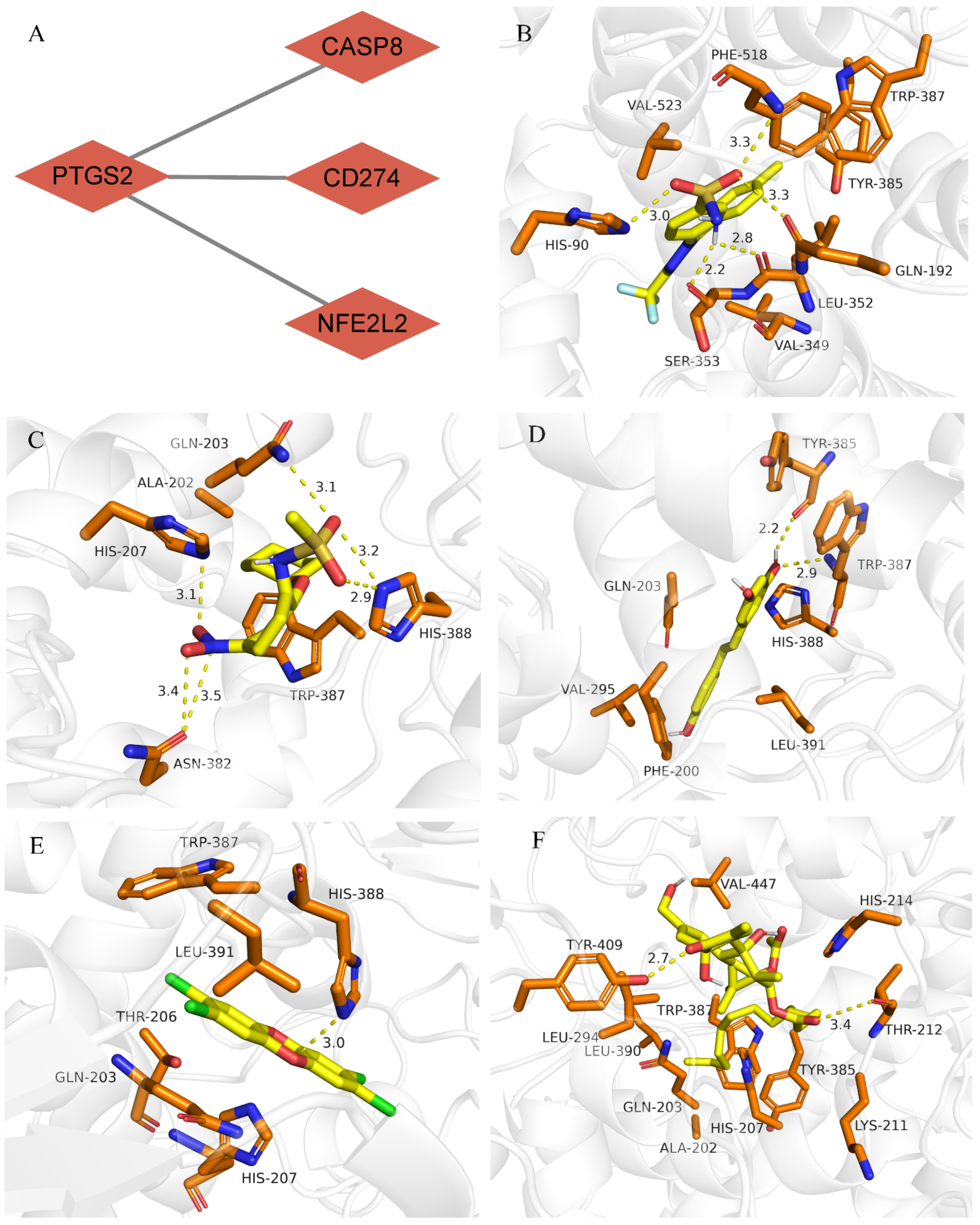
2.8. Drug-Gene Interaction and Molecular Docking Analysis
3. Results
3.1. Expression of Pyroptosis-Related Differentially Expressed Genes
3.2. Functional Annotation of the Target DEGs
3.3. PPI and miRNA-mRNA Networks
3.4. General Information on the Study Population
3.5. Validation of Pyroptosis-Related Genes Associated with Gout
3.6. Drug–Gene Interaction and Molecular Docking Analyses of PTGS2
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Singh, J.A.; Gaffo, A. Gout epidemiology and comorbidities. Semin. Arthritis Rheum. 2020, 50, S11–S16. [Google Scholar] [CrossRef] [PubMed]
- Man, S.M.; Karki, R.; Kanneganti, T.-D. Molecular mechanisms and functions of pyroptosis, inflammatory caspases and inflammasomes in infectious diseases. Immunol. Rev. 2017, 277, 61–75. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Narang, R.K.; Dalbeth, N. Pathophysiology of Gout. Semin. Nephrol. 2020, 40, 550–563. [Google Scholar] [CrossRef] [PubMed]
- Wu, M.; Tian, Y.; Wang, Q.; Guo, C. Gout: A disease involved with complicated immunoinflammatory responses: A narrative review. Clin. Rheumatol. 2020, 39, 2849–2859. [Google Scholar] [CrossRef]
- Dalbeth, N.; Gosling, A.L.; Gaffo, A.; Abhishek, A. Gout. Lancet 2021, 397, 1843–1855. [Google Scholar] [CrossRef]
- Xu, Y.-T.; Leng, Y.-R.; Liu, M.-M.; Dong, R.-F.; Bian, J.; Yuan, L.-L.; Zhang, J.-G.; Xia, Y.-Z.; Kong, L.-Y. MicroRNA and long noncoding RNA involvement in gout and prospects for treatment. Int. Immunopharmacol. 2020, 87, 106842. [Google Scholar] [CrossRef]
- Dhanasekaran, D.N.; Reddy, E.P. JNK-signaling: A multiplexing hub in programmed cell death. Genes Cancer 2017, 8, 682–694. [Google Scholar] [CrossRef] [Green Version]
- Gong, T.; Yang, Y.; Jin, T.; Jiang, W.; Zhou, R. Orchestration of NLRP3 Inflammasome Activation by Ion Fluxes. Trends Immunol. 2018, 39, 393–406. [Google Scholar] [CrossRef]
- Kayagaki, N.; Stowe, I.B.; Lee, B.L.; O’Rourke, K.; Anderson, K.; Warming, S.; Cuellar, T.; Haley, B.; Roose-Girma, M.; Phung, Q.T.; et al. Caspase-11 cleaves gasdermin D for non-canonical inflammasome signalling. Nature 2015, 526, 666–671. [Google Scholar] [CrossRef]
- Al Mamun, A.; Mimi, A.A.; Wu, Y.; Zaeem, M.; Aziz, A.; Suchi, S.A.; Alyafeai, E.; Munir, F.; Xiao, J. Pyroptosis in diabetic nephropathy. Clin. Chim. Acta 2021, 523, 131–143. [Google Scholar] [CrossRef]
