
Low-Ammonium Environment Increases the Nutrient Exchange between Diatom–Diazotroph Association Cells and Facilitates Photosynthesis and N2 Fixation—a Mechanistic Modeling Analysis


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results and Discussion
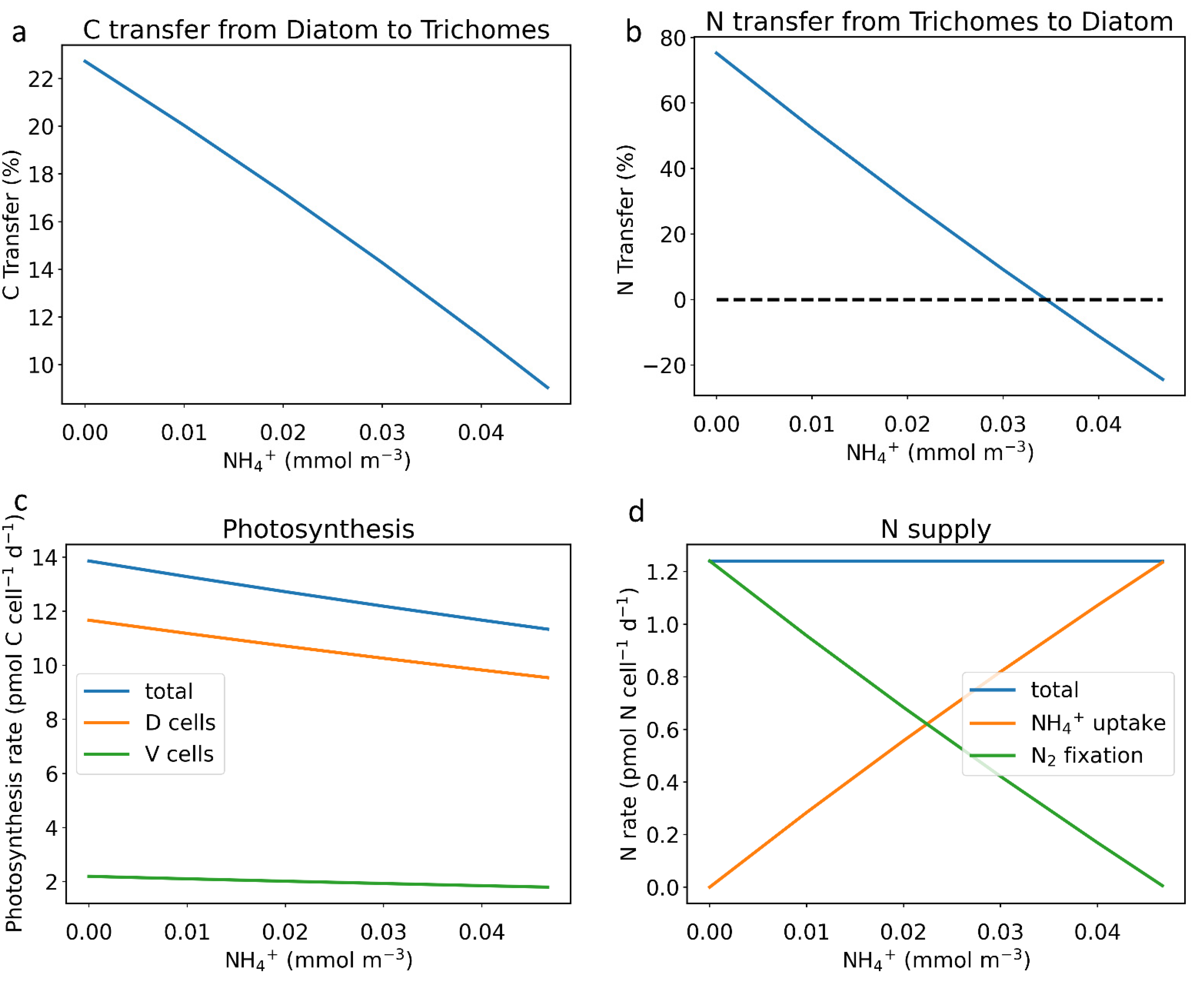
2.1. NH4+ Uptake Influences Nutrient Exchange and Metabolism
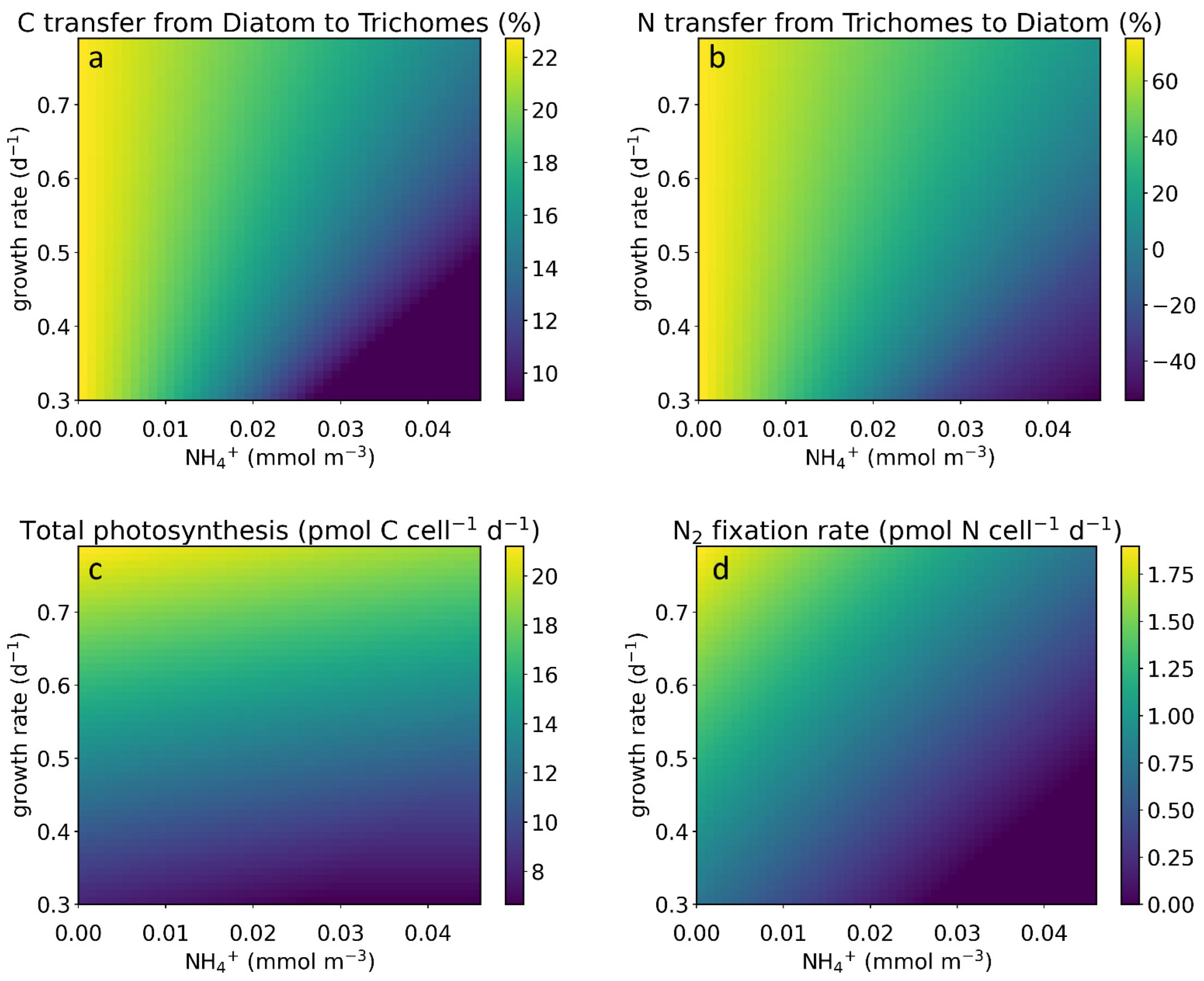
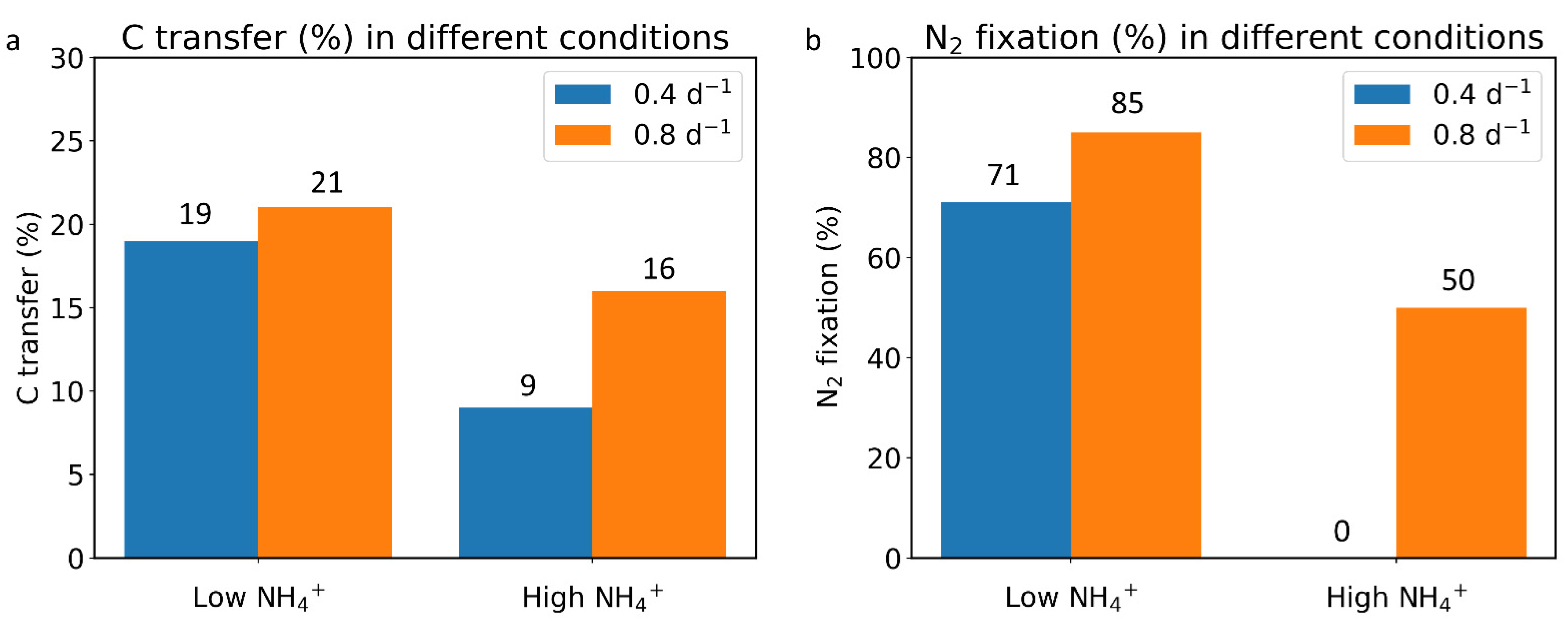
2.2. Effect of Growth Rate
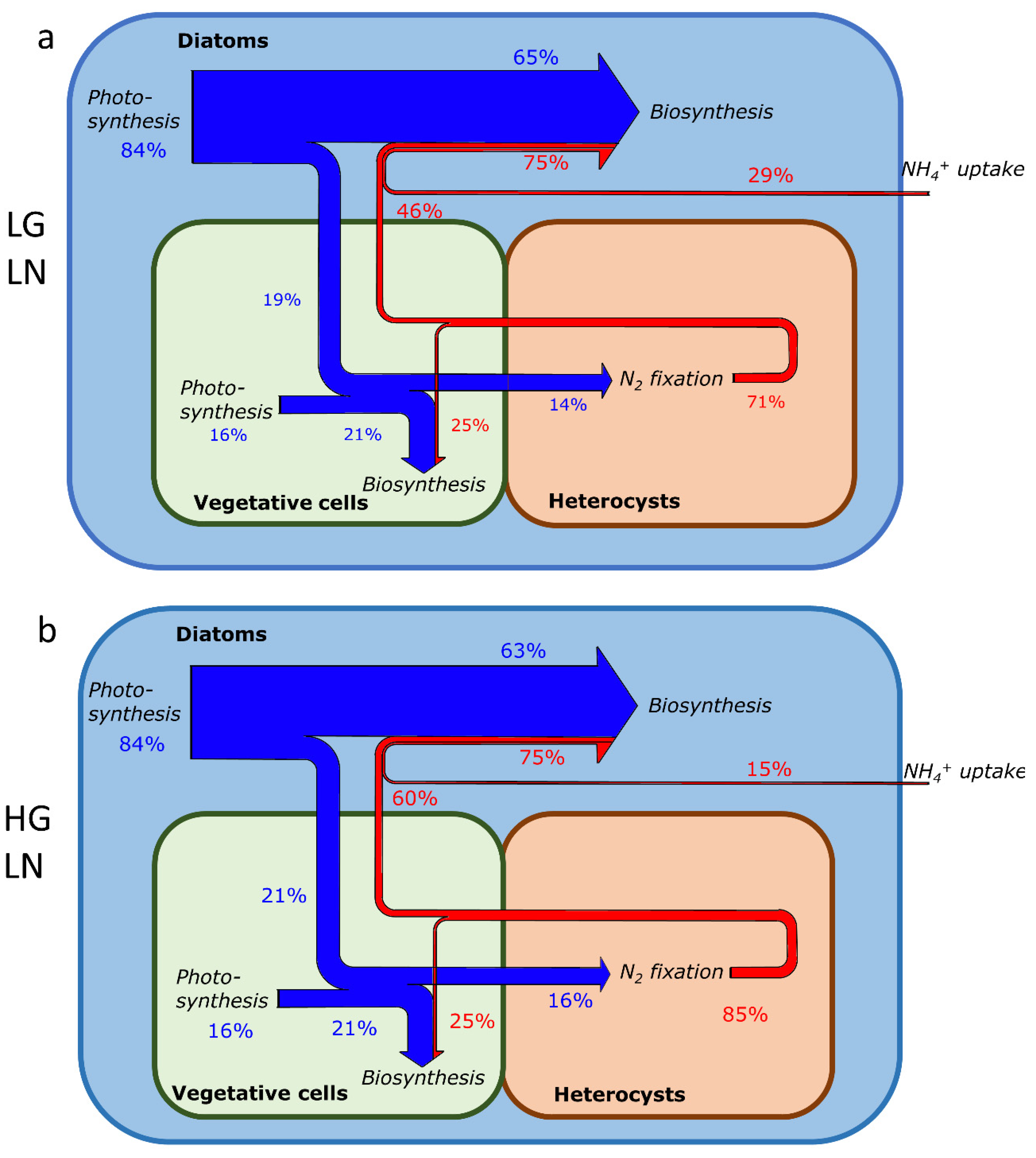
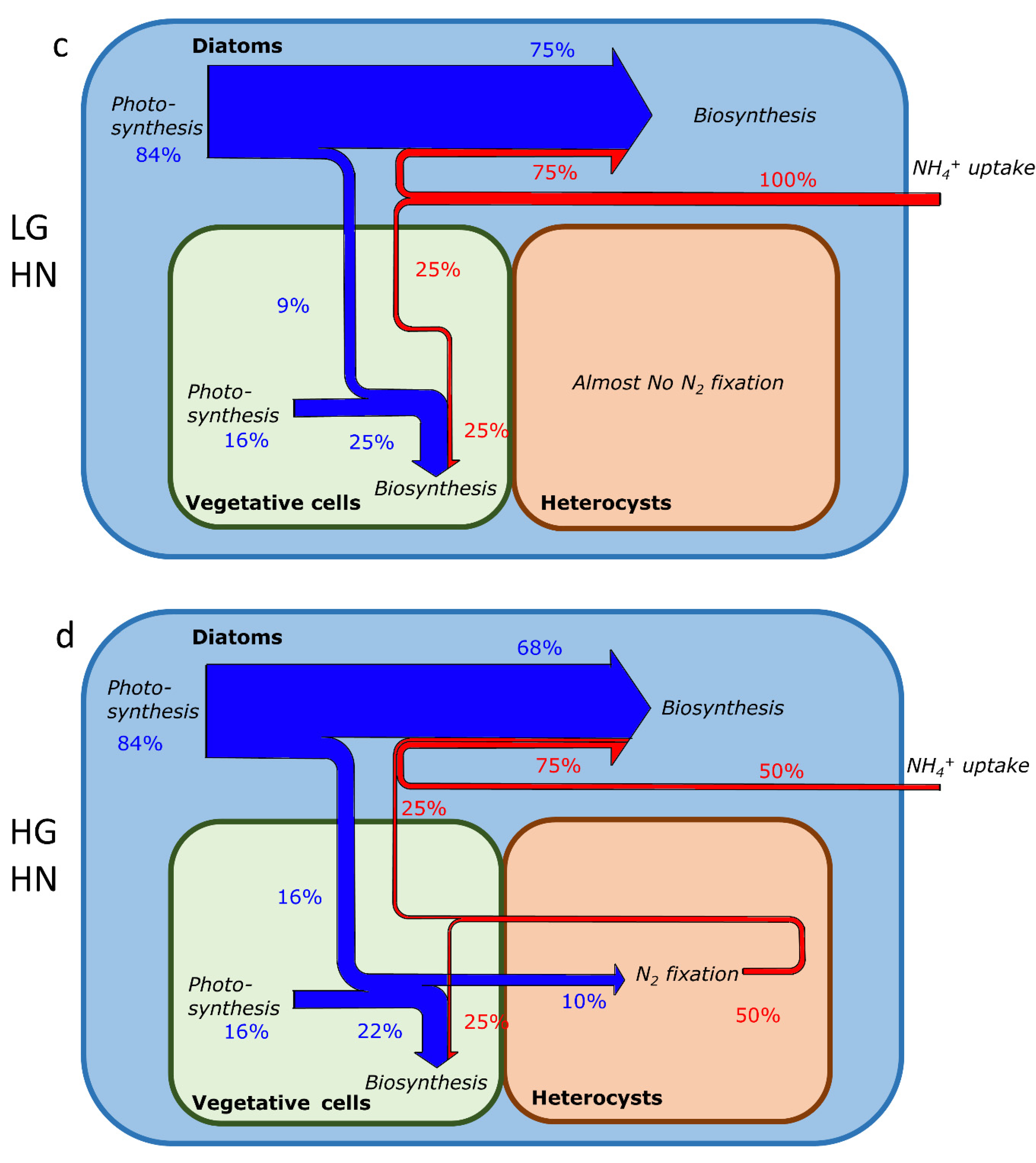
2.3. Element Fate and Flux in Different NH4+ Concentrations and Growth Rates
2.4. Effect of Other Factors: Trichome Number and Light Intensity
2.4.1. Trichome Number
2.4.2. Light Intensity
2.5. Comparison to Previous Studies and Implications for Future Work
3. Conclusions
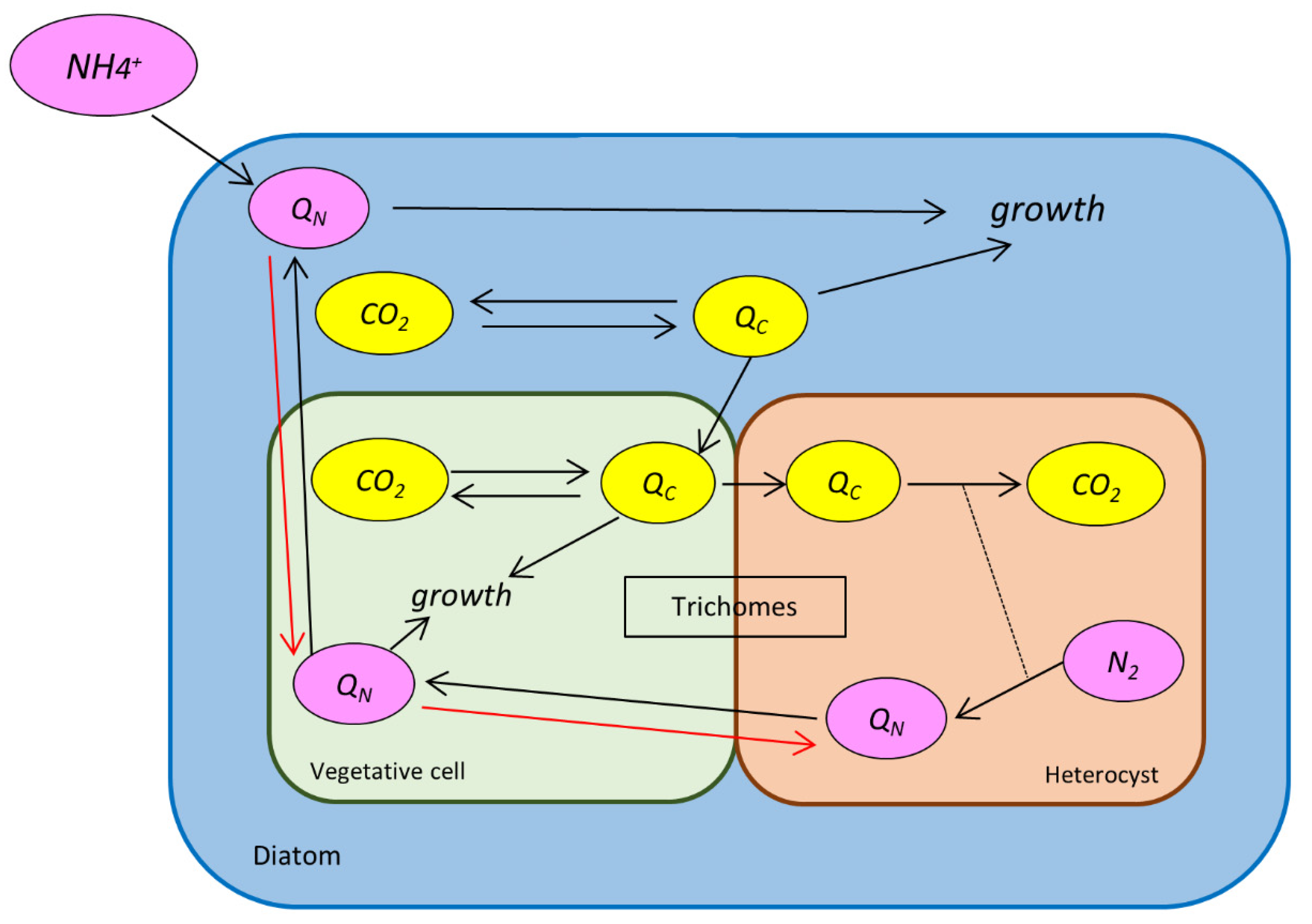
4. Materials and Methods
4.1. C Balance
4.2. N Balance
4.3. Values and Calculations
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Villareal, T.A. Widespread Occurrence of the Hemiaulus-cyanobacterial Symbiosis in the Southwest North Atlantic Ocean. Bull. Mar. Sci. 1994, 54, 7. [Google Scholar]
- Villareal, T.A.; Brown, C.G.; Brzezinski, M.A.; Krause, J.W.; Wilson, C. Summer Diatom Blooms in the North Pacific Subtropical Gyre: 2008–2009. PLoS ONE 2012, 7, 2008–2009. [Google Scholar] [CrossRef] [PubMed]
- Villareal, T.A. Marine Nitrogen-Fixing Diatom-Cyanobacteria Symbioses. In Marine Pelagic Cyanobacteria: Trichodesmium and Other Diazotrophs; Carpenter, E.J., Capone, D.G., Rueter, J.G., Eds.; Springer: Dordrecht, The Netherlands, 1992; pp. 163–175. ISBN 978-94-015-7977-3. [Google Scholar]
- Foster, R.A.; Kuypers, M.M.M.; Vagner, T.; Paerl, R.W.; Musat, N.; Zehr, J.P. Nitrogen Fixation and Transfer in Open Ocean Diatom-Cyanobacterial Symbioses. ISME J. 2011, 5, 1484–1493. [Google Scholar] [CrossRef] [PubMed]
- Tuo, S.H.; Lee Chen, Y.L.; Chen, H.Y. Low Nitrate Availability Promotes Diatom Diazotroph Associations in the Marginal Seas of the Western Pacific. Aquat. Microb. Ecol. 2014, 73, 135–150. [Google Scholar] [CrossRef]
- Inomura, K.; Follett, C.L.; Masuda, T.; Eichner, M.; Prášil, O.; Deutsch, C. Carbon Transfer from the Host Diatom Enables Fast Growth and High Rate of N2 Fixation by Symbiotic Heterocystous Cyanobacteria. Plants 2020, 9, 192. [Google Scholar] [CrossRef]
