
Dermal Papilla Cell-Derived Extracellular Vesicles Increase Hair Inductive Gene Expression in Adipose Stem Cells via β-Catenin Activation

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. FBS EV Depletion and EV Isolation
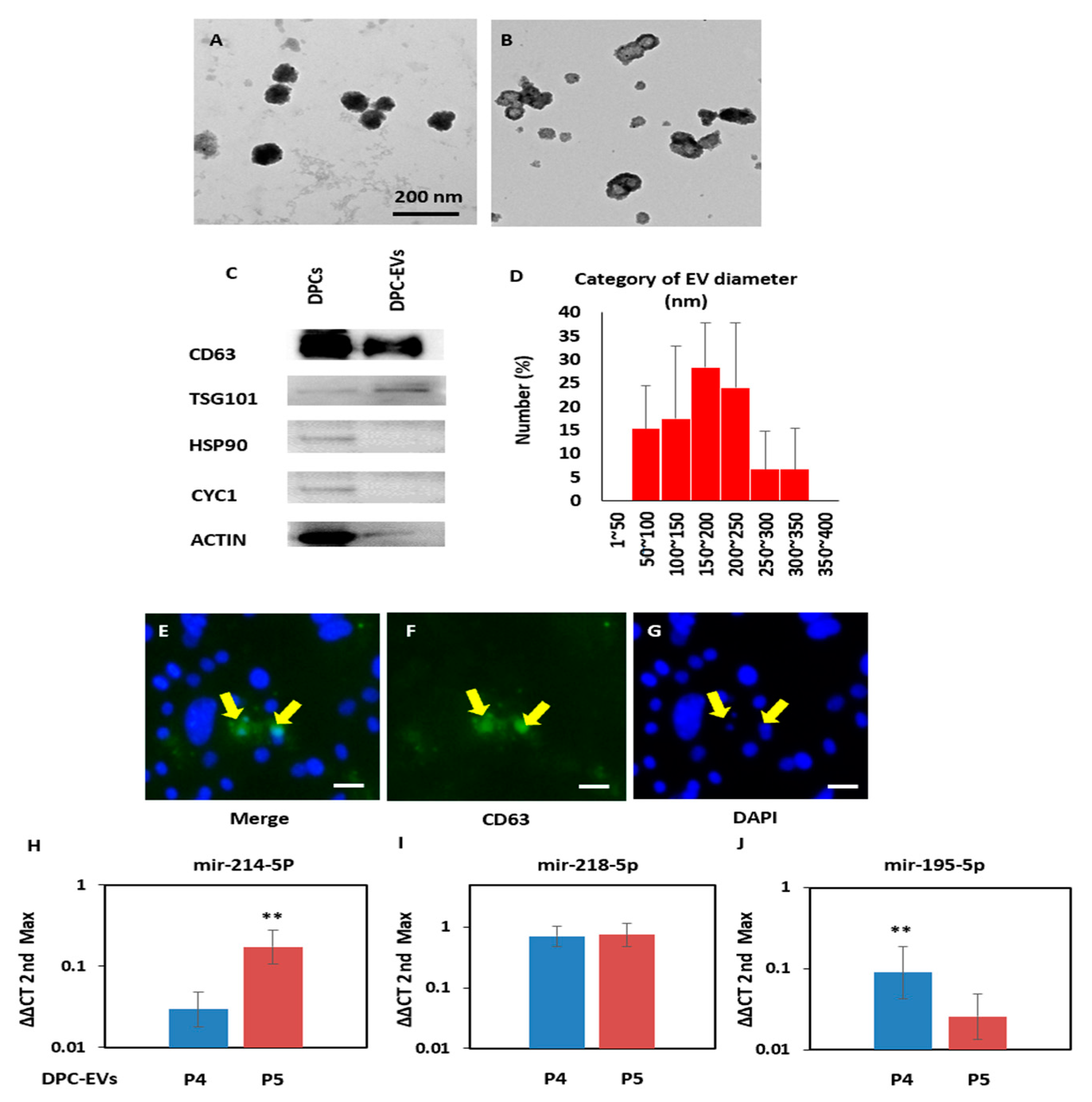
2.2. Transmission Electron Microscope (TEM)
2.3. Cell Culture
2.3.1. Culture Media
2.3.2. Isolation of DPCs
2.3.3. Isolation of ASCs
2.4. Oil O Red and Alkaline Phosphatase (ALP) Staining
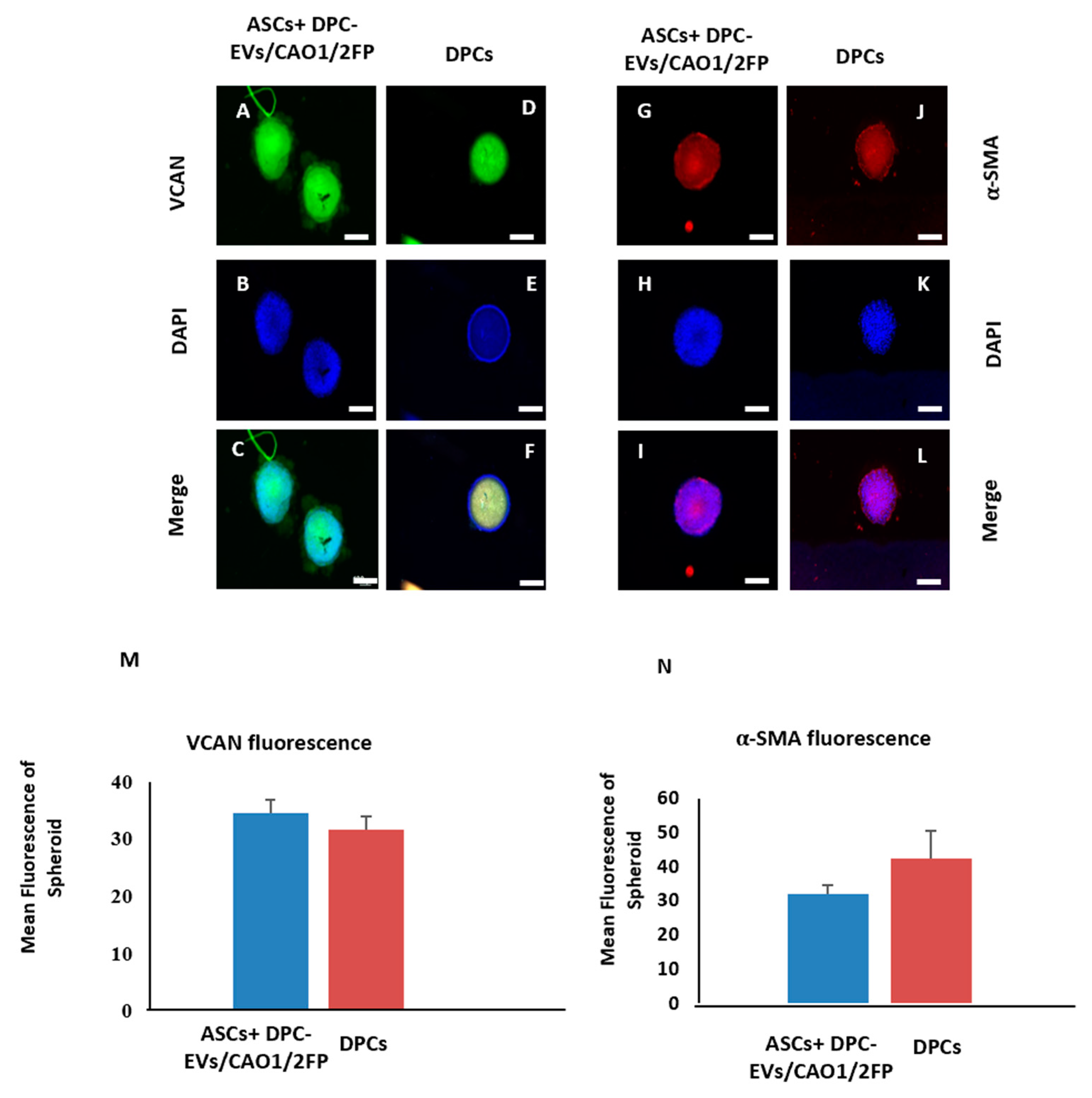
2.5. Immunocytochemistry
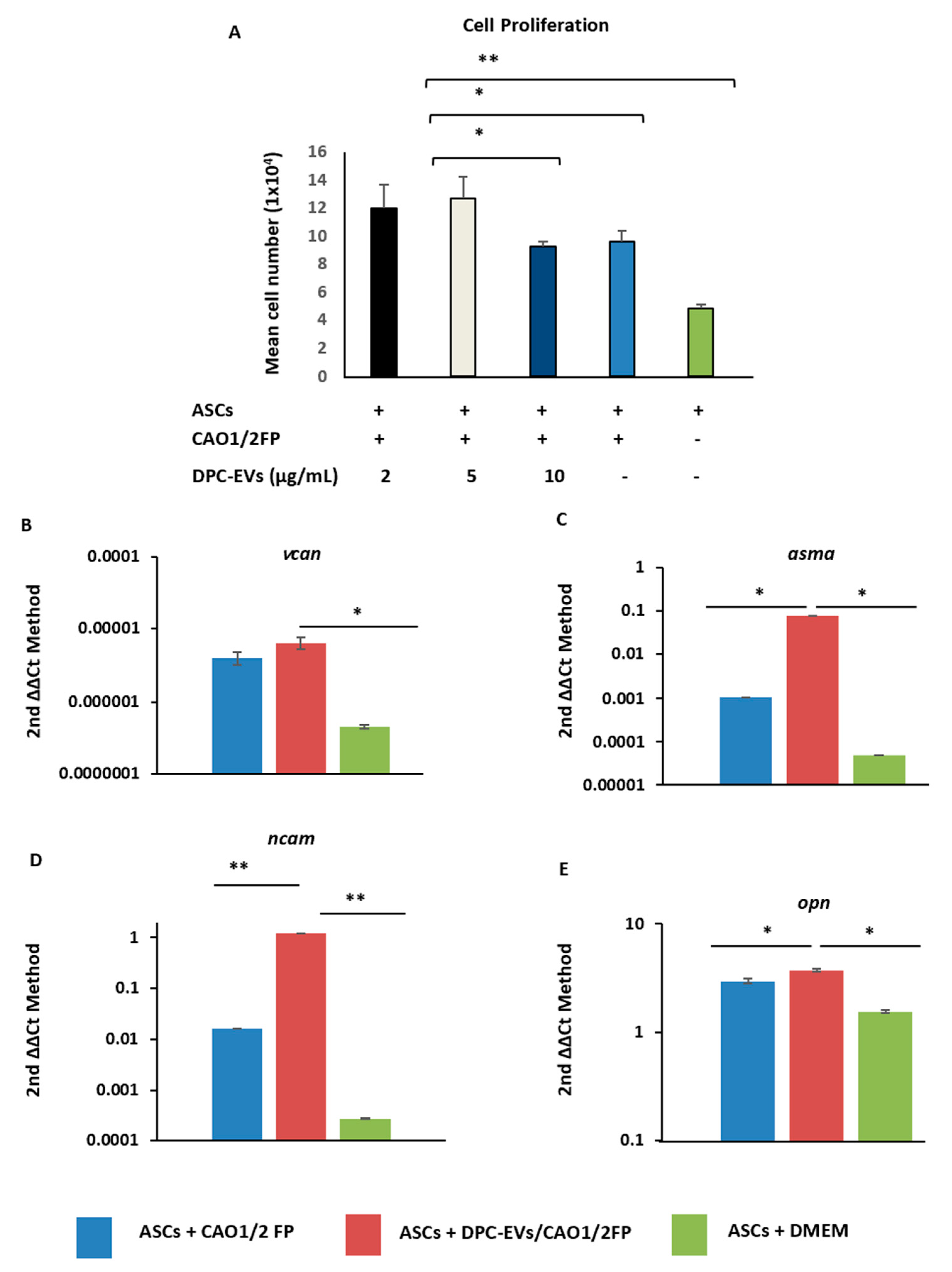
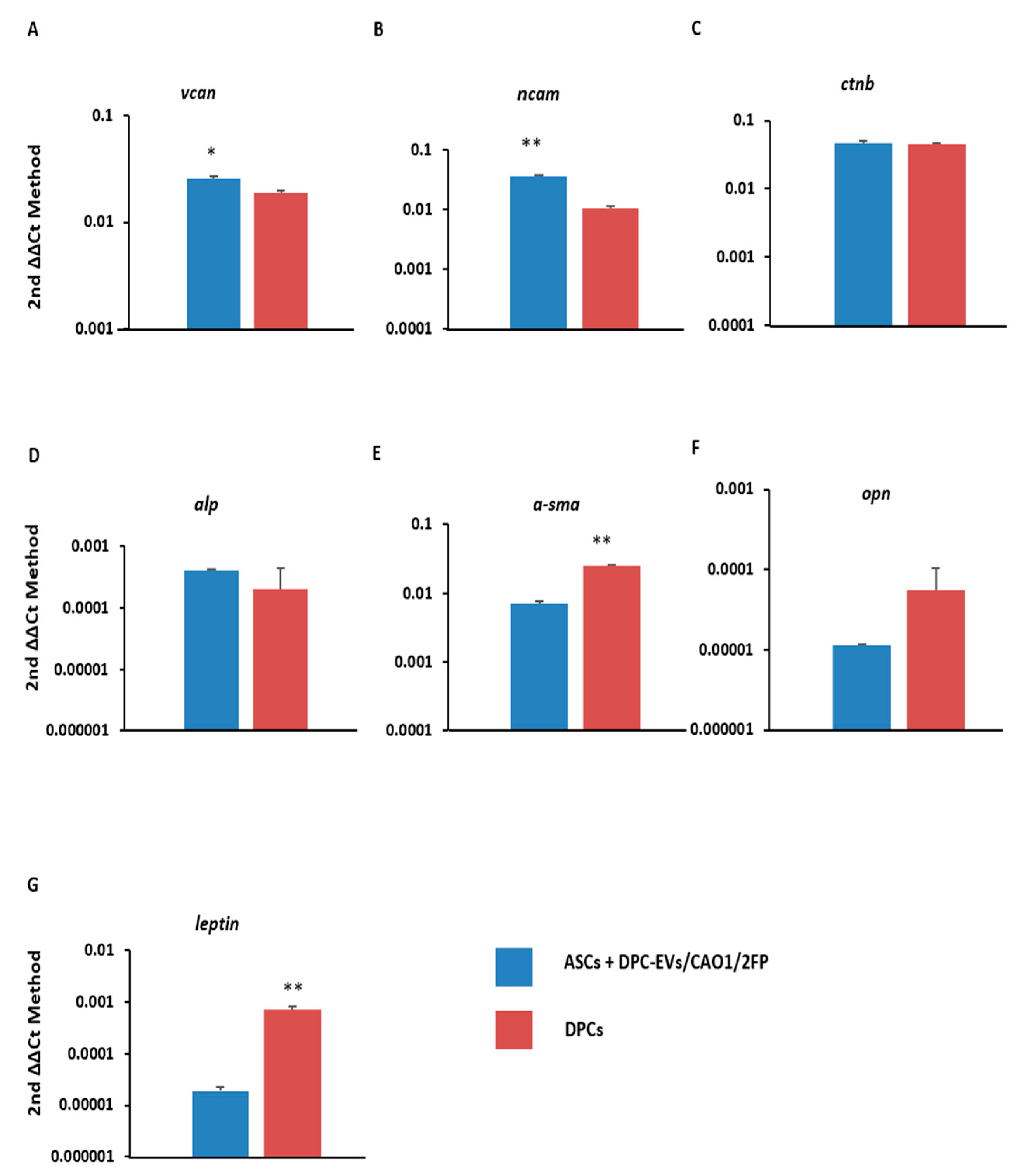
2.6. mRNA Extraction and RT-PCR
2.7. miRNA Extraction and RT-PCR
2.8. Protein Concentration and Western Blot
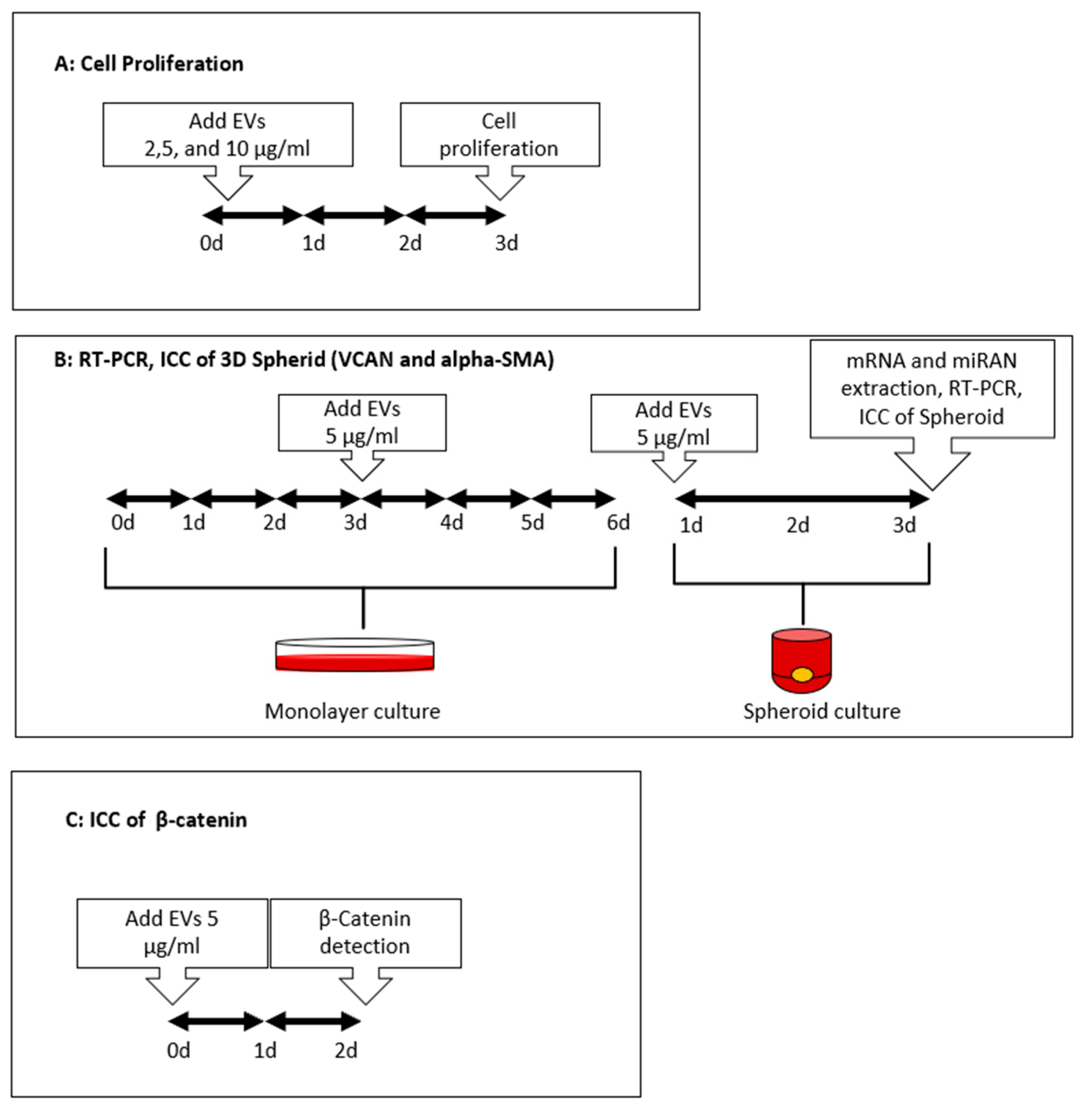
2.9. Cell Proliferation Assay
2.10. Statistical Analysis
3. Results
3.1. Identification of DPC-EVs
3.2. Induction of Cell Proliferation and Inculcation of Hair Induction-Related Gene Expression in ASCs with DPC-EVs
3.3. Transdifferentiation of ASCs into DPC-like Cells through Incubation with DPC-EVs
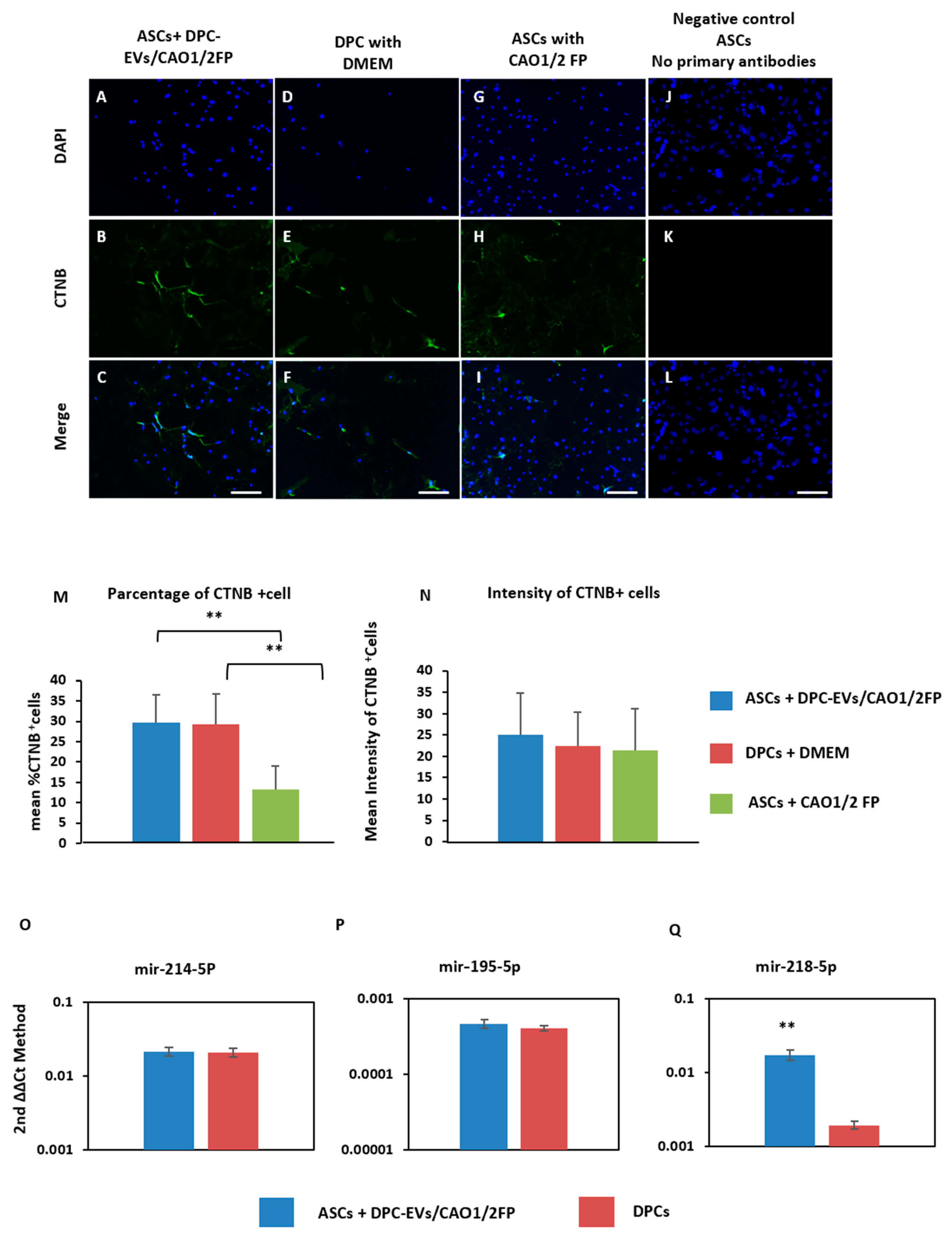
3.4. Induction of the CTNB Pathway in ASCs through Incubation with DPC-EVs and miRNAs
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Chueh, S.C.; Lin, S.J.; Chen, C.C.; Lei, M.; Wang, L.M.; Widelitz, R.; Hughes, M.W.; Jiang, T.X.; Chuong, C.M. Therapeutic strategy for hair regeneration: Hair cycle activation, niche environment modulation, wound-induced follicle neogenesis, and stem cell engineering. Expert Opin. Biol. Therapy 2013, 13, 377–391. [Google Scholar] [CrossRef] [Green Version]
