
Bta-miR-223 Targeting the RHOB Gene in Dairy Cows Attenuates LPS-Induced Inflammatory Responses in Mammary Epithelial Cells


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Cell Culture
2.2. miRNA Mimic, Inhibitors and siRNA
2.3. Dual-Luciferase Reporter Assays
2.4. Transfection of bMECs
2.5. RNA Isolation and Real-Time Quantitative PCR (qPCR)
2.6. Western Blot Analysis
2.7. ELISA for Cytokines
2.8. Statistical Analysis
3. Results
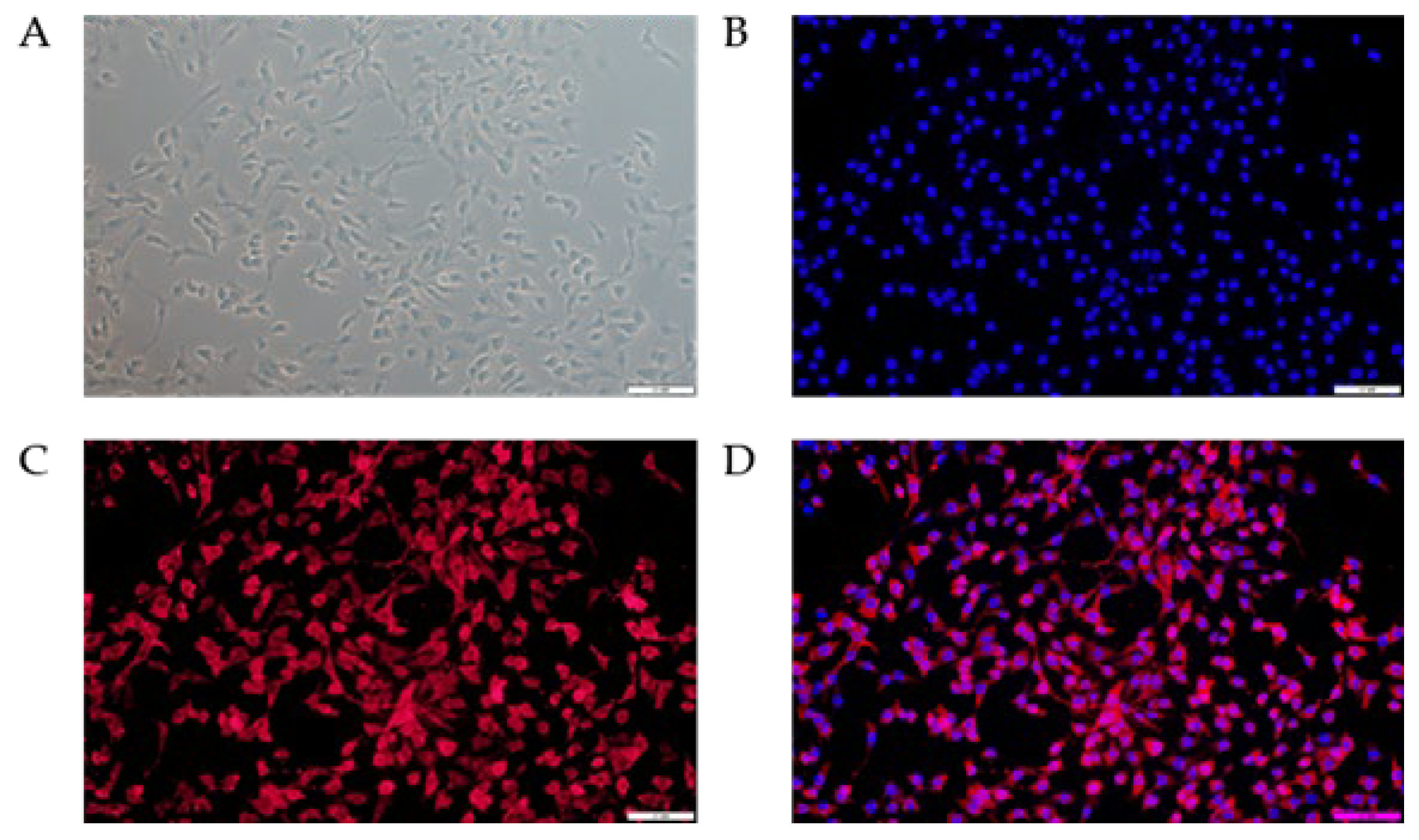
3.1. Phenotypic Verification of bMECs
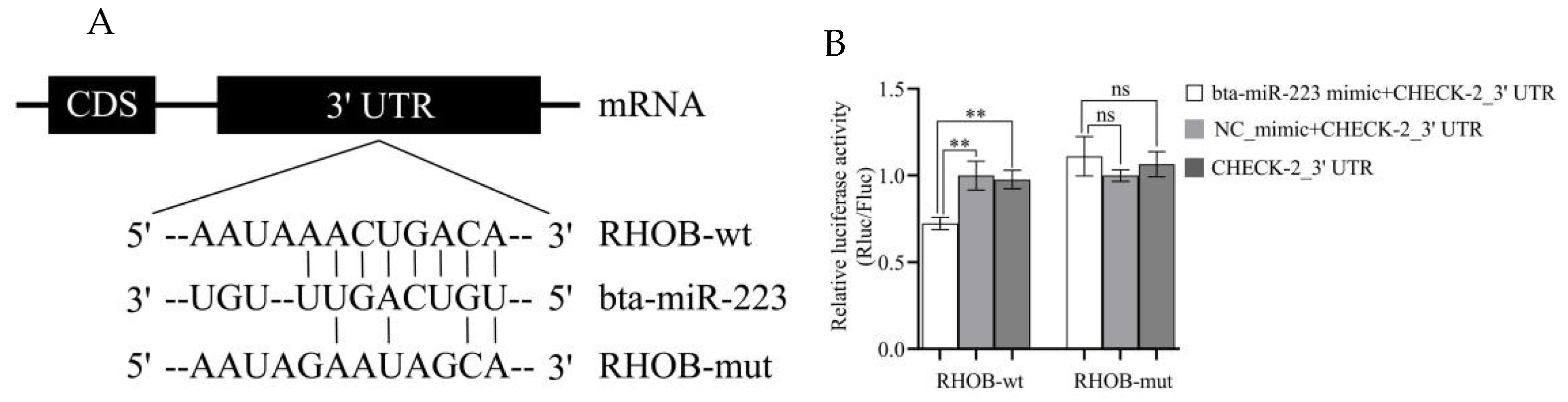
3.2. RHOB Is a Target Gene of bta-miR-223
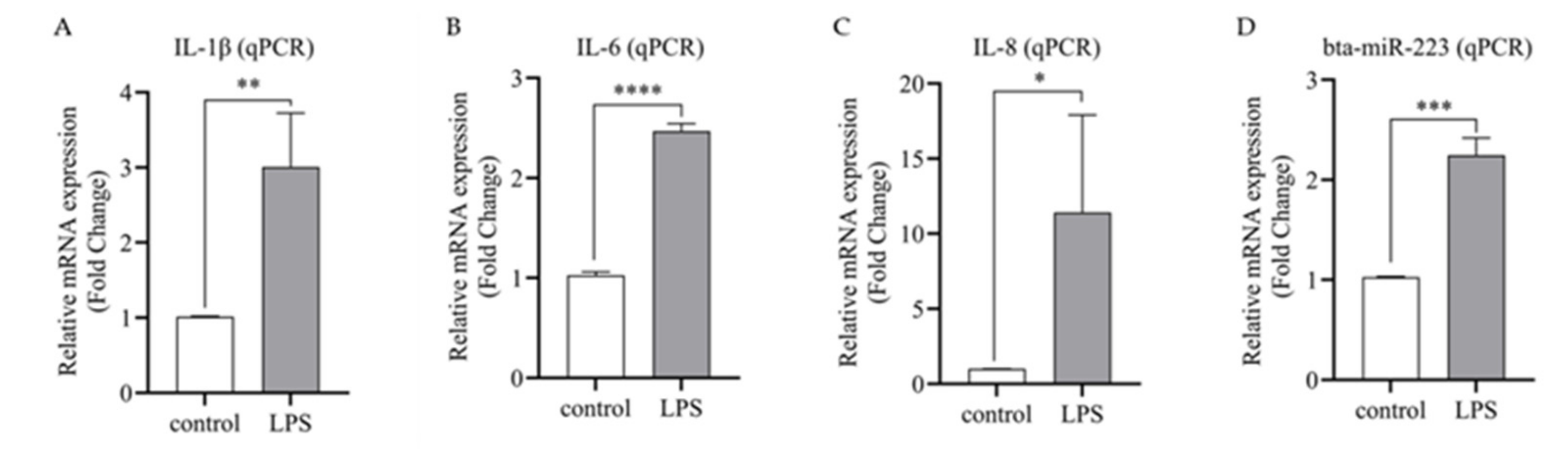
3.3. The Expression of Bta-miR-223 Is Up-Regulated in Inflammatory bMECs
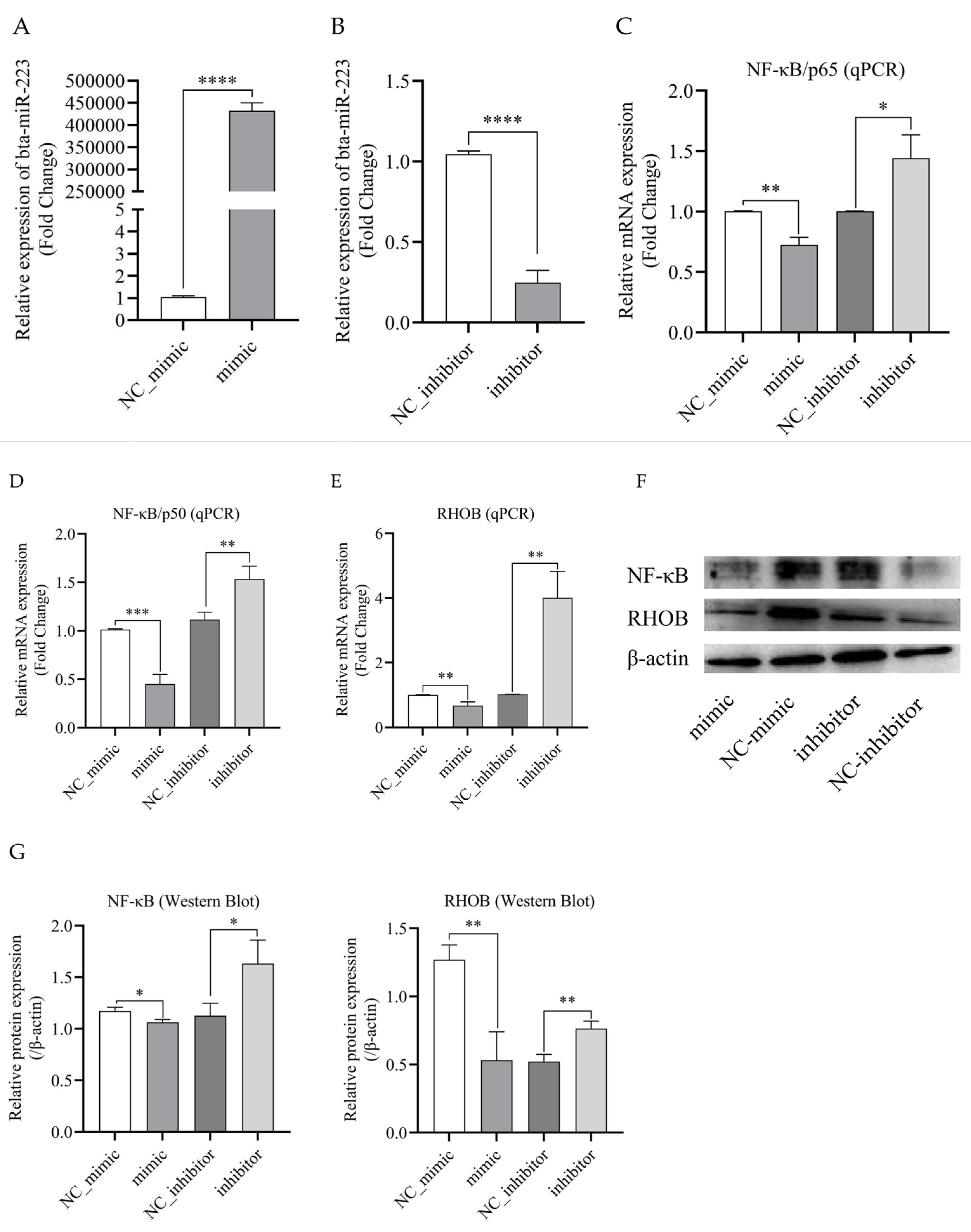
3.4. Bta-miR-223 Inhibits the Expression of RHOB and NF-κB Genes
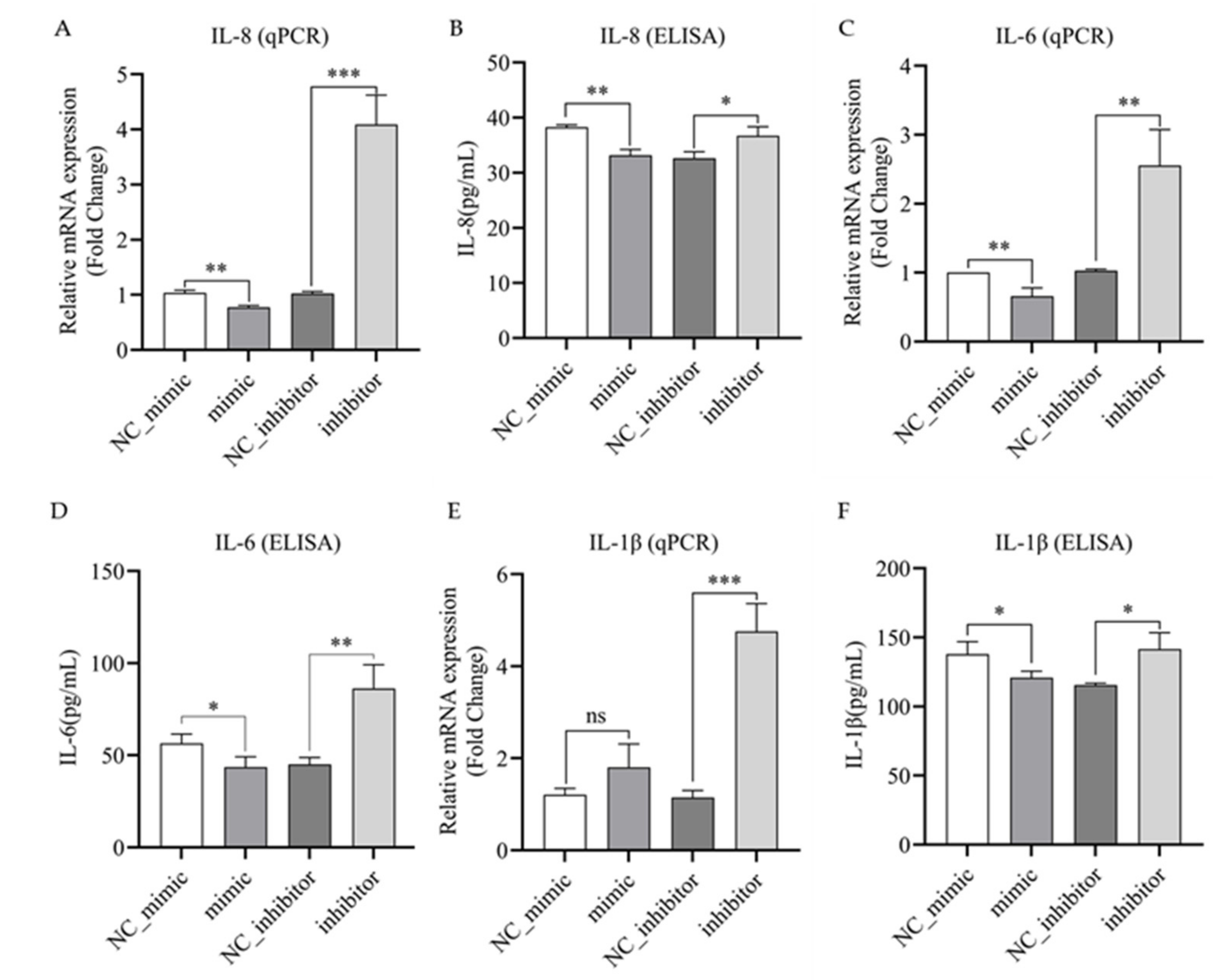
3.5. Bta-miR-223 Negatively Regulates the Expression and Secretion of IL-1β, IL-6 and IL-8 in LPS-Stimulated bMECs
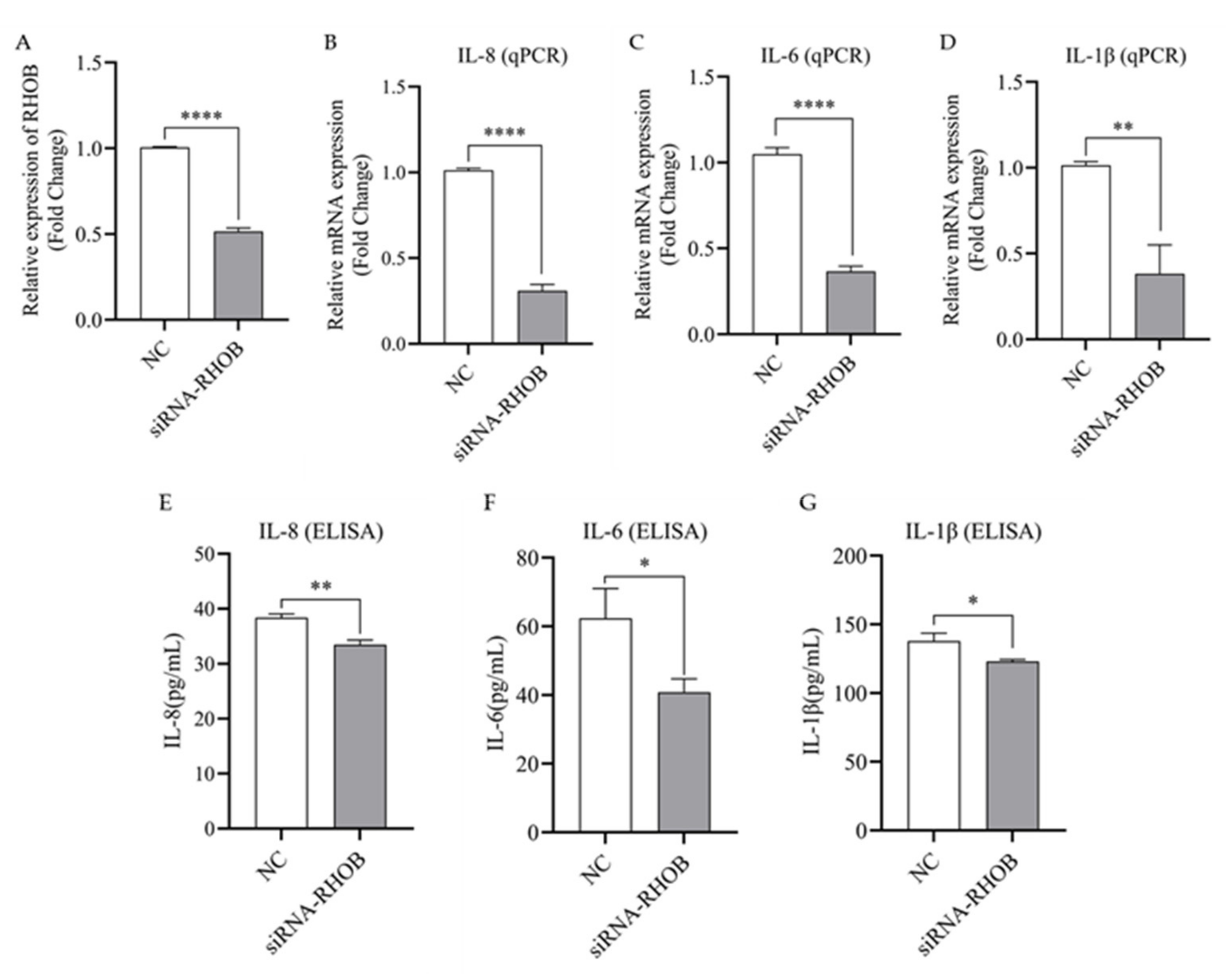
3.6. Silencing of RHOB Inhibits the Expression and Secretion of IL-1β, IL-6 and IL-8 in LPS-Induced bMECs
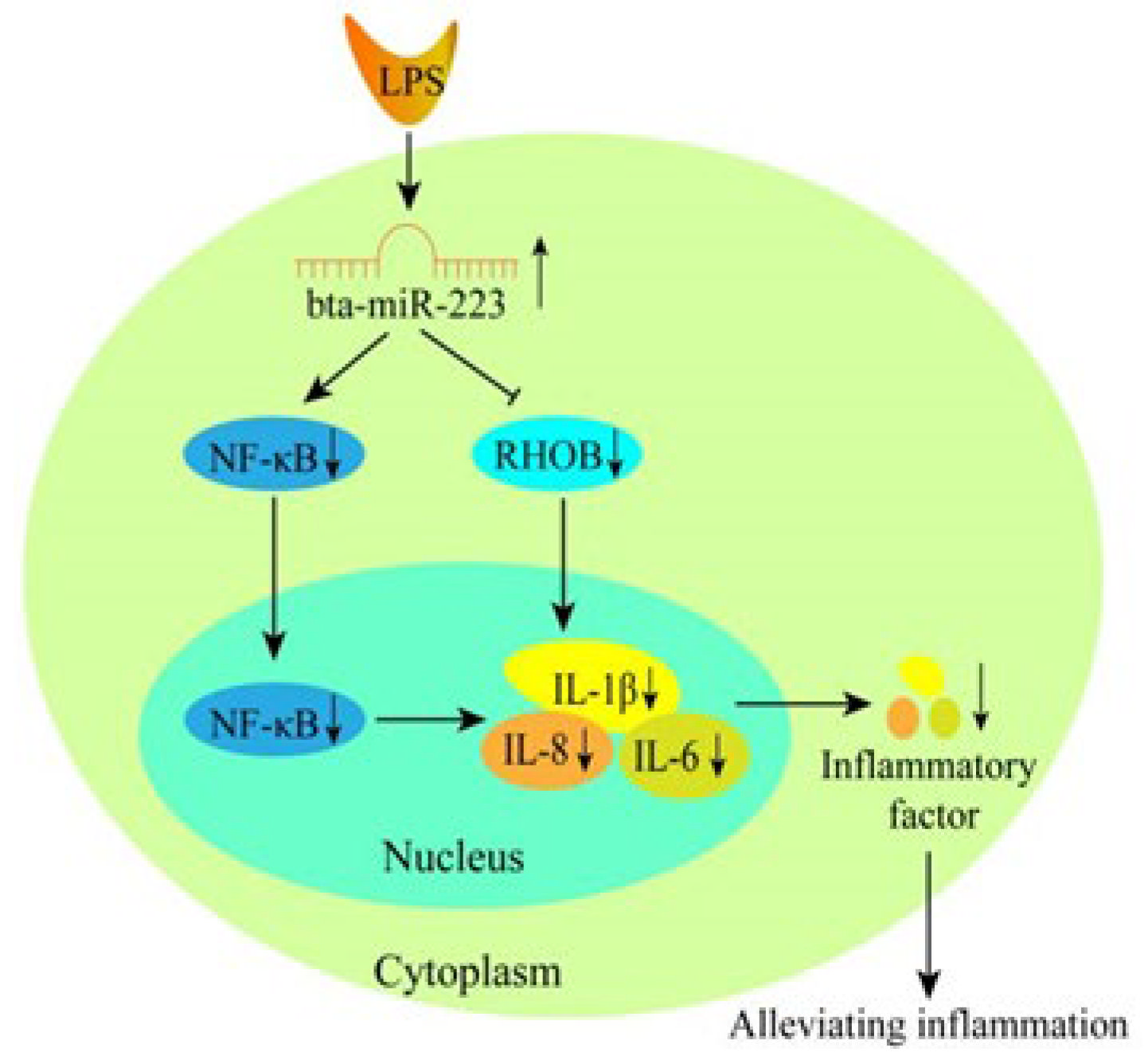
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Heikkilä, A.M.; Liski, E.; Pyörälä, S.; Taponen, S. Pathogen-specific production losses in bovine mastitis. J. Dairy Sci. 2018, 101, 9493–9504. [Google Scholar] [CrossRef] [PubMed]
