
Small Extracellular Vesicles and Their Involvement in Cancer Resistance: An Up-to-Date Review



Abstract
:1. Prima Facie of Extracellular Vesicles (EVs) in Cancer
2. Prima Facie of Extracellular Vesicles (EVs) in Chemoresistance
3. Exosomes as Chemoresistance Mediators
3.1. Resistance to Platinum-Based Therapy
3.2. Resistance to Alkylating Agents
3.3. Resistance to Antimetabolite Agents
4. Exosomes as Chemosensitivity Mediators
5. Conclusions and Perspectives
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Yates, A.G.; Pink, R.C.; Erdbrügger, U.; Siljander, P.R.; Dellar, E.R.; Pantazi, P.; Akbar, N.; Cooke, W.R.; Vatish, M.; Dias-Neto, E.; et al. In sickness and in health: The functional role of extracellular vesicles in physiology and pathology in vivo: Part I: Health and Normal Physiology: Part I: Health and Normal Physiology. J. Extracell. Vesicles 2022, 11, e12151. [Google Scholar] [CrossRef] [PubMed]
- Yates, A.G.; Pink, R.C.; Erdbrügger, U.; Siljander, P.R.; Dellar, E.R.; Pantazi, P.; Akbar, N.; Cooke, W.R.; Vatish, M.; Dias-Neto, E.; et al. In sickness and in health: The functional role of extracellular vesicles in physiology and pathology in vivo: Part II: Pathology: Part II: Pathology. J. Extracell. Vesicles 2022, 11, e12190. [Google Scholar] [CrossRef] [PubMed]
- Chargaff, E.; West, R. The biological significance of the thromboplastic protein of blood. J. Biol. Chem. 1946, 166, 189–197. [Google Scholar] [CrossRef]
- Couch, Y.; Buzàs, E.I.; Di Vizio, D.; Gho, Y.S.; Harrison, P.; Hill, A.F.; Lötvall, J.; Raposo, G.; Stahl, P.D.; Théry, C.; et al. A brief history of nearly EV-erything—The rise and rise of extracellular vesicles. J. Extracell. Vesicles 2021, 10, e12144. [Google Scholar] [CrossRef] [PubMed]
- Chang, W.H.; Cerione, R.A.; Antonyak, M.A. Extracellular Vesicles and Their Roles in Cancer Progression. Methods Mol. Biol. 2021, 2174, 143–170. [Google Scholar] [CrossRef]
- Console, L.; Scalise, M. Extracellular Vesicles and Cell Pathways Involved in Cancer Chemoresistance. Life 2022, 12, 618. [Google Scholar] [CrossRef]
- Zhou, E.; Li, Y.; Wu, F.; Guo, M.; Xu, J.; Wang, S.; Tan, Q.; Ma, P.; Song, S.; Jin, Y. Circulating extracellular vesicles are effective biomarkers for predicting response to cancer therapy. EBioMedicine 2021, 67, 103365. [Google Scholar] [CrossRef]
- Bao, Q.; Huang, Q.; Chen, Y.; Wang, Q.; Sang, R.; Wang, L.; Xie, Y.; Chen, W. Tumor-Derived Extracellular Vesicles Regulate Cancer Progression in the Tumor Microenvironment. Front. Mol. Biosci. 2021, 8, 796385. [Google Scholar] [CrossRef]
- Vu, L.T.; Peng, B.; Zhang, D.X.; Ma, V.; Mathey-Andrews, C.A.; Lam, C.K.; Kiomourtzis, T.; Jin, J.; McReynolds, L.; Huang, L.; et al. Tumor-secreted extracellular vesicles promote the activation of cancer-associated fibroblasts via the transfer of microRNA-125b. J. Extracell. Vesicles 2019, 8, 1599680. [Google Scholar] [CrossRef]
- Khan, M.I.; Alsayed, R.K.M.E.; Choudhry, H.; Ahmad, A. Exosome-Mediated Response to Cancer Therapy: Modulation of Epigenetic Machinery. Int. J. Mol. Sci. 2022, 23, 6222. [Google Scholar] [CrossRef]
- Yamana, K.; Inoue, J.; Yoshida, R.; Sakata, J.; Nakashima, H.; Arita, H.; Kawaguchi, S.; Gohara, S.; Nagao, Y.; Takeshita, H.; et al. Extracellular vesicles derived from radioresistant oral squamous cell carcinoma cells contribute to the acquisition of radioresistance via the miR-503-3p-BAK axis. J. Extracell. Vesicles 2021, 10, e12169. [Google Scholar] [CrossRef] [PubMed]
- Cappello, F.; Fais, S. Extracellular vesicles in cancer pros and cons: The importance of the evidence-based medicine. Semin Cancer Biol. 2022. [Google Scholar] [CrossRef] [PubMed]
- Corvigno, S.; Johnson, A.M.; Wong, K.K.; Cho, M.S.; Afshar-Kharghan, V.; Menter, D.G.; Sood, A.K. Novel Markers for Liquid Biopsies in Cancer Management: Circulating Platelets and Extracellular Vesicles. Mol. Cancer Ther. 2022, 21, 1067–1075. [Google Scholar] [CrossRef] [PubMed]
- Słomka, A.; Mocan, T.; Wang, B.; Nenu, I.; Urban, S.K.; Gonzales-Carmona, M.; Schmidt-Wolf, I.G.H.; Lukacs-Kornek, V.; Strassburg, C.P.; Spârchez, Z.; et al. EVs as Potential New Therapeutic Tool/Target in Gastrointestinal Cancer and HCC. Cancers 2020, 12, 3019. [Google Scholar] [CrossRef]
- Słomka, A.; Wang, B.; Mocan, T.; González-Carmona, M.; Strassburg, C.P.; Lukacs-Kornek, V.; Kornek, M.T. Extracellular vesicles small RNA clusters: Hit the nail on the head of liver cancer detection. Hepatobiliary Surg. Nutr. 2022, 11, 100–102. [Google Scholar] [CrossRef]
- Bunduc, S.; Gede, N.; Váncsa, S.; Lillik, V.; Kiss, S.; Juhász, M.F.; Erőss, B.; Szakács, Z.; Gheorghe, C.; Mikó, A.; et al. Exosomes as prognostic biomarkers in pancreatic ductal adenocarcinoma-a systematic review and meta-analysis. Transl. Res. 2022, 244, 126–136. [Google Scholar] [CrossRef]
- Jafari, D.; Tiyuri, A.; Rezaei, E.; Moradi, Y.; Jafari, R.; Jokar Shoorijeh, F.; Barati, M. Diagnostic accuracy of cerebrospinal fluid and serum-isolated extracellular vesicles for glioblastoma: A systematic review and meta-analysis. Expert Rev. Mol. Diagn 2020, 20, 1075–1085. [Google Scholar] [CrossRef]
- Xiang, H.; Li, F.; Luo, J.; Long, W.; Hong, L.; Hu, Y.; Du, H.; Yuan, Y.; Luo, M. A meta-analysis on the relationship of exosomes and the prognosis of lung cancer. Medicine 2021, 100, e25332. [Google Scholar] [CrossRef]
- Guo, D.; Yuan, J.; Xie, A.; Lin, Z.; Li, X.; Chen, J. Diagnostic performance of circulating exosomes in human cancer: A meta-analysis. J. Clin. Lab. Anal. 2020, 34, e23341. [Google Scholar] [CrossRef]
- Miller, K.D.; Nogueira, L.; Mariotto, A.B.; Rowland, J.H.; Yabroff, K.R.; Alfano, C.M.; Jemal, A.; Kramer, J.L.; Siegel, R.L. Cancer treatment and survivorship statistics, 2019. CA Cancer J. Clin. 2019, 69, 363–385. [Google Scholar] [CrossRef] [Green Version]
- Bleker de Oliveira, M.; Koshkin, V.; Liu, G.; Krylov, S.N. Analytical Challenges in Development of Chemoresistance Predictors for Precision Oncology. Anal. Chem. 2020, 92, 12101–12110. [Google Scholar] [CrossRef] [PubMed]
- Jing, Y.; Liang, W.; Liu, J.; Zhang, L.; Wei, J.; Yang, J.; Zhang, Y.; Huang, Z. Autophagy-mediating microRNAs in cancer chemoresistance. Cell Biol. Toxicol. 2020, 36, 517–536. [Google Scholar] [CrossRef] [PubMed]
- Madden, E.C.; Gorman, A.M.; Logue, S.E.; Samali, A. Tumour Cell Secretome in Chemoresistance and Tumour Recurrence. Trends Cancer 2020, 6, 489–505. [Google Scholar] [CrossRef] [PubMed]