- Banka, N.; Bunagan, M.J.; Shapiro, J. Pattern hair loss in men: Diagnosis and medical treatment. Dermatol. Clin. 2013, 31, 129–140. [Google Scholar] [CrossRef]
- Jahoda, C.A.B.; Oliver, R.F. Vibrissa dermal papilla cell aggregative behaviour in vivo and in vitro. J. Dembryovelogy Exp. Morphol. 1984, 79, 211–224. [Google Scholar] [CrossRef]
- Jahoda, C.; Horne, K.A.; Oliver, R.F. Induction of hair growth by implantation of cultured dermal papilla cells. Nature 1984, 311, 560–562. [Google Scholar] [CrossRef] [PubMed]
- Robinson, M.; Reynolds, A.J.; Jahoda, C.A. Hair Cycle Stage of the Mouse Vibrissa Follicle Determines Subsequent Fiber Growth and Follicle Behavior In Vitro. J. Investig. Dermatol. 1997, 108, 495–500. [Google Scholar] [CrossRef] [Green Version]
- Jung, Y.J.; Kim, H.K.; Cho, Y.; Choi, J.S.; Woo, C.H.; Lee, K.S.; Sul, J.H.; Lee, C.M.; Han, J.; Park, J.H.; et al. Cell reprogramming using extracellular vesicles from differentiating stem cells into white/beige adipocytes. Sci. Adv. 2020, 6, eaay6721. [Google Scholar] [CrossRef] [Green Version]
- Rajendran, R.L.; Gangadaran, P.; Bak, S.S.; Oh, J.M.; Kalimuthu, S.; Lee, H.W.; Baek, S.H.; Zhu, L.; Sung, Y.K.; Jeong, S.Y.; et al. Extracellular vesicles derived from MSCs activates dermal papilla cell in vitro and promotes hair follicle conversion from telogen to anagen in mice. Sci. Rep. 2017, 7, 15560. [Google Scholar] [CrossRef] [Green Version]
