
Rat Hepatic Stellate Cell Line CFSC-2G: Genetic Markers and Short Tandem Repeat Profile Useful for Cell Line Authentication
 , , and
, , and 
Abstract
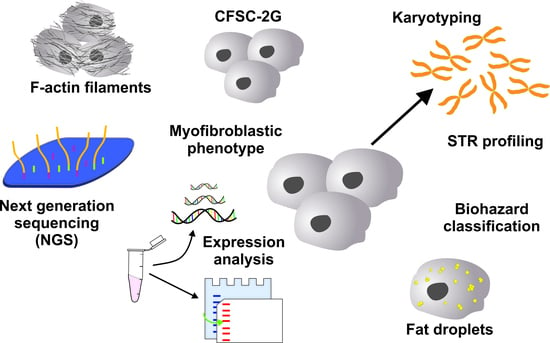
:
1. Introduction
2. Materials and Methods
2.1. Literature Search
2.2. Cell Culture
2.3. Electron Microscopic Analysis
2.4. Preparation of CFSC-2G Metaphase Chromosomes and Karyotyping
2.5. In Situ Hybridization and Spectral Imaging
2.6. Short Tandem Repeat (STR) Profiling
2.7. Next-Generation Sequencing and Data Analysis
2.8. Western Blot Analysis
2.9. Rhodamine–Phalloidin Stain
2.10. Lipid Droplet Staining
3. Results
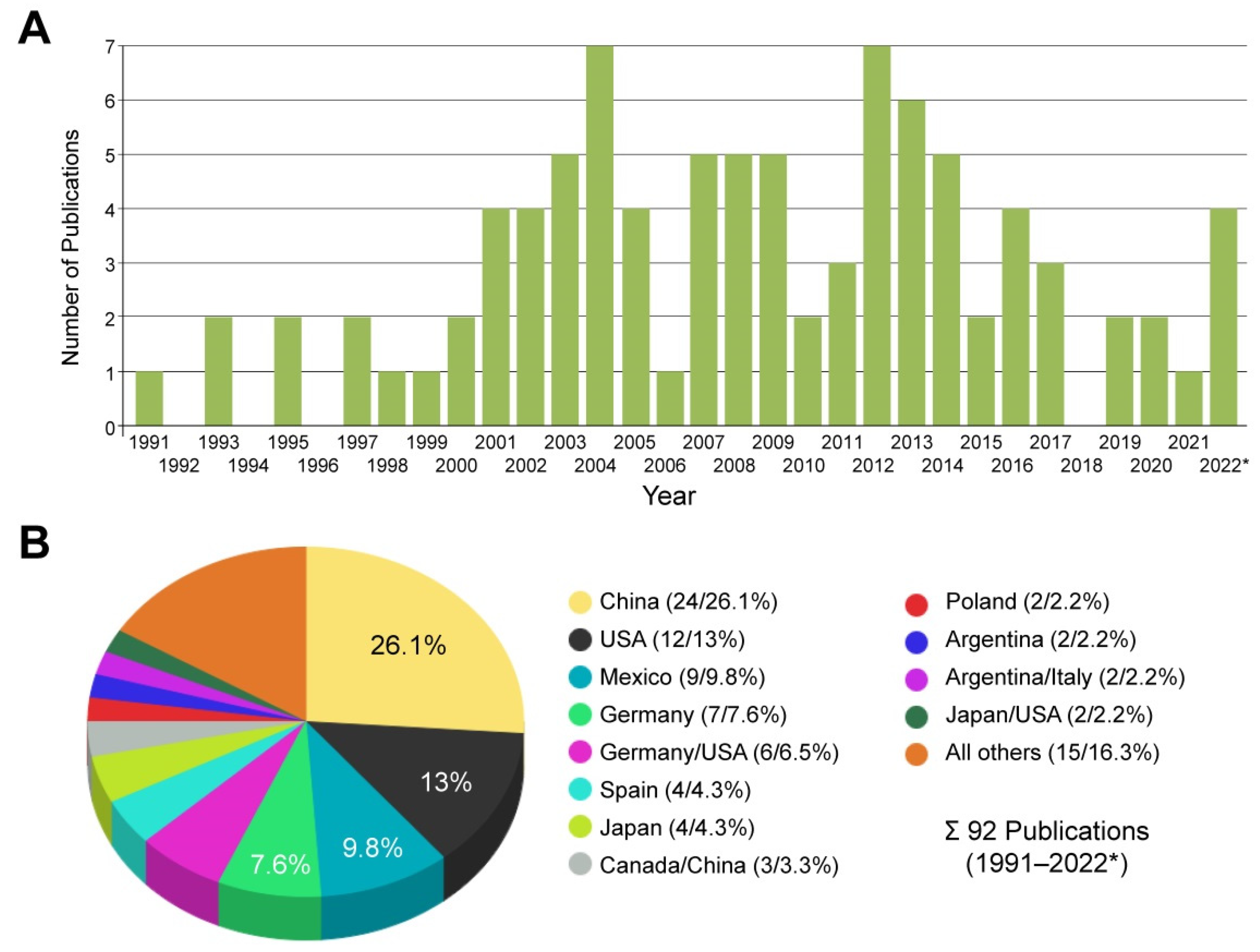
3.1. Usage of CFSC in Biomedical Research
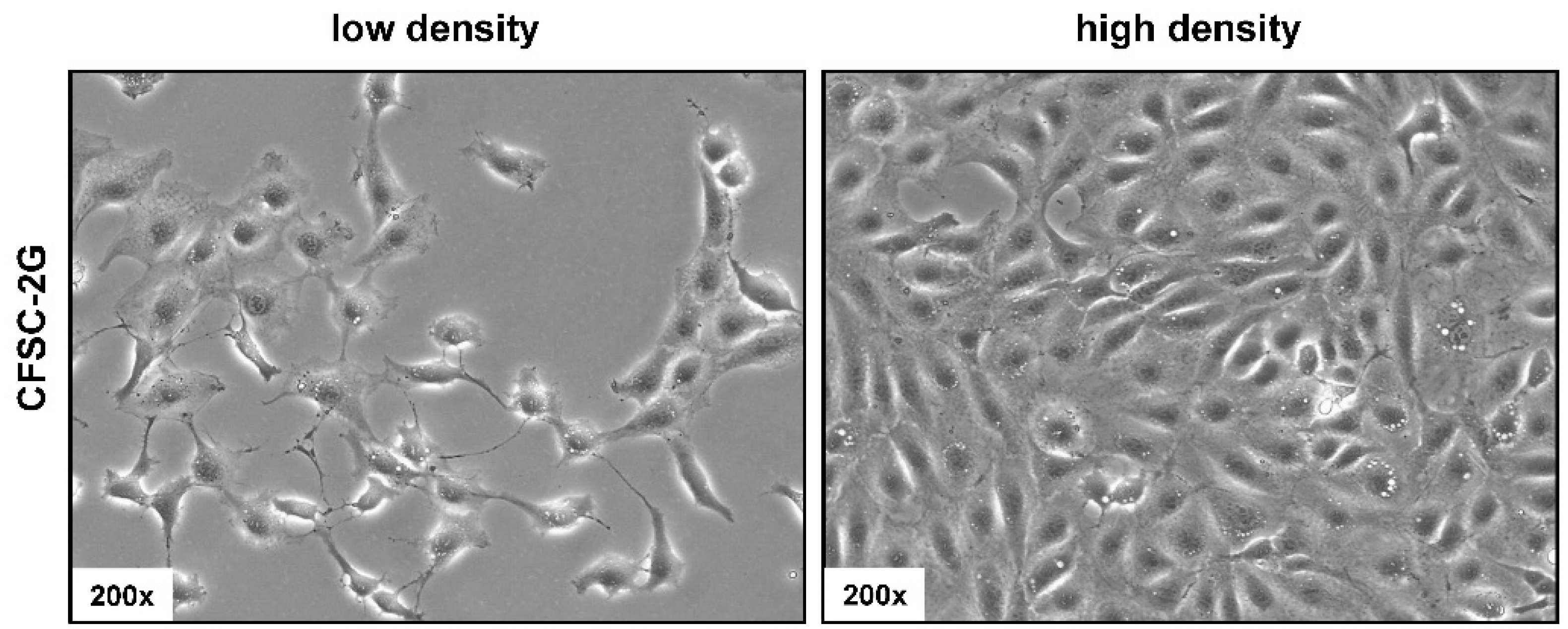
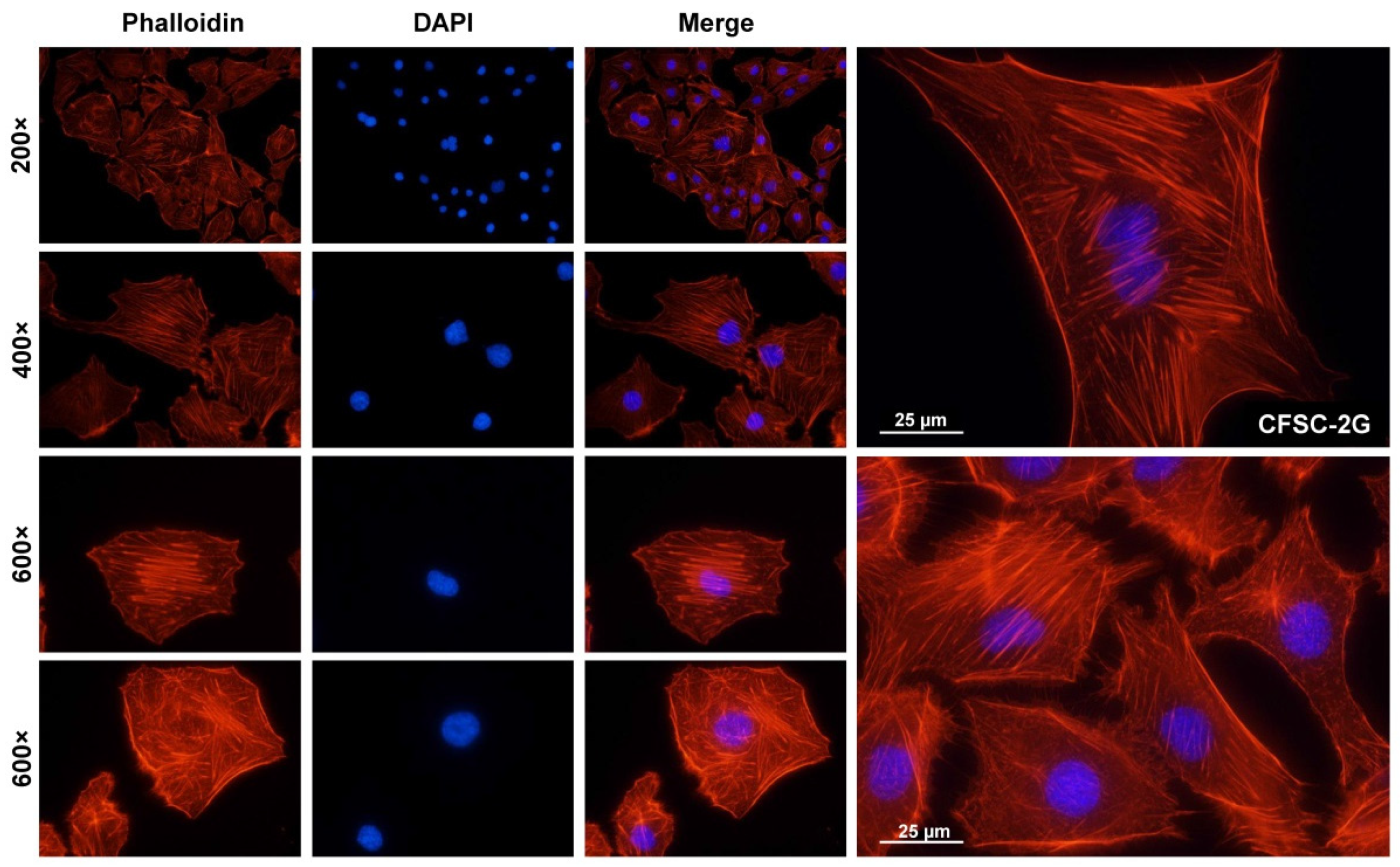
3.2. Phenotypic Characteristics of CFSC-2G Cells
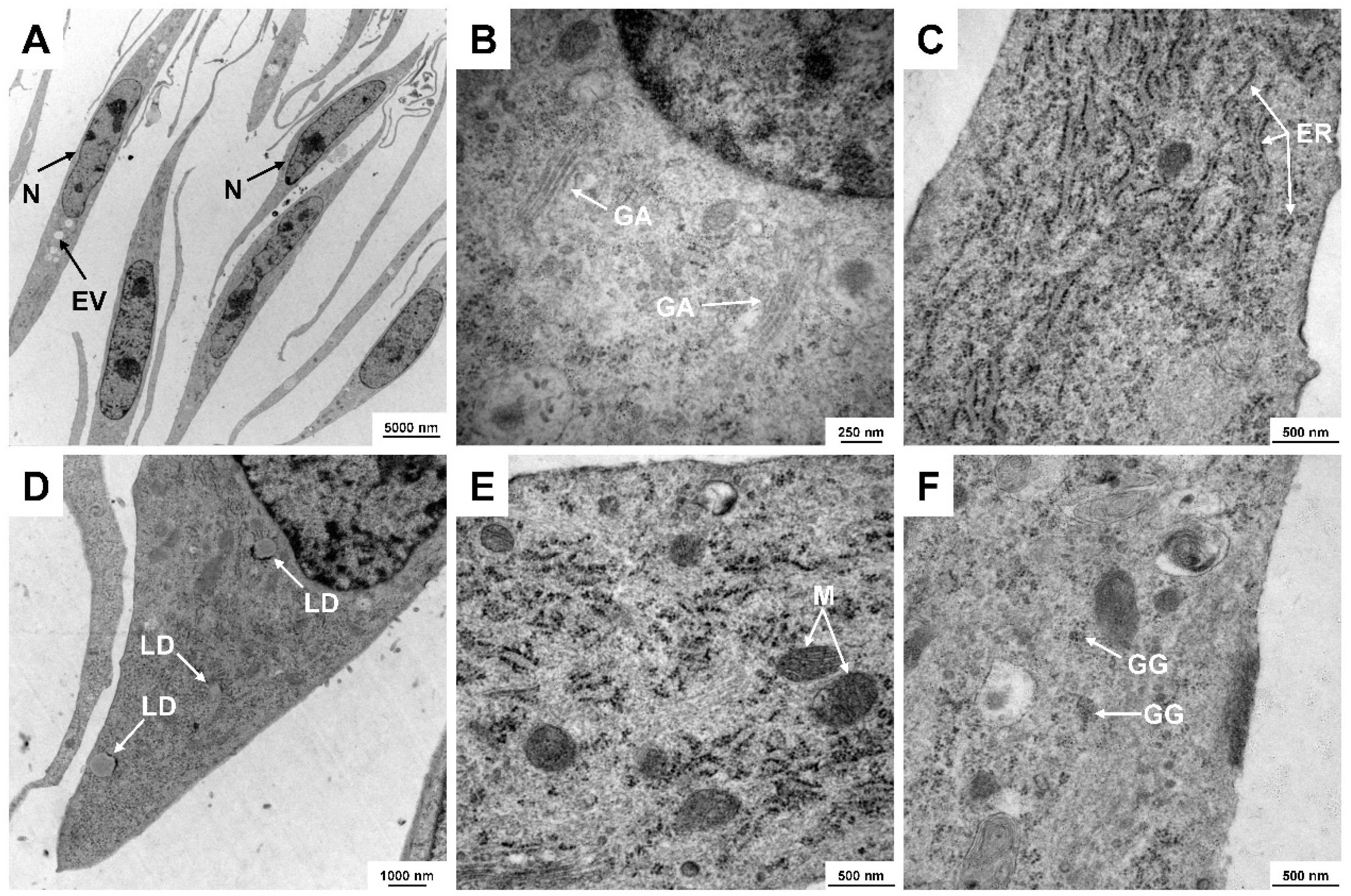
3.3. Ultrastructural Analysis of CFSC-2G
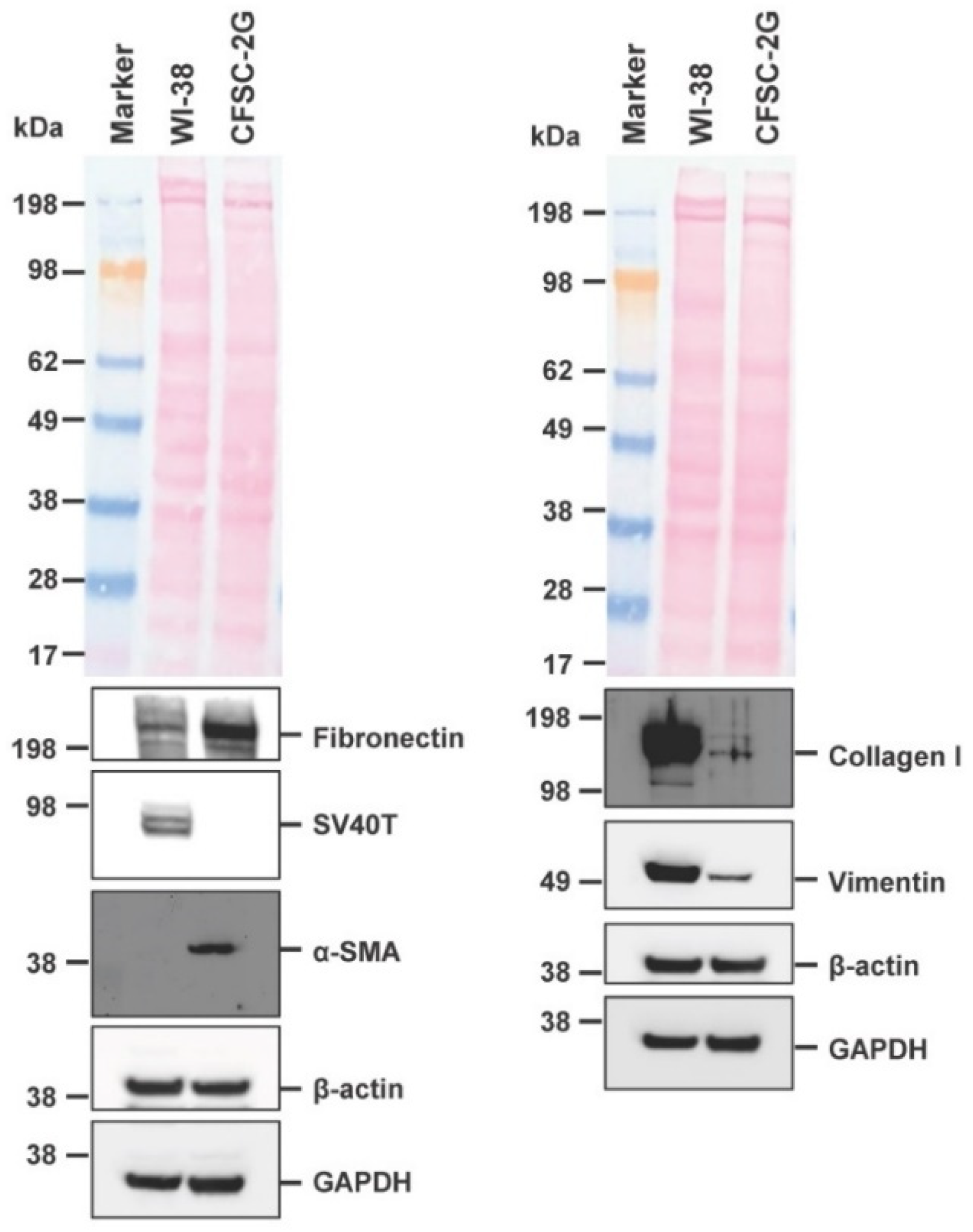
3.4. Expression of Typical Hepatic Stellate Cell Markers in CFSC-2G
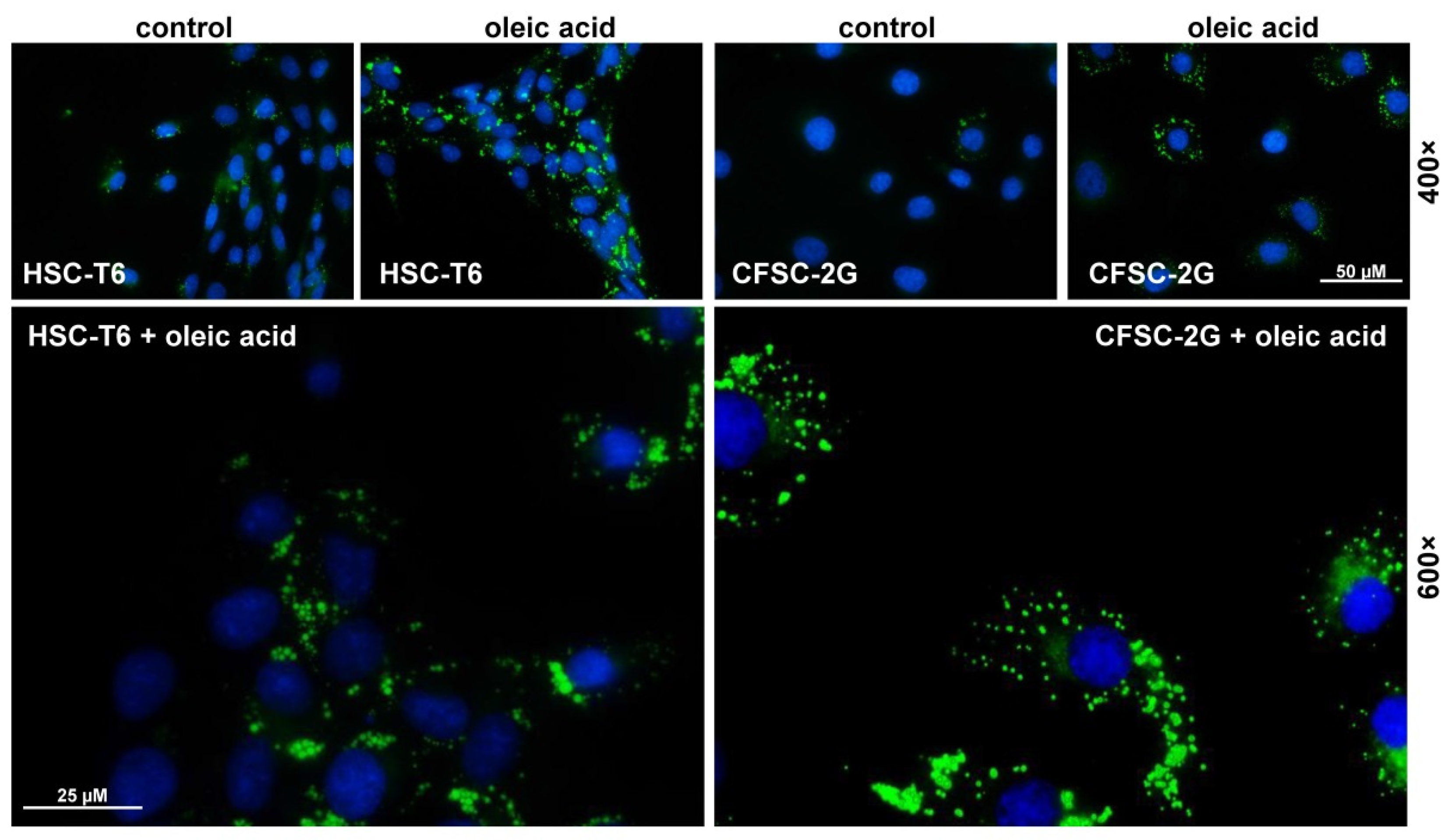
3.5. Fat Uptake and Storage in CFSC-2G
3.6. Genetic Analysis of CFSC-2G Cells
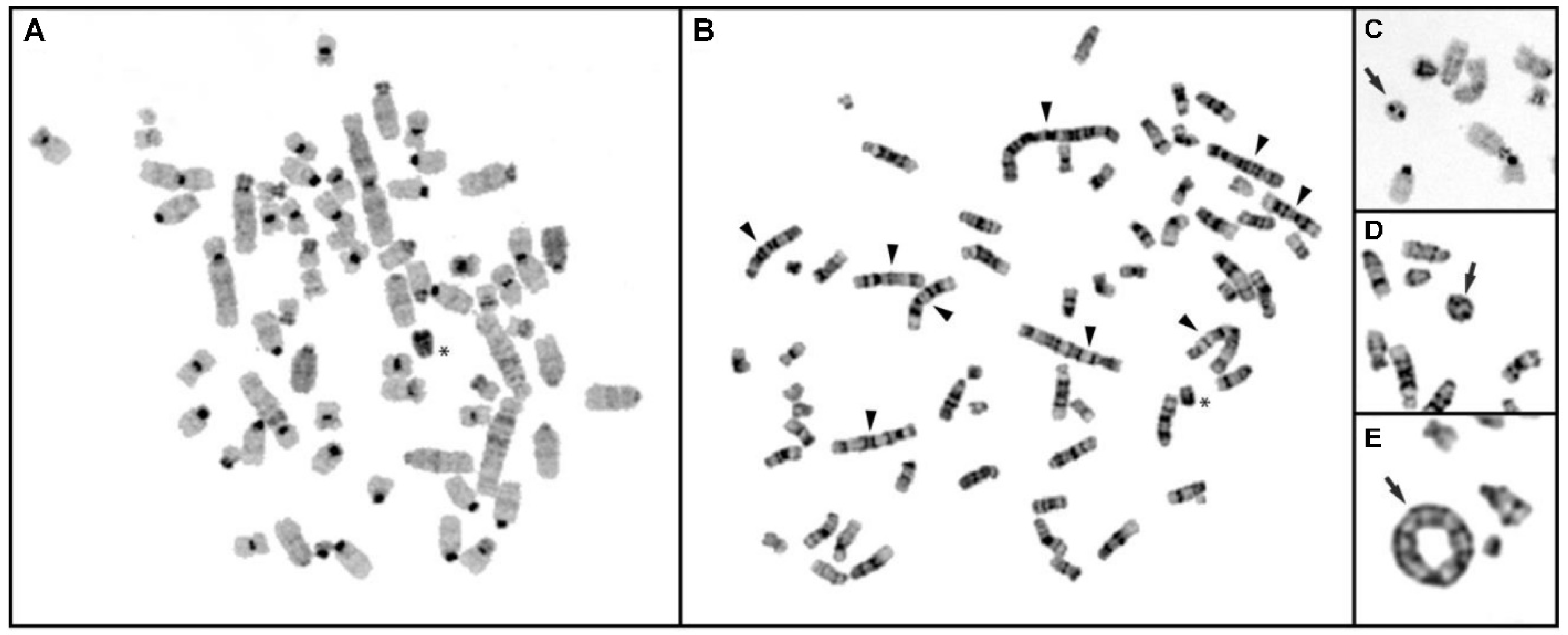
3.6.1. Conventional Cytogenetic Analysis of CFSC-2G Cells
3.6.2. In Situ Hybridization and Spectral Karyotype Analysis
3.6.3. Short Tandem Repeat Analysis
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Herrmann, J.; Gressner, A.M.; Weiskirchen, R. Immortal hepatic stellate cell lines: Useful tools to study hepatic stellate cell biology and function? J. Cell Mol. Med. 2007, 11, 704–722. [Google Scholar] [CrossRef] [PubMed]
- Krenkel, O.; Hundertmark, J.; Ritz, T.P.; Weiskirchen, R.; Tacke, F. Single cell RNA sequencing identifies subsets of hepatic stellate cells and myofibroblasts in liver fibrosis. Cells 2019, 8, 503. [Google Scholar] [CrossRef] [PubMed]
- Weiskirchen, R.; Weimer, J.; Meurer, S.K.; Kron, A.; Seipel, B.; Vater, I.; Arnold, N.; Siebert, R.; Xu, L.; Friedman, S.L.; et al. Genetic characteristics of the human hepatic stellate cell line LX-2. PLoS ONE 2013, 8, e75692. [Google Scholar] [CrossRef]
- Schröder, S.K.; Schüler, H.M.; Petersen, K.V.; Tesauro, C.; Knudsen, B.R.; Pedersen, F.S.; Krus, F.; Buhl, E.M.; Roeb, E.; Roderfeld, M.; et al. Genetic and molecular characterization of the immortalized murine hepatic stellate cell line GRX. Cells 2022, 11, 1504. [Google Scholar] [CrossRef] [PubMed]