- De Schepper, S.; De Ketelaere, A.; Bannerman, D.D.; Paape, M.J.; Peelman, L.; Burvenich, C. The toll-like receptor-4 (TLR-4) pathway and its possible role in the pathogenesis of Escherichia coli mastitis in dairy cattle. Vet. Res. 2008, 39, 5. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.; Tian, R.F.; Zhang, H.L.; Liu, X.L. Anti-mastitis SNV identification of NFκB1 in Chinese Holstein cows and the possible anti-inflammation role of NFκB1/p105 in bovine MECs. Acta Biochim. Biophys. Sin. 2020, 52, 1191–1201. [Google Scholar] [CrossRef] [PubMed]
- Dai, W.Y.; Wang, M.Y.; Wang, P.Q.; Wen, J.; Wang, J.Y.; Cha, S.; Xiao, X.L.; He, Y.R.; Shu, R.; Bai, D. lncRNA NEAT1 ameliorates LPS-induced inflammation in MG63 cells by activating autophagy and suppressing the NLRP3 inflammasome. Int. J. Mol. Med. 2021, 47, 607–620. [Google Scholar] [CrossRef] [PubMed]
- Dang, C.P.; Leelahavanichkul, A. Over-expression of miR-223 induces M2 macrophage through glycolysis alteration and attenuates LPS-induced sepsis mouse model, the cell-based therapy in sepsis. PLoS ONE 2020, 15, e236038. [Google Scholar] [CrossRef] [PubMed]
- Fu, Y.H.; Zhou, E.S.; Liu, Z.C.; Li, F.Y.; Liang, D.J.; Liu, B.; Song, X.J.; Zhao, F.Y.; Fen, X.S.; Li, D.P.; et al. Staphylococcus aureus and Escherichia coli elicit different innate immune responses from bovine mammary epithelial cells. Vet. Immunol. Immunopathol. 2013, 155, 245–252. [Google Scholar] [CrossRef]