- Narayanan, R.; Huang, C.-C.; Ravindran, S. Hijacking the Cellular Mail: Exosome Mediated Differentiation of Mesenchymal Stem Cells. Stem Cells Int. 2016, 2016, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Mizuno, H. Adipose-derived Stem Cells for Tissue Repair and Regeneration: Ten Years of Research and a Literature Review. J. Nippon. Med. Sch. 2009, 76, 56–66. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Takahashi, K.; Yamanaka, S. Induction of Pluripotent Stem Cells from Mouse Embryonic and Adult Fibroblast Cultures by Defined Factors. Cell 2006, 126, 663–676. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Young, F.E. A Time for Restraint. Science 2000, 287, 1424. [Google Scholar] [CrossRef]
- Lenoir, N. Europe Confronts the Embryonic Stem Cell Research Challenge. Science 2000, 287, 1425–1427. [Google Scholar] [CrossRef]
- Stenderup, K.; Justesen, J.; Clausen, C.; Kassem, M. Aging is associated with decreased maximal life span and accelerated senescence of bone marrow stromal cells. Bone 2003, 33, 919–926. [Google Scholar] [CrossRef]
- Zuk, P.A.; Zhu, M.; Ashjian, P.; De Ugarte, D.A.; Huang, J.I.; Mizuno, H.; Alfonso, Z.C.; Fraser, J.K.; Benhaim, P.; Hedrick, M.H. Human Adipose Tissue Is a Source of Multipotent Stem Cells. Mol. Biol. Cell 2002, 13, 4279–4295. [Google Scholar] [CrossRef] [PubMed]