- Subramaniam, A.; Yager, P.L.; Carpenter, E.J.; Mahaffey, C.; Björkman, K.; Cooley, S.; Kustka, A.B.; Montoya, J.P.; Sañudo-Wilhelmy, S.A.; Shipe, R.; et al. Amazon River Enhances Diazotrophy and Carbon Sequestration in the Tropical North Atlantic Ocean. Proc. Natl. Acad. Sci. USA 2008, 105, 10460–10465. [Google Scholar] [CrossRef]
- Carpenter, E.J. Marine Cyanobacterial Symbioses. Biol. Environ. 2002, 102, 15–18. [Google Scholar] [CrossRef]
- Sundström, B.G. Observations on Rhizosolenia Clevei Ostenfeld (Bacillariophyceae) and Richelia intracellularis Schmidt (Cyanophyceae). Bot. Mar. 1984, 27, 345–356. [Google Scholar] [CrossRef]
- Foster, R.A.; Zehr, J.P. Characterization of Diatom-Cyanobacteria Symbioses on the Basis of NifH, HetR and 16S RRNA Sequences. Environ. Microbiol. 2006, 8, 1913–1925. [Google Scholar] [CrossRef]
- Foster, R.A.; Tienken, D.; Littmann, S.; Whitehouse, M.J.; Kuypers, M.M.M.; White, A.E. The Rate and Fate of N2 and C Fixation by Marine Diatom-Diazotroph Symbioses. ISME J. 2022, 16, 477–487. [Google Scholar] [CrossRef]
- Foster, R.A.; Subramaniam, A.; Zehr, J.P. Distribution and Activity of Diazotrophs in the Eastern Equatorial Atlantic. Environ. Microbiol. 2009, 11, 741–750. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Caputo, A.; Stenegren, M.; Pernice, M.C.; Foster, R.A. A Short Comparison of Two Marine Planktonic Diazotrophic Symbioses Highlights an Un-Quantified Disparity. Front. Mar. Sci. 2018, 5, 2. [Google Scholar] [CrossRef]
- Hilton, J.A.; Foster, R.A.; James Tripp, H.; Carter, B.J.; Zehr, J.P.; Villareal, T.A. Genomic Deletions Disrupt Nitrogen Metabolism Pathways of a Cyanobacterial Diatom Symbiont. Nat. Commun. 2013, 4, 1767. [Google Scholar] [CrossRef] [PubMed]
- Martínez-Pérez, C.; Mohr, W.; Löscher, C.R.; Dekaezemacker, J.; Littmann, S.; Yilmaz, P.; Lehnen, N.; Fuchs, B.M.; Lavik, G.; Schmitz, R.A.; et al. The Small Unicellular Diazotrophic Symbiont, UCYN-A, Is a Key Player in the Marine Nitrogen Cycle. Nat. Microbiol. 2016, 1, 16163. [Google Scholar] [CrossRef]
- Sessitsch, A.; Howieson, J.G.; Perret, X.; Antoun, H.; Martínez-Romero, E. Advances in Rhizobium Research. CRC. Crit. Rev. Plant. Sci. 2002, 21, 323–378. [Google Scholar] [CrossRef]