- Meurer, S.K.; Brenner, D.A.; Weiskirchen, R. Multiplex short tandem repeat profiling of immortalized hepatic stellate cell line Col-GFP HSC. PLoS ONE 2022, 17, e0274219. [Google Scholar] [CrossRef]
- Nanda, I.; Steinlein, C.; Haaf, T.; Buhl, E.M.; Grimm, D.G.; Schröder, S.K.; Weiskirchen, R. Genetic characterization of rat hepatic stellate cell line HSC-T6 for in vitro cell line authentication. Cells 2022, 11, 1783. [Google Scholar] [CrossRef]
- Greenwel, P.; Schwartz, M.; Rosas, M.; Peyrol, S.; Grimaud, J.A.; Rojkind, M. Characterization of fat-storing cell lines derived from normal and CCl4-cirrhotic livers. Differences in the production of interleukin-6. Lab. Investig. 1991, 65, 644–653. [Google Scholar]
- Greenwel, P.; Rubin, J.; Schwartz, M.; Hertzberg, E.L.; Rojkind, M. Liver fat-storing cell clones obtained from a CCl4-cirrhotic rat are heterogeneous with regard to proliferation, expression of extracellular matrix components, interleukin-6, and connexin 43. Lab. Investig. 1993, 69, 210–216. [Google Scholar]
- National Library of Medicine. Available online: https://pubmed.ncbi.nlm.nih.gov/ (accessed on 11 May 2022).
- Vogel, S.; Piantedosi, R.; Frank, J.; Lalazar, A.; Rockey, D.C.; Friedman, S.L.; Blaner, W.S. An immortalized rat liver stellate cell line (HSC-T6): A new cell model for the study of retinoid metabolism in vitro. J. Lipid Res. 2000, 41, 882–893. [Google Scholar] [CrossRef]
- Sumner, A.T. A simple technique for demonstrating centromeric heterochromatin. Exp. Cell Res. 1972, 75, 304–306. [Google Scholar] [CrossRef]
- Buwe, A.; Steinlein, C.; Koehler, M.R.; Bar-Am, I.; Katzin, N.; Schmid, M. Multicolor spectral karyotyping of rat chromosomes. Cytogenet. Genome Res. 2003, 103, 163–168. [Google Scholar] [CrossRef] [PubMed]
- Schröder, S.K.; Asimakopoulou, A.; Tillmann, S.; Koschmieder, S.; Weiskirchen, R. TNF-α controls Lipocalin-2 expression in PC-3 prostate cancer cells. Cytokine 2020, 135, 155214. [Google Scholar] [CrossRef] [PubMed]
- Pérez-Tamayo, R. Liver Pathophysiology. In Therapies and Antioxidants, 1st ed.; Muriel, P., Ed.; Foreword: pp. xxix–xxx; Academic Press: Cambridge, MA, USA, 2017; ISBN 978-0-12-804274-8. [Google Scholar]
- Xu, L.; Hui, A.Y.; Albanis, E.; Arthur, M.J.; O’Byrne, S.M.; Blaner, W.S.; Mukherjee, P.; Friedman, S.L.; Eng, F.J. Human hepatic stellate cell lines, LX-1 and LX-2: New tools for analysis of hepatic fibrosis. Gut 2005, 54, 142–151. [Google Scholar] [CrossRef] [PubMed]
