
Supramolecular Hydrogel-Wrapped Gingival Mesenchymal Stem Cells in Cutaneous Radiation Injury
 and
and 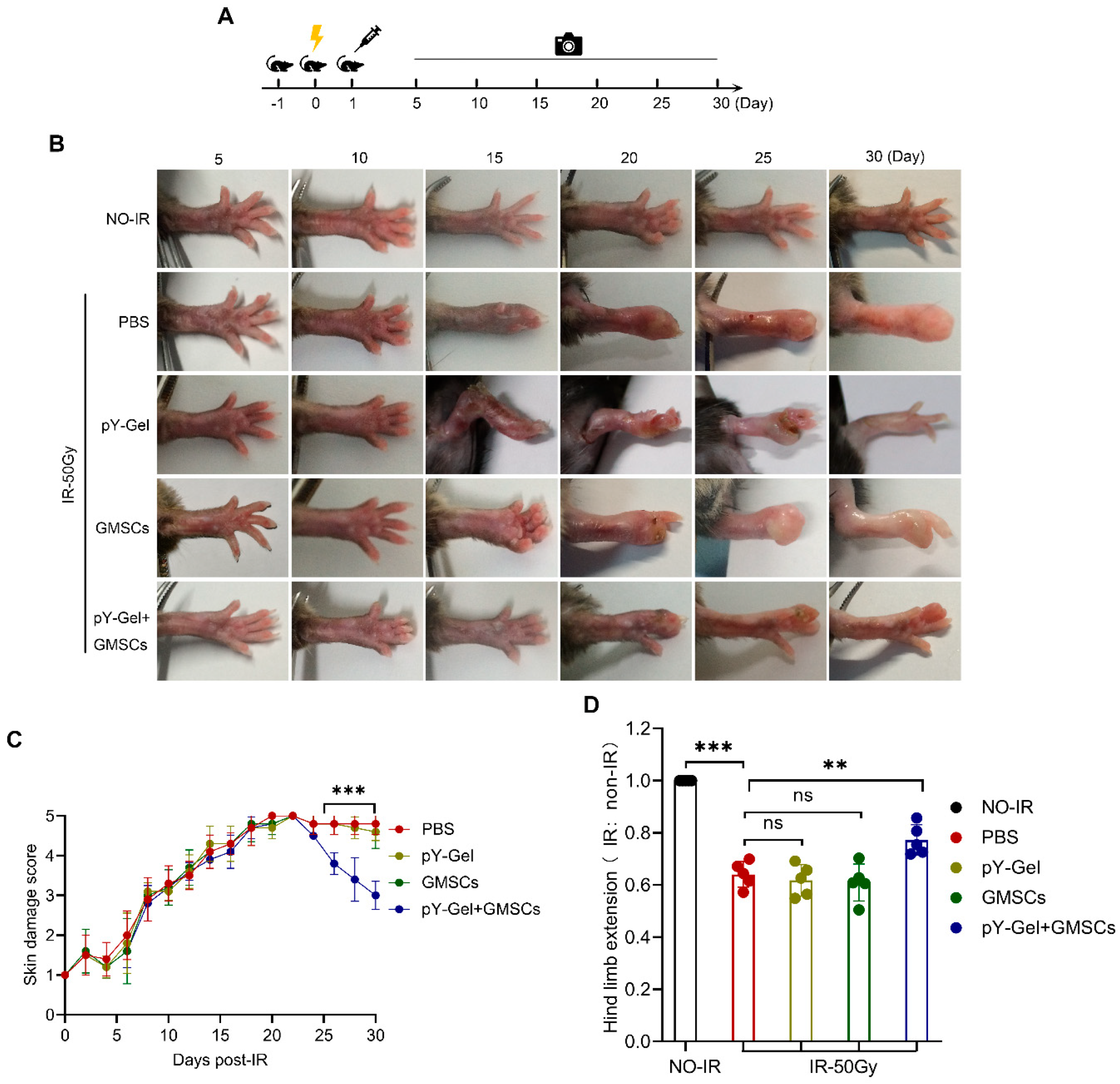{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Cell Culture and Preparation of Conditioned Medium
2.2. Animals and Radiation-Induced Skin Injury Model
2.3. Nap-GDFDFpDY (pY-Gel) Self-Assembling Peptide Hydrogels
2.4. Transplantation of GMSCs in Living Mice and Wound Assessment
2.5. Histological Examination
2.6. Western Blot Analysis
2.7. Quantitative Real-Time PCR Analysis
2.8. Proliferation Assay
2.9. Colony Formation Assay
2.10. Scratch Wound Healing Assay
2.11. Transwell Migration Assay
2.12. Immunofluorescence and Quantitative Analysis of Individual Cells
2.13. Cell Cycle Analysis
2.14. RNA Interference
2.15. Statistical Analysis
3. Results
3.1. Stem Cell Encapsulation by Nap-GDFDFpDY (pY-Gel) Self-Assembling Peptide Hydrogels
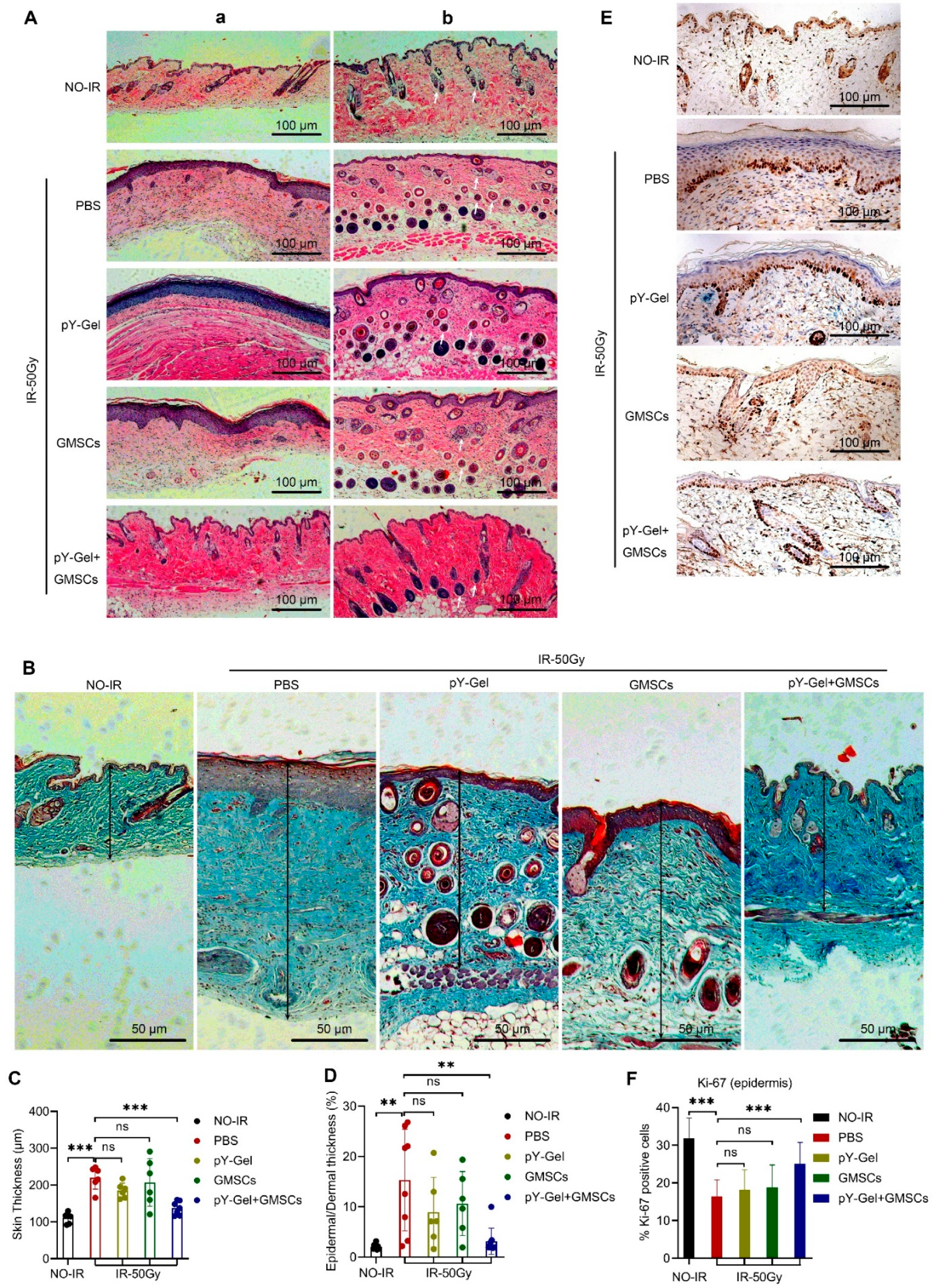
3.2. Evaluation of Healing Quality of the CRI
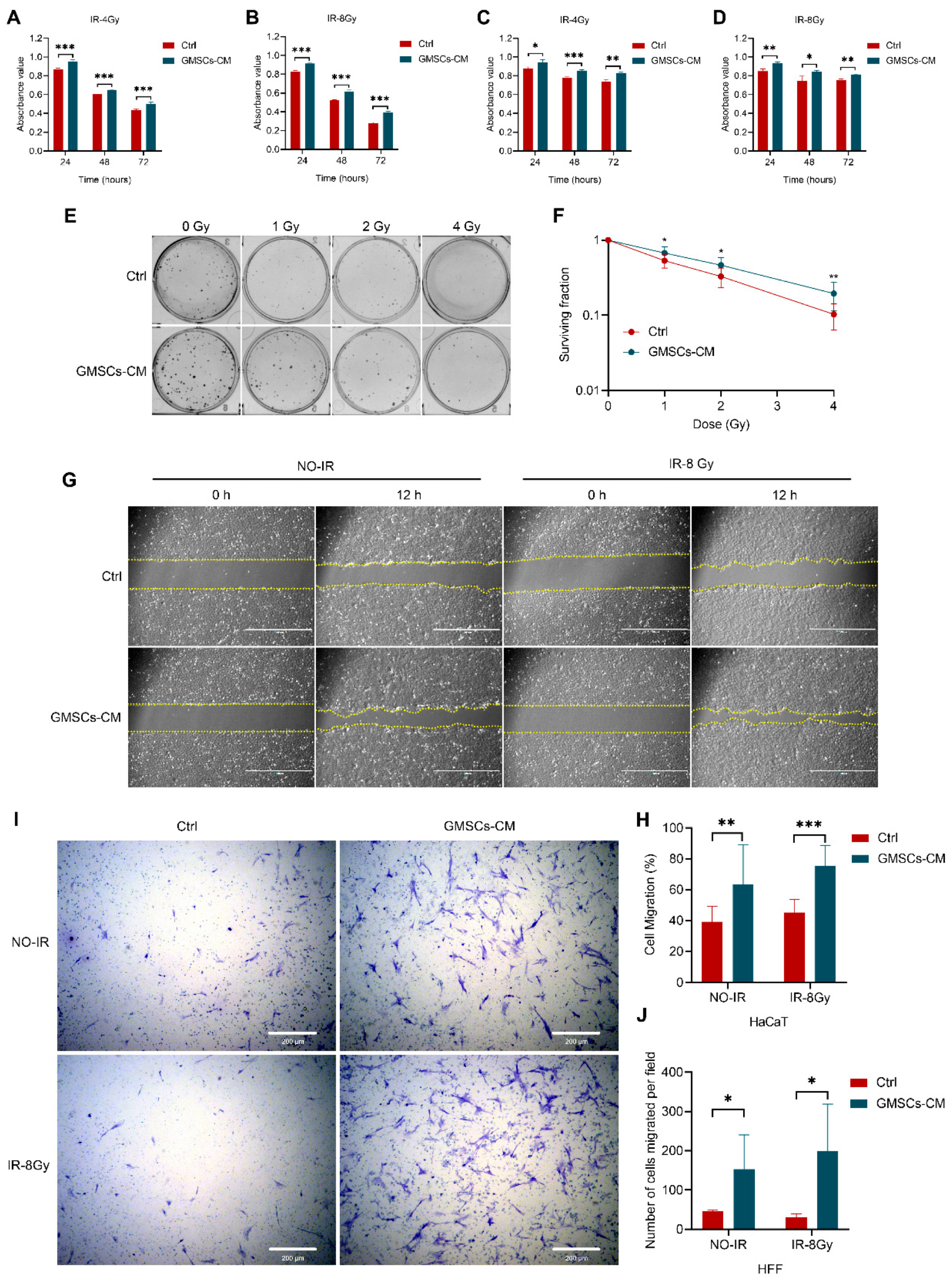
3.3. GMSCs Promote the Proliferation and Migration of Skin Cells
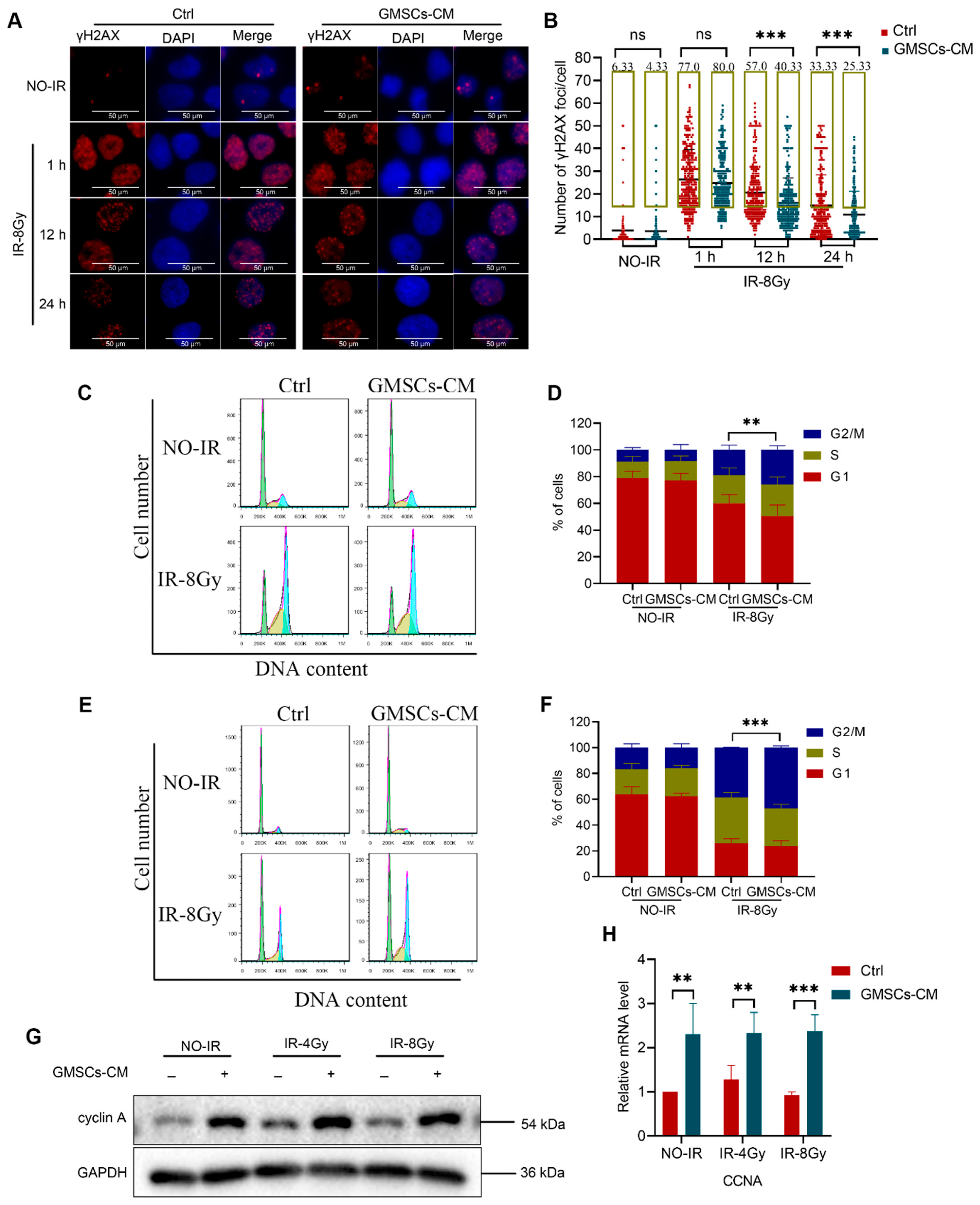
3.4. GMSCs Enhance the Ability of Skin Cells to Repair DNA Damage
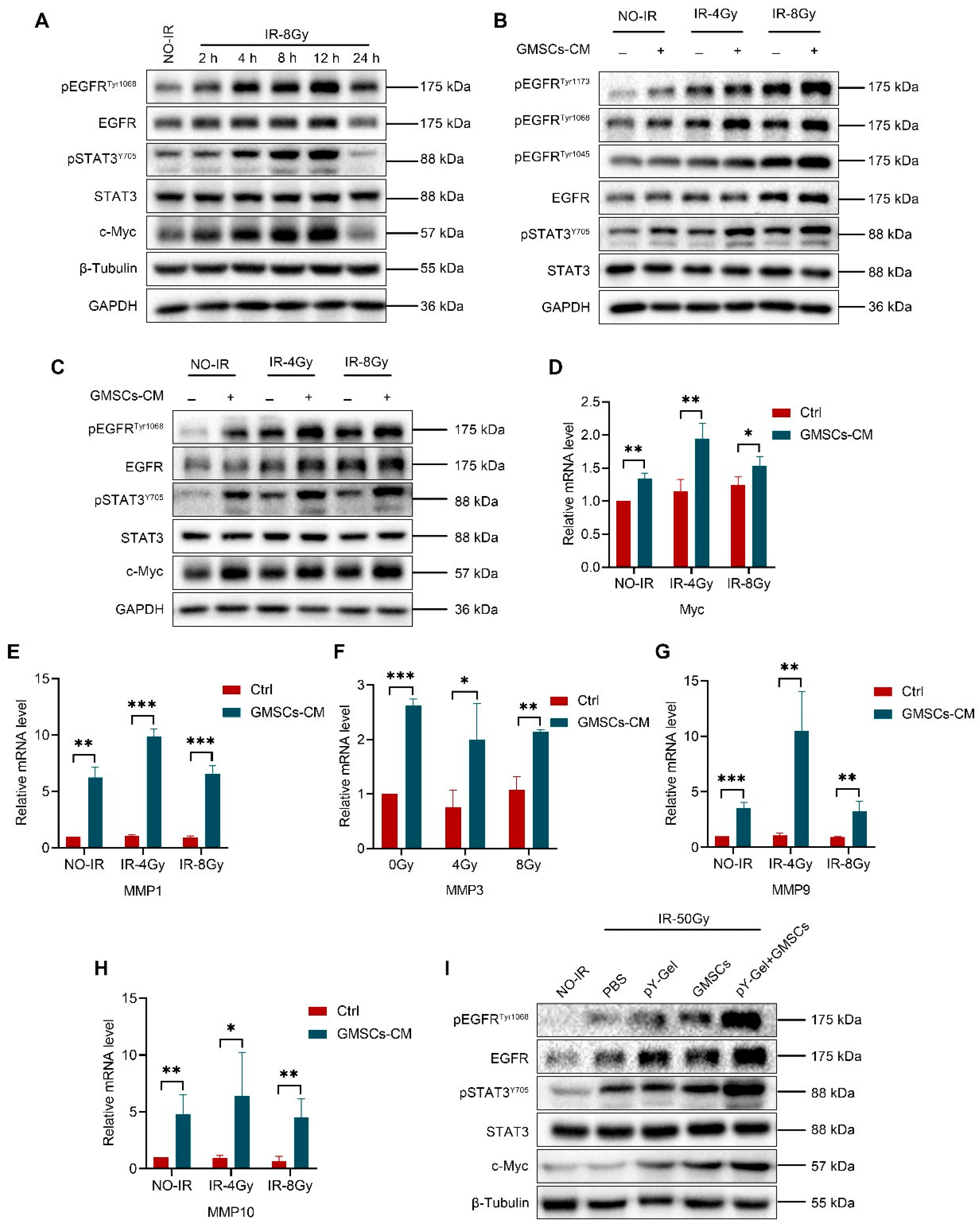
3.5. GMSCs Regulate IR-Induced Activation of the EGFR/STAT3 Signaling Pathway
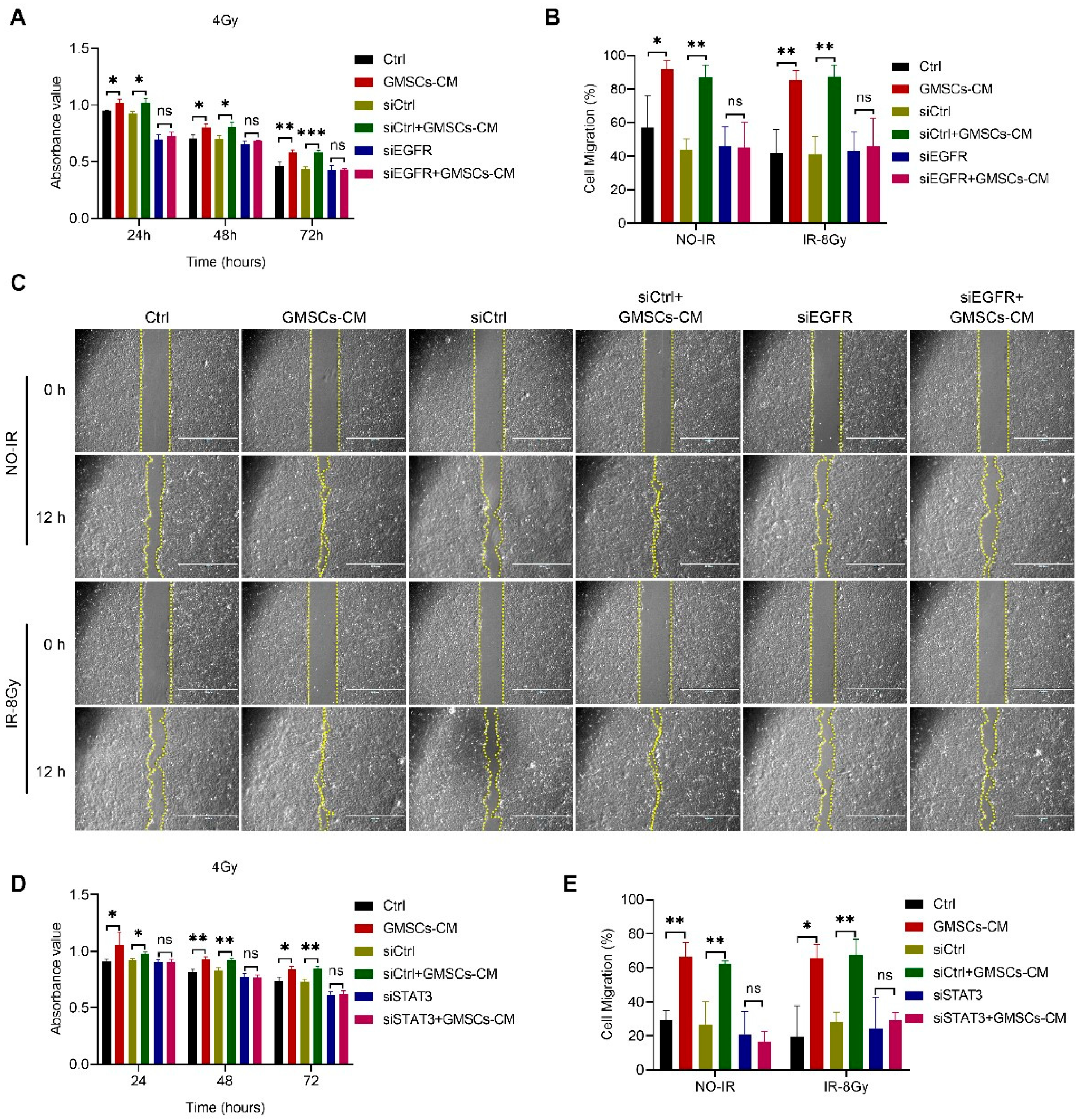
3.6. GMSCs Affect Cell Radiosensitivity by Regulating the EGFR/STAT3 Signaling Pathway
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
References
- Kamiya, K.; Ozasa, K.; Akiba, S.; Niwa, O.; Kodama, K.; Takamura, N.; Zaharieva, E.K.; Kimura, Y.; Wakeford, R. Long-term effects of radiation exposure on health. Lancet 2015, 386, 469–478. [Google Scholar] [CrossRef]
- Soriano, J.L.; Calpena, A.C.; Souto, E.B.; Clares, B. Therapy for prevention and treatment of skin ionizing radiation damage: A review. Int. J. Radiat. Biol. 2019, 95, 537–553. [Google Scholar] [CrossRef] [PubMed]
- Rodgers, K.; Jadhav, S.S. The application of mesenchymal stem cells to treat thermal and radiation burns. Adv. Drug Deliv. Rev. 2018, 123, 75–81. [Google Scholar] [CrossRef] [PubMed]
- Ryan, J.L. Ionizing radiation: The good, the bad, and the ugly. J. Investig. Dermatol. 2012, 132, 985–993. [Google Scholar] [CrossRef] [PubMed]
- Prise, K.M.; Saran, A. Concise review: Stem cell effects in radiation risk. Stem Cells 2011, 29, 1315–1321. [Google Scholar] [CrossRef]