- Zheng, L.H.; Xu, Y.Y.; Lu, J.Y.; Liu, M.; Dai, B.; Miao, J.F.; Yin, Y.L. Variant innate immune responses of mammary epithelial cells to challenge by Staphylococcus aureus, Escherichia coli and the regulating effect of taurine on these bioprocesses. Free Radic. Biol. Med. 2016, 96, 166–180. [Google Scholar] [CrossRef]
- Lim, L.P.; Glasner, M.E.; Yekta, S.; Burge, C.B.; Bartel, D.P. Vertebrate microRNA genes. Science 2003, 299, 1540. [Google Scholar] [CrossRef]
- Han, S.; Li, X.L.; Liu, J.; Zou, Z.W.; Luo, L.; Wu, R.; Zhao, Z.H.; Wang, C.Y.; Shen, B.L. Bta-miR-223 Targeting CBLB Contributes to Resistance to Staphylococcus aureus Mastitis Through the PI3K/AKT/NF-κB Pathway. Front. Vet. Sci. 2020, 7, 529. [Google Scholar] [CrossRef]
- Li, L.M.; Huang, J.M.; Zhang, X.J.; Ju, Z.H.; Qi, C.; Zhang, Y.; Li, Q.L.; Wang, C.F.; Miao, W.R.; Zhong, J.F.; et al. One SNP in the 3’-UTR of HMGB1 gene affects the binding of target bta-miR-223 and is involved in mastitis in dairy cattle. Immunogenetics 2012, 64, 817–824. [Google Scholar] [CrossRef]
- Yan, Y.R.; Lu, K.X.; Ye, T.; Zhang, Z.W. MicroRNA-223 attenuates LPS-induced inflammation in an acute lung injury model via the NLRP3 inflammasome and TLR4/NF-κB signaling pathway via RHOB. Int. J. Mol. Med. 2019, 43, 1467–1477. [Google Scholar] [CrossRef] [PubMed]
- Sun, G.H.; Li, H.T.; Rossi, J.J. Sequence context outside the target region influences the effectiveness of miR-223 target sites in the RhoB 3’ UTR. Nucleic Acids Res. 2010, 38, 239–252. [Google Scholar] [CrossRef] [PubMed]