- Qian, Z.; Zhao, Y.; Wan, C.; Deng, Y.; Zhuang, Y.; Xu, Y.; Zhu, Y.; Lu, S.; Bao, Z. Pyroptosis in the Initiation and Progression of Atherosclerosis. Front. Pharmacol. 2021, 12, 1312. [Google Scholar] [CrossRef]
- Zhang, Q.-B.; Zhu, D.; Dai, F.; Huang, Y.-Q.; Zheng, J.-X.; Tang, Y.-P.; Dong, Z.-R.; Liao, X.; Qing, Y.-F. MicroRNA-223 Suppresses IL-1β and TNF-α Production in Gouty Inflammation by Targeting the NLRP3 Inflammasome. Front. Pharmacol. 2021, 12, 1312. [Google Scholar] [CrossRef]
- Dong, W.; Gao, W.; Cui, J.; Xu, T.; Sun, Y. microRNA-148 is involved in NF-κB signaling pathway regulation after LPS stimulation by targeting IL-1β in miiuy croaker. Fish Shellfish Immunol. 2021, 118, 66–71. [Google Scholar] [CrossRef]
- Lu, Q.; Wu, R.; Zhao, M.; Garcia-Gomez, A.; Ballestar, E. miRNAs as Therapeutic Targets in Inflammatory Disease. Trends Pharmacol. Sci. 2019, 40, 853–865. [Google Scholar] [CrossRef]
- Gebert, L.F.R.; Macrae, I.J. Regulation of microRNA function in animals. Nat. Rev. Mol. Cell Biol. 2018, 20, 21–37. [Google Scholar] [CrossRef]
- Zhou, J.; Yu, L.; Gao, X.; Hu, J.; Wang, J.; Dai, Z.; Wang, J.-F.; Zhang, Z.; Lu, S.; Huang, X.; et al. Plasma MicroRNA Panel to Diagnose Hepatitis B Virus–Related Hepatocellular Carcinoma. J. Clin. Oncol. 2011, 29, 4781–4788. [Google Scholar] [CrossRef] [Green Version]
- Giannopoulou, L.; Zavridou, M.; Kasimir-Bauer, S.; Lianidou, E.S. Liquid biopsy in ovarian cancer: The potential of circulating miRNAs and exosomes. Transl. Res. 2019, 205, 77–91. [Google Scholar] [CrossRef]
- Dieckmann, K.-P.; Radtke, A.; Geczi, L.; Matthies, C.; Anheuser, P.; Eckardt, U.; Sommer, J.; Zengerling, F.; Trenti, E.; Pichler, R.; et al. Serum Levels of MicroRNA-371a-3p (M371 Test) as a New Biomarker of Testicular Germ Cell Tumors: Results of a Prospective Multicentric Study. J. Clin. Oncol. 2019, 37, 1412–1423. [Google Scholar] [CrossRef]
- Qing, Y.-F.; Zheng, J.-X.; Tang, Y.-P.; Dai, F.; Dong, Z.-R.; Zhang, Q.-B. LncRNAs Landscape in the patients of primary gout by microarray analysis. PLoS ONE 2021, 16, e0232918. [Google Scholar] [CrossRef]
- Liu, T.; Li, X.; Cui, Y.; Meng, P.; Zeng, G.; Wang, Q.; Wang, Y. Bioinformatics Analysis Identifies Potential Ferroptosis Key Genes in the Pathogenesis of Intracerebral Hemorrhage. Front. Neurosci. 2021, 15, 529. [Google Scholar] [CrossRef]
