

Diagnostic Potential of microRNAs in Extracellular Vesicles Derived from Bronchoalveolar Lavage Fluid for Pneumonia—A Preliminary Report
Abstract
:1. Introduction
2. Materials and Methods
2.1. Patients and Samples
2.2. Cell Culture and Treatment
2.3. Purification and Identification of Extracellular Vesicles
2.4. Candidate miRNAs Selection
2.5. RNA Extraction and Quantitative Reverse Transcription PCR
2.6. Statistical Analysis
3. Results
3.1. Patient Characteristics
3.2. Characterization of EVs Derived from Different Samples
3.3. Differentially Expressed miRNAs Specifically Showed in Extracellular Vesicles Derived from Bronchoalveolar Lavage Fluid
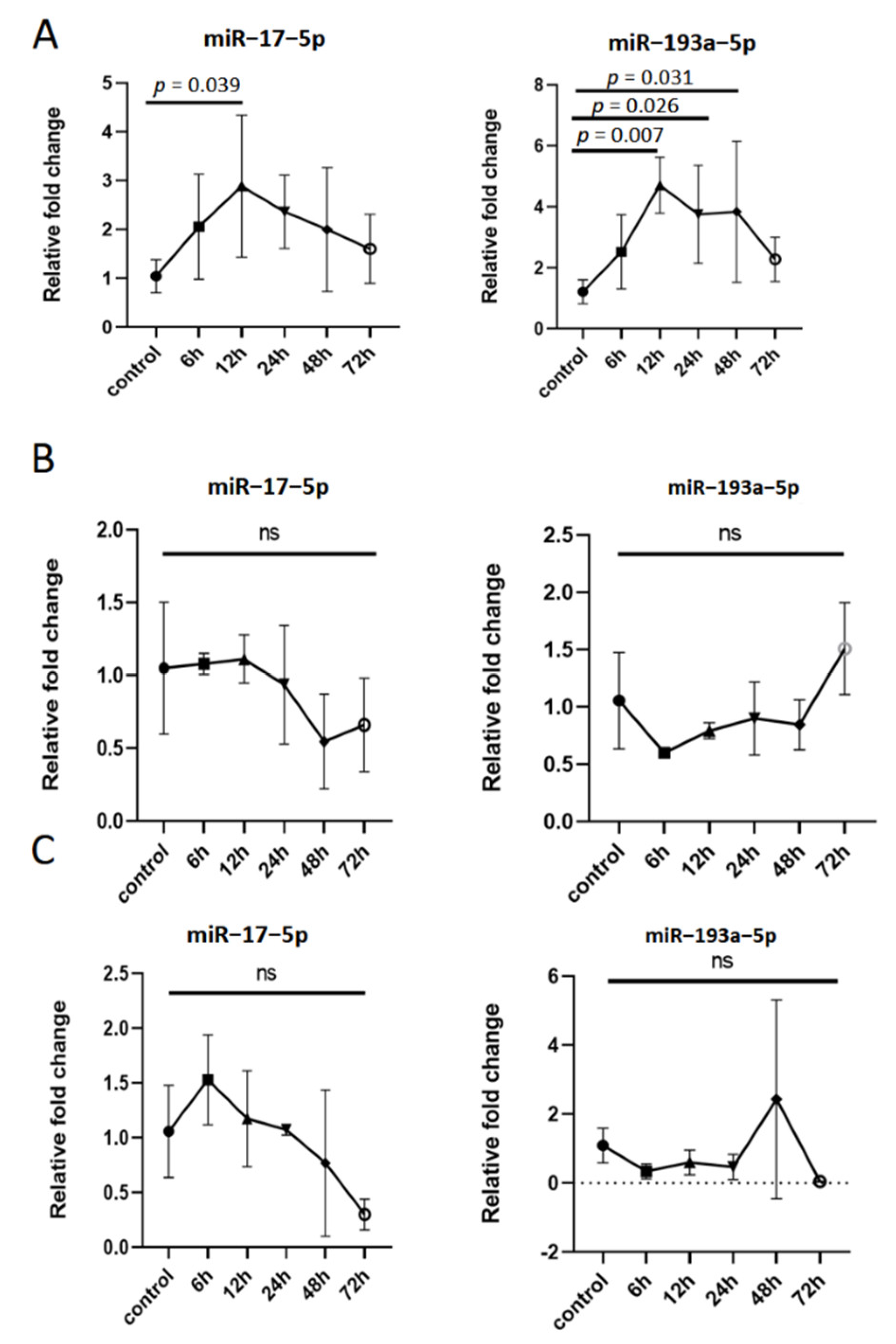
3.4. Inflammatory Stimuli Increases the Expression of miR–17–5p and miR–193a–5p in Extracellular Vesicles Derived from Macrophages
3.5. The Expression of miR–17–5p and miR–193a–5p in Extracellular Vesicles Derived from Macrophages Is Dynamic in Inflammatory Response
3.6. Diagnostic Value of miR–17–5p and miR–193a–5p in BALF-EVs for Pneumonia
3.7. MicroRNAs Target Prediction and Pathway Analysis
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Conflicts of Interest
Abbreviations
References
- World Health Organization. The Top 10 Causes of Death. 2019. Available online: https://www.who.int/news-room/fact-sheets/detail/the-top-10-causes-of-death (accessed on 9 December 2020).
- Zakharkina, T.I.; Martin-Loeches, I.; Matamoros, S.; Povoa, P.; Torres, A.; Kastelijn, J.B.; Hofstra, J.J.; de Wever, B.; de Jong, M.; Schultz, M.J.; et al. The dynamics of the pulmonary microbiome during mechanical ventilation in the intensive care unit and the association with occurrence of pneumonia. Thorax 2017, 72, 803–810. [Google Scholar] [CrossRef] [PubMed]
- Vallés, J.; Martin-Loeches, I.; Torres, A.; Diaz, E.; Seijas, I.; López, M.J.; Garro, P.; Castillo, C.; Garnacho-Montero, J.; Martin, M.; et al. Epidemiology, antibiotic therapy and clinical outcomes of healthcare-associated pneumonia in critically ill patients: A Spanish cohort study. Intensive Care Med. 2014, 40, 572–581. [Google Scholar] [CrossRef] [PubMed]
- Chen, H.; Yin, Y.; Gao, H.; Guo, Y.; Dong, Z.; Wang, X.; Zhang, Y.; Yang, S.; Peng, Q.; Liu, Y.; et al. Clinical Utility of In-house Metagenomic Next-generation Sequencing for the Diagnosis of Lower Respiratory Tract Infections and Analysis of the Host Immune Response. Clin. Infect. Dis. 2020, 71 (Suppl. S4), S416–S426. [Google Scholar] [CrossRef] [PubMed]
- Wunderink, R.G.; Waterer, G. Advances in the causes and management of community acquired pneumonia in adults. BMJ 2017, 358, j2471. [Google Scholar] [CrossRef]