- Su, Y.; Meador, J.A.; Geard, C.R.; Balajee, A.S. Analysis of ionizing radiation-induced DNA damage and repair in three-dimensional human skin model system. Exp. Dermatol. 2010, 19, e16–e22. [Google Scholar] [CrossRef]
- Singh, M.; Alavi, A.; Wong, R.; Akita, S. Radiodermatitis: A review of our current understanding. Am. J. Clin. Dermatol. 2016, 17, 277–292. [Google Scholar] [CrossRef]
- Salvo, N.; Barnes, E.; van Draanen, J.; Stacey, E.; Mitera, G.; Breen, D.; Giotis, A.; Czarnota, G.; Pang, J.; De Angelis, C. Prophylaxis and management of acute radiation-induced skin reactions: A systematic review of the literature. Curr. Oncol. 2010, 17, 94–112. [Google Scholar] [CrossRef]
- Hade, M.D.; Suire, C.N.; Suo, Z. Mesenchymal stem cell-derived exosomes: Applications in regenerative medicine. Cells 2021, 10, 1959. [Google Scholar] [CrossRef]
- Jo, H.; Brito, S.; Kwak, B.M.; Park, S.; Lee, M.G.; Bin, B.H. Applications of mesenchymal stem cells in skin regeneration and rejuvenation. Int. J. Mol. Sci. 2021, 22, 2410. [Google Scholar] [CrossRef]
- Jiang, D.; Qi, Y.; Walker, N.G.; Sindrilaru, A.; Hainzl, A.; Wlaschek, M.; MacNeil, S.; Scharffetter-Kochanek, K. The effect of adipose tissue derived MSCs delivered by a chemically defined carrier on full-thickness cutaneous wound healing. Biomaterials 2013, 34, 2501–2515. [Google Scholar] [CrossRef] [PubMed]
- Xu, G.; Zhang, L.; Ren, G.; Yuan, Z.; Zhang, Y.; Zhao, R.C.; Shi, Y. Immunosuppressive properties of cloned bone marrow mesenchymal stem cells. Cell Res. 2007, 17, 240–248. [Google Scholar] [CrossRef] [PubMed]
- Rustad, K.C.; Wong, V.W.; Sorkin, M.; Glotzbach, J.P.; Major, M.R.; Rajadas, J.; Longaker, M.T.; Gurtner, G.C. Enhancement of mesenchymal stem cell angiogenic capacity and stemness by a biomimetic hydrogel scaffold. Biomaterials 2012, 33, 80–90. [Google Scholar] [CrossRef] [PubMed]
- Su, N.; Gao, P.L.; Wang, K.; Wang, J.Y.; Zhong, Y.; Luo, Y. Fibrous scaffolds potentiate the paracrine function of mesenchymal stem cells: A new dimension in cell-material interaction. Biomaterials 2017, 141, 74–85. [Google Scholar] [CrossRef]
- Selek, O.; Buluç, L.; Muezzinoğlu, B.; Ergün, R.E.; Ayhan, S.; Karaöz, E. Mesenchymal stem cell application improves tendon healing via anti-apoptotic effect (Animal study). Acta Orthop. Traumatol. Turc. 2014, 48, 187–195. [Google Scholar] [CrossRef]