- Fishilevich, S.; Zimmerman, S.; Kohn, A.; Stein, T.I.; Olender, T.; Kolker, E.; Safran, M.; Lancet, R. Genic insights from integrated human proteomics in GeneCards. Database 2016, 2016, baw030. [Google Scholar] [CrossRef] [Green Version]
- Su, T.; Gu, C.; Draga, D.; Zhou, C.; Lhamo, T.; Zheng, Z.; Qiu, Q. Integrative analysis of miRNA–mRNA network in high altitude retinopathy by bioinformatics analysis. Biosci. Rep. 2021, 41, bsr20200776. [Google Scholar] [CrossRef]
- Sun, S.; Shen, Y.; Wang, J.; Li, J.; Cao, J.; Zhang, J. Identification and Validation of Autophagy-Related Genes in Chronic Obstructive Pulmonary Disease. Int. J. Chronic Obstr. Pulm. Dis. 2021, 16, 67–78. [Google Scholar] [CrossRef]
- Zhou, G.; Soufan, O.; Ewald, J.; Hancock, R.E.W.; Basu, N.; Xia, J. NetworkAnalyst 3.0: A visual analytics platform for comprehensive gene expression profiling and meta-analysis. Nucleic Acids Res. 2019, 47, W234–W241. [Google Scholar] [CrossRef] [Green Version]
- Neogi, T.; Jansen, T.L.T.A.; Dalbeth, N.; Fransen, J.; Schumacher, H.R.; Berendsen, D.; Brown, M.; Choi, H.; Edwards, N.L.; Janssens, H.J.E.M.; et al. 2015 Gout classification criteria: An American College of Rheumatology/European League Against Rheumatism collaborative initiative. Ann. Rheum. Dis. 2015, 74, 1789–1798. [Google Scholar] [CrossRef] [Green Version]
- Davis, A.P.; Wiegers, T.C.; Johnson, R.J.; Sciaky, D.; Wiegers, J.; Mattingly, C.J. Comparative Toxicogenomics Database (CTD): Update 2021. Nucleic Acids Res. 2021, 49, D1138–D1143. [Google Scholar] [CrossRef]
- Geoffrey, B.; Sanker, A.; Madaj, R.; Valdés Tresanco, M.S.; Upadhyay, M.; Gracia, J. A program to automate the discovery of drugs for West Nile and Dengue virus-programmatic screening of over a billion compounds on PubChem, generation of drug leads and automated in silico modelling. J. Biomol. Struct. Dyn. 2022, 40, 4293–4300. [Google Scholar] [CrossRef]
- Karuppasamy, M.P.; Venkateswaran, S.; Subbiah, P. PDB-2-PBv3.0: An updated protein block database. J. Bioinform. Comput. Biol. 2020, 18, 2050009. [Google Scholar] [CrossRef]
- Nguyen, N.T.; Nguyen, T.H.; Pham, T.N.H.; Huy, N.T.; Van Bay, M.; Pham, M.Q.; Nam, P.C.; Vu, V.V.; Ngo, S.T. Autodock Vina Adopts More Accurate Binding Poses but Autodock4 Forms Better Binding Affinity. J. Chem. Inf. Model. 2019, 60, 204–211. [Google Scholar] [CrossRef]
- Lam, W.W.T.; Siu, S.W.I. PyMOL mControl: Manipulating molecular visualization with mobile devices. Biochem. Mol. Biol. Educ. 2016, 45, 76–83. [Google Scholar] [CrossRef]