- Sligl, W.I.; Hoang, H.; Eurich, D.T.; Malhotra, A.; Marrie, T.J.; Majumdar, S.R. Macrolide use in the treatment of critically ill patients with pneumonia: Incidence, correlates, timing and outcomes. Can. J. Infect. Dis. Med. Microbiol. 2013, 24, e107–e112. [Google Scholar] [CrossRef]
- Hermann, S.; Brandes, F.; Kirchner, B.; Buschmann, D.; Borrmann, M.; Klein, M.; Kotschote, S.; Bonin, M.; Reithmair, M.; Kaufmann, I.; et al. Diagnostic potential of circulating cell-free microRNAs for community-acquired pneumonia and pneumonia-related sepsis. J. Cell Mol. Med. 2020, 24, 12054–12064. [Google Scholar] [CrossRef]
- Kalil, A.C.; Metersky, M.L.; Klompas, M.; Muscedere, J.; Sweeney, D.A.; Palmer, L.B.; Napolitano, L.M.; O’Grady, N.P.; Bartlett, J.G.; Carratalà, J.; et al. Management of Adults with Hospital-acquired and Ventilator-associated Pneumonia: 2016 Clinical Practice Guidelines by the Infectious Diseases Society of America and the American Thoracic Society. Clin. Infect. Dis. 2016, 63, e61–e111. [Google Scholar] [CrossRef]
- Musher, D.M.; Thorner, A.R. Community-acquired pneumonia. N. Engl. J. Med. 2014, 371, 1619–1628. [Google Scholar] [CrossRef]
- Karakioulaki, M.; Stolz, D. Biomarkers in Pneumonia-Beyond Procalcitonin. Int. J. Mol. Sci. 2019, 20, 2004. [Google Scholar] [CrossRef]
- Canetta, S.; Sourander, A.; Surcel, H.M.; Hinkka-Yli-Salomäki, S.; Leiviskä, J.; Kellendonk, C.; McKeague, I.W.; Brown, A.S. Elevated maternal C-reactive protein and increased risk of schizophrenia in a national birth cohort. Am. J. Psychiatry 2014, 171, 960–968. [Google Scholar] [CrossRef]
- Oprescu, N.; Micheu, M.M.; Scafa-Udriste, A.; Popa-Fotea, N.M.; Dorobantu, M. Inflammatory markers in acute myocardial infarction and the correlation with the severity of coronary heart disease. Ann. Med. 2021, 53, 1041–1047. [Google Scholar] [CrossRef]
- Ito, A.; Ishida, T. Diagnostic markers for community-acquired pneumonia. Ann. Transl. Med. 2020, 8, 609. [Google Scholar] [CrossRef]
- Li, J.C.; Wang, L.Y.; Wang, Y.F.; Mei, M.H.; Shi, L.; Yao, M.L.; Guan, X.D.; Ouyang, B. Serum procalcitonin in patients with pulmonary infection and central nervous system injury. Zhonghua Yi Xue Za Zhi 2017, 97, 2940–2945. [Google Scholar]
- Wussler, D.; Kozhuharov, N.; Oliveira, M.T.; Bossa, A.; Sabti, Z.; Nowak, A.; Murray, K.; de Lavallaz, J.d.F.; Badertscher, P.; Twerenbold, R.; et al. Clinical Utility of Procalcitonin in the Diagnosis of Pneumonia. Clin. Chem. 2019, 65, 1532–1542. [Google Scholar] [CrossRef]