- Khot, V.M.; Salunkhe, A.B.; Pricl, S.; Bauer, J.; Thorat, N.D.; Townley, H. Nanomedicine-driven molecular targeting, drug delivery, and therapeutic approaches to cancer chemoresistance. Drug. Discov. Today 2021, 26, 724–739. [Google Scholar] [CrossRef] [PubMed]
- Namee, N.M.; O’Driscoll, L. Extracellular vesicles and anti-cancer drug resistance. Biochim. Biophys. Acta Rev. Cancer 2018, 1870, 123–136. [Google Scholar] [CrossRef] [PubMed]
- Xavier, C.P.R.; Caires, H.R.; Barbosa, M.A.G.; Bergantim, R.; Guimarães, J.E.; Vasconcelos, M.H. The Role of Extracellular Vesicles in the Hallmarks of Cancer and Drug Resistance. Cells 2020, 9, 1141. [Google Scholar] [CrossRef] [PubMed]
- Bach, D.H.; Hong, J.Y.; Park, H.J.; Lee, S.K. The role of exosomes and miRNAs in drug-resistance of cancer cells. Int. J. Cancer 2017, 141, 220–230. [Google Scholar] [CrossRef]
- Steinbichler, T.B.; Dudás, J.; Skvortsov, S.; Ganswindt, U.; Riechelmann, H.; Skvortsova, I.I. Therapy resistance mediated by exosomes. Mol. Cancer 2019, 18, 58. [Google Scholar] [CrossRef]
- Hayatudin, R.; Fong, Z.; Ming, L.C.; Goh, B.H.; Lee, W.L.; Kifli, N. Overcoming Chemoresistance via Extracellular Vesicle Inhibition. Front. Mol. Biosci. 2021, 8, 629874. [Google Scholar] [CrossRef]
- Słomka, A.; Wang, B.; Mocan, T.; Horhat, A.; Willms, A.G.; Schmidt-Wolf, I.G.H.; Strassburg, C.P.; Gonzalez-Carmona, M.A.; Lukacs-Kornek, V.; Kornek, M.T. Extracellular Vesicles and Circulating Tumour Cells—Complementary liquid biopsies or standalone concepts. Theranostics 2022, 12, 5836–5855. [Google Scholar] [CrossRef]
- Catalanotto, C.; Cogoni, C.; Zardo, G. MicroRNA in Control of Gene Expression: An Overview of Nuclear Functions. Int. J. Mol. Sci. 2016, 17, 1712. [Google Scholar] [CrossRef] [PubMed]
- Statello, L.; Guo, C.J.; Chen, L.L.; Huarte, M. Gene regulation by long non-coding RNAs and its biological functions. Nat. Rev. Mol. Cell Biol. 2021, 22, 96–118. [Google Scholar] [CrossRef] [PubMed]
- Shao, T.; Pan, Y.H.; Xiong, X.D. Circular RNA: An important player with multiple facets to regulate its parental gene expression. Mol. Ther. Nucleic Acids 2021, 23, 369–376. [Google Scholar] [CrossRef] [PubMed]
- Sun, H.; Sun, R.; Song, X.; Gu, W.; Shao, Y. Mechanism and clinical value of exosomes and exosomal contents in regulating solid tumor radiosensitivity. J. Transl. Med. 2022, 20, 189. [Google Scholar] [CrossRef]
- Ye, Q.; Liu, K.; Shen, Q.; Li, Q.; Hao, J.; Han, F.; Jiang, R.W. Reversal of Multidrug Resistance in Cancer by Multi-Functional Flavonoids. Front. Oncol. 2019, 9, 487. [Google Scholar] [CrossRef]
- Shi, S.; Huang, X.; Ma, X.; Zhu, X.; Zhang, Q. Research of the mechanism on miRNA193 in exosomes promotes cisplatin resistance in esophageal cancer cells. PLoS ONE 2020, 15, e0225290. [Google Scholar] [CrossRef]
- Zhang, T.; Zhang, P.; Li, H.X. CAFs-Derived Exosomal miRNA-130a Confers Cisplatin Resistance of NSCLC Cells Through PUM2-Dependent Packaging. Int. J. Nanomed. 2021, 16, 561–577. [Google Scholar] [CrossRef]
- Li, Z.; Niu, H.; Qin, Q.; Yang, S.; Wang, Q.; Yu, C.; Wei, Z.; Jin, Z.; Wang, X.; Yang, A.; et al. lncRNA UCA1 Mediates Resistance to Cisplatin by Regulating the miR-143/FOSL2-Signaling Pathway in Ovarian Cancer. Mol. Ther. Nucleic Acids 2019, 17, 92–101. [Google Scholar] [CrossRef]
- Luo, X.; Wei, J.; Yang, F.L.; Pang, X.X.; Shi, F.; Wei, Y.X.; Liao, B.Y.; Wang, J.L. Exosomal lncRNA HNF1A-AS1 affects cisplatin resistance in cervical cancer cells through regulating microRNA-34b/TUFT1 axis. Cancer Cell Int. 2019, 19, 323. [Google Scholar] [CrossRef]
- Wang, J.; Lv, B.; Su, Y.; Wang, X.; Bu, J.; Yao, L. Exosome-Mediated Transfer of lncRNA HOTTIP Promotes Cisplatin Resistance in Gastric Cancer Cells by Regulating HMGA1/miR-218 Axis. Onco Targets Ther. 2019, 12, 11325–11338. [Google Scholar] [CrossRef] [Green Version]
- Wang, X.; Yu, H.; Yu, Z.; Wang, D. Exosomal lncRNA HEIH promotes cisplatin resistance in tongue squamous cell carcinoma via targeting miR-3619-5p/HDGF axis. Acta Histochem. 2020, 122, 151647. [Google Scholar] [CrossRef] [PubMed]
- Xin, L.; Zhou, L.Q.; Liu, C.; Zeng, F.; Yuan, Y.W.; Zhou, Q.; Li, S.H.; Wu, Y.; Wang, J.L.; Wu, D.Z.; et al. Transfer of LncRNA CRNDE in TAM-derived exosomes is linked with cisplatin resistance in gastric cancer. EMBO Rep. 2021, 22, e52124. [Google Scholar] [CrossRef] [PubMed]
- Seimiya, T.; Otsuka, M.; Iwata, T.; Shibata, C.; Tanaka, E.; Suzuki, T.; Koike, K. Emerging Roles of Exosomal Circular RNAs in Cancer. Front. Cell Dev. Biol. 2020, 8, 568366. [Google Scholar] [CrossRef] [PubMed]
- Xu, X.; Tao, R.; Sun, L.; Ji, X. Exosome-transferred hsa_circ_0014235 promotes DDP chemoresistance and deteriorates the development of non-small cell lung cancer by mediating the miR-520a-5p/CDK4 pathway. Cancer Cell Int. 2020, 20, 552. [Google Scholar] [CrossRef] [PubMed]
- Shi, Q.; Ji, T.; Ma, Z.; Tan, Q.; Liang, J. Serum Exosomes-Based Biomarker circ_0008928 Regulates Cisplatin Sensitivity, Tumor Progression, and Glycolysis Metabolism by miR-488/HK2 Axis in Cisplatin-Resistant Nonsmall Cell Lung Carcinoma. Cancer Biother. Radiopharm. 2021. [Google Scholar] [CrossRef]
- Wang, X.; Wang, H.; Jiang, H.; Qiao, L.; Guo, C. Circular RNAcirc_0076305 Promotes Cisplatin (DDP) Resistance of Non-Small Cell Lung Cancer Cells by Regulating ABCC1 Through miR-186-5p. Cancer Biother. Radiopharm. 2021. [Google Scholar] [CrossRef]
- Luo, Y.; Gui, R. Circulating exosomal circFoxp1 confers cisplatin resistance in epithelial ovarian cancer cells. J. Gynecol. Oncol. 2020, 31, e75. [Google Scholar] [CrossRef]
- Zang, R.; Qiu, X.; Song, Y.; Wang, Y. Exosomes Mediated Transfer of Circ_0000337 Contributes to Cisplatin (CDDP) Resistance of Esophageal Cancer by Regulating JAK2 via miR-377-3p. Front. Cell Dev. Biol. 2021, 9, 673237. [Google Scholar] [CrossRef]
- Liu, S.; Wu, M.; Peng, M. Circ_0000260 Regulates the Development and Deterioration of Gastric Adenocarcinoma with Cisplatin Resistance by Upregulating MMP11 via Targeting MiR-129-5p. Cancer Manag. Res. 2020, 12, 10505–10519. [Google Scholar] [CrossRef]
- Silva, I.L.Z.; Kohata, A.A.; Shigunov, P. Modulation and function of Pumilio proteins in cancer. Semin Cancer Biol. 2022. [Google Scholar] [CrossRef]
- Smialek, M.J.; Ilaslan, E.; Sajek, M.P.; Jaruzelska, J. Role of PUM RNA-Binding Proteins in Cancer. Cancers 2021, 13, 129. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Cho, K.B.; Li, Y.; Tao, G.; Xie, Z.; Guo, B. Long Noncoding RNA (lncRNA)-Mediated Competing Endogenous RNA Networks Provide Novel Potential Biomarkers and Therapeutic Targets for Colorectal Cancer. Int. J. Mol. Sci. 2019, 20, E5758. [Google Scholar] [CrossRef] [PubMed]