- Inomura, K.; Deutsch, C.; Masuda, T.; Prášil, O.; Follows, M.J. Quantitative Models of Nitrogen-Fixing Organisms. Comput. Struct. Biotechnol. J. 2020, 18, 3905–3924. [Google Scholar] [CrossRef]
- Peters, G.A.; Meeks, J.C. The Azolla-Anabaena Symbiosis: Basic Biology. Annu. Rev. Plant. Physiol. Plant. Mol. Biol. 1989, 40, 193–210. [Google Scholar] [CrossRef]
- Zehr, J.P. How Single Cells Work Together. Science 2015, 349, 1163–1164. [Google Scholar] [CrossRef]
- Villareal, T.A.; Adornato, L.; Wilson, C.; Schoenbaechler, C.A. Summer Blooms of Diatom-Diazotroph Assemblages and Surface Chlorophyll in the North Pacific Gyre: A Disconnect. J. Geophys. Res. Ocean. 2011, 116, C03001. [Google Scholar] [CrossRef]
- Kitajima, S.; Furuya, K.; Hashihama, F.; Takeda, S.; Kanda, J. Latitudinal Distribution of Diazotrophs and Their Nitrogen Fixation in the Tropical and Subtropical Western North Pacific. Limnol. Oceanogr. 2009, 54, 537–547. [Google Scholar] [CrossRef]
- Dore, J.E.; Letelier, R.M.; Church, M.J.; Lukas, R.; Karl, D.M. Summer Phytoplankton Blooms in the Oligotrophic North Pacific Subtropical Gyre: Historical Perspective and Recent Observations. Prog. Oceanogr. 2008, 76, 2–38. [Google Scholar] [CrossRef]
- Carpenter, E.J.; Montoya, J.P.; Burns, J.; Mulholland, M.R.; Subramaniam, A.; Capone, D.G. Extensive Bloom of a N2-Fixing Diatom/Cyanobacterial Association in the Tropical Atlantic Ocean. Mar. Ecol. Prog. Ser. 1999, 185, 273–283. [Google Scholar] [CrossRef]
- Devassy, R.P.; El-Sherbiny, M.M.; Al-Sofyani, A.A.; Crosby, M.P.; Al-Aidaroos, A.M. Seasonality and Latitudinal Variability in the Diatom-Cyanobacteria Symbiotic Relationships in the Coastal Waters of the Red Sea, Saudi Arabia. Symbiosis 2019, 78, 215–227. [Google Scholar] [CrossRef]
- Stukel, M.R.; Coles, V.J.; Brooks, M.T.; Hood, R.R. Top-down, Bottom-up and Physical Controls on Diatom-Diazotroph Assemblage Growth in the Amazon River Plume. Biogeosciences 2014, 1, 3259–3278. [Google Scholar] [CrossRef]
- Monteiro, F.M.; Follows, M.J.; Dutkiewicz, S. Distribution of Diverse Nitrogen Fixers in the Global Ocean. Glob. Biogeochem. Cycles 2010, 24, 1–16. [Google Scholar] [CrossRef]