- Liu, Z.; Yan, J.; Tong, L.; Liu, S.; Zhang, Y. The role of exosomes from BALF in lung disease. J. Cell Physiol. 2021. [Google Scholar] [CrossRef]
- Costabel, U.; Guzman, J. Bronchoalveolar lavage in interstitial lung disease. Curr. Opin. Pulm. Med. 2001, 7, 255–261. [Google Scholar] [CrossRef]
- Pastor, L.; Vera, E.; Marin, J.M.; Sanz-Rubio, D. Extracellular Vesicles from Airway Secretions: New Insights in Lung Diseases. Int. J. Mol. Sci. 2021, 22, 583. [Google Scholar] [CrossRef]
- Alipoor, S.D.; Mortaz, E.; Garssen, J.; Movassaghi, M.; Mirsaeidi, M.; Adcock, I.M. Exosomes and Exosomal miRNA in Respiratory Diseases. Mediat. Inflamm. 2016, 2016, 5628404. [Google Scholar] [CrossRef]
- He, L.; Hannon, G.J. MicroRNAs: Small RNAs with a big role in gene regulation. Nat. Rev. Genet. 2004, 5, 522–531. [Google Scholar] [CrossRef]
- Guiot, J.; Cambier, M.; Boeckx, A.; Henket, M.; Nivelles, O.; Gester, F.; Louis, E.; Malaise, M.; Dequiedt, F.; Louis, R.; et al. Macrophage-derived exosomes attenuate fibrosis in airway epithelial cells through delivery of antifibrotic miR–142–3p. Thorax 2020, 75, 870–881. [Google Scholar] [CrossRef]
- Wang, L.; Liao, Y.; Yang, R.; Yu, Z.; Zhang, L.; Zhu, Z.; Wu, X.; Shen, J.; Liu, J.; Xu, L.; et al. Sja-miR–71a in Schistosome egg-derived extracellular vesicles suppresses liver fibrosis caused by schistosomiasis via targeting semaphorin 4D. J. Extracell. Vesicles 2020, 9, 1785738. [Google Scholar] [CrossRef]
- Gonzalo-Calvo, D.; Vea, A.; Bär, C.; Fiedler, J.; Couch, L.S.; Brotons, C.; Llorente-Cortes, V.; Thum, T. Circulating non-coding RNAs in biomarker-guided cardiovascular therapy: A novel tool for personalized medicine? Eur. Heart J. 2019, 40, 1643–1650. [Google Scholar] [CrossRef]
- Quan, Y.; Wang, Z.; Gong, L.; Peng, X.; Richard, M.A.; Zhang, J.; Fornage, M.; Alcorn, J.L.; Wang, D. Exosome miR–371b–5p promotes proliferation of lung alveolar progenitor type II cells by using PTEN to orchestrate the PI3K/Akt signaling. Stem Cell Res. Ther. 2017, 8, 138. [Google Scholar] [CrossRef]
- Zhang, D.; Lee, H.; Wang, X.; Groot, M.; Sharma, L.; Cruz, C.S.D.; Jin, Y. A potential role of microvesicle-containing miR–223/142 in lung inflammation. Thorax 2019, 74, 865–874. [Google Scholar] [CrossRef]
- Metlay, J.P.; Waterer, G.W.; Long, A.C.; Anzueto, A.; Brozek, J.; Crothers, K.; Cooley, L.A.; Dean, N.C.; Fine, M.J.; Flanders, S.A.; et al. Diagnosis and Treatment of Adults with Community-acquired Pneumonia. An Official Clinical Practice Guideline of the American Thoracic Society and Infectious Diseases Society of America. Am. J. Respir. Crit. Care Med. 2019, 200, e45–e67. [Google Scholar] [CrossRef]
