

Extracellular HMGB1 as Inflammatory Mediator in the Progression of Mycoplasma Gallisepticum Infection
 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Mycoplasma Strains
2.2. HD-11 Cells Culture and Treatment
2.3. Mycoplasma Gallisepticum Quantification
2.4. Gene Overexpression and Knockdown Assays
2.5. Immunofluorescent Staining
2.6. Protein Extraction and Western Blot
2.7. ELISA
2.8. RNA Isolation and Quantitative Real-Time PCR
2.9. Experimental Animals and Treatments
2.10. Histopathological Analysis of Spleen
2.11. Terminal Deoxynucleotidyl Transferase—Mediated dUTP Nick Endlabeling Assay
2.12. Statistics
3. Results
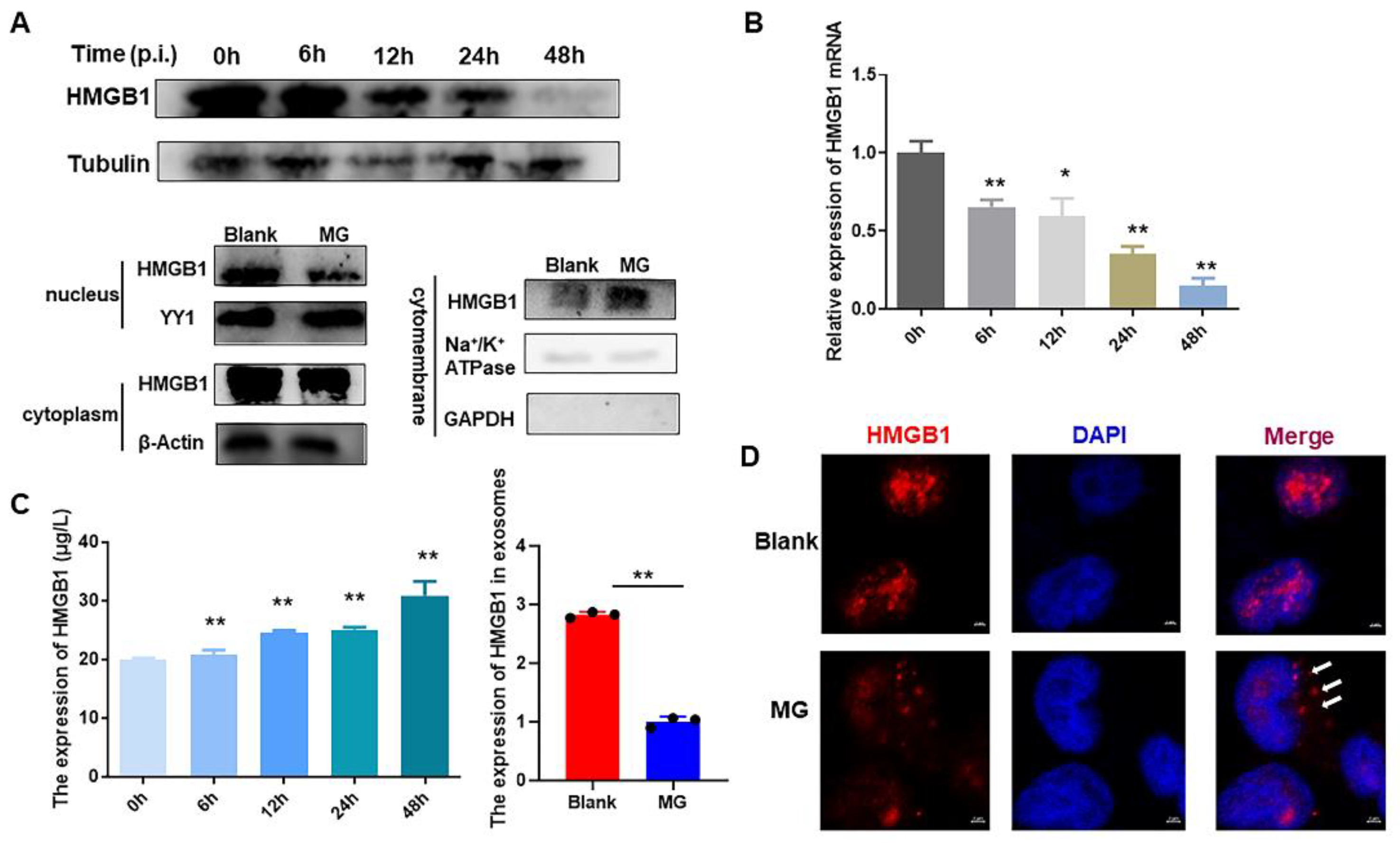
3.1. MG Infection Induces the Secretion of HMGB1
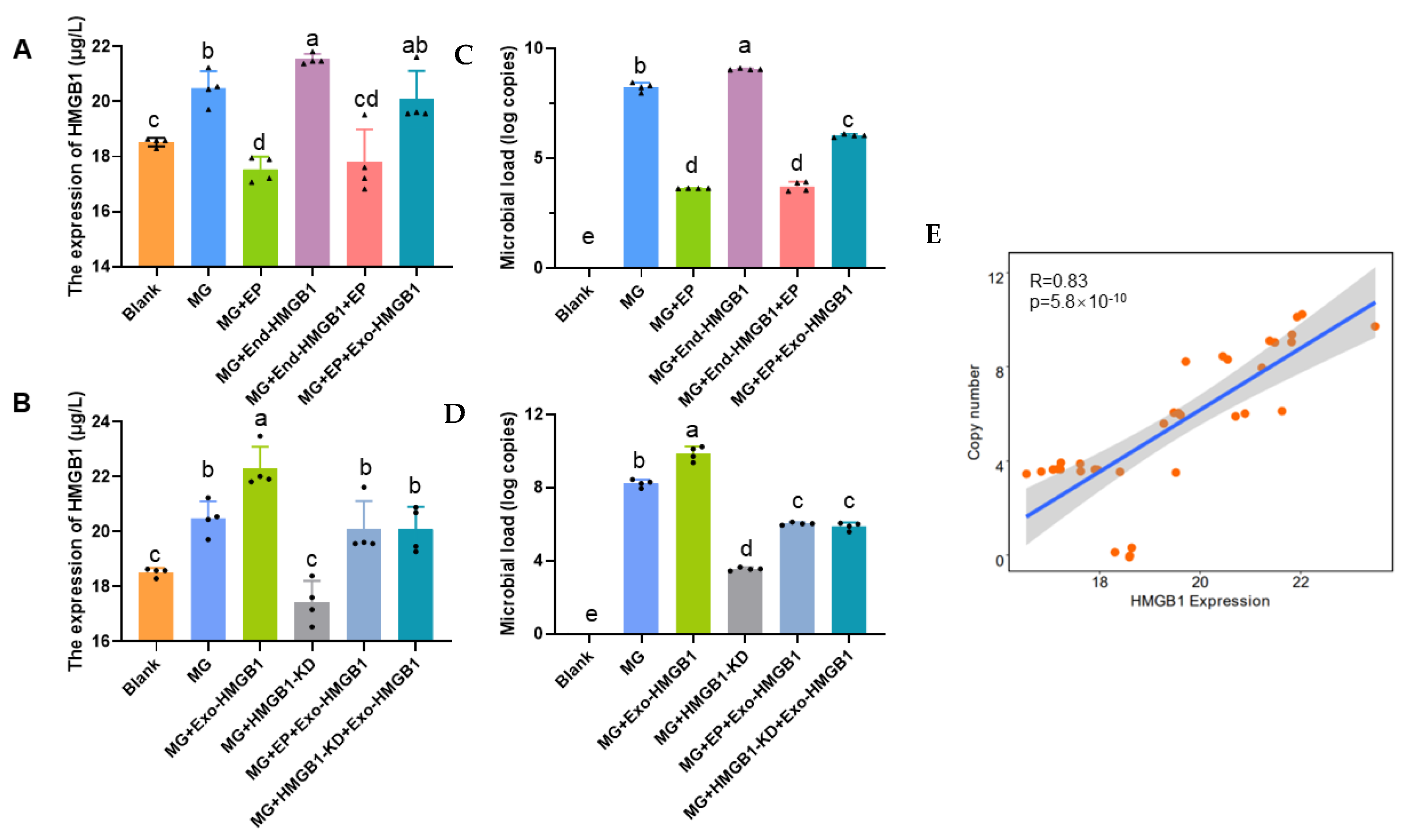
3.2. Extracellular HMGB1 Affects MG Replication
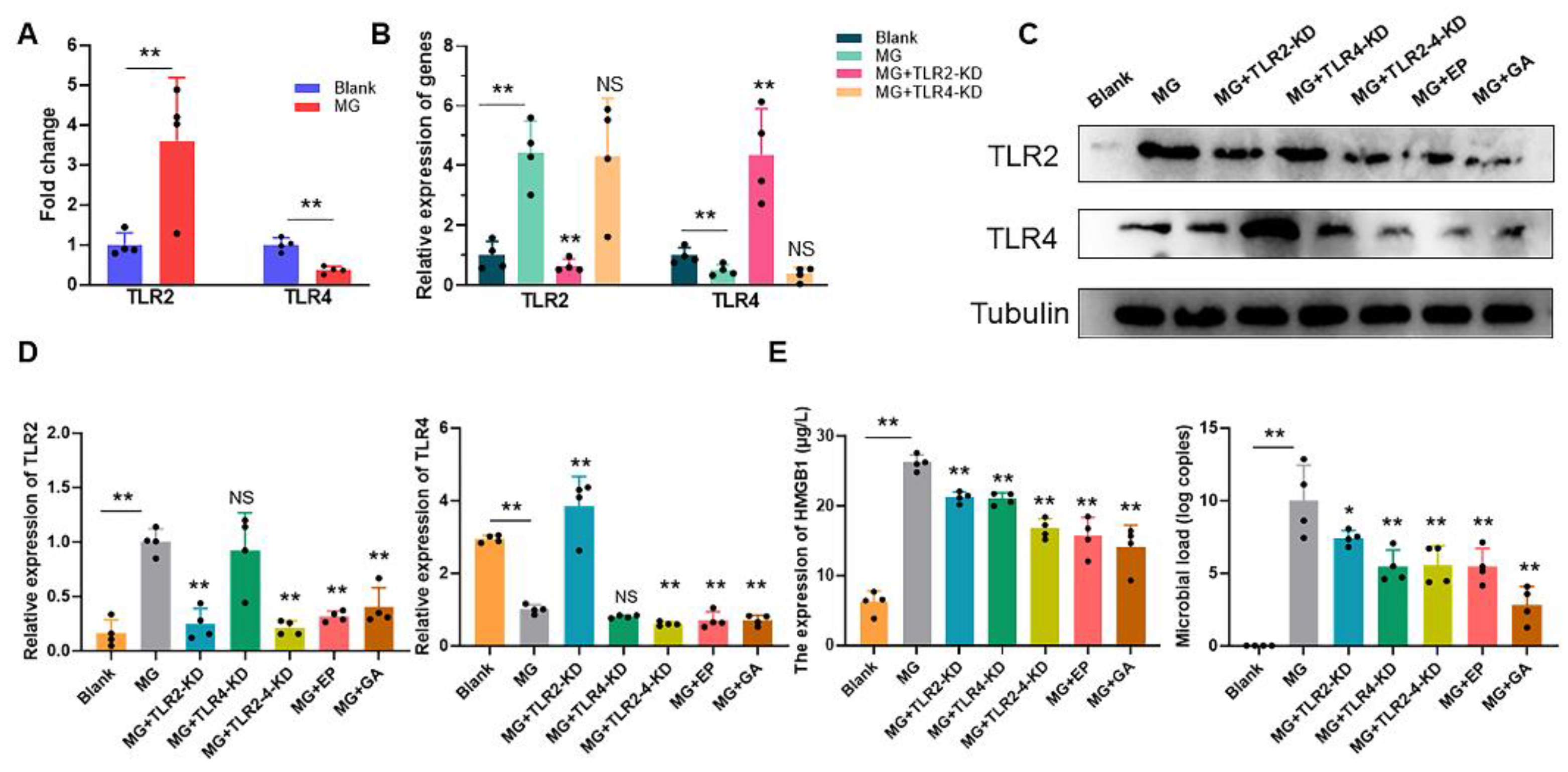
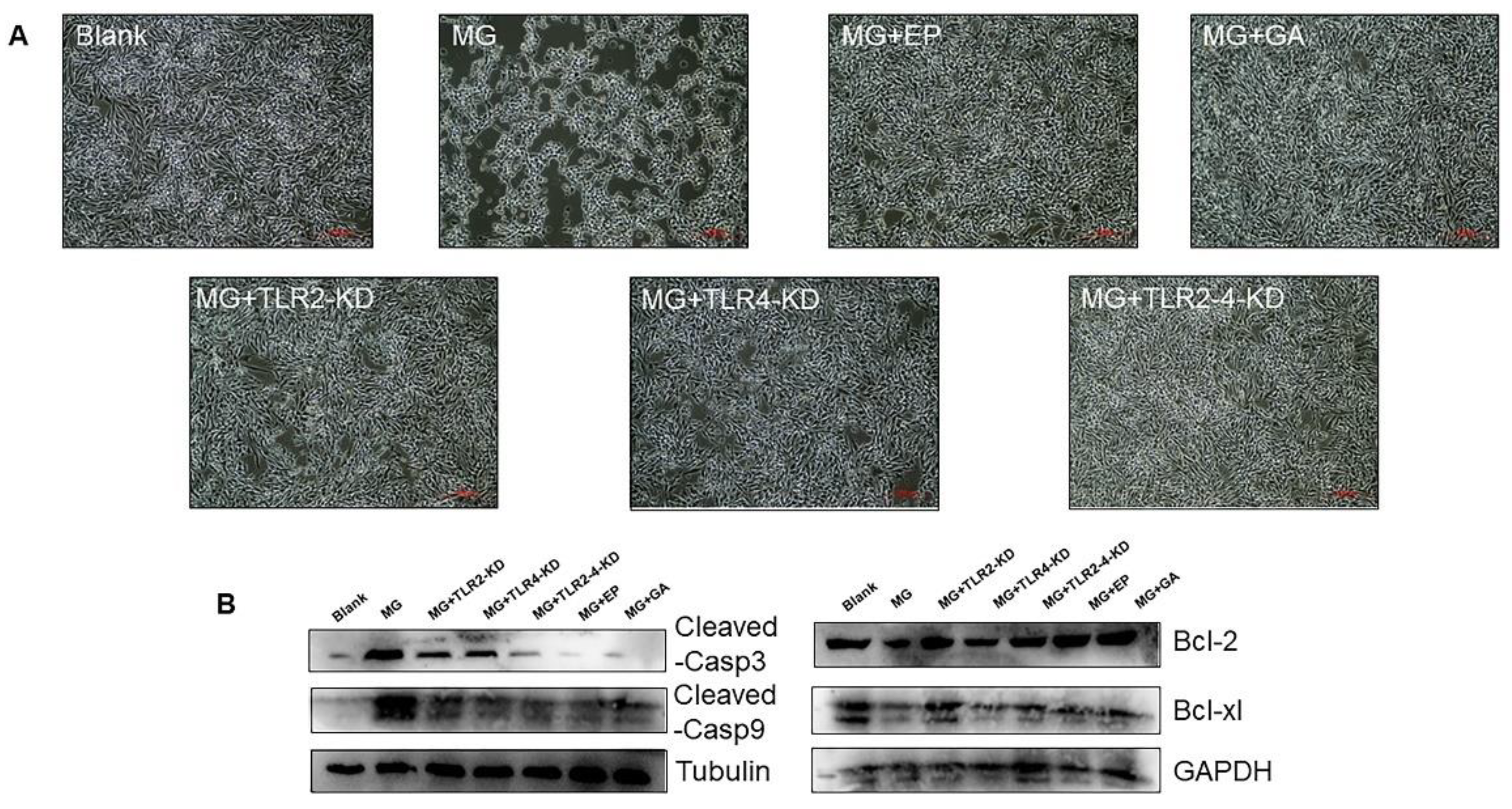
3.3. Extracellular HMGB1 Activates TLR2/4 to Promote MG Proliferation
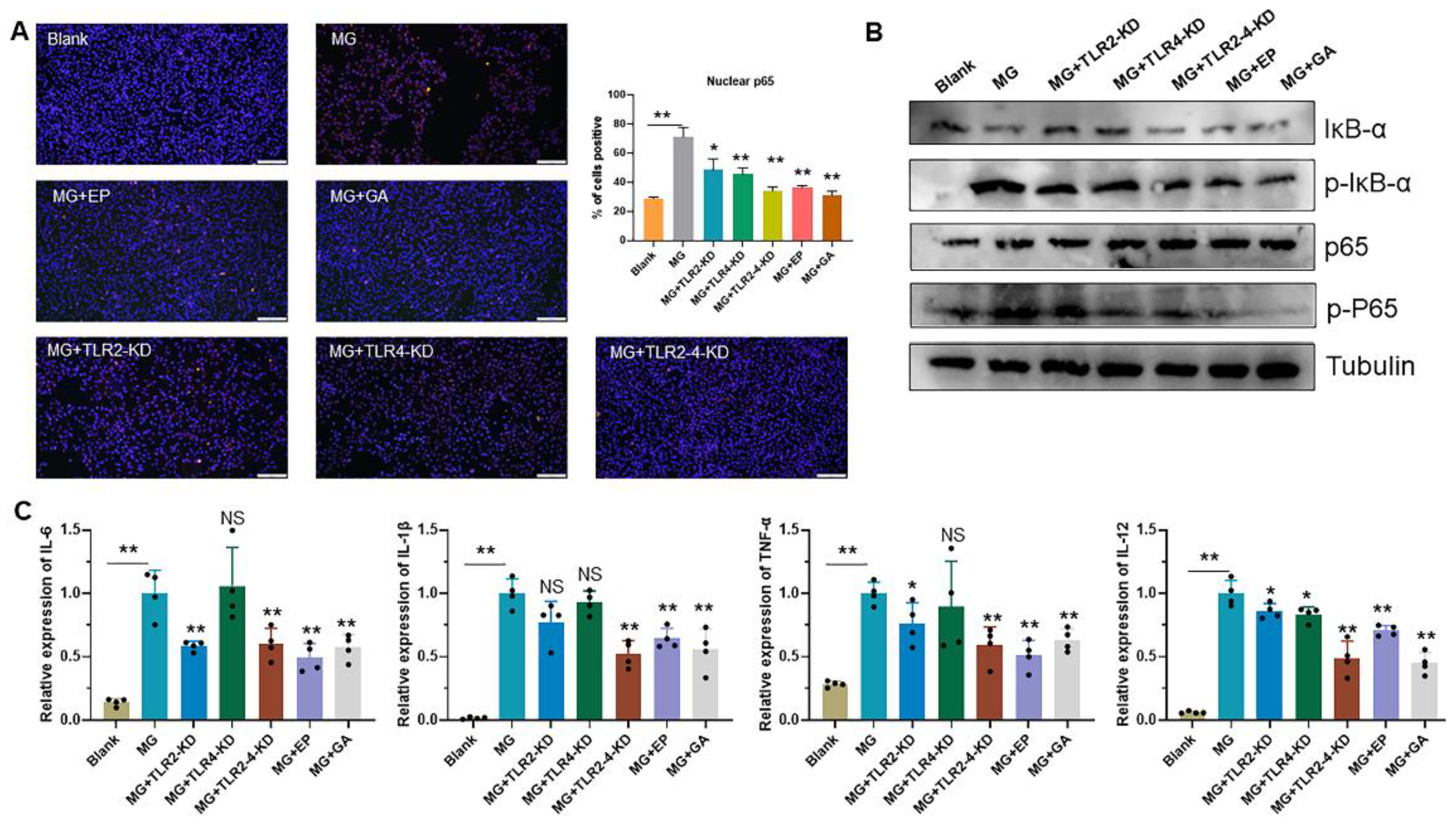
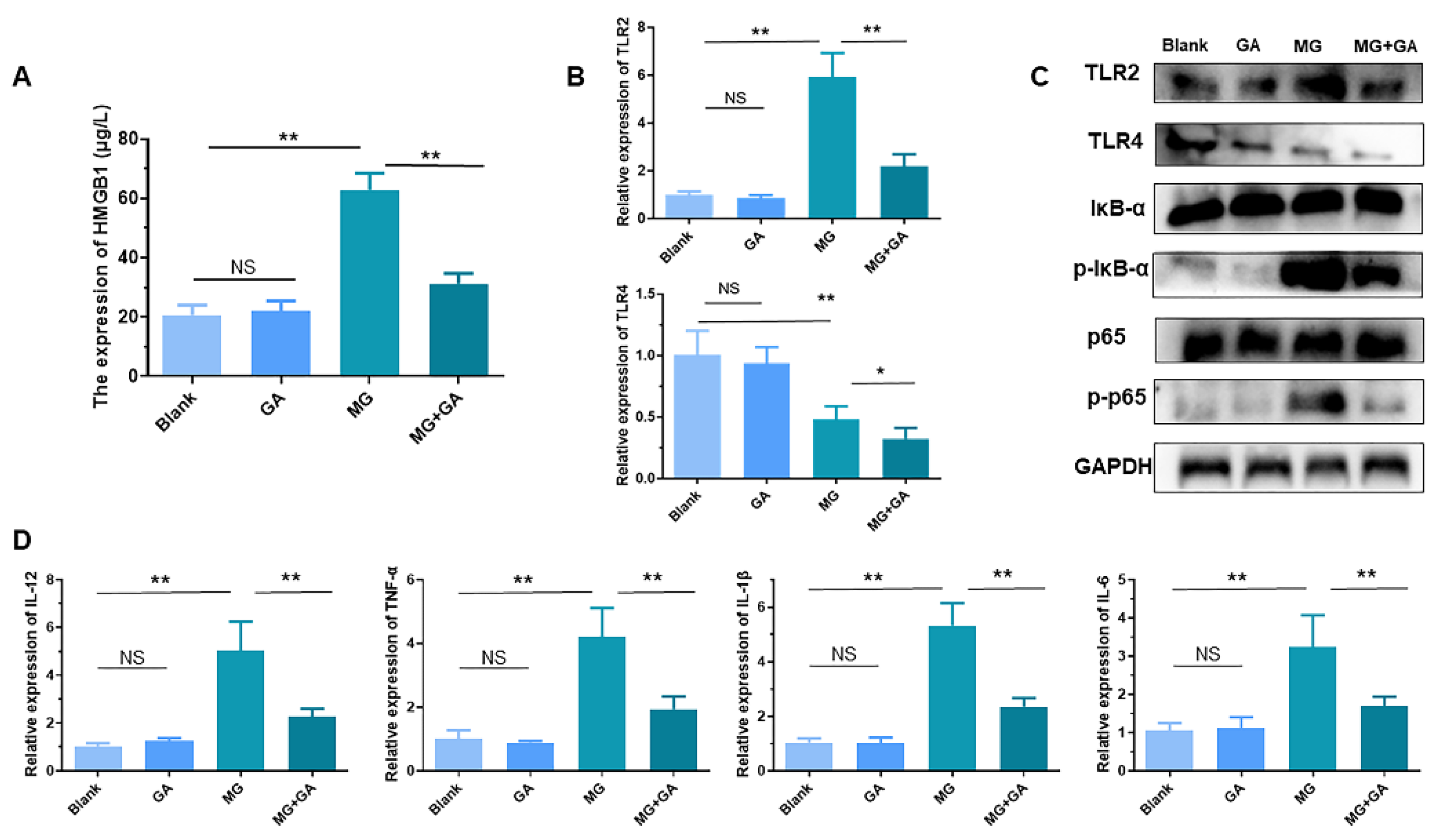
3.4. HMGB1-Activated TLR2-4 Triggers Immune Disorders via NF-κB
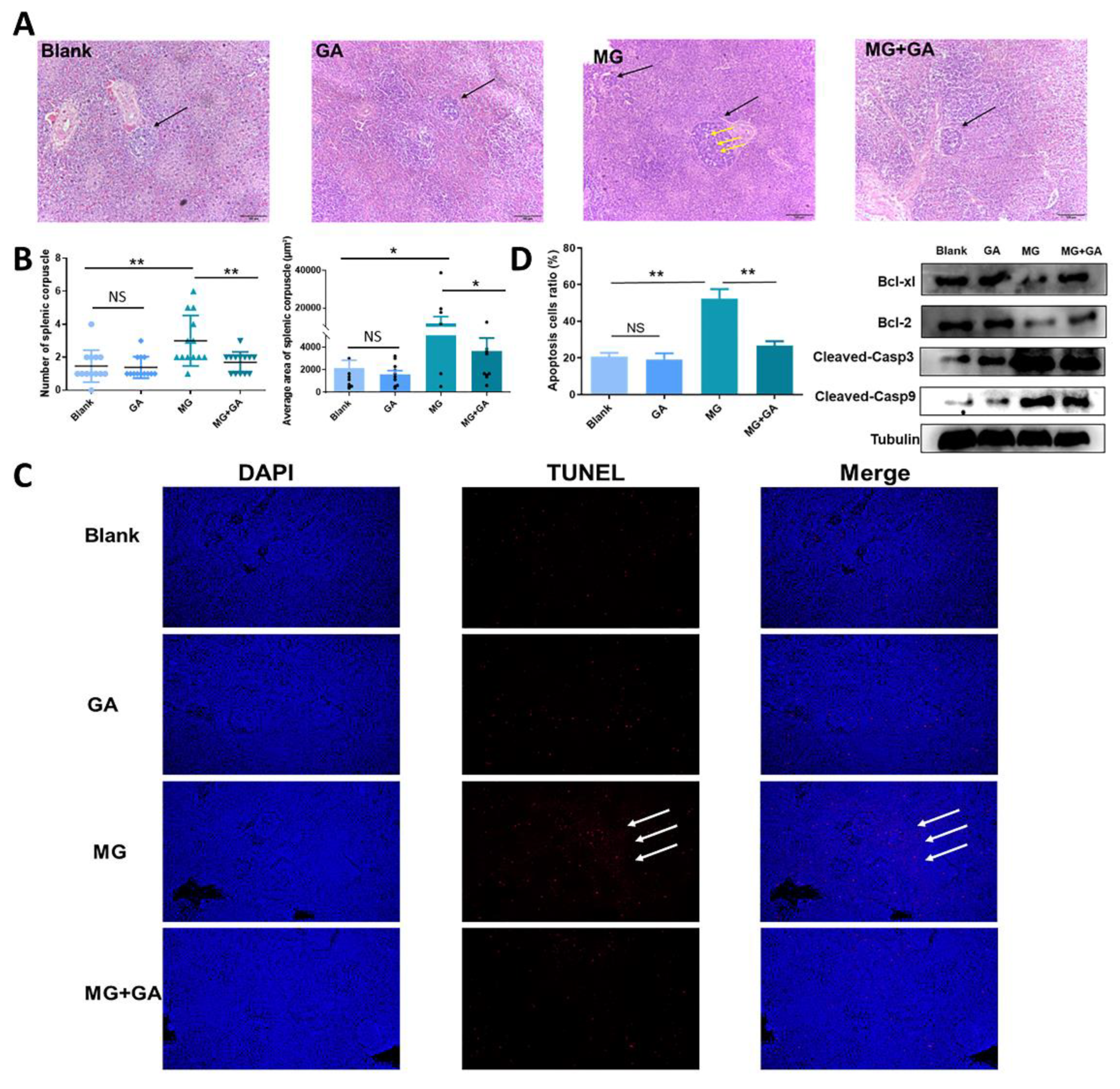
3.5. Disruption of HMGB1 Binding to TLR2-4 Receptors In Vivo Alleviates MG-Induced Immune Organ Damage
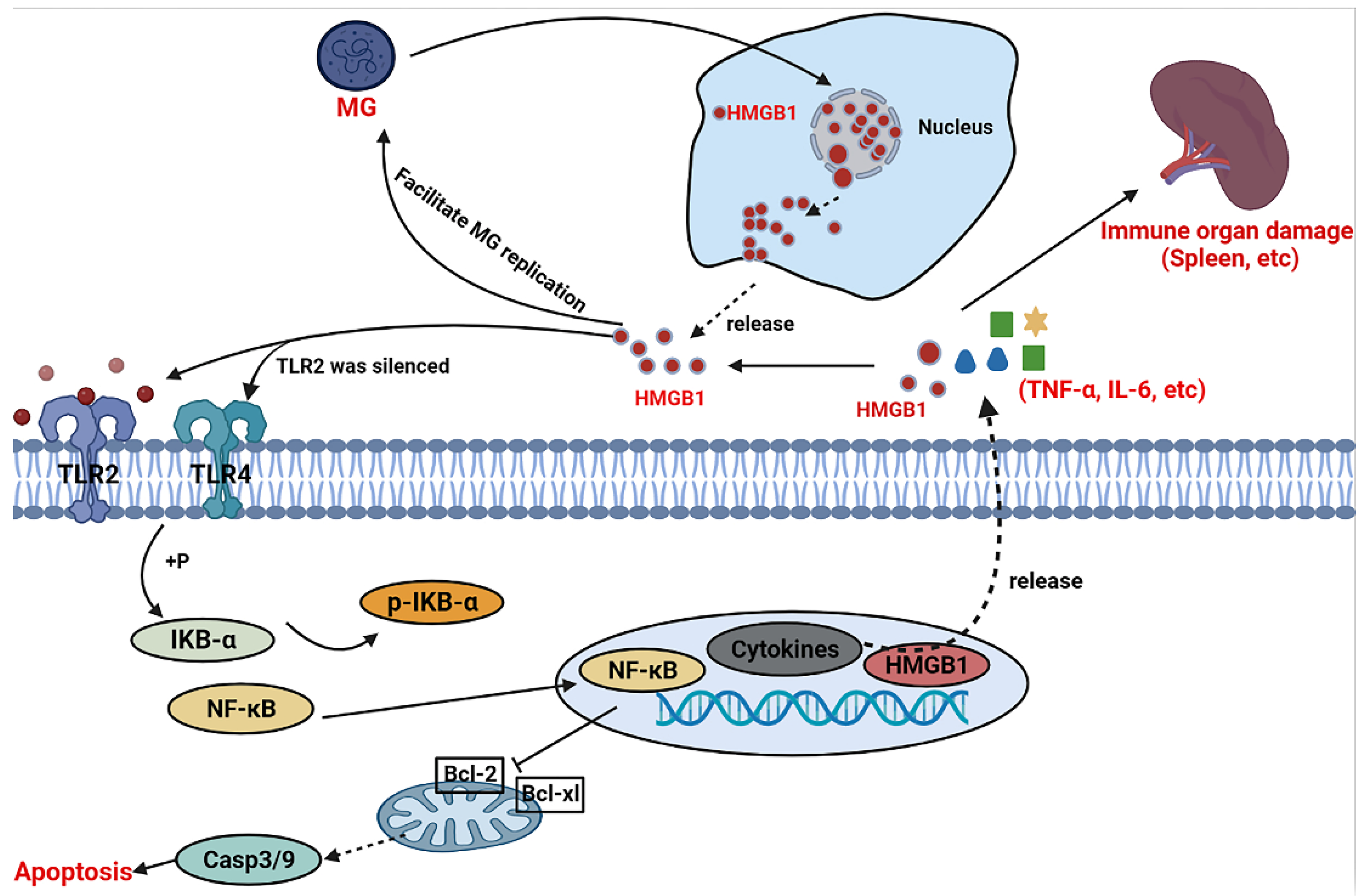
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Cao, X. Self-regulation and cross-regulation of pattern-recognition receptor signalling in health and disease. Nat. Rev. Immunol. 2016, 16, 35–50. [Google Scholar] [CrossRef] [PubMed]
- Gong, T.; Liu, L.; Jiang, W.; Zhou, R. DAMP-sensing receptors in sterile inflammation and inflammatory diseases. Nat. Rev. Immunol. 2020, 20, 95–112. [Google Scholar] [CrossRef] [PubMed]
- Bianchi, M.E. HMGB1 loves company. J. Leukoc. Biol. 2009, 86, 573–576. [Google Scholar] [CrossRef] [PubMed]
- Gardella SAndrei, C.; Ferrera, D.; Lotti, L.V.; Torrisi, M.R.; Bianchi, M.E.; Rubartelli, A. The nuclear protein HMGB1 is secreted by monocytes via a non-classical, vesicle-mediated secretory pathway. EMBO Rep. 2002, 3, 995–1001. [Google Scholar] [CrossRef] [PubMed]
- Chen, R.; Kang, R.; Tang, D. The mechanism of HMGB1 secretion and release. Exp. Mol. Med. 2022, 54, 91–102. [Google Scholar] [CrossRef]
- Tian, J.; Avalos, A.M.; Mao, S.Y.; Chen, B.; Senthil, K.; Wu, H.; Parroche, P.; Drabic, S.; Golenbock, D.; Sirois, C.; et al. Toll-like receptor 9-dependent activation by DNA-containing immune complexes is mediated by HMGB1 and RAGE. Nat. Immunol. 2007, 8, 487–496. [Google Scholar] [CrossRef]