- Monteiro, F.M.; Dutkiewicz, S.; Follows, M.J. Biogeographical Controls on the Marine Nitrogen Fixers. Glob. Biogeochem. Cycles 2011, 25, 1–8. [Google Scholar] [CrossRef]
- Sunda, W.G.; Shertzer, K.W.; Hardison, D.R. Ammonium Uptake and Growth Models in Marine Diatoms: Monod and Droop Revisited. Mar. Ecol. Prog. Ser. 2009, 386, 29–41. [Google Scholar] [CrossRef]
- Goldman, J.C.; McCarthy, J.J. Steady State Growth and Ammonium Uptake of a Fast-growing Marine Diatom. Limnol. Oceanogr. 1978, 23, 695–703. [Google Scholar] [CrossRef]
- Saros, J.E.; Fritz, S.C. Changes in the Growth Rates of Saline-Lake Diatoms in Response to Variation in Salinity, Brine Type and Nitrogen Form. J. Plankton Res. 2000, 22, 1071–1083. [Google Scholar] [CrossRef]
- Popovich, C.A.; Spetter, C.V.; Marcovecchio, J.E.; Freije, R.H. Dissolved Nutrient Availability during Winter Diatom Bloom in a Turbid and Shallow Estuary (Bahía Blanca, Argentina). J. Coast. Res. 2008, 24, 95–102. [Google Scholar] [CrossRef]
- Dekaezemacker, J.; Bonnet, S. Sensitivity of N2 Fixation to Combined Nitrogen Forms (NO3− and NH4+) in Two Strains of the Marine Diazotroph Crocosphaera Watsonii (Cyanobacteria). Mar. Ecol. Prog. Ser. 2011, 438, 33–46. [Google Scholar] [CrossRef] [Green Version]
- Boatman, T.G.; Davey, P.A.; Lawson, T.; Geider, R.J. The Physiological Cost of Diazotrophy for Trichodesmium erythraeum IMS101. PLoS ONE 2018, 13, e0195638. [Google Scholar] [CrossRef]
- Pyle, A.E.; Johnson, A.M.; Villareal, T.A. Isolation, Growth, and Nitrogen Fixation Rates of the Hemiaulus-Richelia (Diatom-Cyanobacterium) Symbiosis in Culture. PeerJ 2020, 8, e10115. [Google Scholar] [CrossRef] [PubMed]
- Vaillancourt, R.D.; Marra, J.; Seki, M.P.; Parsons, M.L.; Bidigare, R.R. Impact of a Cyclonic Eddy on Phytoplankton Community Structure and Photosynthetic Competency in the Subtropical North Pacific Ocean. Deep. Res. Part. I Oceanogr. Res. Pap. 2003, 50, 829–847. [Google Scholar] [CrossRef]
- Zeev, E.B.; Yogev, T.; Man-Aharonovich, D.; Kress, N.; Herut, B.; Béjà, O.; Berman-Frank, I. Seasonal Dynamics of the Endosymbiotic, Nitrogen-Fixing Cyanobacterium Richelia intracellularis in the Eastern Mediterranean Sea. ISME J. 2008, 2, 911–923. [Google Scholar] [CrossRef] [PubMed]