- Hu, Y.; Yi, B.; Chen, X.; Xu, L.; Zhou, X.J.; Zhu, X.G. miR-223 promotes tumor progression via targeting RhoB in gastric cancer. J. Oncol. 2022, 2022, 6708871. [Google Scholar] [CrossRef] [PubMed]
- Li, S.F.; Feng, Y.P.; Huang, Y.X.; Liu, Y.; Wang, Y.X.; Liang, Y.; Zeng, H.; Qu, H.; Wei, L. MiR-223-3p regulates cell viability, migration, invasion, and apoptosis of non-small cell lung cancer cells by targeting RHOB. Open Life Sci. 2020, 15, 389–399. [Google Scholar] [CrossRef]
- Tang, H.C.; Lai, Y.Y.; Zheng, J.; Jiang, H.Y.; Xu, G. MiR-223-3p inhibits antigen endocytosis and presentation and promotes the tolerogenic potential of dendritic cells through targeting mannose receptor signaling and Rhob. J. Immunol. Res. 2020, 2020, 1379458. [Google Scholar] [CrossRef]
- Zhang, N.J.; Fu, L.Y.; Bu, Y.H.; Yao, Y.; Wang, Y.J. Downregulated expression of miR-223 promotes toll-like receptor-activated inflammatory responses in macrophages by targeting RhoB. Mol. Immunol. 2017, 91, 42–48. [Google Scholar] [CrossRef]
- Yang, J.; Hu, Q.C.; Wang, J.P.; Ren, Q.Q.; Wang, X.P.; Luoreng, Z.M.; Wei, D.W.; Ma, Y. RNA-Seq reveals the role of miR-29c in regulating inflammation and oxidative stress of bovine mammary epithelial cells. Front. Vet. Sci. 2022, 9, 865415. [Google Scholar] [CrossRef]
- Wang, X.P.; Luoreng, Z.M.; Zan, L.S.; Li, F.; Li, N. Bovine miR-146a regulates inflammatory cytokines of bovine mammary epithelial cells via targeting the TRAF6 gene. J. Dairy Sci. 2017, 100, 7648–7658. [Google Scholar] [CrossRef]