- Paraskevopoulou, M.D.; Hatzigeorgiou, A.G. Analyzing MiRNA-LncRNA Interactions. Methods Mol. Biol. 2016, 1402, 271–286. [Google Scholar] [CrossRef]
- Li, Y.; Wang, T.; Li, Y.; Chen, D.; Yu, Z.; Jin, L.; Ni, L.; Yang, S.; Mao, X.; Gui, Y.; et al. Identification of long-non coding RNA UCA1 as an oncogene in renal cell carcinoma. Mol. Med. Rep. 2016, 13, 3326–3334. [Google Scholar] [CrossRef] [PubMed]
- Liu, Z.; Wei, X.; Zhang, A.; Li, C.; Bai, J.; Dong, J. Long non-coding RNA HNF1A-AS1 functioned as an oncogene and autophagy promoter in hepatocellular carcinoma through sponging hsa-miR-30b-5p. Biochem. Biophys. Res. Commun. 2016, 473, 1268–1275. [Google Scholar] [CrossRef] [PubMed]
- Ghafouri-Fard, S.; Dashti, S.; Taheri, M. The HOTTIP (HOXA transcript at the distal tip) lncRNA: Review of oncogenic roles in human. Biomed. Pharmacother. 2020, 127, 110158. [Google Scholar] [CrossRef]
- Chen, C.; Gu, C.; Ren, Q.; Ding, F.; Pan, Q.; Niu, Y.; Ma, D.; Wu, L. lncRNA HEIH, an indicator of high malignancy and poor prognosis, functions as an oncogene in breast cancer. Mol. Med. Rep. 2020, 22, 2869–2877. [Google Scholar] [CrossRef]
- Lu, Y.; Sha, H.; Sun, X.; Zhang, Y.; Wu, Y.; Zhang, J.; Zhang, H.; Wu, J.; Feng, J. CRNDE: An oncogenic long non-coding RNA in cancers. Cancer Cell Int. 2020, 20, 162. [Google Scholar] [CrossRef]
- Li, S.; Fang, X.D.; Wang, X.Y.; Fei, B.Y. Fos-like antigen 2 (FOSL2) promotes metastasis in colon cancer. Exp. Cell Res. 2018, 373, 57–61. [Google Scholar] [CrossRef]
- Wan, X.; Guan, S.; Hou, Y.; Qin, Y.; Zeng, H.; Yang, L.; Qiao, Y.; Liu, S.; Li, Q.; Jin, T.; et al. FOSL2 promotes VEGF-independent angiogenesis by transcriptionnally activating Wnt5a in breast cancer-associated fibroblasts. Theranostics 2021, 11, 4975–4991. [Google Scholar] [CrossRef]
- Li, S.; Liu, Z.; Fang, X.D.; Wang, X.Y.; Fei, B.Y. MicroRNA (miR)-597-5p Inhibits Colon Cancer Cell Migration and Invasion by Targeting FOS-Like Antigen 2 (FOSL2). Front. Oncol. 2019, 9, 495. [Google Scholar] [CrossRef]
- He, J.; Mai, J.; Li, Y.; Chen, L.; Xu, H.; Zhu, X.; Pan, Q. miR-597 inhibits breast cancer cell proliferation, migration and invasion through FOSL2. Oncol. Rep. 2017, 37, 2672–2678. [Google Scholar] [CrossRef]
- Hein, S.; Mahner, S.; Kanowski, C.; Löning, T.; Jänicke, F.; Milde-Langosch, K. Expression of Jun and Fos proteins in ovarian tumors of different malignant potential and in ovarian cancer cell lines. Oncol. Rep. 2009, 22, 177–183. [Google Scholar] [CrossRef] [PubMed]
- Sun, L.; Guo, Z.; Sun, J.; Li, J.; Dong, Z.; Zhang, Y.; Chen, J.; Kan, Q.; Yu, Z. MiR-133a acts as an anti-oncogene in Hepatocellular carcinoma by inhibiting FOSL2 through TGF-β/Smad3 signaling pathway. Biomed. Pharmacother. 2018, 107, 168–176. [Google Scholar] [CrossRef] [PubMed]
- Sun, X.; Dai, G.; Yu, L.; Hu, Q.; Chen, J.; Guo, W. miR-143-3p inhibits the proliferation, migration and invasion in osteosarcoma by targeting FOSL2. Sci. Rep. 2018, 8, 606. [Google Scholar] [CrossRef] [PubMed]
- Zhou, B.; Zhan, H.; Tin, L.; Liu, S.; Xu, J.; Dong, Y.; Li, X.; Wu, L.; Guo, W. TUFT1 regulates metastasis of pancreatic cancer through HIF1-Snail pathway induced epithelial-mesenchymal transition. Cancer Lett. 2016, 382, 11–20. [Google Scholar] [CrossRef]
- Kawasaki, N.; Isogaya, K.; Dan, S.; Yamori, T.; Takano, H.; Yao, R.; Morishita, Y.; Taguchi, L.; Morikawa, M.; Heldin, C.H.; et al. TUFT1 interacts with RABGAP1 and regulates mTORC1 signaling. Cell Discov. 2018, 4, 1. [Google Scholar] [CrossRef]
- Lin, H.; Zeng, W.; Lei, Y.; Chen, D.; Nie, Z. Tuftelin 1 (TUFT1) Promotes the Proliferation and Migration of Renal Cell Carcinoma via PI3K/AKT Signaling Pathway. Pathol. Oncol. Res. 2021, 27, 640936. [Google Scholar] [CrossRef]
- Zhu, L.; Zhou, K.X.; Ma, M.Z.; Yao, L.L.; Zhang, Y.L.; Li, H.; Du, C.; Yang, X.M. Tuftelin 1 Facilitates Hepatocellular Carcinoma Progression through Regulation of Lipogenesis and Focal Adhesion Maturation. J. Immunol. Res. 2022, 2022, 1590717. [Google Scholar] [CrossRef]
- Liu, H.; Zhu, J.; Mao, Z.; Zhang, G.; Hu, X.; Chen, F. Tuft1 promotes thyroid carcinoma cell invasion and proliferation and suppresses apoptosis through the Akt-mTOR/GSK3β signaling pathway. Am. J. Transl. Res. 2018, 10, 4376–4384. [Google Scholar]
- Wu, M.N.; Zheng, W.J.; Ye, W.X.; Wang, L.; Chen, Y.; Yang, J.; Yao, D.F.; Yao, M. Oncogenic tuftelin 1 as a potential molecular-targeted for inhibiting hepatocellular carcinoma growth. World J. Gastroenterol. 2021, 27, 3327–3341. [Google Scholar] [CrossRef] [PubMed]
- Liu, W.; Zhang, L.; Jin, Z.; Zhao, M.; Li, Z.; Chen, G.; Sun, L.; Chen, B. TUFT1 is expressed in breast cancer and involved in cancer cell proliferation and survival. Oncotarget 2017, 8, 74962–74974. [Google Scholar] [CrossRef] [PubMed]
- Pallante, P.; Sepe, R.; Puca, F.; Fusco, A. High mobility group a proteins as tumor markers. Front. Med. (Lausanne) 2015, 2, 15. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Hu, L.; Zheng, Y.; Guo, L. HMGA1 in cancer: Cancer classification by location. J. Cell Mol. Med. 2019, 23, 2293–2302. [Google Scholar] [CrossRef] [PubMed]
- De Martino, M.; Esposito, F.; Fusco, A. Critical role of the high mobility group A proteins in hematological malignancies. Hematol. Oncol. 2022, 40, 2–10. [Google Scholar] [CrossRef]
- Milella, M.; Falcone, I.; Conciatori, F.; Cesta Incani, U.; Del Curatolo, A.; Inzerilli, N.; Nuzzo, C.M.; Vaccaro, V.; Vari, S.; Cognetti, F.; et al. PTEN: Multiple Functions in Human Malignant Tumors. Front. Oncol. 2015, 5, 24. [Google Scholar] [CrossRef]
- Misra, S.; Ghosh, G.; Chowdhury, S.G.; Karmakar, P. Non-canonical function of nuclear PTEN and its implication on tumorigenesis. DNA Repair. (Amst.) 2021, 107, 103197. [Google Scholar] [CrossRef]
- Cai, B.; Yang, L.; Do Jung, Y.; Zhang, Y.; Liu, X.; Zhao, P.; Li, J. PTEN: An Emerging Potential Target for Therapeutic Intervention in Respiratory Diseases. Oxid. Med. Cell. Longev. 2022, 2022, 4512503. [Google Scholar] [CrossRef]
- Wang, K.; Liu, J.; Li, Y.L.; Li, J.P.; Zhang, R. Ubiquitination/de-ubiquitination: A promising therapeutic target for PTEN reactivation in cancer. Biochim. Biophys. Acta Rev. Cancer 2022, 1877, 188723. [Google Scholar] [CrossRef]
- Hu, T.H.; Huang, C.C.; Liu, L.F.; Lin, P.R.; Liu, S.Y.; Chang, H.W.; Changchien, C.S.; Lee, C.M.; Chuang, J.H.; Tai, M.H. Expression of hepatoma-derived growth factor in hepatocellular carcinoma. Cancer 2003, 98, 1444–1456. [Google Scholar] [CrossRef]