- Sunda, W.G.; Hardison, D.R. Ammonium Uptake and Growth Limitation in Marine Phytoplankton. Limnol. Oceanogr. 2007, 52, 2496–2506. [Google Scholar] [CrossRef]
- Follett, C.L.; Dutkiewicz, S.; Karl, D.M.; Inomura, K.; Follows, M.J. Seasonal Resource Conditions Favor a Summertime Increase in North Pacific Diatom-Diazotroph Associations. ISME J. 2018, 12, 1543–1557. [Google Scholar] [CrossRef]
- Inomura, K.; Bragg, J.; Riemann, L.; Follows, M.J. A Quantitative Model of Nitrogen Fixation in the Presence of Ammonium. PLoS ONE 2018, 13, e0208282. [Google Scholar] [CrossRef]
- Pahlow, M.; Oschlies, A. Chain Model of Phytoplankton P, N and Light Colimitation. Mar. Ecol. Prog. Ser. 2009, 376, 69–83. [Google Scholar] [CrossRef]
- Pahlow, M.; Dietze, H.; Oschlies, A. Optimality-Based Model of Phytoplankton Growth and Diazotrophy. Mar. Ecol. Prog. Ser. 2013, 489, 1–16. [Google Scholar] [CrossRef]
- Sarkar, D.; Landa, M.; Bandyopadhyay, A.; Pakrasi, H.B.; Zehr, J.P.; Maranas, C.D. Elucidation of Trophic Interactions in an Unusual Single-Cell Nitrogen-Fixing Symbiosis Using Metabolic Modeling. PLoS Comput. Biol. 2021, 17, e1008983. [Google Scholar] [CrossRef]
- Malatinszky, D.; Steuer, R.; Jones, P.R. A Comprehensively Curated Genome-Scale Two-Cell Model for the Heterocystous Cyanobacterium Anabaena Sp. PCC 7120. Plant. Physiol. 2017, 173, 509–523. [Google Scholar] [CrossRef] [PubMed]
- Gardner, J.J.; Boyle, N.R. The Use of Genome-Scale Metabolic Network Reconstruction to Predict Fluxes and Equilibrium Composition of N-Fixing versus C-Fixing Cells in a Diazotrophic Cyanobacterium, Trichodesmium Erythraeum. BMC Syst. Biol. 2017, 11, 4. [Google Scholar] [CrossRef] [PubMed]
- Villareal, T.A. Laboratory Culture and Preliminary Characterization of the Nitrogen-Fixing Rhizosolenia-Richelia Symbiosis. Mar. Ecol. 1990, 11, 117–132. [Google Scholar] [CrossRef]
- Mulholland, M.R.; Bernhardt, P.W. The Effect of Growth Rate, Phosphorus Concentration, and Temperature on N2 Fixation, Carbon Fixation, and Nitrogen Release in Continuous Cultures of Trichodesmium IMS101. Limnol. Oceanogr. 2005, 50, 839–849. [Google Scholar] [CrossRef]
- Hutchins, D.A.; Fu, F.X.; Zhang, Y.; Warner, M.E.; Feng, Y.; Portune, K.; Bernhardt, P.W.; Mulholland, M.R. CO2 Control of Trichodesmium N2 Fixation, Photosynthesis, Growth Rates, and Elemental Ratios: Implications for Past, Present, and Future Ocean Biogeochemistry. Limnol. Oceanogr. 2007, 52, 1293–1304. [Google Scholar] [CrossRef]