- Strandberg, Y.; Gray, C.; Vuocolo, T.; Donaldson, L.; Broadway, M.; Tellam, R. Lipopolysaccharide and lipoteichoic acid induce different innate immune responses in bovine mammary epithelial cells. Cytokine 2005, 31, 72–86. [Google Scholar] [CrossRef]
- Jiao, P.; Wang, X.P.; Luoreng, Z.M.; Yang, J.; Jia, L.; Ma, Y.; Wei, D.W. MiR-223: An effective regulator of immune cell differentiation and inflammation. Int. J. Biol. Sci. 2021, 17, 2308–2322. [Google Scholar] [CrossRef]
- Naeem, A.; Zhong, K.; Moisá, S.J.; Drackley, J.K.; Moyes, K.M.; Loor, J.J. Bioinformatics analysis of microRNA and putative target genes in bovine mammary tissue infected with Streptococcus uberis. J. Dairy Sci. 2012, 95, 6397–6408. [Google Scholar] [CrossRef] [PubMed]
- Saenz-De-Juano, M.D.; Silvestrelli, G.; Bauersachs, S.; Ulbrich, S.E. Determining extracellular vesicles properties and miRNA cargo variability in bovine milk from healthy cows and cows undergoing subclinical mastitis. BMC Genomics 2022, 23, 189. [Google Scholar] [CrossRef] [PubMed]
- Mazières, J.; Tillement, V.; Allal, C.; Clanet, C.; Bobin, L.; Chen, Z.; Sebti, S.M.; Favre, G.; Pradines, A. Geranylgeranylated, but not farnesylated, RhoB suppresses Ras transformation of NIH-3T3 cells. Exp. Cell Res. 2005, 304, 354–364. [Google Scholar] [CrossRef] [PubMed]