- Östman, J.; Arner, P.; Engfeldt, P.; Kager, L. Regional differences in the control of lipolysis in human adipose tissue. Metabolism 1979, 28, 1198–1205. [Google Scholar] [CrossRef]
- Lundgren, M.; Burén, J.; Ruge, T.; Myrnäs, T.; Eriksson, J.W. Glucocorticoids Down-Regulate Glucose Uptake Capacity and Insulin-Signaling Proteins in Omental But Not Subcutaneous Human Adipocytes. J. Clin. Endocrinol. Metab. 2004, 89, 2989–2997. [Google Scholar] [CrossRef] [PubMed]
- Schmidt, B.; Horsley, V. Unravelling hair follicle-adipocyte communication. Exp. Dermatol. 2012, 21, 827–830. [Google Scholar] [CrossRef] [Green Version]
- Festa, E.; Fretz, J.; Berry, R.; Schmidt, B.; Rodeheffer, M.; Horowitz, M.; Horsley, V. Adipocyte Lineage Cells Contribute to the Skin Stem Cell Niche to Drive Hair Cycling. Cell 2011, 146, 761–771. [Google Scholar] [CrossRef] [Green Version]
- Tian, Y.; Pamela, J.J.; Robert, M.L.; Da-Wen, Y.; Tadashige, S.; Yi, G.; Qiong, C.; Kevin, G.; Irwin, M.F.; Tung, T.S. Osteopontin Gene is Expressed in the Dermal Papilla of Pelage Follicles in a Hair-Cycle-Dependent Manner. J. Investig. Dermatol. 2001, V-117, 1554–1558. [Google Scholar] [CrossRef]
- Peerani, R.; Rao, B.; Bauwens, C.; Yin, T.; Wood, G.; Nagy, A.; Kumacheva, E.; Zandstra, P.W. Niche-mediated control of human embryonic stem cell self-renewal and differentiation. EMBO J. 2007, 26, 4744–4755. [Google Scholar] [CrossRef] [Green Version]