- Horie, S.; Kitamura, Y.; Kawasaki, H.; Terada, T. Inhibitory effects of antisense oligonucleotides on the expression of procollagen type III gene in mouse hepatic stellate cells transformed by simian virus 40. Pathol. Int. 2000, 50, 937–944. [Google Scholar] [CrossRef]
- Kitamura, Y.; Tanigawa, T.; Katsumoto, T.; Tomita, K.; Wang, H.R.; Hirai, K.; Ichihara, K.; Terada, T. Cell growth and differentiation of a novel mouse Ito (fat-storing) cell line transformed by a temperature-sensitive mutant of simian virus 40. Hepatology 1997, 26, 323–329. [Google Scholar] [CrossRef]
- Miura, N.; Kanayama, Y.; Nagai, W.; Hasegawa, T.; Seko, Y.; Kaji, T.; Naganuma, A. Characterization of an immortalized hepatic stellate cell line established from metallothionein-null mice. J. Toxicol. Sci. 2006, 31, 391–398. [Google Scholar] [CrossRef]
- Meurer, S.K.; Alsamman, M.; Sahin, H.; Wasmuth, H.E.; Kisseleva, T.; Brenner, D.A.; Trautwein, C.; Weiskirchen, R.; Scholten, D. Overexpression of endoglin modulates TGF-β1-signalling pathways in a novel immortalized mouse hepatic stellate cell line. PLoS ONE 2013, 8, e56116. [Google Scholar] [CrossRef] [PubMed]
- Greenspan, P.; Mayer, E.P.; Fowler, S.D. Nile red: A selective fluorescent stain for intracellular lipid droplets. J. Cell Biol. 1985, 100, 965–973. [Google Scholar] [CrossRef]
- Pakhomov, A.A.; Deyev, I.E.; Ratnikova, N.M.; Chumakov, S.P.; Mironiuk, V.B.; Kononevich, Y.N.; Muzafarov, A.M.; Martynov, V.I. BODIPY-based dye for no-wash live-cell staining and imaging. Biotechniques 2017, 63, 77–80. [Google Scholar] [CrossRef]
- Hong, Y.; Li, S.; Wang, J.; Li, Y. In vitro inhibition of hepatic stellate cell activation by the autophagy-related lipid droplet protein ATG2A. Sci. Rep. 2018, 8, 9232, Erratum in Sci. Rep. 2018, 8, 14569. [Google Scholar] [CrossRef]
- Ben-David, U.; Siranosian, B.; Ha, G.; Tang, H.; Oren, Y.; Hinohara, K.; Strathdee, C.A.; Dempster, J.; Lyons, N.J.; Burns, R.; et al. Genetic and transcriptional evolution alters cancer cell line drug response. Nature 2018, 560, 325–330. [Google Scholar] [CrossRef] [PubMed]
- Hazra, S.; Miyahara, T.; Rippe, R.A.; Tsukamoto, H. PPAR gamma and hepatic stellate cells. Comp. Hepatol. 2004, 3 (Suppl. 1). [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Khomich, O.; Ivanov, A.V.; Bartosch, B. Metabolic hallmarks of hepatic stellate cells in liver fibrosis. Cells 2019, 9, 24. [Google Scholar] [CrossRef]
- Horbach, S.P.J.M.; Halffman, W. The ghosts of HeLa: How cell line misidentification contaminates the scientific literature. PLoS ONE 2017, 12, e0186281. [Google Scholar] [CrossRef] [PubMed]