- Wei, L.J.; Li, J.A.; Bai, D.M.; Song, Y. MiR-223-RhoB signaling pathway regulates the proliferation and apoptosis of colon adenocarcinoma. Chem. Biol. Interact. 2018, 289, 9–14. [Google Scholar] [CrossRef] [PubMed]
- Fritz, G.; Kaina, B. Ras-related GTPase Rhob represses NF-κB signaling. J. Biol. Chem. 2001, 276, 3115–3122. [Google Scholar] [CrossRef]
- Chen, Y.X.; Li, Z.B.; Diao, F.; Cao, D.M.; Fu, C.C.; Lu, J. Up-regulation of RhoB by glucocorticoids and its effects on the cell proliferation and NF-kappaB transcriptional activity. J. Steroid Biochem. Mol. Biol. 2006, 101, 179–187. [Google Scholar] [CrossRef]
- Wang, X.H.; Wang, Y.; Diao, F.; Lu, J. RhoB is involved in lipopolysaccharide-induced inflammation in mouse in vivo and in vitro. J. Physiol. Biochem. 2013, 69, 189–197. [Google Scholar] [CrossRef]
- Huang, G.X.; Su, J.; Zhang, M.Z.; Jin, Y.D.; Wang, Y.; Zhou, P.; Lu, J. RhoB regulates the function of macrophages in the hypoxia-induced inflammatory response. Cell. Mol. Immunol. 2017, 14, 265–275. [Google Scholar] [CrossRef]
- Kroon, J.; Tol, S.; van Amstel, S.; Elias, J.A.; Fernandez-Borja, M. The small GTPase RhoB regulates TNFα signaling in endothelial cells. PLoS ONE 2013, 8, e75031. [Google Scholar] [CrossRef]
- Wei, Z.K.; Wang, J.J.; Wang, Y.N.; Wang, C.Q.; Liu, X.; Han, Z.; Fu, Y.H.; Yang, Z.T. Effects of neutrophil extracellular traps on bovine mammary epithelial cells in vitro. Front. Immunol. 2019, 10, 1003. [Google Scholar] [CrossRef]