- Ren, H.; Tang, X.; Lee, J.J.; Feng, L.; Everett, A.D.; Hong, W.K.; Khuri, F.R.; Mao, L. Expression of hepatoma-derived growth factor is a strong prognostic predictor for patients with early-stage non-small-cell lung cancer. J. Clin. Oncol. 2004, 22, 3230–3237. [Google Scholar] [CrossRef] [PubMed]
- Yamamoto, S.; Tomita, Y.; Hoshida, Y.; Takiguchi, S.; Fujiwara, Y.; Yasuda, T.; Doki, Y.; Yoshida, K.; Aozasa, K.; Nakamura, H.; et al. Expression of hepatoma-derived growth factor is correlated with lymph node metastasis and prognosis of gastric carcinoma. Clin. Cancer Res. 2006, 12, 117–122. [Google Scholar] [CrossRef]
- Shetty, A.; Dasari, S.; Banerjee, S.; Gheewala, T.; Zheng, G.; Chen, A.; Kajdacsy-Balla, A.; Bosland, M.C.; Munirathinam, G. Hepatoma-derived growth factor: A survival-related protein in prostate oncogenesis and a potential target for vitamin K2. Urol. Oncol. 2016, 34, e1–e483. [Google Scholar] [CrossRef]
- Enomoto, H.; Nakamura, H.; Nishikawa, H.; Nishiguchi, S.; Iijima, H. Hepatoma-Derived Growth Factor: An Overview and Its Role as a Potential Therapeutic Target Molecule for Digestive Malignancies. Int. J. Mol. Sci. 2020, 21, E4216. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.; Zhou, M.; Yant, L.; Huang, C. Circular RNA in disease: Basic properties and biomedical relevance. Wiley Interdisci. Rev. RNA 2022, e1723. [Google Scholar] [CrossRef] [PubMed]
- Goel, S.; DeCristo, M.J.; McAllister, S.S.; Zhao, J.J. CDK4/6 Inhibition in Cancer: Beyond Cell Cycle Arrest. Trends Cell Biol. 2018, 28, 911–925. [Google Scholar] [CrossRef]
- Goel, S.; Bergholz, J.S.; Zhao, J.J. Targeting CDK4 and CDK6 in cancer. Nat. Rev. Cancer 2022, 22, 356–372. [Google Scholar] [CrossRef]
- Adon, T.; Shanmugarajan, D.; Kumar, H.Y. CDK4/6 inhibitors: A brief overview and prospective research directions. RSC Adv. 2021, 11, 29227–29246. [Google Scholar] [CrossRef]
- Fassl, A.; Geng, Y.; Sicinski, P. CDK4 and CDK6 kinases: From basic science to cancer therapy. Science 2022, 375, eabc1495. [Google Scholar] [CrossRef]
- Patra, K.C.; Wang, Q.; Bhaskar, P.T.; Miller, L.; Wang, Z.; Wheaton, W.; Chandel, N.; Laakso, M.; Muller, W.J.; Allen, E.L.; et al. Hexokinase 2 is required for tumor initiation and maintenance and its systemic deletion is therapeutic in mouse models of cancer. Cancer Cell 2013, 24, 213–228. [Google Scholar] [CrossRef]
- Anderson, M.; Marayati, R.; Moffitt, R.; Yeh, J.J. Hexokinase 2 promotes tumor growth and metastasis by regulating lactate production in pancreatic cancer. Oncotarget 2017, 8, 56081–56094. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, L.; Yan, X.; Chen, J.; Zhan, Q.; Hua, Y.; Xu, S.; Li, Z.; Wang, Z.; Dong, Y.; Zuo, D.; et al. Hexokinase 2 discerns a novel circulating tumor cell population associated with poor prognosis in lung cancer patients. Proc. Natl. Acad. Sci. USA 2021, 118, e2012228118. [Google Scholar] [CrossRef] [PubMed]
- Sun, Z.; Tan, Z.; Peng, C.; Yi, W. HK2 is associated with the Warburg effect and proliferation in liver cancer: Targets for effective therapy with glycyrrhizin. Mol. Med. Rep. 2021, 23, 343. [Google Scholar] [CrossRef] [PubMed]
- Ciscato, F.; Ferrone, L.; Masgras, I.; Laquatra, C.; Rasola, A. Hexokinase 2 in Cancer: A Prima Donna Playing Multiple Characters. Int. J. Mol. Sci. 2021, 22, 4716. [Google Scholar] [CrossRef] [PubMed]
- Zhang, D.; Wang, H.; Yu, W.; Qiao, F.; Su, X.; Xu, H. Downregulation of hexokinase 2 improves radiosensitivity of breast cancer. Transl. Cancer Res. 2019, 8, 290–297. [Google Scholar] [CrossRef]
- Shangguan, X.; He, J.; Ma, Z.; Zhang, W.; Ji, Y.; Shen, K.; Yue, Z.; Li, W.; Xin, Z.; Zheng, Q.; et al. SUMOylation controls the binding of hexokinase 2 to mitochondria and protects against prostate cancer tumorigenesis. Nat. Commun. 2021, 12, 1812. [Google Scholar] [CrossRef]
- Cole, S.P. Multidrug resistance protein 1 (MRP1, ABCC1), a “multitasking” ATP-binding cassette (ABC) transporter. J. Biol. Chem. 2014, 289, 30880–30888. [Google Scholar] [CrossRef]
- Zhang, Y.K.; Wang, Y.J.; Gupta, P.; Chen, Z.S. Multidrug Resistance Proteins (MRPs) and Cancer Therapy. AAPS J. 2015, 17, 802–812. [Google Scholar] [CrossRef]
- Suzuki, T.; Nishio, K.; Tanabe, S. The MRP family and anticancer drug metabolism. Curr. Drug Metab. 2001, 2, 367–377. [Google Scholar] [CrossRef]
- Kruh, G.D.; Zeng, H.; Rea, P.A.; Liu, G.; Chen, Z.S.; Lee, K.; Belinsky, M.G. MRP subfamily transporters and resistance to anticancer agents. J. Bioenerg. Biomembr. 2001, 33, 493–501. [Google Scholar] [CrossRef]
- Kruh, G.D.; Belinsky, M.G. The MRP family of drug efflux pumps. Oncogene 2003, 22, 7537–7552. [Google Scholar] [CrossRef]
- Haimeur, A.; Conseil, G.; Deeley, R.G.; Cole, S.P. The MRP-related and BCRP/ABCG2 multidrug resistance proteins: Biology, substrate specificity and regulation. Curr. Drug Metab. 2004, 5, 21–53. [Google Scholar] [CrossRef]
- Alberich-Jordà, M.; Wouters, B.; Balastik, M.; Shapiro-Koss, C.; Zhang, H.; Di Ruscio, A.; DiRuscio, A.; Radomska, H.S.; Ebralidze, A.K.; Amabile, G.; et al. C/EBPγ deregulation results in differentiation arrest in acute myeloid leukemia. J. Clin. Investig. 2012, 122, 4490–4504. [Google Scholar] [CrossRef]
- Huang, Y.; Lin, L.; Shen, Z.; Li, Y.; Cao, H.; Peng, L.; Qiu, Y.; Cheng, X.; Meng, M.; Lu, D.; et al. CEBPG promotes esophageal squamous cell carcinoma progression by enhancing PI3K-AKT signaling. Am. J. Cancer Res. 2020, 10, 3328–3344. [Google Scholar]
- Shi, H.; Niimi, A.; Takeuchi, T.; Shiogama, K.; Mizutani, Y.; Kajino, T.; Inada, K.; Hase, T.; Hatta, T.; Shibata, H.; et al. CEBPγ facilitates lamellipodia formation and cancer cell migration through CERS6 upregulation. Cancer Sci. 2021, 112, 2770–2780. [Google Scholar] [CrossRef]
- Labat-de-Hoz, L.; Alonso, M.A. Formins in Human Disease. Cells 2021, 10, 2554. [Google Scholar] [CrossRef]
- Wu, Y.; Shen, Z.; Wang, K.; Ha, Y.; Lei, H.; Jia, Y.; Ding, R.; Wu, D.; Gan, S.; Li, R.; et al. High FMNL3 expression promotes nasopharyngeal carcinoma cell metastasis: Role in TGF-β1-induced epithelia-to-mesenchymal transition. Sci. Rep. 2017, 7, 42507. [Google Scholar] [CrossRef]
- Zeng, Y.F.; Xiao, Y.S.; Lu, M.Z.; Luo, X.J.; Hu, G.Z.; Deng, K.Y.; Wu, X.M.; Xin, H.B. Increased expression of formin-like 3 contributes to metastasis and poor prognosis in colorectal carcinoma. Exp. Mol. Pathol. 2015, 98, 260–267. [Google Scholar] [CrossRef]