- Acharya, P.; Chouhan, K.; Weiskirchen, S.; Weiskirchen, R. Cellular mechanisms of liver fibrosis. Front. Pharmacol. 2021, 12, 671640. [Google Scholar] [CrossRef] [PubMed]
- Ye, Y.; Dan, Z. All-trans retinoic acid diminishes collagen production in a hepatic stellate cell line via suppression of active protein-1 and c-Jun N-terminal kinase signal. J. Huazhong Uni. Sci. Technol. Med. Sci. 2010, 30, 726–733. [Google Scholar] [CrossRef]
- Brennan, R.J.; Schiestl, R.H. Chloroform and carbon tetrachloride induce intrachromosomal recombination and oxidative free radicals in Saccharomyces cerevisiae. Mutat. Res. 1998, 397, 271–278. [Google Scholar] [CrossRef]
- Liu, J.; Li, H.; Nomura, K.; Dofuku, R.; Kitagawa, T. Cytogenetic analysis of hepatic cell lines derived from SV40-T antigen gene-harboring transgenic mice. Cancer Genet. Cytogenet. 1991, 55, 207–216. [Google Scholar] [CrossRef]
- Barros, B.; Morais, M.; Teixeira, A.L.; Medeiros, R. Loss of chromosome Y and its potential applications as biomarker in health and forensic sciences. Cytogenet. Genome Res. 2020, 160, 225–237. [Google Scholar] [CrossRef]
- Sano, S.; Horitani, K.; Ogawa, H.; Halvardson, J.; Chavkin, N.W.; Wang, Y.; Sano, M.; Mattisson, J.; Hata, A.; Danielsson, M.; et al. Hematopoietic loss of Y chromosome leads to cardiac fibrosis and heart failure mortality. Science 2022, 377, 292–297. [Google Scholar] [CrossRef]
- Forsberg, L.A.; Rasi, C.; Malmqvist, N.; Davies, H.; Pasupulati, S.; Pakalapati, G.; Sandgren, J.; Diaz de Ståhl, T.; Zaghlool, A.; Giedraitis, V.; et al. Mosaic loss of chromosome Y in peripheral blood is associated with shorter survival and higher risk of cancer. Nat. Genet. 2014, 46, 624–628. [Google Scholar] [CrossRef] [PubMed]
- Prokop, J.W.; Deschepper, C.F. Chromosome Y genetic variants: Impact in animal models and on human disease. Physiol. Genomics 2015, 47, 525–537. [Google Scholar] [CrossRef] [PubMed] [Green Version]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Antibody | Cat. No. | Company | Expected Size (kDa) | Dilution | Clonality |
|---|---|---|---|---|---|
| α-SMA | CBL171-I | Sigma-Aldrich, Taufkirchen, Germany | 45 | 1000 | r mAb |
| β-actin | A5441 | Sigma-Aldrich | 43 | 10,000 | m mAb |
| Collagen I | NB600-408 | Novus Biologicals, Wiesbaden, Germany | 139 | 1000 | r mAb |
| Fibronectin | AB1954 | Sigma-Aldrich | 262 | 3000 | r pAb |
| GAPDH (6C5) | sc-32233 | Santa Cruz Biotech., Santa Cruz, CA, USA | 37 | 1000 | m mAb |
| SV40T (v-300) | sc-20800 | Santa Cruz | 94 | 1000 | r pAb |