- Zhang, J.; Mao, F.; Zhao, G.; Wang, H.; Yan, X.; Zhang, Q. Long non-coding RNA SNHG16 promotes lipopolysaccharides-induced acute pneumonia in A549 cells via targeting miR–370–3p/IGF2 axis. Int. Immunopharmacol. 2020, 78, 106065. [Google Scholar] [CrossRef]
- Wei, Z.; Zhang, X.; Yong, T.; Bie, N.; Zhan, G.; Li, X.; Liang, Q.; Li, J.; Yu, J.; Huang, G.; et al. Boosting anti-PD-1 therapy with metformin-loaded macrophage-derived microparticles. Nat. Commun. 2021, 12, 440. [Google Scholar] [CrossRef]
- Crescitelli, R.; Lässer, C.; Szabó, T.G.; Kittel, A.; Eldh, M.; Dianzani, I.; Buzás, E.I.; Lötvall, J. Distinct RNA profiles in subpopulations of extracellular vesicles: Apoptotic bodies, microvesicles and exosomes. J. Extracell Vesicles 2013, 2, 20677. [Google Scholar] [CrossRef] [PubMed]
- Whipple, A.J.; Breton-Provencher, V.; Jacobs, H.N.; Chitta, U.K.; Sur, M.; Sharp, P.A. Imprinted Maternally Expressed microRNAs Antagonize Paternally Driven Gene Programs in Neurons. Mol. Cell 2020, 78, 85–95.e8. [Google Scholar] [CrossRef]
- Galván-Román, J.M.; Lancho-Sánchez, Á.; Luquero-Bueno, S.; Vega-Piris, L.; Curbelo, J.; Manzaneque-Pradales, M.; Gómez, M.; De La Fuente, H.; Ortega-Gómez, M.; Aspa, J. Usefulness of circulating microRNAs miR–146a and miR–16–5p as prognostic biomarkers in community-acquired pneumonia. PLoS ONE 2020, 15, e0240926. [Google Scholar] [CrossRef]
- Wu, X.; Wu, C.; Gu, W.; Ji, H.; Zhu, L. Serum Exosomal MicroRNAs Predict Acute Respiratory Distress Syndrome Events in Patients with Severe Community-Acquired Pneumonia. Biomed Res. Int. 2019, 2019, 3612020. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huang, S.; Feng, C.; Zhai, Y.Z.; Zhou, X.; Li, B.; Wang, L.L.; Chen, W.; Lv, F.Q.; Li, T.S. Identification of miRNA biomarkers of pneumonia using RNA-sequencing and bioinformatics analysis. Exp. Ther. Med. 2017, 13, 1235–1244. [Google Scholar] [CrossRef]
- Scheller, N.; Herold, S.; Kellner, R.; Bertrams, W.; Jung, A.L.; Janga, H.; Greulich, T.; Schulte, L.N.; Vogelmeier, C.F.; Lohmeyer, J.; et al. Proviral MicroRNAs Detected in Extracellular Vesicles from Bronchoalveolar Lavage Fluid of Patients with Influenza Virus-Induced Acute Respiratory Distress Syndrome. J. Infect. Dis. 2019, 219, 540–543. [Google Scholar] [CrossRef] [PubMed]
- Lee, H.; Zhang, D.; Zhu, Z.; Cruz, C.S.D.; Jin, Y. Epithelial cell-derived microvesicles activate macrophages and promote inflammation via microvesicle-containing microRNAs. Sci. Rep. 2016, 6, 35250. [Google Scholar] [CrossRef]