- Fu, F.X.; Mulholland, M.R.; Garcia, N.S.; Beck, A.; Bernhardt, P.W.; Warner, M.E.; Sañudo-Wilhelmy, S.A.; Hutchins, D.A. Interactions between Changing pCO2, N2 Fixation, and Fe Limitation in the Marine Unicellular Cyanobacterium Crocosphaera. Limnol. Oceanogr. 2008, 53, 2472–2484. [Google Scholar] [CrossRef]
- Bale, N.J.; Villareal, T.A.; Hopmans, E.C.; Brussaard, C.P.D.; Besseling, M.; Dorhout, D.; Sinninghe Damsté, J.S.; Schouten, S. C5 Glycolipids of Heterocystous Cyanobacteria Track Symbiont Abundance in the Diatom Hemiaulus Hauckii across the Tropical North Atlantic. Biogeosciences 2018, 15, 1229–1241. [Google Scholar] [CrossRef]
- Capone, D.G.; Bronk, D.A.; Mulholland, M.R.; Carpenter, E.J. Nitrogen in the Marine Environment, 2nd ed.; Elsevier: Amsterdam, The Netherlands, 2008; ISBN 978-0-12-372522-6. [Google Scholar]
- Smith, E. Limiting Factors in Photosynthesis: Light and Carbon Dioxide. J. Gen. Physiol. 1938, 22, 21–35. [Google Scholar] [CrossRef]
- Ryther, J.H. Photosynthesis in the Ocean as a Function of Light Intensity. Limnol. Oceanogr. 1956, 1, 61–70. [Google Scholar] [CrossRef]
- Falciatore, A. Exploring the Molecular Basis of Responses to Light in Marine Diatoms. J. Exp. Bot. 2012, 63, 1575–1591. [Google Scholar] [CrossRef] [Green Version]
- Ingebrigtsen, R.A.; Hansen, E.; Andersen, J.H.; Eilertsen, H.C. Light and Temperature Effects on Bioactivity in Diatoms. J. Appl. Phycol. 2016, 28, 939–950. [Google Scholar] [CrossRef] [PubMed]
- Grossman, A.R.; Schaefer, M.R.; Chiang, G.G.; Collier, J.L. The Responses of Cyanobacteria to Environmental Conditions: Light and Nutrients. In The Molecular Biology of Cyanobacteria; Bryant, D.A., Ed.; Springer: Dordrecht, The Netherlands, 1994; pp. 641–675. ISBN 978-94-011-0227-8. [Google Scholar]
- Papers, J.B.C.; Grossman, A.R.; Bhaya, D. Tracking the Light Environment by Cyanobacteria and the Dynamic Nature of Light Harvesting. J. Biol. Chem. 2001, 276, 11449–11452. [Google Scholar] [CrossRef]