- Zeng, Y.F.; Xiao, Y.S.; Liu, Y.; Luo, X.J.; Wen, L.D.; Liu, Q.; Chen, M. Formin-like 3 regulates RhoC/FAK pathway and actin assembly to promote cell invasion in colorectal carcinoma. World J. Gastroenterol. 2018, 24, 3884–3897. [Google Scholar] [CrossRef]
- Gardberg, M.; Heuser, V.D.; Koskivuo, I.; Koivisto, M.; Carpén, O. FMNL2/FMNL3 formins are linked with oncogenic pathways and predict melanoma outcome. J. Pathol. Clin. Res. 2016, 2, 41–52. [Google Scholar] [CrossRef]
- Liu, J.; Chen, S.; Chen, Y.; Geng, N.; Feng, C. High expression of FMNL3 associates with cancer cell migration, invasion, and unfavorable prognosis in tongue squamous cell carcinoma. J. Oral. Pathol. Med. 2019, 48, 459–467. [Google Scholar] [CrossRef]
- González de Vega, R.; Clases, D.; Fernández-Sánchez, M.L.; Eiró, N.; González, L.O.; Vizoso, F.J.; Doble, P.A.; Sanz-Medel, A. MMP-11 as a biomarker for metastatic breast cancer by immunohistochemical-assisted imaging mass spectrometry. Anal. Bioanal. Chem. 2019, 411, 639–646. [Google Scholar] [CrossRef]
- Lee, J.; Lee, J.; Kim, J.H. Identification of Matrix Metalloproteinase 11 as a Prognostic Biomarker in Pancreatic Cancer. Anticancer Res. 2019, 39, 5963–5971. [Google Scholar] [CrossRef]
- Kim, H.S.; Kim, M.G.; Min, K.W.; Jung, U.S.; Kim, D.H. High MMP-11 expression associated with low CD8+ T cells decreases the survival rate in patients with breast cancer. PLoS ONE 2021, 16, e0252052. [Google Scholar] [CrossRef]
- Ma, B.; Ran, R.; Liao, H.Y.; Zhang, H.H. The paradoxical role of matrix metalloproteinase-11 in cancer. Biomed. Pharmacother. 2021, 141, 111899. [Google Scholar] [CrossRef]
- Wang, X.; Zhang, H.; Yang, H.; Bai, M.; Ning, T.; Deng, T.; Liu, R.; Fan, Q.; Zhu, K.; Li, J.; et al. Exosome-delivered circRNA promotes glycolysis to induce chemoresistance through the miR-122-PKM2 axis in colorectal cancer. Mol. Oncol. 2020, 14, 539–555. [Google Scholar] [CrossRef]
- Zhong, Y.; Wang, D.; Ding, Y.; Tian, G.; Jiang, B. Circular RNA circ_0032821 contributes to oxaliplatin (OXA) resistance of gastric cancer cells by regulating SOX9 via miR-515-5p. Biotechnol. Lett. 2021, 43, 339–351. [Google Scholar] [CrossRef]
- Xu, Y.; Zhu, M. Novel exosomal miR-46146 transfer oxaliplatin chemoresistance in colorectal cancer. Clin. Transl. Oncol. 2020, 22, 1105–1116. [Google Scholar] [CrossRef]
- Ning, T.; Li, J.; He, Y.; Zhang, H.; Wang, X.; Deng, T.; Liu, R.; Li, H.; Bai, M.; Fan, Q.; et al. Exosomal miR-208b related with oxaliplatin resistance promotes Treg expansion in colorectal cancer. Mol. Ther. 2021, 29, 2723–2736. [Google Scholar] [CrossRef]
- Deng, X.; Ruan, H.; Zhang, X.; Xu, X.; Zhu, Y.; Peng, H.; Zhang, X.; Kong, F.; Guan, M. Long noncoding RNA CCAL transferred from fibroblasts by exosomes promotes chemoresistance of colorectal cancer cells. Int. J. Cancer 2020, 146, 1700–1716. [Google Scholar] [CrossRef]
- Ghafouri-Fard, S.; Hussen, B.M.; Mohaqiq, M.; Shoorei, H.; Baniahmad, A.; Taheri, M.; Jamali, E. Interplay Between Non-Coding RNAs and Programmed Cell Death Proteins. Front. Oncol. 2022, 12, 808475. [Google Scholar] [CrossRef] [PubMed]
- Schmid, T.; Jansen, A.P.; Baker, A.R.; Hegamyer, G.; Hagan, J.P.; Colburn, N.H. Translation inhibitor Pdcd4 is targeted for degradation during tumor promotion. Cancer Res. 2008, 68, 1254–1260. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hayashi, A.; Aishima, S.; Miyasaka, Y.; Nakata, K.; Morimatsu, K.; Oda, Y.; Nagai, E.; Oda, Y.; Tanaka, M.; Tsuneyoshi, M. Pdcd4 expression in intraductal papillary mucinous neoplasm of the pancreas: Its association with tumor progression and proliferation. Hum. Pathol. 2010, 41, 1507–1515. [Google Scholar] [CrossRef]
- Zhu, Y.; Zhao, K.; Prinz, A.; Keyvani, K.; Lambertz, N.; Kreitschmann-Andermahr, I.; Lei, T.; Sure, U. Loss of endothelial programmed cell death 10 activates glioblastoma cells and promotes tumor growth. Neuro. Oncol. 2016, 18, 538–548. [Google Scholar] [CrossRef]
- Nickel, A.C.; Wan, X.Y.; Saban, D.V.; Weng, Y.L.; Zhang, S.; Keyvani, K.; Sure, U.; Zhu, Y. Loss of programmed cell death 10 activates tumor cells and leads to temozolomide-resistance in glioblastoma. J. Neurooncol. 2019, 141, 31–41. [Google Scholar] [CrossRef] [PubMed]
- Wan, X.; Saban, D.V.; Kim, S.N.; Weng, Y.; Dammann, P.; Keyvani, K.; Sure, U.; Zhu, Y. PDCD10-Deficiency Promotes Malignant Behaviors and Tumor Growth via Triggering EphB4 Kinase Activity in Glioblastoma. Front. Oncol. 2020, 10, 1377. [Google Scholar] [CrossRef] [PubMed]
- Dayton, T.L.; Jacks, T.; Vander Heiden, M.G. PKM2, cancer metabolism, and the road ahead. EMBO Rep. 2016, 17, 1721–1730. [Google Scholar] [CrossRef]
- Zahra, K.; Dey, T.; Ashish; Mishra, S.P.; Pandey, U. Pyruvate kinase M2 and cancer: The role of PKM2 in promoting tumorigenesis. Front. Oncol. 2020, 10, 159. [Google Scholar] [CrossRef]
- Aldaz, P.; Otaegi-Ugartemendia, M.; Saenz-Antoñanzas, A.; Garcia-Puga, M.; Moreno-Valladares, M.; Flores, J.M.; Gerovska, D.; Arauzo-Bravo, M.J.; Samprón, N.; Matheu, A.; et al. SOX9 promotes tumor progression through the axis BMI1-p21CIP. Sci. Rep. 2020, 10, 357. [Google Scholar] [CrossRef]
- Chen, X.; Chen, S.; Yu, D. Protein kinase function of pyruvate kinase M2 and cancer. Cancer Cell Int. 2020, 20, 523. [Google Scholar] [CrossRef]
- Ma, Y.; Shepherd, J.; Zhao, D.; Bollu, L.R.; Tahaney, W.M.; Hill, J.; Zhang, Y.; Mazumdar, A.; Brown, P.H. SOX9 Is Essential for Triple-Negative Breast Cancer Cell Survival and Metastasis. Mol. Cancer Res. 2020, 18, 1825–1838. [Google Scholar] [CrossRef] [PubMed]
- Qin, H.; Yang, Y.; Jiang, B.; Pan, C.; Chen, W.; Diao, W.; Ding, M.; Cao, W.; Zhang, Z.; Chen, M.; et al. SOX9 in prostate cancer is upregulated by cancer-associated fibroblasts to promote tumor progression through HGF/c-Met-FRA1 signaling. FEBS J. 2021, 288, 5406–5429. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.F.; Dang, H.F.; Luo, X.; Wang, Q.Q.; Gao, C.; Tian, Y.X. Downregulation of SOX9 suppresses breast cancer cell proliferation and migration by regulating apoptosis and cell cycle arrest. Oncol. Lett. 2021, 22, 517. [Google Scholar] [CrossRef] [PubMed]
- Carrasco-Garcia, E.; Lopez, L.; Moncho-Amor, V.; Carazo, F.; Aldaz, P.; Collado, M.; Bell, D.; Gaafar, A.; Karamitopoulou, E.; Tzankov, A.; et al. SOX9 Triggers Different Epithelial to Mesenchymal Transition States to Promote Pancreatic Cancer Progression. Cancers 2022, 14, 916. [Google Scholar] [CrossRef]
- Sardar, D.; Chen, H.C.; Reyes, A.; Varadharajan, S.; Jain, A.; Mohila, C.; Curry, R.; Lozzi, B.; Rajendran, K.; Cervantes, A.; et al. Sox9 directs divergent epigenomic states in brain tumor subtypes. Proc. Natl. Acad. Sci. USA 2022, 119, e2202015119. [Google Scholar] [CrossRef]