- Li, Y.; Dalli, J.; Chiang, N.; Baron, R.M.; Quintana, C.; Serhan, C.N. Plasticity of leukocytic exudates in resolving acute inflammation is regulated by MicroRNA and proresolving mediators. Immunity 2013, 39, 885–898. [Google Scholar] [CrossRef]
- Feigin, M.; Garvin, T.; Bailey, P.; Waddell, N.; Chang, D.K.; Kelley, D.R.; Shuai, S.; Gallinger, S.; McPherson, J.; Grimmond, S.; et al. Recurrent noncoding regulatory mutations in pancreatic ductal adenocarcinoma. Nat. Genet. 2017, 49, 825–833. [Google Scholar] [CrossRef]
- Zhou, J.; Lin, J.; Zhao, Y.; Sun, X. Deregulated Expression of miR–483–3p Serves as a Diagnostic Biomarker in Severe Pneumonia Children with Respiratory Failure and Its Predictive Value for the Clinical Outcome of Patients. Mol. Biotechnol. 2021, 64, 311–319. [Google Scholar] [CrossRef]
- Guo, J.; Zhang, N.; Liu, G.; Zhang, A.; Liu, X.; Zheng, J. Upregulated expression of long non-coding RNA MEG3 serves as a prognostic biomarker in severe pneumonia children and its regulatory mechanism. Bioengineered 2021, 12, 7120–7131. [Google Scholar] [CrossRef]
- Li, Q.; Wu, T.; Li, S. MiR–181b serves as diagnosis and prognosis biomarker in severe community-acquired pneumonia. Genet. Mol. Biol. 2021, 44, e20200431. [Google Scholar] [CrossRef]
- Valadi, H.; Ekström, K.; Bossios, A.; Sjöstrand, M.; Lee, J.J.; Lötvall, J.O. Exosome-mediated transfer of mRNAs and microRNAs is a novel mechanism of genetic exchange between cells. Nat. Cell Biol. 2007, 9, 654–659. [Google Scholar] [CrossRef]
- Li, Y.; Zheng, Q.; Bao, C.; Li, S.; Guo, W.; Zhao, J.; Chen, D.; Gu, J.; He, X.; Huang, S. Circular RNA is enriched and stable in exosomes: A promising biomarker for cancer diagnosis. Cell Res. 2015, 25, 981–984. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huang, F.; Bai, J.; Zhang, J.; Yang, D.; Fan, H.; Huang, L.; Shi, T.; Lu, G. Identification of potential diagnostic biomarkers for pneumonia caused by adenovirus infection in children by screening serum exosomal microRNAs. Mol. Med. Rep. 2019, 19, 4306–4314. [Google Scholar] [CrossRef] [PubMed]
- Njock, M.S.; Cheng, H.S.; Dang, L.T.; Nazari-Jahantigh, M.; Lau, A.C.; Boudreau, E.; Roufaiel, M.; Cybulsky, M.I.; Schober, A.; Fish, E.J. Endothelial cells suppress monocyte activation through secretion of extracellular vesicles containing antiinflammatory microRNAs. Blood 2015, 125, 3202–3212. [Google Scholar] [CrossRef] [PubMed]