- Pawitan, J.A. Prospect of Stem Cell Conditioned Medium in Regenerative Medicine. BioMed Res. Int. 2014, 2014, 1–14. [Google Scholar] [CrossRef] [Green Version]
- Kazi, T.; Niibe, I.; Nishikawa, A.; Matsuzaki, T. Optimal stimulation toward the dermal papilla lineage can be promoted by combined use of osteogenic and adipogenic inducers. FEBS Open Bio 2019, 10, 197–210. [Google Scholar] [CrossRef]
- Marx, V. A better brew. Nature 2013, 496, 253–258. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ahmed, M.I.; Alam, M.; Emelianov, V.U.; Poterlowicz, K.; Patel, A.; Sharov, A.A.; Mardaryev, A.; Botchkareva, N.V.; Guiu, J.; Bergen, D.J.; et al. MicroRNA-214 controls skin and hair follicle development by modulating the activity of the Wnt pathway. J. Cell Biol. 2014, 207, 549–567. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Han, L.; Liu, B.; Chen, X.; Chen, H.; Deng, W.; Yang, C.; Ji, B.; Wan, M. Activation of Wnt/β-catenin signaling is involved in hair growth-promoting effect of 655-nm red light and LED in in vitro culture model. Lasers Med Sci. 2018, 33, 637–645. [Google Scholar] [CrossRef] [PubMed]
- Zhu, N.; Huang, K.; Liu, Y.; Zhang, H.; Lin, E.; Zeng, Y.; Li, H.; Xu, Y.; Cai, B.; Yuan, Y.; et al. miR-195-5p Regulates Hair Follicle Inductivity of Dermal Papilla Cells by Suppressing Wnt/β-Catenin Activation. BioMed Res. Int. 2018, 2018, 1–13. [Google Scholar] [CrossRef] [Green Version]