- Wang, J.; Li, T.; Wang, B. Exosomal transfer of miR-25-3p promotes the proliferation and temozolomide resistance of glioblastoma cells by targeting FBXW7. Int. J. Oncol. 2021, 59, 64. [Google Scholar] [CrossRef]
- Zhang, Z.; Yin, J.; Lu, C.; Wei, Y.; Zeng, A.; You, Y. Exosomal transfer of long non-coding RNA SBF2-AS1 enhances chemoresistance to temozolomide in glioblastoma. J. Exp. Clin. Cancer Res. 2019, 38, 166. [Google Scholar] [CrossRef]
- Li, Z.; Meng, X.; Wu, P.; Zha, C.; Han, B.; Li, L.; Sun, N.; Qi, T.; Qin, J.; Zhang, Y.; et al. Glioblastoma Cell-Derived lncRNA-Containing Exosomes Induce Microglia to Produce Complement C5, Promoting Chemotherapy Resistance. Cancer Immunol. Res. 2021, 9, 1383–1399. [Google Scholar] [CrossRef]
- Si, J.; Li, W.; Li, X.; Cao, L.; Chen, Z.; Jiang, Z. Heparanase confers temozolomide resistance by regulation of exosome secretion and circular RNA composition in glioma. Cancer Sci. 2021, 112, 3491–3506. [Google Scholar] [CrossRef]
- Han, C.; Wang, S.; Wang, H.; Zhang, J. Exosomal circ-HIPK3 Facilitates Tumor Progression and Temozolomide Resistance by Regulating miR-421/ZIC5 Axis in Glioma. Cancer Biother. Radiopharm. 2021, 36, 537–548. [Google Scholar] [CrossRef]
- Tong, S.; Wang, Y.; Wu, J.; Long, J.; Zhong, P.; Wang, B. Comprehensive pharmacogenomics characterization of temozolomide response in gliomas. Eur. J. Pharmacol. 2021, 912, 174580. [Google Scholar] [CrossRef] [PubMed]
- Yeh, C.H.; Bellon, M.; Nicot, C. FBXW7: A critical tumor suppressor of human cancers. Mol. Cancer 2018, 17, 115. [Google Scholar] [CrossRef] [PubMed]
- Lan, H.; Sun, Y. Tumor Suppressor FBXW7 and Its Regulation of DNA Damage Response and Repair. Front. Cell Dev. Biol. 2021, 9, 751574. [Google Scholar] [CrossRef]
- Shteingauz, A.; Boyango, I.; Naroditsky, I.; Hammond, E.; Gruber, M.; Doweck, I.; Ilan, N.; Vlodavsky, I. Heparanase Enhances Tumor Growth and Chemoresistance by Promoting Autophagy. Cancer Res. 2015, 75, 3946–3957. [Google Scholar] [CrossRef] [PubMed]
- de Boer, C.; Armstrong, Z.; Lit, V.A.J.; Barash, U.; Ruijgrok, G.; Boyango, I.; Weitzenberg, M.M.; Schröder, S.P.; Sarris, A.J.C.; Meeuwenoord, N.J.; et al. Mechanism-based heparanase inhibitors reduce cancer metastasis in vivo. Proc. Natl. Acad. Sci. USA 2022, 119, e2203167119. [Google Scholar] [CrossRef] [PubMed]
- Coombe, D.R.; Gandhi, N.S. Heparanase: A Challenging Cancer Drug Target. Front. Oncol. 2019, 9, 1316. [Google Scholar] [CrossRef]
- Jayatilleke, K.M.; Hulett, M.D. Heparanase and the hallmarks of cancer. J. Transl. Med. 2020, 18, 453. [Google Scholar] [CrossRef]
- Satow, R.; Inagaki, S.; Kato, C.; Shimozawa, M.; Fukami, K. Identification of zinc finger protein of the cerebellum 5 as a survival factor of prostate and colorectal cancer cells. Cancer Sci. 2017, 108, 2405–2412. [Google Scholar] [CrossRef]
- Maimaiti, A.; Aizezi, A.; Anniwaer, J.; Ayitula, A.; Ali, B.; Dilixiati, M. Zinc finger of the cerebellum 5 promotes colorectal cancer cell proliferation and cell cycle progression through enhanced Cdk1/CDC25C signaling. Arch. Med. Sci. 2021, 17, 449–461. [Google Scholar] [CrossRef]
- Song, W.; Yu, W.; Li, D.; Cheng, C.; Wu, X.; Chen, J.; Zhang, W. ZIC5 promotes human hepatocellular carcinoma cell proliferation through upregulating COL1A1. J. Gastrointest. Oncol. 2022, 13, 1237–1247. [Google Scholar] [CrossRef]
- Satow, R.; Aiga, Y.; Watanabe, T.; Ishizuka, N.; Yoneda, A.; Fukami, K. Zic family member 5 promotes survival in human pancreatic cancer and cholangiocarcinoma cells. Biochem. Biophys. Rep. 2022, 31, 101289. [Google Scholar] [CrossRef] [PubMed]
- Mao, G.; Mu, Z.; Wu, D.A. Exosomal lncRNA FOXD3-AS1 upregulates ELAVL1 expression and activates PI3K/Akt pathway to enhance lung cancer cell proliferation, invasion, and 5-fluorouracil resistance. Acta Biochim. Biophys. Sin. (Shanghai) 2021, 53, 1484–1494. [Google Scholar] [CrossRef] [PubMed]
- Zhao, K.; Cheng, X.; Ye, Z.; Li, Y.; Peng, W.; Wu, Y.; Xing, C. Exosome-Mediated Transfer of circ_0000338 Enhances 5-Fluorouracil Resistance in Colorectal Cancer through Regulating MicroRNA 217 (miR-217) and miR-485-3p. Mol. Cell Biol. 2021, 41, e00517-20. [Google Scholar] [CrossRef] [PubMed]
- Zeng, Z.; Zhao, Y.; Chen, Q.; Zhu, S.; Niu, Y.; Ye, Z.; Hu, P.; Chen, D.; Xu, P.; Chen, J.; et al. Hypoxic exosomal HIF-1α-stabilizing circZNF91 promotes chemoresistance of normoxic pancreatic cancer cells via enhancing glycolysis. Oncogene 2021, 40, 5505–5517. [Google Scholar] [CrossRef]
- Hon, K.W.; Ab-Mutalib, N.S.; Abdullah, N.M.A.; Jamal, R.; Abu, N. Extracellular Vesicle-derived circular RNAs confers chemoresistance in Colorectal cancer. Sci. Rep. 2019, 9, 16497. [Google Scholar] [CrossRef]
- Monaco, F.; Gaetani, S.; Alessandrini, F.; Tagliabracci, A.; Bracci, M.; Valentino, M.; Neuzil, J.; Amati, M.; Bovenzi, M.; Tomasetti, M.; et al. Exosomal transfer of miR-126 promotes the anti-tumour response in malignant mesothelioma: Role of miR-126 in cancer-stroma communication. Cancer Lett. 2019, 463, 27–36. [Google Scholar] [CrossRef]
- Zhao, Z.; Ji, M.; Wang, Q.; He, N.; Li, Y. Circular RNA Cdr1as Upregulates SCAI to Suppress Cisplatin Resistance in Ovarian Cancer via miR-1270 Suppression. Mol. Ther. Nucleic Acids 2019, 18, 24–33. [Google Scholar] [CrossRef]
- Jiang, L.; Zhang, Y.; Guo, L.; Liu, C.; Wang, P.; Ren, W. Exosomal microRNA-107 reverses chemotherapeutic drug resistance of gastric cancer cells through HMGA2/mTOR/P-gp pathway. BMC Cancer 2021, 21, 1290. [Google Scholar] [CrossRef]
- Xu, Y.; Qiu, A.; Peng, F.; Tan, X.; Wang, J.; Gong, X. Exosomal transfer of circular RNA FBXW7 ameliorates the chemoresistance to oxaliplatin in colorectal cancer by sponging miR-18b-5p. Neoplasma 2021, 68, 108–118. [Google Scholar] [CrossRef]
- Zeng, A.; Wei, Z.; Yan, W.; Yin, J.; Huang, X.; Zhou, X.; Li, R.; Shen, F.; Wu, W.; Wang, X.; et al. Exosomal transfer of miR-151a enhances chemosensitivity to temozolomide in drug-resistant glioblastoma. Cancer Lett. 2018, 436, 10–21. [Google Scholar] [CrossRef]
- Cui, J.; Wang, H.; Zhang, X.; Sun, X.; Zhang, J.; Ma, J. Exosomal miR-200c suppresses chemoresistance of docetaxel in tongue squamous cell carcinoma by suppressing TUBB3 and PPP2R1B. Aging 2020, 12, 6756–6773. [Google Scholar] [CrossRef] [PubMed]
- Retraction: Exosomes function as nanoparticles to transfer miR-199a-3p to reverse chemoresistance to cisplatin in hepatocellular carcinoma. BioSci. Rep. 2022, 42, BSR-2019-4026_RET. [CrossRef] [PubMed]