- Benavides, M.; Bonnet, S.; Le Moigne, F.A.C.; Armin, G.; Inomura, K.; Hallstrøm, S.; Riemann, L.; Berman-Frank, I.; Poletti, E.; Garel, M.; et al. Sinking Trichodesmium Fixes Nitrogen in the Dark Ocean. ISME J. 2022, 16, 2398–2405. [Google Scholar] [CrossRef]
- Herrero, A.; Flores, E. Genetic Responses to Carbon and Nitrogen Availability in Anabaena. Environ. Microbiol. 2019, 21, 1–17. [Google Scholar] [CrossRef]
- Zehr, J.P.; Ward, B.B. Nitrogen Cycling in the Ocean: New Perspectives on Processes and Paradigms. Appl. Environ. Microbiol. 2002, 68, 1015–1024. [Google Scholar] [CrossRef]
- Capone, D.G.; Zehr, J.P.; Paerl, H.W.; Bergman, B.; Carpenter, E.J. Trichodesmium, a Globally Significant Marine Cyanobacterium. Science 1997, 276, 1221–1229. [Google Scholar] [CrossRef]
- Gruber, N.; Galloway, J.N. An Earth-System Perspective of the Global Nitrogen Cycle. Nature 2008, 451, 10–13. [Google Scholar] [CrossRef]
- Letscher, R.T.; Villareal, T.A. Evaluation of the Seasonal Formation of Subsurface Negative Preformed Nitrate Anomalies in the Subtropical North Pacific and North Atlantic. Biogeosciences 2018, 15, 6461–6480. [Google Scholar] [CrossRef]
- Nieves-morión, M.; Flores, E.; Foster, R.A. Minireview Predicting Substrate Exchange in Marine Diatom-Heterocystous Cyanobacteria Symbioses. Environ. Microbiol. 2020, 22, 2027–2052. [Google Scholar] [CrossRef]
- Foster, R.A.; Goebel, N.L.; Zehr, J.P.; Foster, R.A.; Goebel, N.L.; Zehr, J.P. Isolation of Calothrix Rhizosoleniae (Cyanobacteria) Strain Sc01 From Chaetoceros (Bacillariophyta) Spp. Diatoms of the Subtropical North Pacific Ocean. Phycol. Soc. Am. 2010, 1037, 1028–1037. [Google Scholar] [CrossRef]
- Foster, R.A.; Zehr, J.P. Diversity, Genomics, and Distribution of Phytoplankton-Cyanobacterium Single-Cell Symbiotic Associations. Annu. Rev. Microbiol. 2019, 73, 435–456. [Google Scholar] [CrossRef] [PubMed]
- Harke, M.J.; Frischkorn, K.R.; Haley, S.T.; Aylward, F.O.; Zehr, J.P.; Dyhrman, S.T. Periodic and Coordinated Gene Expression between a Diazotroph and Its Diatom Host. ISME J. 2019, 13, 118–131. [Google Scholar] [CrossRef] [PubMed]
- Menden-Deuer, S.; Lessard, E.J. Carbon to Volume Relationships for Dinoflagellates, Diatoms, and Other Protist Plankton. Limnol. Oceanogr. 2000, 45, 569–579. [Google Scholar] [CrossRef]