- Belay, N.; Mohammed, N.; Seyoum, W. Bovine mastitis: Prevalence, risk factors, and bacterial pathogens isolated in lactating cows in gamo zone, southern ethiopia. Vet. Med. 2022, 13, 9–19. [Google Scholar] [CrossRef] [PubMed]
- Gilbert, F.B.; Cunha, P.; Jensen, K.; Glass, E.J.; Foucras, G.; Robert-Granié, C.; Rupp, R.; Rainard, P. Differential response of bovine mammary epithelial cells to Staphylococcus aureus or Escherichia coli agonists of the innate immune system. Vet. Res. 2013, 44, 40. [Google Scholar] [CrossRef] [PubMed]
- Liu, M.J.; Song, S.X.; Li, H.R.; Jiang, X.Y.; Yin, P.; Wan, C.R.; Liu, X.X.; Liu, F.H.; Xu, J.Q. The protective effect of caffeic acid against inflammation injury of primary bovine mammary epithelial cells induced by lipopolysaccharide. J. Dairy Sci. 2014, 97, 2856–2865. [Google Scholar] [CrossRef] [PubMed]
- Sun, L.T.; Chen, L.; Wang, F.G.; Zheng, X.; Yuan, C.F.; Niu, Q.G.; Li, Z.; Deng, L.; Zheng, B.B.; Li, C.J.; et al. Exogenous hydrogen sulfide prevents lipopolysaccharide-induced inflammation by blocking the TLR4/NF-κB pathway in MAC-T cells. Gene 2019, 710, 114–121. [Google Scholar] [CrossRef]
- Wang, J.J.; Guo, C.M.; Wei, Z.K.; He, X.X.; Kou, J.H.; Zhou, E.S.; Yang, Z.T.; Fu, Y.H. Morin suppresses inflammatory cytokine expression by downregulation of nuclear factor-κB and mitogen-activated protein kinase (MAPK) signaling pathways in lipopolysaccharide-stimulated primary bovine mammary epithelial cells. J. Dairy Sci. 2016, 99, 3016–3022. [Google Scholar] [CrossRef]