- Andersson, U.; Yang, H.; Harris, H. High-mobility group box 1 protein (HMGB1) operates as an alarmin outside as well as inside cells. Semin. Immunol. 2018, 38, 40–48. [Google Scholar] [CrossRef]
- Andersson, U.; Tracey, K.J. HMGB1 is a therapeutic target for sterile inflammation and infection. Annu. Rev. Immunol. 2011, 29, 139–162. [Google Scholar] [CrossRef]
- Mahmoudi, J.; Sabermarouf, B.; Baradaran, B.; Sadat-Hatamnezhad, L.; Shotorbani, S.S. Up-regulation of TLR2 and TLR4 in high mobility group Box1-stimulated macrophages in pulpitis patients. Iran. J. Basic Med. Sci. 2017, 20, 209–215. [Google Scholar]
- Cai, J.; Yuan, H.; Wang, Q.; Yang, H.; Al-Abed, Y.; Hua, Z.; Wang, J.; Chen, D.; Wu, J.; Lu, B.; et al. HMGB1-Driven Inflammation and Intimal Hyperplasia After Arterial Injury Involves Cell-Specific Actions Mediated by TLR4. Arterioscler. Thromb. Vasc. Biol. 2015, 35, 2579–2593. [Google Scholar] [CrossRef]
- Ding, J.; Cui, X.; Liu, Q. Emerging role of HMGB1 in lung diseases: Friend or foe. J. Cell. Mol. Med. 2017, 21, 1046–1057. [Google Scholar] [CrossRef]
- Park, J.S.; Svetkauskaite, D.; He, Q.; Kim, J.Y.; Strassheim, D.; Ishizaka, A.; Abraham, E. Involvement of toll-like receptors 2 and 4 in cellular activation by high mobility group box 1 protein. J. Biol. Chem. 2004, 279, 7370–7377. [Google Scholar] [CrossRef] [PubMed]
- Attiq, A.; Yao, L.J.; Afzal, S.; Khan, M.A. The triumvirate of NF-κB, inflammation and cytokine storm in COVID-19. Int. Immunopharmacol. 2021, 101, 108255. [Google Scholar] [CrossRef] [PubMed]
- Ferreira, G.; Blasina, F.; Rodríguez Rey, M.; Anesetti, G.; Sapiro, R.; Chavarría, L.; Cardozo, R.; Rey, G.; Sobrevia, L.; Nicolson, G.L. Pathophysiological and molecular considerations of viral and bacterial infections during maternal-fetal and -neonatal interactions of SARS-CoV-2, Zika, and Mycoplasma infectious diseases. Biochim. Biophys. Acta Mol. Basis Dis. 2022, 1868, 166285. [Google Scholar] [CrossRef]
- Luo, R.; Wang, Y.; Guo, Q.; Fan, C.; Jiang, G.; Wang, L.; Zou, M.; Wang, T.; Sun, Y.; Peng, X. Andrographolide attenuates Mycoplasma gallisepticum-induced inflammation and apoptosis by the JAK/PI3K/AKT signal pathway in the chicken lungs and primary alveolar type II epithelial cells. Int. Immunopharmacol. 2022, 109, 108819. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Tong, D.; Sun, Y.; Sun, H.; Liu, F.; Zou, M.; Luo, R.; Peng, X. DF-1 cells prevent MG-HS infection through gga-miR-24-3p/RAP1B mediated decreased proliferation and increased apoptosis. Res. Vet. Sci. 2021, 141, 164–173. [Google Scholar] [CrossRef]