- Rittmann, B.E.; McCarty, P.L. Environmental Biotechnology: Principles and Applications; previous paper Inomura 2020; McGraw-Hill Education: New York, NY, USA, 2001; ISBN 1260440591. [Google Scholar]
- Villareal, T. Nitrogen-Fixation by the Cyanobacterial Symbiont of the Diatom Genus Hemiaulus. Mar. Ecol. Prog. Ser. 1991, 76, 201–204. [Google Scholar] [CrossRef]
- Redfield, A.C. The Biological Control of Chemical Factors in the Environment. Am. Sci. 1958, 46, 205–221. [Google Scholar]
- Sohm, J.A.; Edwards, B.R.; Wilson, B.G.; Webb, E.A. Constitutive Extracellular Polysaccharide (EPS) Production by Specific Isolates of Crocosphaera watsonii. Front. Microbiol. 2011, 2, 229. [Google Scholar] [CrossRef]
- Großkopf, T.; Laroche, J. Direct and Indirect Costs of Dinitrogen Fixation in Crocosphaera Watsonii WH8501 and Possible Implications for the Nitrogen Cycle. Front. Microbiol. 2012, 3, 236. [Google Scholar] [CrossRef]
- Paulot, F.; Jacob, D.J.; Johnson, M.T.; Bell, T.G.; Baker, A.R.; Keene, W.C.; Lima, I.D.; Doney, S.C.; Stock, C.A. Global Oceanic Emission of Ammonia: Constraints from Seawater and Atmospheric Observations. Glob. Biogeochem. Cycles 2015, 29, 1165–1178. [Google Scholar] [CrossRef]
- Inomura, K.; Wilson, S.T.; Deutsch, C. Mechanistic Model for the Coexistence of Nitrogen Fixation and Photosynthesis in Marine Trichodesmium. mSystems 2019, 4, e00210-19. [Google Scholar] [CrossRef] [Green Version]






Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gao, M.; Armin, G.; Inomura, K. Low-Ammonium Environment Increases the Nutrient Exchange between Diatom–Diazotroph Association Cells and Facilitates Photosynthesis and N2 Fixation—a Mechanistic Modeling Analysis. Cells 2022, 11, 2911. https://doi.org/10.3390/cells11182911
Gao M, Armin G, Inomura K. Low-Ammonium Environment Increases the Nutrient Exchange between Diatom–Diazotroph Association Cells and Facilitates Photosynthesis and N2 Fixation—a Mechanistic Modeling Analysis. Cells. 2022; 11(18):2911. https://doi.org/10.3390/cells11182911
Chicago/Turabian StyleGao, Meng, Gabrielle Armin, and Keisuke Inomura. 2022. "Low-Ammonium Environment Increases the Nutrient Exchange between Diatom–Diazotroph Association Cells and Facilitates Photosynthesis and N2 Fixation—a Mechanistic Modeling Analysis" Cells 11, no. 18: 2911. https://doi.org/10.3390/cells11182911