| Vimentin | ab92547 | Abcam, Berlin, Germany | 54 | 3000 | r mAb |
| g anti-rIgG (H + L), HRP | #31460 | Invitrogen, ThermoFisher, Dreieich, Germany | NA | 5000 | g pAb |
| g anti-mIgG (H + L), HRP | #31430 | Invitrogen | NA | 5000 | g pAb |
| Transcript Id | Gene Id | Gene | Gene Description | TPM (CFSC-2G/HSC-T6) | Remarks * |
|---|---|---|---|---|---|
| ENSRNOT00000083468.1 | ENSRNOG00000058039.1 | Acta2 | actin alpha 2, smooth muscle | 10.9245/2.377 | HSC marker |
| ENSRNOT00000005311.6 | ENSRNOG00000003897.6 | Col1a1 | collagen type I alpha 1 chain | 1408.42/2033.74 | HSC marker |
| ENSRNOT00000004956.4 | ENSRNOG00000003357.4 | Col3a1 | collagen type III alpha 1 chain | 64.4932/2128.9 | HSC marker |
| ENSRNOT00000024430.5 | ENSRNOG00000018087.5 | Vim | Vimentin | 3853.69/2054.81 | HSC marker |
| ENSRNOT00000057585.4 | ENSRNOG00000014288.8 | Fn1 | Fibronectin 1 | 1383.81/143.953 | HSC marker |
| ENSRNOT00000019772.6 | 702.835/352.585 | ||||
| ENSRNOT00000013538.6 | ENSRNOG00000009884.6 | Lgals1 | galectin 1 | 8557.92/4801.86 | HSC marker |
| ENSRNOT00000017486.7 | ENSRNOG00000012840.7 | Sparc | secreted protein acidic and cysteine rich | 1040.23/2188.34 | HSC marker |
| ENSRNOT00000067011.2 | ENSRNOG00000003772.7 | Csrp2 | cysteine and glycine-rich protein 2 | 62.7395/195.244 | HSC marker |
| ENSRNOT00000013745.7 | ENSRNOG00000010208.7 | Timp1 | TIMP metallopeptidase inhibitor | 714.162/5518.57 | inhibitor of MMPs |
| ENSRNOT00000004290.4 | ENSRNOG00000003148.5 | Timp2 | TIMP metallopeptidase inhibitor 2 | 1030.84/369.667 | inhibitor of MMPs |
| ENSRNOT00000010180.5 | ENSRNOG00000007650.5 | Cd63 | Cd63 molecule | 2732.07/1639.48 | TIMP1 receptor |
| ENSRNOT00000011208.7 | ENSRNOG00000008301.7 | Tagln2 | transgelin 2 | 642.605/754.93 | smooth muscle marker |
| ENSRNOT00000015962.6 | ENSRNOG00000020660.7 | Cfl1 | cofilin 1 | 1961.22/1670.37 | cytoskeleton |
| ENSRNOT00000042459.4 | ENSRNOG00000034254.4 | Actb | actin, beta | 3869.05/4379.5 | cytoskeleton |
| ENSRNOT00000080216.1 | 1503.75/1820.7 | ||||
| ENSRNOT00000078250.1 | ENSRNOG00000056836.1 | Cav1 | caveolin 1 | 486.234/167.988 | scaffolding protein |
| ENSRNOT00000008659.4 | ENSRNOG00000009972.7 | Rara | retinoic acid receptor, alpha | 26.5339/24.8697 | nuclear receptor |
| ENSRNOT00000084644.1 | 20.4423/23.8544 | ||||
| ENSRNOT00000033048.6 | ENSRNOG00000024061.7 | Rarb | retinoic acid receptor, beta | 0.00872314/0.541197 | nuclear receptor |