- Ishfaq, M.; Zhang, W.; Liu, Y.; Wang, J.; Wu, Z.; Shah, S.W.; Li, R.; Miao, Y.; Chen, C.; Li, J. Baicalin attenuated Mycoplasma gallisepticum-induced immune impairment in chicken bursa of fabricius through modulation of autophagy and inhibited inflammation and apoptosis. J. Sci. Food Agric. 2021, 101, 880–890. [Google Scholar] [CrossRef]
- Wang, J.; Ishfaq, M.; Li, J. Baicalin ameliorates Mycoplasma gallisepticum-induced inflammatory injury in the chicken lung through regulating the intestinal microbiota and phenylalanine metabolism. Food Funct. 2021, 12, 4092–4104. [Google Scholar] [CrossRef]
- Miao, Y.; Niu, D.; Wang, Z.; Wang, J.; Wu, Z.; Bao, J.; Hu, W.; Guo, Y.; Li, R.; Ishfaq, M.; et al. Mycoplasma gallisepticum induced inflammation-mediated Th1/Th2 immune imbalance via JAK/STAT signaling pathway in chicken trachea: Involvement of respiratory microbiota. Vet. Microbiol. 2022, 265, 109330. [Google Scholar] [CrossRef]
- Bi, D.; Ji, X. The isolation and identification of the Mycoplasma gallisepticum. Acta Vet. Zootechnol. Sin. 1988, 1, 146–148. [Google Scholar]
- Calus, D.; Maes, D.; Vranckx, K.; Villareal, I.; Pasmans, F.; Haesebrouck, F. Validation of ATP luminometry for rapid and accurate titration of Mycoplasma hyopneumoniae in Friis medium and a comparison with the color changing units assay. J. Microbiol. Methods 2010, 83, 335–340. [Google Scholar] [CrossRef] [PubMed]
- Miao, Y.; Niu, D.; Wang, Z.; Wang, J.; Wu, Z.; Bao, J.; Jin, X.; Li, R.; Ishfaq, M.; Li, J. Methylsulfonylmethane ameliorates inflammation via NF-κB and ERK/JNK-MAPK signaling pathway in chicken trachea and HD11 cells during Mycoplasma gallisepticum infection. Poult. Sci. 2022, 101, 101706. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Wang, L.; Luo, R.; Sun, Y.; Zou, M.; Wang, T.; Guo, Q.; Peng, X. Glycyrrhizic Acid against Mycoplasma gallisepticum-Induced Inflammation and Apoptosis Through Suppressing the MAPK Pathway in Chickens. J. Agric. Food Chem. 2022, 70, 1996–2009. [Google Scholar] [CrossRef]
- Ji, J.; Fu, T.; Dong, C.; Zhu, W.; Yang, J.; Kong, X.; Zhang, Z.; Bao, Y.; Zhao, R.; Ge, X.; et al. Targeting HMGB1 by ethyl pyruvate ameliorates systemic lupus erythematosus and reverses the senescent phenotype of bone marrow-mesenchymal stem cells. Aging 2019, 11, 4338–4353. [Google Scholar] [CrossRef] [PubMed]