- Wang, Y.; Zhao, Y.-R.; Zhang, A.-Y.; Ma, J.; Wang, Z.-Z.; Zhang, X. Targeting of miR-20a against CFLAR to potentiate TRAIL-induced apoptotic sensitivity in HepG2 cells. Eur. Rev. Med Pharmacol. Sci. 2017, 21, 2087–2097. [Google Scholar] [PubMed]
- Lin, Y.; Wu, Z. MicroRNA-128 inhibits proliferation and invasion of glioma cells by targeting COX-2. Gene 2018, 658, 63–69. [Google Scholar] [CrossRef] [PubMed]
- Huang, J.; Weng, Q.; Shi, Y.; Mao, W.; Zhao, Z.; Wu, R.; Ren, J.; Fang, S.; Lu, C.; Du, Y.; et al. MicroRNA-155-5p suppresses PD-L1 expression in lung adenocarcinoma. FEBS Open Bio 2020, 10, 1065–1071. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Caggiano, R.; Cattaneo, F.; Moltedo, O.; Esposito, G.; Perrino, C.; Trimarco, B.; Ammendola, R.; Faraonio, R. miR-128 Is Implicated in Stress Responses by Targeting MAFG in Skeletal Muscle Cells. Oxidative Med. Cell. Longev. 2017, 2017, 1–13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wicherska-Pawłowska, K.; Wróbel, T.; Rybka, J. Toll-Like Receptors (TLRs), NOD-Like Receptors (NLRs), and RIG-I-Like Receptors (RLRs) in Innate Immunity. TLRs, NLRs, and RLRs Ligands as Immunotherapeutic Agents for Hematopoietic Diseases. Int. J. Mol. Sci. 2021, 22, 13397. [Google Scholar] [CrossRef] [PubMed]
- Kinra, M.; Nampoothiri, M.; Arora, D.; Mudgal, J. Reviewing the importance of TLR-NLRP3-pyroptosis pathway and mechanism of experimental NLRP3 inflammasome inhibitors. Scand. J. Immunol. 2021, 95, e13124. [Google Scholar] [CrossRef] [PubMed]
- Wei, Q.; Lv, F.; Zhang, H.; Wang, X.; Geng, Q.; Zhang, X.; Li, T.; Wang, S.; Wang, Y.; Cui, Y. MicroRNA-101-3p inhibits fibroblast-like synoviocyte proliferation and inflammation in rheumatoid arthritis by targeting PTGS2. Biosci. Rep. 2020, 40, BSR20191136. [Google Scholar] [CrossRef] [Green Version]
- Fritsch, M.; Günther, S.D.; Schwarzer, R.; Albert, M.-C.; Schorn, F.; Werthenbach, J.P.; Schiffmann, L.M.; Stair, N.; Stocks, H.; Seeger, J.M.; et al. Caspase-8 is the molecular switch for apoptosis, necroptosis and pyroptosis. Nature 2019, 575, 683–687. [Google Scholar] [CrossRef]
- Tummers, B.; Mari, L.; Guy, C.S.; Heckmann, B.L.; Rodriguez, D.A.; Rühl, S.; Moretti, J.; Crawford, J.C.; Fitzgerald, P.; Kanneganti, T.-D.; et al. Caspase-8-Dependent Inflammatory Responses Are Controlled by Its Adaptor, FADD, and Necroptosis. Immunity 2020, 52, 994–1006.e8. [Google Scholar] [CrossRef]