- Hu, S.; Li, Z.; Lutz, H.; Huang, K.; Su, T.; Cores, J.; Dinh, P.-U.C.; Cheng, K. Dermal exosomes containing miR-218-5p promote hair regeneration by regulating β-catenin signaling. Sci. Adv. 2020, 6, eaba1685. [Google Scholar] [CrossRef] [PubMed]
- Valadi, H.; Ekstrom, K.; Bossios, A.; Sjöstrand, M.; Lee, J.J.; Lötvall, J.O. Exosome-mediated transfer of mRNAs and microRNAs is a novel mechanism of genetic exchange between cells. Nat. Cell Biol. 2007, 9, 654–659. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- O’Brien, K.; Breyne, K.; Ughetto, S.; Laurent, L.C.; Breakefield, X.O. RNA delivery by extracellular vesicles in mammalian cells and its applications. Nat. Rev. Mol. Cell Biol. 2020, 21, 585–606. [Google Scholar] [CrossRef]
- Jahoda, C.; Reynolds, A.; Chaponnier, C.; Forester, J.; Gabbiani, G. Smooth muscle alpha-actin is a marker for hair follicle dermis in vivo and in vitro. J. Cell Sci. 1991, 99 Pt 3, 627–636. [Google Scholar] [CrossRef]
- Driskell, R.; Giangreco, A.; Jensen, K.; Mulder, K.W.; Watt, F.M. Sox2-positive dermal papilla cells specify hair follicle type in mammalian epidermis. Development 2009, 136, 2815–2823. [Google Scholar] [CrossRef] [Green Version]
- Feng, R.; Wen, J. Overview of the roles of Sox2 in stem cell and development. Biol. Chem. 2015, 396, 883–891. [Google Scholar] [CrossRef] [PubMed]