- Behl, T.; Sharma, E.; Sehgal, A.; Kaur, I.; Kumar, A.; Arora, R.; Pal, G.; Kakkar, M.; Kumar, R.; Bungau, S. Expatiating the molecular approaches of HMGB1 in diabetes mellitus: Highlighting signalling pathways via RAGE and TLRs. Mol. Biol. Rep. 2021, 48, 1869–1881. [Google Scholar] [CrossRef] [PubMed]
- Li, C.; Peng, S.; Liu, X.; Han, C.; Wang, X.; Jin, T.; Liu, S.; Wang, W.; Xie, X.; He, X.; et al. Glycyrrhizin, a Direct HMGB1 Antagonist, Ameliorates Inflammatory Infiltration in a Model of Autoimmune Thyroiditis via Inhibition of TLR2-HMGB1 Signalin. Thyroid. Off. J. Am. Thyroid. Assoc. 2017, 27, 722–731. [Google Scholar] [CrossRef] [PubMed]
- Kawai, T.; Akira, S. Signaling to NF-kappaB by Toll-like receptors. Trends Mol. Med. 2007, 13, 460–469. [Google Scholar] [CrossRef]
- Yang, Y.; Wang, H.; Kouadir, M.; Song, H.; Shi, F. Recent advances in the mechanisms of NLRP3 inflammasome activation and its inhibitors. Cell Death Dis. 2019, 10, 128. [Google Scholar] [CrossRef]
- Lange, S.S.; Mitchell, D.L.; Vasquez, K.M. High mobility group protein B1 enhances DNA repair and chromatin modification after DNA damage. Proc. Natl. Acad. Sci. USA 2008, 105, 10320–10325. [Google Scholar] [CrossRef]
- Lu, B.; Wang, H.; Andersson, U.; Tracey, K.J. Regulation of HMGB1 release by inflammasomes. Protein Cell 2013, 4, 163–167. [Google Scholar] [CrossRef]
- Wang, C.Q.; Huang, B.F.; Wang, Y.; Tang, C.H.; Jin, H.C.; Shao, F.; Shao, J.K.; Wang, Q.; Zeng, Y. Subcellular localization of HMGB1 in colorectal cancer impacts on tumor grade and survival prognosis. Sci. Rep. 2020, 10, 18587. [Google Scholar] [CrossRef] [PubMed]
- Volchuk, A.; Ye, A.; Chi, L.; Steinberg, B.E.; Goldenberg, N.M. Indirect regulation of HMGB1 release by gasdermin D. Nat. Commun. 2020, 11, 4561. [Google Scholar] [CrossRef] [PubMed]
- Lu, B.; Wang, C.; Wang, M.; Li, W.; Chen, F.; Tracey, K.J.; Wang, H. Molecular mechanism and therapeutic modulation of high mobility group box 1 release and action: An updated review. Expert. Rev. Clin. Immunol. 2014, 10, 713–727. [Google Scholar] [CrossRef] [PubMed]
- Ding, X.; Li, S.; Zhu, L. Potential effects of HMGB1 on viral replication and virus infection-induced inflammatory responses: A promising therapeutic target for virus infection-induced inflammatory diseases. Cytokine Growth Factor Rev. 2021, 62, 54–61. [Google Scholar] [CrossRef] [PubMed]
- Yu, R.; Yang, D.; Lei, S.; Wang, X.; Meng, X.; Xue, B.; Zhu, H. Hmgb1 promotes hepatitis c virus replication by interaction with stem-loop 4 in the viral 5′ untranslated region. J. Virol. 2015, 90, 2332–2344. [Google Scholar] [CrossRef] [PubMed]