- Mazini, L.; Rochette, L.; Admou, B.; Amal, S.; Malka, G. Hopes and limits of Adipose-Derived Stem Cells (ADSCs) and Mesenchymal Stem Cells (MSCs) in wound healing. Int. J. Mol. Sci. 2020, 21, 1306. [Google Scholar] [CrossRef]
- Sasaki, M.; Abe, R.; Fujita, Y.; Ando, S.; Inokuma, D.; Shimizu, H. Mesenchymal stem cells are recruited into wounded skin and contribute to wound repair by transdifferentiation into multiple skin cell type. J. Immunol. 2008, 180, 2581–2587. [Google Scholar] [CrossRef]
- Kim, J.W.; Lee, J.H.; Lyoo, Y.S.; Jung, D.I.; Park, H.M. The effects of topical mesenchymal stem cell transplantation in canine experimental cutaneous wounds. Vet. Dermatol. 2013, 24, 242-e53. [Google Scholar] [CrossRef]
- Maria, A.T.; Toupet, K.; Bony, C.; Pirot, N.; Vozenin, M.C.; Petit, B.; Roger, P.; Batteux, F.; Le Quellec, A.; Jorgensen, C.; et al. Antifibrotic, antioxidant, and immunomodulatory effects of mesenchymal stem cells in HOCl-induced systemic sclerosis. Arthritis Rheumatol. 2016, 68, 1013–1025. [Google Scholar] [CrossRef]
- Jiang, M.; Yu, Y.; Luo, J.; Gao, Q.; Zhang, L.; Wang, Q.; Zhao, J. Bone marrow-derived mesenchymal stem cells expressing thioredoxin 1 attenuate bleomycin-induced skin fibrosis and oxidative stress in scleroderma. J. Investig. Dermatol. 2017, 137, 1223–1233. [Google Scholar] [CrossRef] [Green Version]
- Liu, S.; Jiang, L.; Li, H.; Shi, H.; Luo, H.; Zhang, Y.; Yu, C.; Jin, Y. Mesenchymal stem cells prevent hypertrophic scar formation via inflammatory regulation when undergoing apoptosis. J. Investig. Dermatol. 2014, 134, 2648–2657. [Google Scholar] [CrossRef] [PubMed]
- Guillamat-Prats, R. The Role of MSC in wound healing, scarring and regeneration. Cells 2021, 10, 1729. [Google Scholar] [CrossRef] [PubMed]
- Casado-Díaz, A.; Quesada-Gómez, J.M.; Dorado, G. Extracellular vesicles derived from Mesenchymal Stem Cells (MSC) in regenerative medicine: Applications in skin wound healing. Front. Bioeng. Biotechnol. 2020, 8, 146. [Google Scholar] [CrossRef] [PubMed]
- Ahangar, P.; Mills, S.J.; Smith, L.E.; Gronthos, S.; Cowin, A.J. Human gingival fibroblast secretome accelerates wound healing through anti-inflammatory and pro-angiogenic mechanisms. NPJ Regen. Med. 2020, 5, 24. [Google Scholar] [CrossRef] [PubMed]