| ENSRNOT00000016801.5 | ENSRNOG00000012499.7 | Rarg | retinoic acid receptor, gamma | 49.0507/64.5919 | nuclear receptor |
| ENSRNOT00000017096.7 | 25.6328/39.1824 | ||||
| ENSRNOT00000012892.4 | ENSRNOG00000009446.4 | Rxra | retinoid X receptor alpha | 41.2102/18.9052 | nuclear receptor |
| ENSRNOT00000041613.5 | ENSRNOG00000000464.7 | Rxrb | retinoid X receptor beta | 15.9738/20.0384 | nuclear receptor |
| ENSRNOT00000087670.1 | 1.53037/1.79029 | ||||
| ENSRNOT00000086978.1 | 1.02833/1.08069 | ||||
| ENSRNOT00000081588.1 | 1.61434/0.804912 | ||||
| ENSRNOT00000091182.1 | 0.347194/0.723898 | ||||
| ENSRNOT00000087895.1 | 0.0953106/0.688452 | ||||
| ENSRNOT00000084638.1 | 0.615142/0.499241 | ||||
| ENSRNOT00000012137.5 ENSRNOT00000051858.5 ENSRNOT00000082969.1 | ENSRNOG00000008839.7 | Pparg | peroxisome proliferation-activated receptor-γ | 0.883663/0.157304 0.386135/0.248569 0/6.50011 | nuclear receptor |
| ENSRNOT00000077227.1 | ENSRNOG00000004537.6 | Rxrg | retinoid X receptor gamma | 0/0 | nuclear receptor |
| ENSRNOT00000006117.5 | 0.271271/0 | ||||
| ENSRNOT00000019571.3 | ENSRNOG00000014090.3 | Retsat | retinol saturase | 58.3177/52.476 | retinoid metabolism |
| ENSRNOT00000081756.1 | ENSRNOG00000053850.1 | Rdh5 | retinol dehydrogenase 5 | 0.693011/0.0383589 | retinoid metabolism |
| ENSRNOT00000032076.4 | ENSRNOG00000025767.4 | Rdh8 | retinol dehydrogenase 8 | 0.0525309/0.112568 | retinoid metabolism |
| ENSRNOT00000009096.4 | ENSRNOG00000006681.4 | Rdh10 | retinol dehydrogenase 10 | 21.0453/3.87175 | retinoid metabolism |
| ENSRNOT00000085927.1 | ENSRNOG00000054770.1 | Rdh11 | retinol dehydrogenase 11 | 35.9222/26.8725 | retinoid metabolism |
| ENSRNOT00000078436.1 | 0.245147/0.15904 | ||||
| ENSRNOT00000089162.1 | ENSRNOG00000056553.1 | Rdh12 | retinol dehydrogenase 12 | 0/1.06243 | retinoid metabolism |
| ENSRNOT00000031462.5 | ENSRNOG00000027919.5 | Rdh13 | retinol dehydrogenase 13 | 1.19281/1.96789 | retinoid metabolism |
| ENSRNOT00000006020.6 | ENSRNOG00000039551.3 | Rdh14 | retinol dehydrogenase 14 | 19.6884/8.16116 | retinoid metabolism |
| ENSRNOT00000093003.1 | 5.89539/2.97058 | ||||
| ENSRNOT00000092948.1 | 1.27751/0.857514 | ||||
| ENSRNOT00000018622.4 | ENSRNOG00000013794.4 | Rbp1 | retinol-binding protein 1 | 0/25.3842 | retinoid metabolism |
| ENSRNOT00000018755.6 | ENSRNOG00000013932.6 | Rbp2 | retinol-binding protein 2 | 1.64899/0.309974 | retinoid metabolism |
| ENSRNOT00000021055.7 | ENSRNOG00000015518.7 | Rbp4 | retinol-binding protein 4 | 0.0677519/0.0363529 | retinoid metabolism |