- Lou, J.; Hao, Y.; Lin, K.; Lyu, Y.; Chen, M.; Wang, H.; Zou, D.; Jiang, X.; Wang, R.; Jin, D.; et al. Circular RNA CDR1as disrupts the p53/MDM2 complex to inhibit Gliomagenesis. Mol. Cancer 2020, 19, 138. [Google Scholar] [CrossRef]
- Jiang, J.; Li, R.; Wang, J.; Hou, J.; Qian, H.; Xu, W. Circular RNA CDR1as Inhibits the Metastasis of Gastric Cancer through Targeting miR-876-5p/GNG7 Axis. Gastroenterol. Res. Pract. 2021, 2021, 5583029. [Google Scholar] [CrossRef] [PubMed]
- Liu, T.; Zhang, X.; Du, L.; Wang, Y.; Liu, X.; Tian, H.; Wang, L.; Li, P.; Zhao, Y.; Duan, W.; et al. Retraction Note: Exosome-transmitted miR-128-3p increase chemosensitivity of oxaliplatin-resistant colorectal cancer. Mol. Cancer 2022, 21, 157. [Google Scholar] [CrossRef]
- Mocan, T.; Simão, A.L.; Castro, R.E.; Rodrigues, C.M.P.; Słomka, A.; Wang, B.; Strassburg, C.; Wöhler, A.; Willms, A.G.; Kornek, M. Liquid Biopsies in Hepatocellular Carcinoma: Are We Winning. J. Clin. Med. 2020, 9, 1541. [Google Scholar] [CrossRef]
- Urban, S.K.; Sänger, H.; Krawczyk, M.; Julich-Haertel, H.; Willms, A.; Ligocka, J.; Azkargorta, M.; Mocan, T.; Kahlert, C.; Kruk, B.; et al. Synergistic effects of extracellular vesicle phenotyping and AFP in hepatobiliary cancer differentiation. Liver Int. 2020, 40, 3103–3116. [Google Scholar] [CrossRef]
- Hekmatirad, S.; Moloudizargari, M.; Moghadamnia, A.A.; Kazemi, S.; Mohammadnia-Afrouzi, M.; Baeeri, M.; Moradkhani, F.; Asghari, M.H. Inhibition of Exosome Release Sensitizes U937 Cells to PEGylated Liposomal Doxorubicin. Front. Immunol. 2021, 12, 692654. [Google Scholar] [CrossRef]
- Liang, Y.; Duan, L.; Lu, J.; Xia, J. Engineering exosomes for targeted drug delivery. Theranostics 2021, 11, 3183–3195. [Google Scholar] [CrossRef]
- Chen, H.; Wang, L.; Zeng, X.; Schwarz, H.; Nanda, H.S.; Peng, X.; Zhou, Y. Exosomes, a New Star for Targeted Delivery. Front. Cell Dev. Biol. 2021, 9, 751079. [Google Scholar] [CrossRef]
- Jayasinghe, M.K.; Pirisinu, M.; Yang, Y.; Peng, B.; Pham, T.T.; Lee, C.Y.; Tan, M.; Vu, L.T.; Dang, X.T.; Pham, T.C.; et al. Surface-engineered extracellular vesicles for targeted delivery of therapeutic RNAs and peptides for cancer therapy. Theranostics 2022, 12, 3288–3315. [Google Scholar] [CrossRef] [PubMed]
- Murphy, D.E.; de Jong, O.G.; Brouwer, M.; Wood, M.J.; Lavieu, G.; Schiffelers, R.M.; Vader, P. Extracellular vesicle-based therapeutics: Natural versus engineered targeting and trafficking. Exp. Mol. Med. 2019, 51, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Claridge, B.; Lozano, J.; Poh, Q.H.; Greening, D.W. Development of Extracellular Vesicle Therapeutics: Challenges, Considerations, and Opportunities. Front. Cell Dev. Biol. 2021, 9, 734720. [Google Scholar] [CrossRef] [PubMed]
- Silva, A.K.A.; Morille, M.; Piffoux, M.; Arumugam, S.; Mauduit, P.; Larghero, J.; Bianchi, A.; Aubertin, K.; Blanc-Brude, O.; Noël, D.; et al. Development of extracellular vesicle-based medicinal products: A position paper of the group “Extracellular Vesicle translatiOn to clinicaL perspectiVEs—EVOLVE France”. Adv. Drug Deliv. Rev. 2021, 179, 114001. [Google Scholar] [CrossRef]
- Dooley, K.; McConnell, R.E.; Xu, K.; Lewis, N.D.; Haupt, S.; Youniss, M.R.; Martin, S.; Sia, C.L.; McCoy, C.; Moniz, R.J.; et al. A versatile platform for generating engineered extracellular vesicles with defined therapeutic properties. Mol. Ther. 2021, 29, 1729–1743. [Google Scholar] [CrossRef]


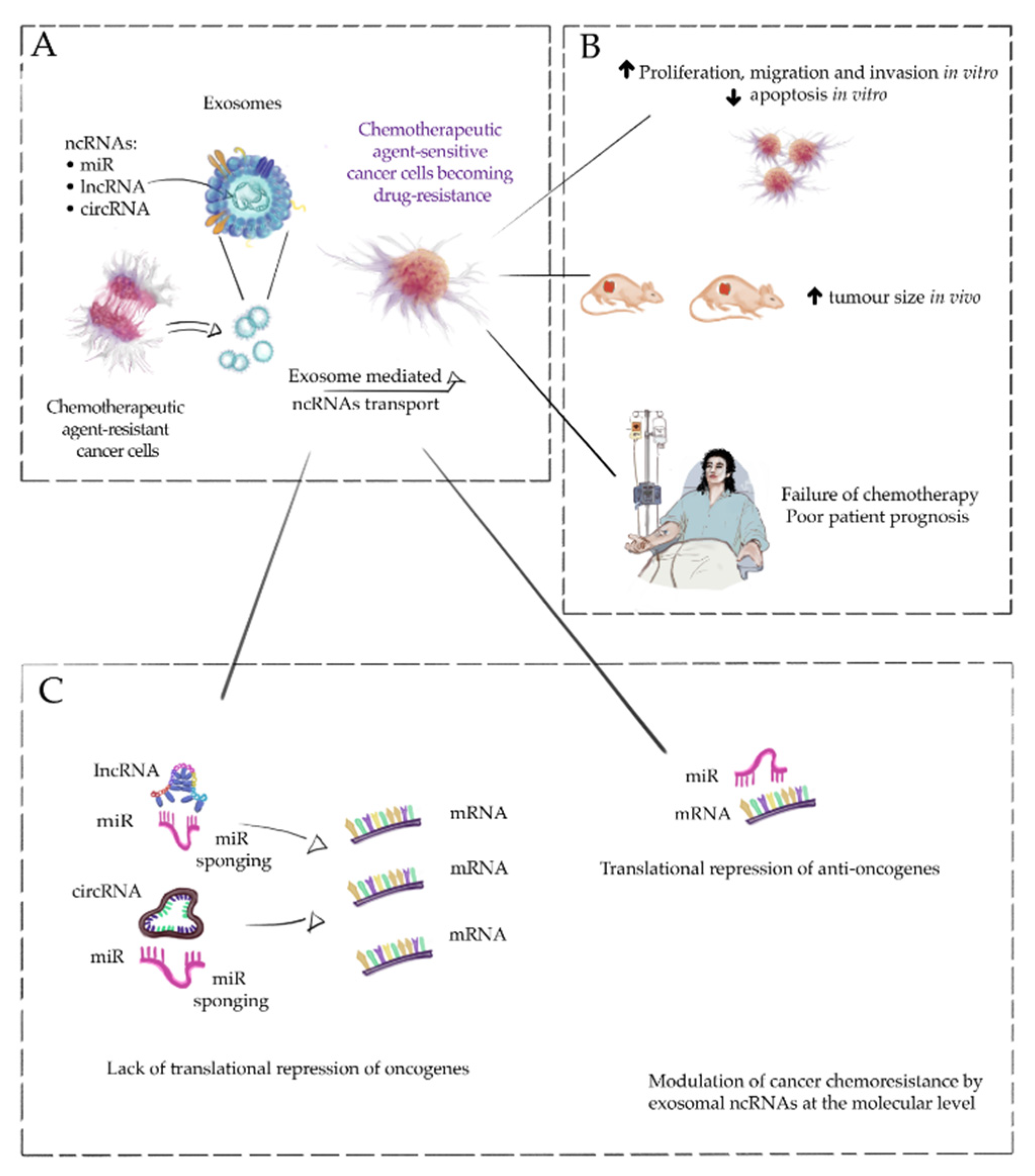{kind=link}
| Type of Cancer | Anticancer Agent | EXO Cellular Source | EXO Cargo | Targeted Regulatory Network | Main In Vitro ncRNAs/EXO Effect | Main In Vivo ncRNAs/EXO Effect | Ref. |
|---|---|---|---|---|---|---|---|
| Esophageal cancer | Cisplatin | TE-1/DDP | miR-193 | TFAP2C | (1) Inhibition of cisplatin-induced cancer cell cycle arrest (2) Inhibition of cancer cell apoptosis | (1) Increasing the tumor size in a mouse model | [36] |
| Non-small cell lung cancer | Cisplatin | CAFs | miR-130a | ND | (1) Increasing the cancer cell survival rate | (1) Increasing the tumor size in a mouse mode | [37] |
| Ovarian cancer | Cisplatin | ND* | lncRNA UCA1 | miR-143/FOSL2 | (1) Increasing the cancer cell proliferation (2) Inhibition of cancer cell apoptosis (3) Decreasing cisplatin-induced cytotoxicity | (1) Increasing the tumor size in a mouse mode | [38] |
| Cervical cancer | Cisplatin | HeLa/DDP | lncRNA HNF1A-AS1 | miR-34b/TUFT1 | (1) Increasing the cancer cell proliferation (2) Inhibition of the cancer cell apoptosis | (1) Increasing the tumor size in a mouse mode | [39] |
| Gastric cancer | Cisplatin | MGC-803/DDP MKN-45/DDP | lncRNA HOTTIP | miR-218/HMGA1 | (1) Increasing the cancer cell proliferation, migration, and invasion | (1) Chemotherapy failure of GC patients was associated with high lncRNA HOTTIP expression | [40] |
| Tongue squamous cell carcinoma | Cisplatin | SCC4/DDP | lncRNA HEIH | miR-3619-5p/HDGF | (1) Increasing the cancer cell proliferation (2) Inhibition of cancer cell apoptosis | ND | [41] |
| Gastric cancer | Cisplatin | TAMs | lncRNA CRNDE | NEDD4-1/PTEN | (1) Increasing the cancer cell survival rate and proliferation (2) Inhibition of cancer cell apoptosis | (1) Increasing the tumor size in a mouse mode | [42] |
| Non-small cell lung cancer | Cisplatin | ND * | hsa_circ_0014235 | miR-520a-5p/CDK4 | (1) Increasing the cancer cell proliferation, migration, and invasion | (1) Increasing the tumor size in a mouse mode | [44] |
| Non-small cell lung cancer | Cisplatin | ND * | circ_0008928 | miR-488/HK2 | (1) Increasing the cancer cell proliferation, migration, and invasion | (1) Chemotherapy failure of NSCLC patients was associated with high circ_0008928 expression | [45] |
| Non-small cell lung cancer | Cisplatin | ND * | circ_0076305 | miR-186-5p/ABCC1 | (1) Increasing the cancer cell survival rate | (1) Increasing the tumor size in a mouse model | [46] |
| Epithelial ovarian cancer | Cisplatin | ND * | circFoxp1 | miR-22/miR-150-3p/CEBPG/FMNL3 | (1) Increasing the cancer cell survival rate and proliferation | (1) Increasing the tumor size in a mouse mode (2) Chemotherapy failure of EOC patients was associated with high circFoxp1 expression | [47] |
| Esophageal cancer | Cisplatin | EC9706/DDP KYSE30/DDP | circ_0000337 | miR-377-3p/JAK2 | (1) Increasing the cancer cell proliferation, migration, and invasion (2) Inhibition of cancer cell apoptosis | (1) Increasing the tumor size in a mouse model | [48] |