- Ma, Q. Role of Nrf2 in Oxidative Stress and Toxicity. Annu. Rev. Pharmacol. Toxicol. 2013, 53, 401–426. [Google Scholar] [CrossRef]
- Ahmed, S.M.U.; Luo, L.; Namani, A.; Wang, X.J.; Tang, X. Nrf2 signaling pathway: Pivotal roles in inflammation. Biochim. Biophys. Acta Mol. Basis Dis. 2017, 1863, 585–597. [Google Scholar] [CrossRef]
- Liu, W.; Zang, X. Structures of Immune Checkpoints: An Overview on the CD28-B7 Family. Adv. Exp. Med. Biol. 2019, 1172, 63–78. [Google Scholar] [CrossRef]
- Garo, L.P.; Ajay, A.K.; Fujiwara, M.; Beynon, V.; Kuhn, C.; Gabriely, G.; Sadhukan, S.; Raheja, R.; Rubino, S.; Weiner, H.L.; et al. Smad7 Controls Immunoregulatory PDL2/1-PD1 Signaling in Intestinal Inflammation and Autoimmunity. Cell Rep. 2019, 28, 3353–3366.e5. [Google Scholar] [CrossRef] [Green Version]
- Song, M.-Y.; Hong, C.-P.; Park, S.J.; Kim, J.-H.; Yang, B.-G.; Park, Y.; Kim, S.W.; Kim, K.S.; Lee, J.Y.; Lee, S.-W.; et al. Protective effects of Fc-fused PD-L1 on two different animal models of colitis. Gut 2014, 64, 260–271. [Google Scholar] [CrossRef]
- Plotnikova, O.M.; Skoblov, M.Y. Efficiency of the miRNA-mRNA Interaction Prediction Programs. Mol. Biol. 2018, 52, 543–554. [Google Scholar] [CrossRef]
- Chen, S.; Li, B. MiR-128-3p Post-Transcriptionally Inhibits WISP1 to Suppress Apoptosis and Inflammation in Human Articular Chondrocytes via the PI3K/AKT/NF-κB Signaling Pathway. Cell Transplant. 2020, 29, 0963689720939131. [Google Scholar] [CrossRef]
- Fu, X.; Qie, J.; Fu, Q.; Chen, J.; Jin, Y.; Ding, Z. miR-20a-5p/TGFBR2 Axis Affects Pro-inflammatory Macrophages and Aggravates Liver Fibrosis. Front. Oncol. 2020, 10, 107. [Google Scholar] [CrossRef]
- Zhang, J.; Zhu, L.; Shi, H.; Zheng, H. Protective effects of miR-155-5p silencing on IFN-γ-induced apoptosis and inflammation in salivary gland epithelial cells. Exp. Ther. Med. 2021, 22, 1–11. [Google Scholar] [CrossRef]
- Wang, J.; Zhao, Q. LncRNA LINC-PINT increases SOCS1 expression by sponging miR-155-5p to inhibit the activation of ERK signaling pathway in rheumatoid arthritis synovial fibroblasts induced by TNF-α. Int. Immunopharmacol. 2020, 84, 106497. [Google Scholar] [CrossRef]
- Lei, J.; Fu, Y.; Zhuang, Y.; Zhang, K.; Lu, D. LncRNA SNHG1 alleviates IL-1β-induced osteoarthritis by inhibiting miR-16-5p-mediated p38 MAPK and NF-κB signaling pathways. Biosci. Rep. 2019, 39, BSR20191523. [Google Scholar] [CrossRef]