- Shi, Q.; Qian, Z.; Liu, D.; Sun, J.; Wang, X.; Liu, H.; Xu, J.; Guo, X. GMSC-derived exosomes combined with a chitosan/silk hydrogel sponge accelerates wound healing in a diabetic rat skin defect model. Front. Physiol. 2017, 8, 904. [Google Scholar] [CrossRef] [PubMed]
- Xu, X.; Chen, C.; Akiyama, K.; Chai, Y.; Le, A.D.; Wang, Z.; Shi, S. Gingivae contain neural-crest- and mesoderm-derived mesenchymal stem cells. J. Dent. Res. 2013, 92, 825–832. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Q.; Shi, S.; Liu, Y.; Uyanne, J.; Shi, Y.; Shi, S.; Le, A.D. Mesenchymal stem cells derived from human gingiva are capable of immunomodulatory functions and ameliorate inflammation-related tissue destruction in experimental colitis. J. Immunol. 2009, 183, 7787–7798. [Google Scholar] [CrossRef]
- Tomar, G.B.; Srivastava, R.K.; Gupta, N.; Barhanpurkar, A.P.; Pote, S.T.; Jhaveri, H.M.; Mishra, G.C.; Wani, M.R. Human gingiva-derived mesenchymal stem cells are superior to bone marrow-derived mesenchymal stem cells for cell therapy in regenerative medicine. Biochem. Biophys. Res. Commun. 2010, 393, 377–383. [Google Scholar] [CrossRef]
- Tang, L.; Li, N.; Xie, H.; Jin, Y. Characterization of mesenchymal stem cells from human normal and hyperplastic gingiva. J. Cell Physiol. 2011, 226, 832–842. [Google Scholar] [CrossRef]
- Kou, X.; Xu, X.; Chen, C.; Sanmillan, M.L.; Cai, T.; Zhou, Y.; Giraudo, C.; Le, A.; Shi, S. The Fas/Fap-1/Cav-1 complex regulates IL-1RA secretion in mesenchymal stem cells to accelerate wound healing. Sci. Transl. Med. 2018, 10, eaai8524. [Google Scholar] [CrossRef] [Green Version]
- Moshaverinia, A.; Xu, X.; Chen, C.; Ansari, S.; Zadeh, H.H.; Snead, M.L.; Shi, S. Application of stem cells derived from the periodontal ligament or gingival tissue sources for tendon tissue regeneration. Biomaterials 2014, 35, 2642–2650. [Google Scholar] [CrossRef] [PubMed]
- Ansari, S.; Pouraghaei Sevari, S.; Chen, C.; Sarrion, P.; Moshaverinia, A. RGD-modified alginate-GelMA hydrogel sheet containing gingival mesenchymal stem cells: A unique platform for wound healing and soft tissue regeneration. ACS Biomater. Sci. Eng. 2021, 7, 3774–3782. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Luo, Z.; Wang, Y.; He, T.; Yang, C.; Ren, C.; Ma, L.; Gong, C.; Li, X.; Yang, Z. Enzyme-catalyzed formation of supramolecular hydrogels as promising vaccine adjuvants. Adv. Funct. Mater. 2016, 26, 1822–1829. [Google Scholar] [CrossRef]