- Makiko, I.; Setsuya, A.; Yumiko, Y.; Hachiro, T. Human Follicular Papilla Cells Carry Out Nonadipose Tissue Production of Leptin. J. Investig. Dermatol. 2001, 117, 1349–1356. [Google Scholar] [CrossRef]
- Lin, W.H.; Xiang, L.J.; Shi, H.X.; Zhang, J.; Jiang, L.P.; Cai, P.T.; Lin, Z.L.; Lin, B.B.; Huang, Y.; Zhang, H.L.; et al. Fibroblast growth factors stimulate hair growth through β-catenin and Shh expression in C57BL/6 mice. BioMed Res. Int. 2015, 2015, 730139. [Google Scholar] [PubMed] [Green Version]
- Cao, L.; Tian, T.; Huang, Y.; Tao, S.; Zhu, X.; Yang, M.; Gu, J.; Feng, G.; Ma, Y.; Xia, R.; et al. Neural progenitor cell-derived nanovesicles promote hair follicle growth via miR-100. J. Nanobiotechnology 2021, 19, 1–13. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Primer Name | Forward Primer (5′-3′) | Reverse Primer (5′-3′) |
|---|---|---|
| versican | ACTAACCCATGCACTACATCAAG | ACTTTTCCAGACAGAGAGCCTT |
| α-smooth muscle actin | GGCACCACTGAACCCTAAGG | ACAATACCAGTTGTACGTCCAGA |
| alpl | GTGACTACCACTCGGGTGAAC | CTCTGGTGGCATCTCGTTATC |
| Osteopontine | TGCAAGCCCTTTGGCAATG | GCCGTCCATATAGCAGCCC |
| leptin | CCAGTTGGTAACAATGCCATGT | TTCAAAGCCGAGGCATTGTTT |
| beta-catenin (ctnb) | ATGGAGCCGGACAGAAAAGC | CTTGCCACTCAGGGAAGGA |
| ncam | AGAAATCAGCGTTGGAGAGTCC- | TCGTCATCATTCCACACCACT |
| CD44 | CACCATTGCCTCAACTGTGC | TTGTGGGCTCCTGAGTCTGA |
| CD45 | ACCACCAGGTGAATGTCAATTT | CTTGCTTTCCCTCGGTTCTTT |