- Pillinger, M.H.; Mandell, B.F. Therapeutic approaches in the treatment of gout. Semin. Arthritis Rheum. 2020, 50, S24–S30. [Google Scholar] [CrossRef]
- Robinson, P.C.; Terkeltaub, R.; Pillinger, M.H.; Shah, B.; Karalis, V.; Karatza, E.; Liew, D.; Imazio, M.; Cornel, J.H.; Thompson, P.L.; et al. Consensus Statement Regarding the Efficacy and Safety of Long-Term Low-Dose Colchicine in Gout and Cardiovascular Disease. Am. J. Med. 2021, 135, 32–38. [Google Scholar] [CrossRef]
- Li, H.; Ou, G.; He, Y.; Ren, L.; Yang, X.; Zeng, M. Resveratrol attenuates the MSU crystal-induced inflammatory response through the inhibition of TAK1 activity. Int. Immunopharmacol. 2018, 67, 62–68. [Google Scholar] [CrossRef]
- Yang, Q.-B.; He, Y.-L.; Zhong, X.-W.; Xie, W.-G.; Zhou, J.-G. Resveratrol ameliorates gouty inflammation via upregulation of sirtuin 1 to promote autophagy in gout patients. Inflammopharmacology 2019, 27, 47–56. [Google Scholar] [CrossRef]
- Schumacher, H.R.; Berger, M.F.; Li-Yu, J.; Perez-Ruiz, F.; Burgos-Vargas, R.; Li, C. Efficacy and Tolerability of Celecoxib in the Treatment of Acute Gouty Arthritis: A Randomized Controlled Trial. J. Rheumatol. 2012, 39, 1859–1866. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Primer Sequence (5′–3′) |
|---|---|
| CASP8 (FORWARD) | GCCTTGATGTTATTCCAGAGAC |
| CASP8 (REVERSE) | TCTGAAGTTCCCTTTCCATCTC |
| PTGS2 (FORWARD) | ATCCTCCCACAGTCAAAGATAC |
| PTGS2 (REVERSE) | CGCATACTCTGTTGTGTTCC |
| CD274 (FORWARD) | TAGGAAGACGGGTTGAGAATC |
| CD274 (REVERSE) | CACACTCACATGACAAGAAGAC |
| NFE2L2 (FORWARD) | TCTCTTCTGTGCTGTCAAGG |
| NFE2L2 (REVERSE) | AGCTCATACTCTTTCCGTCG |
| hsa-miR-128-3p (FORWARD) | CGTCACAGTGAACCGGTCTCT |
| hsa-miR-16-5p FORWARD) | GCTAGCAGCACGTAAATATTGGCG |
| hsa-miR-20a-5p (FORWARD) | GGGCTAAAGTGCTTATAGTGCAGGT |
| hsa-miR-155-5p (FORWARD) | CGCTTAATGCTAATCGTGATAGGGGT |
| U6 (FORWARD) | GCTTCGGCAGCACATATACTAAAAT |
| U6 (REVERSE) | CGCTTCACGAATTTGCGTGTCAT |
| GAPDH (FORWARD) | TGAGGCCGGTGCTGAGTATGT |
| GAPDH (REVERSE) | CAGTCTTCTGGGTGGCAGTGAT |
| Gene Symbol | logFC | Adj. p-Value | p-Value | Changes |
|---|---|---|---|---|
| NLRP3 | 1.45234415 | 0.0015 | 7.53 × 10−5 | UP |
| IL1B | 3.09126135 | 0.00366 | 0.00026 | UP |
| DDX3X | 1.80012202 | 0.0155 | 0.00184 | UP |
| PRDM1 | 1.84476925 | 0.000238 | 5.77 × 10−6 | UP |