- Luo, Z.; Wu, Q.; Yang, C.; Wang, H.; He, T.; Wang, Y.; Wang, Z.; Chen, H.; Li, X.; Gong, C.; et al. A powerful CD8+ T-cell stimulating D-tetra-peptide hydrogel as a very promising vaccine adjuvant. Adv. Mater. 2017, 29, 1601776. [Google Scholar] [CrossRef]
- Wang, Y.; Li, X.; Zhang, Y.; Wang, L.; Yang, Z. A supramolecular hydrogel to boost the production of antibodies for phosphorylated proteins. Chem. Commun. 2019, 55, 12388–12391. [Google Scholar] [CrossRef]
- Liu, M.; Wang, Z.; Feng, D.; Shang, Y.; Li, X.; Liu, J.; Li, C.; Yang, Z. An insulin-inspired supramolecular hydrogel for prevention of type 1 diabetes. Adv. Sci. 2021, 8, 2003599. [Google Scholar] [CrossRef]
- Xiao, Z.; Su, Y.; Yang, S.; Yin, L.; Wang, W.; Yi, Y.; Fenton, B.M.; Zhang, L.; Okunieff, P. Protective effect of esculentoside A on radiation-induced dermatitis and fibrosis. Int. J. Radiat. Oncol. Biol. Phys. 2006, 65, 8829. [Google Scholar] [CrossRef]
- Borrelli, M.R.; Diaz Deleon, N.M.; Adem, S.; Patel, R.A.; Mascharak, S.; Shen, A.H.; Irizarry, D.; Nguyen, D.; Momeni, A.; Longaker, M.T.; et al. Fat grafting rescues radiation-induced joint contracture. Stem Cells 2020, 38, 382–389. [Google Scholar] [CrossRef]
- Wang, Z.; Chen, Z.; Jiang, Z.; Luo, P.; Liu, L.; Huang, Y.; Wang, H.; Wang, Y.; Long, L.; Tan, X.; et al. Cordycepin prevents radiation ulcer by inhibiting cell senescence via NRF2 and AMPK in rodents. Nat. Commun. 2019, 10, 2538. [Google Scholar] [CrossRef]
- Rousselle, P.; Braye, F.; Dayan, G. Re-epithelialization of adult skin wounds: Cellular mechanisms and therapeutic strategies. Adv. Drug Deliv. Rev. 2019, 146, 344–365. [Google Scholar] [CrossRef]
- Miyamoto, T.; Akutsu, S.N.; Tauchi, H.; Kudo, Y.; Tashiro, S.; Yamamoto, T.; Matsuura, S. Exploration of genetic basis underlying individual differences in radiosensitivity within human populations using genome editing technology. J. Radiat. Res. 2018, 59, ii75–ii82. [Google Scholar] [CrossRef]
- Miyake, T.; Shimada, M.; Matsumoto, Y.; Okino, A. DNA damage response after ionizing radiation exposure in skin keratinocytes derived from human-induced pluripotent stem cells. Int. J. Radiat. Oncol. Biol. Phys. 2019, 105, 193–205. [Google Scholar] [CrossRef] [PubMed]
- Kinner, A.; Wu, W.; Staudt, C.; Iliakis, G. Gamma-H2AX in recognition and signaling of DNA double-strand breaks in the context of chromatin. Nucleic Acids Res. 2008, 36, 5678–5694. [Google Scholar] [CrossRef] [PubMed]