| CD90 | TGCTCTCAGTCTTGCAGGTG | TGGATGGAGTTATCCTTGGTGTT |
| Pdgf-a | ACATGAGGGGACTCTGGGAA | ACAGTGAAGCCCAACAGCTT |
| Pdgf-b | ATTGTGCGAAAGAAGCCCATC | GGGTCACTACTGTCTCACACTT |
| beta-Actin | GGCTGTATTCCCCTCCATCG | CCAGTTGGTAACAATGCCATGT |
| miRNA Name | Mature miRNA Sequence |
|---|---|
| mir-218-5p | UUGUGCUUGAUCUAACCAUGU |
| mir-214-5p | UGCCUGUCUACACUUGCUGUGC |
| mir-195-5p | UAGCAGCACAGAAAUAUUGGC |
| Internal control U6sn | GTGCTCGCTTCGGCAGCACATATACTAAAATTGGAACGATACAGAGAAATTACATGGCCCCTGCGCAAGGATGACACGCAAATTCGTGAAGCGTTCCATATTTT |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kazi, T.; Nagata, A.; Nakagawa, T.; Matsuzaki, T.; Inui, S. Dermal Papilla Cell-Derived Extracellular Vesicles Increase Hair Inductive Gene Expression in Adipose Stem Cells via β-Catenin Activation. Cells 2022, 11, 202. https://doi.org/10.3390/cells11020202
Kazi T, Nagata A, Nakagawa T, Matsuzaki T, Inui S. Dermal Papilla Cell-Derived Extracellular Vesicles Increase Hair Inductive Gene Expression in Adipose Stem Cells via β-Catenin Activation. Cells. 2022; 11(2):202. https://doi.org/10.3390/cells11020202
Chicago/Turabian StyleKazi, Taheruzzaman, Abir Nagata, Takatoshi Nakagawa, Takashi Matsuzaki, and Shigeki Inui. 2022. "Dermal Papilla Cell-Derived Extracellular Vesicles Increase Hair Inductive Gene Expression in Adipose Stem Cells via β-Catenin Activation" Cells 11, no. 2: 202. https://doi.org/10.3390/cells11020202