- Rizk, N.I.; Sallam, A.M.; El-Ansary, A.R.; El-Mesallamy, H.O. HMGB1 and SEPP1 as predictors of hepatocellular carcinoma in patients with viral C hepatitis: Effect of DAAs. Clin. Biochem. 2019, 70, 8–13. [Google Scholar] [CrossRef]
- Kagele, D.; Rossetto, C.C.; Tarrant, M.T.; Pari, G.S. Analysis of the interactions of viral and cellular factors with human cytomegalovirus lytic origin of replication, oriLyt. Virology 2012, 424, 106–114. [Google Scholar] [CrossRef]
- Li, R.; Zhang, J.; Pan, S.; Yuan, Y.; Qi, H.; Shu, H.; Hu, Y.; Ren, L.; Jiang, Y.; Yuan, S. HMGB1 aggravates lipopolysaccharide-induced acute lung injury through suppressing the activity and function of Tregs. Cell. Immunol. 2020, 356, 104192. [Google Scholar] [CrossRef]
- Qu, Y.; Zhan, Y.; Yang, S.; Ren, S.; Qiu, X.; Rehamn, Z.U.; Tan, L.; Sun, Y.; Meng, C.; Song, C.; et al. Newcastle disease virus infection triggers HMGB1 release to promote the inflammatory response. Virology 2018, 525, 19–31. [Google Scholar] [CrossRef]
- Duan, E.; Wang, D.; Luo, R.; Luo, J.; Gao, L.; Chen, H.; Fang, L.; Xiao, S. Porcine reproductive and respiratory syndrome virus infection triggers HMGB1 release to promote inflammatory cytokine production. Virology 2014, 468, 1–9. [Google Scholar] [CrossRef]
- Cheng, B.Q.; Jia, C.Q.; Liu, C.T.; Lu, X.F.; Zhong, N.; Zhang, Z.L.; Fan, W.; Li, Y.Q. Serum high mobility group box chromosomal protein 1 is associated with clinicopathologic features in patients with hepatocellular carcinoma. Dig. Liver Dis. Off. J. Ital. Soc. Gastroenterol. Ital. Assoc. Study Liver. 2008, 40, 446–452. [Google Scholar] [CrossRef] [PubMed]
- Sasahira, T.; Akama, Y.; Fujii, K.; Kuniyasu, H. Expression of receptor for advanced glycation end products and HMGB1/amphoterin in colorectal adenomas. Virchows Arch. Int. J. Pathol. 2005, 446, 411–415. [Google Scholar] [CrossRef] [PubMed]
- Suda, K.; Kitagawa, Y.; Ozawa, S.; Saikawa, Y.; Ueda, M.; Abraham, E.; Kitajima, M.; Ishizaka, A. Serum concentrations of high-mobility group box chromosomal protein 1 before and after exposure to the surgical stress of thoracic esophagectomy: A predictor of clinical course after surgery? Dis. Esophagus Off. J. Int. Soc. Dis. Esophagus 2006, 19, 5–9. [Google Scholar] [CrossRef] [PubMed]
- Momonaka, H.; Hasegawa, S.; Matsushige, T.; Inoue, H.; Kajimoto, M.; Okada, S.; Nakatsuka, K.; Morishima, T.; Ichiyama, T. High mobility group box 1 in patients with 2009 pandemic H1N1 influenza-associated encephalopathy. Brain Dev. 2014, 36, 484–488. [Google Scholar] [CrossRef] [PubMed]