- Zhu, Z.; Zhang, D.; Lee, H.; Menon, A.A.; Wu, J.; Hu, K.; Jin, Y. Macrophage-derived apoptotic bodies promote the proliferation of the recipient cells via shuttling microRNA-221/222. J. Leukoc. Biol. 2017, 101, 1349–1359. [Google Scholar] [CrossRef]
- Xu, H.; Liu, Y.; Cheng, P.; Wang, C.; Liu, Y.; Zhou, W.; Xu, Y.; Ji, G. CircRNA_0000392 promotes colorectal cancer progression through the miR–193a–5p/PIK3R3/AKT axis. J. Exp. Clin. Cancer Res. 2020, 39, 283. [Google Scholar] [CrossRef]
- Wang, J.T.; Wang, Z.H. Role of miR–193a–5p in the proliferation and apoptosis of hepatocellular carcinoma. Eur. Rev. Med. Pharmacol. Sci. 2018, 22, 7233–7239. [Google Scholar]
- Azar, M.; Aghazadeh, H.; Mohammed, H.N.; Sara, M.R.S.; Hosseini, A.; Shomali, N.; Tamjidifar, R.; Tarzi, S.; Mansouri, M.; Serand, S.P.; et al. miR–193a–5p as a promising therapeutic candidate in colorectal cancer by reducing 5-FU and Oxaliplatin chemoresistance by targeting CXCR4. Int. Immunopharmacol. 2021, 92, 107355. [Google Scholar] [CrossRef]
- Duan, B.; Shi, S.; Yue, H.; You, B.; Shan, Y.; Zhu, Z.; Bao, L.; You, Y. Exosomal miR–17–5p promotes angiogenesis in nasopharyngeal carcinoma via targeting BAMBI. J. Cancer. 2019, 10, 6681–6692. [Google Scholar] [CrossRef]
- Song, J.; Liu, Y.; Wang, T.; Li, B.; Zhang, S. MiR–17–5p promotes cellular proliferation and invasiveness by targeting RUNX3 in gastric cancer. Biomed. Pharmacother. 2020, 128, 110246. [Google Scholar] [CrossRef]
- Zhang, Y.; Wang, S.; Lai, Q.; Fang, Y.; Wu, C.; Liu, Y.; Li, Q.; Wang, X.; Gu, C.; Chen, J.; et al. Cancer-associated fibroblasts-derived exosomal miR–17–5p promotes colorectal cancer aggressive phenotype by initiating a RUNX3/MYC/TGF-β1 positive feedback loop. Cancer Lett. 2020, 491, 22–35. [Google Scholar] [CrossRef]
- Murtaza, G.; Khan, A.K.; Rashid, R.; Muneer, S.; Hasan, S.M.F.; Chen, J. FOXO Transcriptional Factors and Long-Term Living. Oxidative Med. Cell Longev. 2017, 2017, 3494289. [Google Scholar] [CrossRef]
- Artham, S.; Gao, F.; Verma, A.; Alwhaibi, A.; Sabbineni, H.; Hafez, S.; Ergul, A.; Somanath, P.R. Endothelial stromelysin1 regulation by the forkhead box-O transcription factors is crucial in the exudative phase of acute lung injury. Pharmacol. Res. 2019, 141, 249–263. [Google Scholar] [CrossRef]
- Bagam, P.; Kaur, G.; Singh, D.P.; Batra, S. In vitro study of the role of FOXO transcription factors in regulating cigarette smoke extract-induced autophagy. Cell Biol. Toxicol. 2021, 37, 531–553. [Google Scholar] [CrossRef]
- Moon, H.W.; Han, H.G.; Jeon, Y.J. Protein Quality Control in the Endoplasmic Reticulum and Cancer. Int. J. Mol. Sci. 2018, 19, 3020. [Google Scholar] [CrossRef]