| ENSRNOT00000021348.5 | ENSRNOG00000015850.5 | Rbp7 | retinol-binding protein 7 | 0.398306/0.244971 | retinoid metabolism |
| ENSRNOT00000082156.1 | ENSRNOG00000025608.4 | Lrat | lecithin retinol acyltransferase | 0.0883818/0.0125274 | retinoid metabolism |
| ENSRNOT00000022113.4 | ENSRNOG00000016275.4 | Ttr | transthyretin | 0.049935/0.161195 | RBP transporter |
| ENSRNOT00000030919.5 | ENSRNOG00000022619.6 | Fth1 | ferritin heavy chain 1 | 5047.69/9890.55 | iron storage |
| ENSRNOT00000087162.1 | ENSRNOG00000059900.1 | Bst2 | bone marrow stromal cell antigen 2 | 291.2/1681.66 | antiviral response |
| ENSRNOT00000091906.1 | 127.083/589.283 | ||||
| ENSRNOT00000080988.1 | ENSRNOG00000052802.1 | Aldoa | Aldolase, fructose-bisphosphate | 1914.86/1471.21 | glycolysis |
| ENSRNOT00000087928.1 | 745.559/492.926 | ||||
| ENSRNOT00000088473.1 | 308.32/183.94 | ||||
| ENSRNOT00000015332.7 | ENSRNOG00000011329.7 | Pkm | pyruvate kinase M1/2 | 1952.24/1437.52 | glycolysis |
| ENSRNOT00000083666.1 | 596.921/455.466 | ||||
| ENSRNOT00000077604.1 | ENSRNOG00000058249.1 | Pgk1 | Phosphoglycerate kinase 1 | 2609.96/1404.31 | glycolysis |
| ENSRNOT00000085653.1 | 0/0.204793 | ||||
| ENSRNOT00000050443.4 | ENSRNOG00000018630.7 | LOC108351137 | Glyceraldehyde-3-phosphate dehydrogenase | 3223.93/4598.86 | glycolysis |
| ENSRNOT00000041328.3 | ENSRNOG00000030963.3 | 2962.7/4243.2 |
| Cell | Chromosome Numbers | der(1)t(1;11) | der(1)t(1;12) | der(1)t(1;14) | der(1)t(1;16) | der(1)t(1;20) | rob(2;2) | der(2)t(2;5) | der(2)t(2;6) | der(2)t(2;12) | der(2)t(2;15) | der(2)t(2;17) | der(3)t(3;7) | der(4)t(4;12) | der(4)t(4;12;16 | der(5)t(5;Y) | der(6)t(6;19) | der(6)t(7;6;19) | der(7)t(7;10) | der(8)t(8;15) | der(9)t(9;12) | der(10)t(10;Y) | der(10)t(10;15) | der(10)t(10;16) | der(12)t(12;19) | der(12(t(12;20) | der(13)t(13;18) | der(14)t(14;16) | der(15)t(15;19) | der(17)t(17:19) | der(18)t(18;20) | der(19)t(19;20) |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 1 | 63, XXY | ● | ● | ● | ● | ● | ● | ● | ● | ● | ● | ● | ● | ● | ● | ● | ● | |||||||||||||||
| 2 | 62, XXY | ● | ● | ● | ● | ● | ● | ● | ● | ● | ● | ● | ● | ● | ● | |||||||||||||||||
| 3 | 62, XXY | ● | ● | ● | ● | ● | ● | ● | ● | ● | ● | ● | ● | ● | ● | |||||||||||||||||
| 4 | 62, XXY | ● | ● | ● | ● | ● | ● | ● | ● | ● | ● | ● | ● | ● | ● | ● | ● | |||||||||||||||
| 5 | 58, XXY | ● | ● | ● | ● | ● | ● | ● | ● | ● | ● | ● | ||||||||||||||||||||
| 6 | 62, XXYY | ● | ● | ● | ● | ● | ● | ● | ● | ● | ● | ● | ● | ● | ||||||||||||||||||