- Ito, Y.; Torii, Y.; Ohta, R.; Imai, M.; Hara, S.; Kawano, Y.; Matsubayashi, T.; Inui, A.; Yoshikawa, T.; Nishimura, N.; et al. Increased levels of cytokines and high-mobility group box 1 are associated with the development of severe pneumonia, but not acute encephalopathy, in 2009 H1N1 influenza-infected children. Cytokine 2011, 56, 180–187. [Google Scholar] [CrossRef]
- Zhang, Y.; Zhang, M.; Wang, C.Y.; Shen, A. Ketamine alleviates LPS induced lung injury by inhibiting HMGB1-RAGE level. Eur. Rev. Med. Pharmacol. Sci. 2018, 22, 1830–1836. [Google Scholar]
- Andersson, U.; Yang, H.; Harris, H. Extracellular HMGB1 as a therapeutic target in inflammatory diseases. Expert Opin. Ther. Targets 2018, 22, 263–277. [Google Scholar] [CrossRef]
- Zainal, N.; Chang, C.P.; Cheng, Y.L.; Wu, Y.W.; Anderson, R.; Wan, S.W.; Chen, C.L.; Ho, T.S.; AbuBakar, S.; Lin, Y.S. Treatment reveals a novel role for HMGB1 in regulation of the type 1 interferon response in dengue virus infection. Sci. Rep. 2017, 7, 42998. [Google Scholar] [CrossRef]
- Nosaka, N.; Yashiro, M.; Yamada, M.; Fujii, Y.; Tsukahara, H.; Liu, K.; Nishibori, M.; Matsukawa, A.; Morishima, T. Anti-high mobility group box-1 monoclonal antibody treatment provides protection against influenza A virus (H1N1)-induced pneumonia in mice. Crit. Care 2015, 19, 249. [Google Scholar] [CrossRef]
- Hou, X.Q.; Qin, J.L.; Zheng, X.X.; Wang, L.; Yang, S.T.; Gao, Y.W.; Xia, X.Z. Potential role of high-mobility group box 1 protein in the pathogenesis of influenza H5N1 virus infection. Acta Virol. 2014, 58, 69–75. [Google Scholar] [CrossRef]
- Smolarczyk, R.; Cichoń, T.; Matuszczak, S.; Mitrus, I.; Lesiak, M.; Kobusińska, M.; Kamysz, W.; Jarosz, M.; Sieroń, A.; Szala, S. The role of Glycyrrhizin, an inhibitor of HMGB1 protein, in anticancer therapy. Arch. Immunol. Ther. Exp. 2012, 60, 391–399. [Google Scholar] [CrossRef] [PubMed]
- Sitia, G.; Iannacone, M.; Müller, S.; Bianchi, M.E.; Guidotti, L.G. Treatment with HMGB1 inhibitors diminishes CTL-induced liver disease in HBV transgenic mice. J. Leukoc. Biol. 2007, 81, 100–107. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Xu, Q.; Mei, L.; Lei, H.; Wen, Q.; Miao, J.; Huang, H.; Chen, D.; Du, S.; Zhang, S.; et al. Paeonol attenuates acute lung injury by inhibiting HMGB1 in lipopolysaccharide-induced shock rats. Int. Immunopharmacol. 2018, 61, 169–177. [Google Scholar] [CrossRef] [PubMed]
- He, Z.W.; Qin, Y.H.; Wang, Z.W.; Chen, Y.; Shen, Q.; Dai, S.M. HMGB1 acts in synergy with lipopolysaccharide in activating rheumatoid synovial fibroblasts via p38 MAPK and NF-κB signaling pathways. Mediat. Inflamm. 2013, 2013, 596716. [Google Scholar] [CrossRef]