- López-Otín, C.; Blasco, M.A.; Partridge, L.; Serrano, M.; Kroemer, G. The hallmarks of aging. Cell 2013, 153, 1194–1217. [Google Scholar] [CrossRef]
- Wang, M.; Kaufman, R.J. Protein misfolding in the endoplasmic reticulum as a conduit to human disease. Nature 2016, 529, 326–335. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Control Group (n = 13) | Pneumonia Group (n = 61) | p Value | |
|---|---|---|---|
| Age, years, mean ± SD | 50.38 ± 13.16 | 57.70 ± 18.84 | 0.107 |
| Sex | 1.000 | ||
| Male | 11 | 49 | |
| Female | 2 | 12 | |
| Any comorbidity, n (%) | 12 (92.3) | 53 (0.87) | 0.940 |
| Hypertension | 9 (69.2) | 26 (42.6) | 0.081 |
| Diabetes | 3 (23.1) | 14 (23.0) | 1.000 |
| Malignancy | 0 (0.0) | 5 (8.2) | 0.579 |
| Chronic liver disease | 2 (15.4) | 14 (23) | 0.818 |
| Cardiovascular disease | 3 (23.1) | 10 (10.6) | 0.862 |
| Renal disease | 1 (7.7) | 9 (14.8) | 0.819 |
| Craniocerebral trauma | 9 (69.2) | 18 (29.5) | 0.017 |
| hsCRP, mean (range), (mg/L) | 52.4 (0.9–151.7) | 76.5 (1.7–328.7) | 0.236 |
| PCT, mean (range), (ng/mL) | 1.1 (0.1–4.1) | 4.9 (0.02–45.0) | 0.004 |
| Index | Cutoff Value | AUC | 95% CI | Specificity (%) | Sensitivity (%) | p |
|---|---|---|---|---|---|---|
| miR–17–5p | 2.32 | 0.753 | 0.639–0.846 | 84.62 | 59.02 | 0.0002 |
| miR–193a–5p | 2.40 | 0.692 | 0.574–0.794 | 100 | 50.82 | 0.0028 |
| miR–17–5p + miR–193a–5p | − | 0.748 | 0.633–0.842 | 92.31 | 57.38 | 0.0001 |
| hsCRP | 28.80 mg/L | 0.651 | 0.531–0.758 | 61.54 | 80.33 | 0.1420 |
| PCT | 0.89 ng/mL | 0.685 | 0.566–0.788 | 69.23 | 62.30 | 0.0254 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sun, Y.; Xian, Y.; Duan, Z.; Wan, Z.; Li, J.; Liao, Y.; Bi, X.; Wu, Z.; Wang, L.; Zhang, K. Diagnostic Potential of microRNAs in Extracellular Vesicles Derived from Bronchoalveolar Lavage Fluid for Pneumonia—A Preliminary Report. Cells 2022, 11, 2961. https://doi.org/10.3390/cells11192961
Sun Y, Xian Y, Duan Z, Wan Z, Li J, Liao Y, Bi X, Wu Z, Wang L, Zhang K. Diagnostic Potential of microRNAs in Extracellular Vesicles Derived from Bronchoalveolar Lavage Fluid for Pneumonia—A Preliminary Report. Cells. 2022; 11(19):2961. https://doi.org/10.3390/cells11192961
Chicago/Turabian StyleSun, Yinfang, Ying Xian, Zhiqin Duan, Zhiping Wan, Jianwei Li, Yao Liao, Xiaogang Bi, Zhongdao Wu, Lifu Wang, and Kouxing Zhang. 2022. "Diagnostic Potential of microRNAs in Extracellular Vesicles Derived from Bronchoalveolar Lavage Fluid for Pneumonia—A Preliminary Report" Cells 11, no. 19: 2961. https://doi.org/10.3390/cells11192961