- Pawlik, T.M.; Keyomarsi, K. Role of cell cycle in mediating sensitivity to radiotherapy. Int. J. Radiat. Oncol. Biol. Phys. 2004, 59, 928–942. [Google Scholar] [CrossRef] [PubMed]
- Sun, X.; Wang, Y.; Ji, K.; Liu, Y.; Kong, Y.; Nie, S.; Li, N.; Hao, J.; Xie, Y.; Xu, C.; et al. NRF2 preserves genomic integrity by facilitating ATR activation and G2 cell cycle arrest. Nucleic Acids Res. 2020, 48, 9109–9123. [Google Scholar] [CrossRef]
- He, P.; Li, Z.; Xu, F.; Ru, G.; Huang, Y.; Lin, E.; Peng, S. AMPK activity contributes to G2 arrest and DNA damage decrease via p53/p21 pathways in oxidatively damaged mouse zygotes. Front. Cell Dev. Biol. 2020, 8, 539485. [Google Scholar] [CrossRef]
- Woodburn, J.R. The epidermal growth factor receptor and its inhibition in cancer therapy. Pharmacol. Ther. 1999, 82, 241–250. [Google Scholar] [CrossRef]
- Schneider, M.R.; Werner, S.; Paus, R.; Wolf, E. Beyond wavy hairs: The epidermal growth factor receptor and its ligands in skin biology and pathology. Am. J. Pathol. 2008, 173, 14–24. [Google Scholar] [CrossRef]
- Pastore, S.; Lulli, D.; Fidanza, P.; Potapovich, A.I.; Kostyuk, V.A.; De Luca, C.; Mikhal’chik, E.; Korkina, L.G. Plant polyphenols regulate chemokine expression and tissue repair in human keratinocytes through interaction with cytoplasmic and nuclear components of epidermal growth factor receptor system. Antioxid. Redox Signal. 2012, 16, 314–328. [Google Scholar] [CrossRef]
- Williams, J.P.; Brown, S.L.; Georges, G.E.; Hauer-Jensen, M.; Hill, R.P.; Huser, A.K.; Kirsch, D.G.; Macvittie, T.J.; Mason, K.A.; Medhora, M.M.; et al. Animal models for medical countermeasures to radiation exposure. Radiat. Res. 2010, 173, 557–578. [Google Scholar] [CrossRef] [Green Version]
- Graczyk-Jarzynka, A.; Sobiak, B.; Mlącki, M.; Wilanowski, T.; Leśniak, W. S100A6 activates EGFR and its downstream signaling in HaCaT keratinocytes. J. Cell Physiol. 2019, 234, 17561–17569. [Google Scholar] [CrossRef] [PubMed]
- Niyonsaba, F.; Ushio, H.; Nakano, N.; Ng, W.; Sayama, K.; Hashimoto, K.; Nagaoka, I.; Okumura, K.; Ogawa, H. Antimicrobial peptides human beta-defensins stimulate epidermal keratinocyte migration, proliferation and production of proinflammatory cytokines and chemokines. J. Invest. Dermatol. 2007, 127, 594–604. [Google Scholar] [CrossRef]