- Rayavara, K.; Kurosky, A.; Stafford, S.J.; Garg, N.J.; Brasier, A.R.; Garofalo, R.P.; Hosakote, Y.M. Proinflammatory Effects of Respiratory Syncytial Virus-Induced Epithelial HMGB1 on Human Innate Immune Cell Activation. J. Immunol. 2018, 201, 2753–2766. [Google Scholar] [CrossRef]
- Yang, H.; Wang, H.; Andersson, U. Targeting Inflammation Driven by HMGB1. Front. Immunol. 2020, 11, 484. [Google Scholar] [CrossRef]
- Kumar, M.; Majumder, D.; Mal, S.; Chakraborty, S.; Gupta, P.; Jana, K.; Gupta, U.D.; Ghosh, Z.; Kundu, M.; Basu, J. Activating transcription factor 3 modulates the macrophage immune response to Mycobacterium tuberculosis infection via reciprocal regulation of inflammatory genes and lipid body formation. Cell. Microbiol. 2020, 22, e13142. [Google Scholar] [CrossRef]
- Kang, R.; Chen, R.; Zhang, Q.; Hou, W.; Wu, S.; Cao, L.; Huang, J.; Yu, Y.; Fan, X.G.; Yan, Z.; et al. HMGB1 in health and disease. Mol. Asp. Med. 2014, 40, 1–116. [Google Scholar]
- Bi, X.; Yan, X.; Jiang, B.; Liang, J.; Zhou, J.; Lu, S.; Liu, J.; Luo, L.; Yin, Z. Indoprofen exerts a potent therapeutic effect against sepsis by alleviating high mobility group box 1-mediated inflammatory responses. Toxicol. Appl. Pharmacol. 2021, 433, 115778. [Google Scholar] [CrossRef]
- Li, R.; Shang, Y.; Yu, Y.; Zhou, T.; Xiong, W.; Zou, X. High-mobility group box 1 protein participates in acute lung injury by activating protein kinase R and inducing M1 polarization. Life Sci. 2020, 246, 117415. [Google Scholar] [CrossRef]
- Senthilkumar, D.; Rajukumar, K.; Kumar, M.; Kalaiyarasu, S.; Shrivastava, D.; Katare, M.; Kulkarni, D.D.; Singh, V.P. Porcine reproductive and respiratory syndrome virus induces concurrent elevation of High Mobility Group Box-1 protein and pro-inflammatory cytokines in experimentally infected piglets. Cytokine 2019, 113, 21–30. [Google Scholar] [CrossRef] [PubMed]
- Bualeong, T.; Kebir, S.; Hof, D.; Goelz, L.; Graewe, M.; Ehrentraut, S.F.; Knuefermann, P.; Baumgarten, G.; Meyer, R.; Ehrentraut, H. Tlr2 deficiency does not limit the development of left ventricular hypertrophy in a model of transverse aortic constriction induced pressure overload. J. Negat. Results Biomed. 2016, 15, 9. [Google Scholar] [CrossRef] [PubMed]