| 7 | 57, XX | ● | ● | ● | ● | ● | ● | ● | ● | ● | ● | ● | ||||||||||||||||||||
| 8 | 65, XXY | ● | ● | ● | ● | ● | ● | ● | ● | ● | ● | ● | ● | ● | ● | ● | ||||||||||||||||
| 9 | 63, XXY | ● | ● | ● | ● | ● | ● | ● | ● | ● | ● | ● | ● | ● | ● | |||||||||||||||||
| 10 | 64, XXY | ● | ● | ● | ● | ● | ● | ● | ● | ● | ● | ● | ● | ● | ● | |||||||||||||||||
| 11 | 64, XXY | ● | ● | ● | ● | ● | ● | ● | ● | ● | ● | ● | ● | ● | ● |
| SN | Marker Name | Chromosomal Location | Allele Sizes (bp) in CFSC-2G | Allele Sizes (bp) in HSC-T6 * |
|---|---|---|---|---|
| 1 | 73 | 1 | 194, 203 | 194 |
| 2 | 8 | 2 | 236 | 234 |
| 3 | 2 | 2 | 126 | 127 |
| 4 | 4 | 3 | 268, 270 | 238 |
| 5 | 3 | 3 | 160, 182 | 160, 162 |
| 6 | 26 | 4 | 150 | 166 |
| 7 | 19 | 4 | 180 | 175 |
| 8 | 81 | 5 | 130, 134 | 130, 132 |
| 9 | 34 | 6 | 184, 189 | 188 |
| 10 | 30 | 7 | 188, 192 | 186, 192 |
| 11 | 24 | 8 | 260 | 249, 253 |
| 12 | 59 | 9 | 145 | 143, 146, 180 |
| 13 | 62 | 9 | 166 | 177 |
| 14 | 1 | 10 | 105 | 96 |
| 15 | 55 | 10 | 210, 214 | 210, 218 |
| 16 | 36 | 11 | 222 | 234 |
| 17 | 67 | 11 | 154, 156 | 165 |
| 18 | 13 | 12 | 121 | 121, 135 |
| 19 | 35 | 13 | 197 | 197, 203 |
| 20 | 42 | 13 | 125 | 127 |
| 21 | 70 | 14 | 158, 175 | 175, 179 |
| 22 | 61 | 15 | 128 | 128 |
| 23 | 79 | 15 | 172, 180 | 172 |
| 24 | 90 | 16 | 159, 161 | 174 |
| 25 | 69 | 16 | 138 | 139 |
| 26 | 78 | 17 | 136, 151 | 147, 151 |
| 27 | 15 | 18 | 232 | 232 |
| 28 | 16 | 18 | 251, 260 | 247, 251 |
| 29 | 75 | 19 | 144 | 144, 184 |
| 30 | 96 | 20 | 210 | 210, 212 |
| 31 | 91 | 20 | 221 | 205, 211 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Nanda, I.; Schröder, S.K.; Steinlein, C.; Haaf, T.; Buhl, E.M.; Grimm, D.G.; Weiskirchen, R. Rat Hepatic Stellate Cell Line CFSC-2G: Genetic Markers and Short Tandem Repeat Profile Useful for Cell Line Authentication. Cells 2022, 11, 2900. https://doi.org/10.3390/cells11182900
Nanda I, Schröder SK, Steinlein C, Haaf T, Buhl EM, Grimm DG, Weiskirchen R. Rat Hepatic Stellate Cell Line CFSC-2G: Genetic Markers and Short Tandem Repeat Profile Useful for Cell Line Authentication. Cells. 2022; 11(18):2900. https://doi.org/10.3390/cells11182900
Chicago/Turabian StyleNanda, Indrajit, Sarah K. Schröder, Claus Steinlein, Thomas Haaf, Eva M. Buhl, Domink G. Grimm, and Ralf Weiskirchen. 2022. "Rat Hepatic Stellate Cell Line CFSC-2G: Genetic Markers and Short Tandem Repeat Profile Useful for Cell Line Authentication" Cells 11, no. 18: 2900. https://doi.org/10.3390/cells11182900