| Gastric adenocarcinoma | Cisplatin | ND * | circ_0000260 | miR-129-5p/MMP11 | (1) Increasing the cancer cell proliferation, migration, and invasion (2) Inhibition of cancer cell apoptosis | (1) Increasing the tumor size in a mouse mode (2) Chemotherapy failure of GAC patients was associated with high circ_0000260 expression | [49] |
| Colorectal cancer | Oxaliplatin | SW480/OXA | hsa_circ_0005963 (ciRS-122) | miR-122/PKM2 | (1) Increasing the cancer cell survival rate (2) Inhibition of cancer cell apoptosis | (1) Increasing the tumor size in a mouse model | [116] |
| Gastric cancer | Oxaliplatin | HGC27/OXA AGS/OXA | circ_0032821 | miR-515-5p/SOX9 | (1) Increasing the cancer cell proliferation, migration, and invasion | (1) Increasing the tumor size in a mouse model | [117] |
| Colorectal cancer | Oxaliplatin | HCT116/OXA HT29/OXA | miR-46146 | PDCD10 | (1) Increasing the cancer cell proliferation (2) Inhibition of cancer cell apoptosis | ND | [118] |
| Colorectal cancer | Oxaliplatin | SW480/OXA | miR-208b | PDCD4 | (1) Tregs expansion | (1) Increasing the tumor size in a mouse model | [119] |
| Colorectal cancer | Oxaliplatin | CAFs | lncRNA CCAL | HuR/ Wnt/β-catenin | (1) Increasing the cancer cell survival rate (2) Inhibition of cancer cell apoptosis | (1) Increasing the tumor size in a mouse model | [120] |
| Type of Cancer | Anticancer Agent | EXO Cellular Source | EXO Cargo | Targeted Regulatory Network | Main In Vitro ncRNAs/EXO Effect | Main In Vivo ncRNAs/EXO Effect | Ref. |
|---|---|---|---|---|---|---|---|
| Glioblastoma | Temozolomide | A172/TMZ | miR-25-3p | FBXW7 | (1) Increasing the cancer cell proliferation (2) Inhibition of cancer cell apoptosis | (1) Increasing the tumor size in a mouse model (2) Chemotherapy failure of GBM patients was associated with high miR-25-3p expression | [136] |
| Glioblastoma | Temozolomide | Rec GBM N3T3rd | lncRNA SBF2-AS1 | miR-151a-3p/XRCC4 | (1) Increasing the cancer cell proliferation (2) Inhibition of cancer cell apoptosis (3) Increasing the DNA damage repair | (1) Increasing the tumor size in a mouse model (2) Chemotherapy failure of GBM patients was associated with high lncRNA SBF2-AS1 expression | [137] |
| Glioblastoma | Temozolomide | LN229/TMZ | lncRNA TALC | ENO1/p38 MAPK/MEF2C/C5 | (1) Inhibition of cancer cell apoptosis (2) Increasing the DNA damage repair | (1) Increasing the tumor size in a mouse model (2) High expression of C5 induced by lncRNA TALC was associated with a poor prognosis in mice and GBM patients | [138] |
| Glioblastoma | Temozolomide | U251/TMZ | hsa_circ_0042003 | Heparanase | (1) Increasing the cancer cell proliferation (2) Inhibition of cancer cell apoptosis | (1) Increasing the tumor size in a mouse model (2) Chemotherapy failure of GBM patients was associated with high hsa_circ_0042003 expression | [139] |
| Glioblastoma | Temozolomide | A172/TMZ U251/TMZ | circ-HIPK3 | miR-421/ZIC5 | (1) Increasing the cancer cell proliferation and invasion (2) Inhibition of cancer cell apoptosis | (1) Increasing the tumor size in a mouse model (2) Chemotherapy failure of GBM patients was associated with high circ-HIPK3 expression | [140] |
| Type of Cancer | Anticancer Agent | EXO Cellular Source | EXO Cargo | Targeted Regulatory Network | Main In Vitro ncRNAs/EXO Effect | Main In Vivo ncRNAs/EXO Effect | Ref. |
|---|---|---|---|---|---|---|---|
| Lung cancer | 5-fluorouracil | A549 SPC-A1 | lncRNA FOXD3-AS1 | ELAVL1/PI3K/Akt | (1) Increasing the cancer cell proliferation and invasion (2) Inhibition of cancer cell apoptosis | ND * | [152] |
| Colorectal cancer | 5-fluorouracil | SW480/5-FU HCT116/5-FU | circ_0000338 | miR-217/miR-485-3p | (1) Increasing the cancer cell proliferation (2) Inhibition of cancer cell apoptosis | (1) Increasing the tumor size in a mouse mode (2) Chemotherapy failure of CRC patients was associated with high circ_0000338 expression | [153] |
| Pancreatic cancer | Gemcitabine | SW1990 BxPC-3 | circZNF91 | miR-23b-3p/SIRT1 | (1) Increasing the cancer cell proliferation (1) Increasing HIF-1α-dependent glycolysis in cancer cell | (1) Increasing the tumor size in a mouse mode (2) High expression of circZNF91was associated with a poor prognosis in PC patients | [154] |
| Type of Cancer | Anticancer Agent | EXO Cellular Source | EXO Cargo | Targeted Regulatory Network | Main In Vitro ncRNAs/EXO Effect | Main In Vivo ncRNAs/EXO Effect | Ref. |
|---|---|---|---|---|---|---|---|
| Ovarian cancer | Cisplatin | ND * | circRNA Cdr1as | miR-1270/SCAI | (1) Inhibiting the cancer cell proliferation and migration (2) Increasing the cancer cell apoptosis | (1) Decreasing the tumor size in a mouse model (2) Chemotherapy failure of OC patients was associated with low circRNA Cdr1as expression | [157] |
| Gastric cancer | Cisplatin | SGC-7901 MGC-803 | miR-107 | HMGA2/mTOR/P-gp | (1) Inhibiting the cancer cell proliferation | ND | [158] |
| Colorectal cancer | Oxaliplatin | FHC | circRNA FBXW7 | miR-18b-5p | (1) Inhibiting the cancer cell proliferation migration, and invasion (2) Inhibiting OXA efflux (3) Increasing the cancer cell apoptosis | (1) Decreasing the tumor size in a mouse model (2) Chemotherapy failure of OC patients was associated with low circRNA FBXW7 expression | [159] |
| Glioblastoma | Temozolomide | GBM cell lines | miR-151a | XRCC4 | (1) Inhibiting the cancer cell proliferation (2) Inhibiting the DNA damage repair (3) Increasing the cancer cell apoptosis | 1) Decreasing the tumor size in a mouse model (2) Low expression of miR-151a was associated with a poor prognosis in GBM patients | [160] |
| Tongue squamous cell carcinoma | Docetaxel | NTECs | miR-200c | TUBB3/PPP2R1B | (1) Inhibiting the cancer cell viability, migration, invasion, and motility (2) Inhibiting the DNA damage repair (3) Increasing the cancer cell apoptosis | (1) Decreasing the tumor size in a mouse model | [161] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Słomka, A.; Kornek, M.; Cho, W.C. Small Extracellular Vesicles and Their Involvement in Cancer Resistance: An Up-to-Date Review. Cells 2022, 11, 2913. https://doi.org/10.3390/cells11182913
Słomka A, Kornek M, Cho WC. Small Extracellular Vesicles and Their Involvement in Cancer Resistance: An Up-to-Date Review. Cells. 2022; 11(18):2913. https://doi.org/10.3390/cells11182913
Chicago/Turabian StyleSłomka, Artur, Miroslaw Kornek, and William C. Cho. 2022. "Small Extracellular Vesicles and Their Involvement in Cancer Resistance: An Up-to-Date Review" Cells 11, no. 18: 2913. https://doi.org/10.3390/cells11182913