- Ishfaq, M.; Chen, C.; Bao, J.; Zhang, W.; Wu, Z.; Wang, J.; Liu, Y.; Tian, E.; Hamid, S.; Li, R.; et al. Baicalin ameliorates oxidative stress and apoptosis by restoring mitochondrial dynamics in the spleen of chickens via the opposite modulation of nf-κb and nrf2/ho-1 signaling pathway during mycoplasma gallisepticum infection. Poult. Sci. 2019, 98, 6296–6310. [Google Scholar] [CrossRef] [PubMed]
- Chen, C.; Li, J.; Zhang, W.; Shah, S.W.A.; Ishfaq, M. Mycoplasma gallisepticum triggers immune damage in the chicken thymus by activating the tlr-2/myd88/nf-κb signaling pathway and nlrp3 inflammasome. Vet. Res. 2020, 51, 52. [Google Scholar] [CrossRef] [Green Version]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Name | Primer Sequence (5′-3′) |
|---|---|
| Primers for CDS Cloning | |
| HMGB1-CDS-F | CCGCTCGAGCGGATGGGCAAAGGTGATCCCAA |
| HMGB1-CDS-R | CCGGAATTCCGGTTATTCATCATCATCATCATCTTCC |
| Primers for RT-qPCR | |
| GAPDH-F | GAGGGTAGTGAAGGCTGCTG |
| GAPDH-R | CACAACACGGTTGCTGTATC |
| TLR-2-F | CGCAAGCTTATGTTCAACCAAAG |
| TLR-2-R | CGCCTCGAGCTATGACTTCAAGG |
| TLR-4-F | ATCTTTCAAGGTGCCACATC |
| TLR-4-R | GGATATGCTTGTTTCCACCA |
| TNF-α-F | GGACAGCCTATGCCAACAAG |
| TNF-α-R | ACACGACAGCCAAGTCAACG |
| IL-1β-F | ACTGGGCATCAAGGGCTACA |
| IL-1β-R | GCTGTCCAGGCGGTAGAAGA |
| IL-12-F | TGGAACGATGAGACACCAGC |
| IL-12-R | AGACAGGCAGGTGTAGTTGC |
| IL-6-F | CTCCTCGCCAATCTGAAGTC |
| IL-6-R | CCCTCACGGTCTTCTCCATA |
| 16 SrRNA-F | AGCTAATCTGTAAAGTTGGTC |
| 16SrRNA-R | CGCTTCCTTGCGGTTAGCAAC |
| HMGB1-F | AAGGTGATCCCAAGAAGCCG |
| HMGB1-R | GAAGCTTGTCAGCCTTTGCC |
| Name | Sequences (5′-3′) |
|---|---|
| Si-TLR2 | GCCAUGCAAACUUUCACAATT |
| UUGUGAAAGUUUGCAUGGCTT | |
| Si-TLR4 | GCAGCCUUCCAUGGCUUAATT |
| UUAAGCCAUGGAAGGCUGCTT | |
| Si-HMGB1 | GCAGAUGAUAAACAGCCUUTT |
| AAGGCUGUUUAUCAUCUGCTT | |
| NC | UUCUCCGAACGUGUCACGUTT |
| ACGUGACACGUUCGGAGAATT |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, Y.; Wang, L.; Hu, F.; Zou, M.; Luo, R.; Sun, Y.; Wang, T.; Guo, Q.; Peng, X. Extracellular HMGB1 as Inflammatory Mediator in the Progression of Mycoplasma Gallisepticum Infection. Cells 2022, 11, 2817. https://doi.org/10.3390/cells11182817
Wang Y, Wang L, Hu F, Zou M, Luo R, Sun Y, Wang T, Guo Q, Peng X. Extracellular HMGB1 as Inflammatory Mediator in the Progression of Mycoplasma Gallisepticum Infection. Cells. 2022; 11(18):2817. https://doi.org/10.3390/cells11182817
Chicago/Turabian StyleWang, Yingjie, Lulu Wang, Fuli Hu, Mengyun Zou, Ronglong Luo, Yingfei Sun, Tengfei Wang, Qiao Guo, and Xiuli Peng. 2022. "Extracellular HMGB1 as Inflammatory Mediator in the Progression of Mycoplasma Gallisepticum Infection" Cells 11, no. 18: 2817. https://doi.org/10.3390/cells11182817