- Li, W.; Wang, X.; Cheng, J.; Li, J.; Wang, Q.; Zhou, Q.; Li, H.; Xue, J.; Zhang, Y.; Yang, L.; et al. Leucine-rich α-2-glycoprotein-1 promotes diabetic corneal epithelial wound healing and nerve regeneration via regulation of matrix metalloproteinases. Exp. Eye Res. 2020, 196, 108060. [Google Scholar] [CrossRef] [PubMed]
- Melchionna, R.; Bellavia, G.; Romani, M.; Straino, S.; Germani, A.; Di Carlo, A.; Capogrossi, M.C.; Napolitano, M. C/EBPγ regulates wound repair and EGF receptor signaling. J. Investig. Dermatol. 2012, 132, 1908–1917. [Google Scholar] [CrossRef] [PubMed]
- Zheng, X.; Ding, Z.; Cheng, W.; Lu, Q.; Kong, X.; Zhou, X.; Lu, G.; Kaplan, D.L. Microskin-inspired injectable MSC-laden hydrogels for scarless wound healing with hair follicles. Adv. Healthc. Mater. 2020, 9, e2000041. [Google Scholar] [CrossRef]
- Jiang, C.M.; Liu, J.; Zhao, J.Y.; Xiao, L.; An, S.; Gou, Y.C.; Quan, H.X.; Cheng, Q.; Zhang, Y.L.; He, W.; et al. Effects of hypoxia on the immunomodulatory properties of human gingiva-derived mesenchymal stem cells. J. Dent. Res. 2015, 94, 69–77. [Google Scholar] [CrossRef] [PubMed]
- Hasani-Sadrabadi, M.M.; Sarrion, P.; Pouraghaei, S.; Chau, Y.; Ansari, S.; Li, S.; Aghaloo, T.; Moshaverinia, A. An engineered cell-laden adhesive hydrogel promotes craniofacial bone tissue regeneration in rats. Sci. Transl. Med. 2020, 12, eaay6853. [Google Scholar] [CrossRef]
- Silina, E.; Manturova, N.; Stupin, V. Mesenchymal stem cells application in wound tissue healing in old animals. Stem Cells Cloning 2020, 13, 103–116. [Google Scholar] [CrossRef]
- Huang, R.; Wang, J.; Chen, H.; Shi, X.; Wang, X.; Zhu, Y.; Tan, Z. The topography of fibrous scaffolds modulates the paracrine function of Ad-MSCs in the regeneration of skin tissues. Biomater. Sci. 2019, 7, 4248–4259. [Google Scholar] [CrossRef]
- De Gregorio, C.; Contador, D.; Díaz, D.; Cárcamo, C.; Santapau, D.; Lobos-Gonzalez, L.; Acosta, C.; Campero, M.; Carpio, D.; Gabriele, C.; et al. Human adipose-derived mesenchymal stem cell-conditioned medium ameliorates polyneuropathy and foot ulceration in diabetic BKS db/db mice. Stem Cell Res. Ther. 2020, 11, 168. [Google Scholar] [CrossRef]
- Kim, Y.J.; Ahn, H.J.; Lee, S.H.; Lee, M.H.; Kang, K.S. Effects of conditioned media from human umbilical cord blood-derived mesenchymal stem cells in the skin immune response. Biomed. Pharmacother. 2020, 131, 110789. [Google Scholar] [CrossRef] [PubMed]