| NFE2L2 | 1.73936101 | 0.00017 | 3.46 × 10−6 | UP |
| PGF | 1.03779492 | 5.53 × 10−5 | 6.64 × 10−7 | UP |
| PTGS2 | 2.70238972 | 2.24 × 10−5 | 1.83 × 10−7 | UP |
| ADORA2A | 1.42834031 | 0.000129 | 2.36 × 10−6 | UP |
| TNF | 2.10325641 | 0.00709 | 0.00064 | UP |
| BHLHE40 | 2.62218684 | 8.26 × 10−6 | 3.92 × 10−8 | UP |
| NINJ1 | 1.72377655 | 0.00119 | 5.56 × 10−5 | UP |
| CXCL8 | 5.62812965 | 2.40 × 10−7 | 1.18 × 10−10 | UP |
| CASP8 | −1.07127595 | 0.000251 | 6.19 × 10−6 | DOWN |
| TXNIP | −1.04520555 | 0.00707 | 0.000637 | DOWN |
| CD274 | −1.59880911 | 0.00231 | 0.000138 | DOWN |
| ADORA3 | −1.36258112 | 0.0155 | 0.00185 | DOWN |
| FADD | −2.26130412 | 1.20 × 10−6 | 2.24 × 10−9 | DOWN |
| Gene | Degree Score | MCC Score | MNC Score |
|---|---|---|---|
| TNF | 14 | 330 | 14 |
| IL1B | 12 | 326 | 12 |
| NLRP3 | 9 | 290 | 9 |
| CXCL8 | 8 | 270 | 8 |
| PTGS2 | 7 | 264 | 7 |
| NFE2L2 | 6 | 144 | 6 |
| CASP8 | 6 | 144 | 6 |
| CD274 | 6 | 32 | 6 |
| TXNIP | 4 | 24 | 4 |
| FADD | 4 | 24 | 4 |
| PGF | 3 | 6 | 3 |
| ADORA2A | 3 | 6 | 3 |
| DDX3X | 2 | 2 | 2 |
| PRDM1 | 2 | 2 | 2 |
| ADORA3 | 2 | 2 | 2 |
| Mean ± Standard Deviation | Gout (n = 5) | Control (n = 5) | p Value |
|---|---|---|---|
| Age (years) | 46.6 ± 6.43 | 48.2 ± 1.10 | 0.5981 |
| Gender (male/female) | 5/0 | 5/0 | |
| Uric acid (mmol/L) | 552.64 ± 76.82 | 341.8 ± 32.87 | 0.0005 |
| Corresponding Receptor Protein PTGS2 | Binding Energy (kcal mol−1) |
|---|---|
| Celecoxib | −11 |
| N-(2-cyclohexyloxy-4-nitrophenyl)methanesulfonamide | −7.8 |
| Resveratrol | −8.1 |
| Tetrachlorodibenzodioxin | −8.4 |
| Tetradecanoylphorbol Acetate | −7 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bai, B.; Liu, Y.; Abudukerimu, A.; Tian, T.; Liang, M.; Li, R.; Sun, Y. Key Genes Associated with Pyroptosis in Gout and Construction of a miRNA-mRNA Regulatory Network. Cells 2022, 11, 3269. https://doi.org/10.3390/cells11203269
Bai B, Liu Y, Abudukerimu A, Tian T, Liang M, Li R, Sun Y. Key Genes Associated with Pyroptosis in Gout and Construction of a miRNA-mRNA Regulatory Network. Cells. 2022; 11(20):3269. https://doi.org/10.3390/cells11203269
Chicago/Turabian StyleBai, Bing, Yezhou Liu, Azierguli Abudukerimu, Tingting Tian, Meiting Liang, Rui Li, and Yuping Sun. 2022. "Key Genes Associated with Pyroptosis in Gout and Construction of a miRNA-mRNA Regulatory Network" Cells 11, no. 20: 3269. https://doi.org/10.3390/cells11203269