- Sun, J.; Zhang, Y.; Song, X.; Zhu, J.; Zhu, Q. The healing effects of conditioned medium derived from mesenchymal stem cells on radiation-induced skin wounds in rats. Cell Transpl. 2019, 28, 105–115. [Google Scholar] [CrossRef] [PubMed]
- Saheli, M.; Bayat, M.; Ganji, R.; Hendudari, F.; Kheirjou, R.; Pakzad, M.; Najar, B.; Piryaei, A. Human mesenchymal stem cells-conditioned medium improves diabetic wound healing mainly through modulating fibroblast behaviors. Arch. Dermatol. Res. 2020, 312, 325–336. [Google Scholar] [CrossRef] [PubMed]
- Sadoughi, F.; Hallajzadeh, J.; Asemi, Z.; Mansournia, M.A.; Alemi, F.; Yousefi, B. Signaling pathways involved in cell cycle arrest during the DNA breaks. DNA Repair 2021, 98, 103047. [Google Scholar] [CrossRef] [PubMed]
- Mi, B.; Liu, J.; Liu, Y.; Hu, L.; Liu, Y.; Panayi, A.C.; Zhou, W.; Liu, G. The designer antimicrobial peptide A-hBD-2 facilitates skin wound healing by stimulating keratinocyte migration and proliferation. Cell Physiol. Biochem. 2018, 51, 647–663. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.; Zhou, Z.; Tian, S.; Zhang, K.; An, G.; Zhang, Y.; Ma, R.; Sheng, B.; Wang, T.; Yang, H.; et al. RPRM deletion preserves hematopoietic regeneration by promoting EGFR-dependent DNA repair and hematopoietic stem cell proliferation post ionizing radiation. Cell Biol. Int. 2022. [Google Scholar] [CrossRef]
- Kim, D.; Lee, A.E.; Xu, Q.; Zhang, Q.; Le, A.D. Gingiva-derived mesenchymal stem cells: Potential application in tissue engineering and regenerative medicine—A comprehensive review. Front. Immunol. 2021, 12, 667221. [Google Scholar] [CrossRef]






Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Nie, S.; Ren, C.; Liang, X.; Cai, H.; Sun, H.; Liu, F.; Ji, K.; Wang, Y.; Liu, Q. Supramolecular Hydrogel-Wrapped Gingival Mesenchymal Stem Cells in Cutaneous Radiation Injury. Cells 2022, 11, 3089. https://doi.org/10.3390/cells11193089
Nie S, Ren C, Liang X, Cai H, Sun H, Liu F, Ji K, Wang Y, Liu Q. Supramolecular Hydrogel-Wrapped Gingival Mesenchymal Stem Cells in Cutaneous Radiation Injury. Cells. 2022; 11(19):3089. https://doi.org/10.3390/cells11193089
Chicago/Turabian StyleNie, Shasha, Chunhua Ren, Xin Liang, Hui Cai, Hao Sun, Fengting Liu, Kaihua Ji, Yan Wang, and Qiang Liu. 2022. "Supramolecular Hydrogel-Wrapped Gingival Mesenchymal Stem Cells in Cutaneous Radiation Injury" Cells 11, no. 19: 3089. https://doi.org/10.3390/cells11193089