- Akhtar, M.; Guo, S.; Guo, Y.F.; Zahoor, A.; Shaukat, A.; Chen, Y.; Umar, T.; Deng, P.G.; Guo, M. Upregulated-gene expression of pro-inflammatory cytokines (TNF-α, IL-1β and IL-6) via TLRs following NF-κB and MAPKs in bovine mastitis. Acta Trop. 2020, 207, 105458. [Google Scholar] [CrossRef]
- Wang, X.P.; Luoreng, Z.M.; Zan, L.S.; Raza, S.H.; Li, F.; Li, N.; Liu, S. Expression patterns of miR-146a and miR-146b in mastitis infected dairy cattle. Mol. Cell. Probes 2016, 30, 342–344. [Google Scholar] [CrossRef]
- Donado, C.A.; Cao, A.B.; Simmons, D.P.; Croker, B.A.; Brennan, P.J.; Brenner, M.B. A two-cell model for IL-1βrelease mediated by death-receptor signaling. Cell. Rep. 2020, 31, 107466. [Google Scholar] [CrossRef]
- Harada, A.; Sekido, N.; Akahoshi, T.; Wada, T.; Mukaida, N.; Matsushima, K. Essential involvement of interleukin-8 (IL-8) in acute inflammation. J. Leukoc. Biol. 1994, 56, 559–564. [Google Scholar] [CrossRef]
- Weber, A.; Wasiliew, P.; Kracht, M. Interleukin-1beta (IL-1beta) processing pathway. Sci. Signal. 2010, 3, 2. [Google Scholar]
- Le, J.M.; Vilcek, J. Interleukin 6: A multifunctional cytokine regulating immune reactions and the acute phase protein response. Lab. Investig. 1989, 61, 588–602. [Google Scholar] [PubMed]







Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jiao, P.; Wang, J.; Yang, J.; Wang, X.; Luoreng, Z. Bta-miR-223 Targeting the RHOB Gene in Dairy Cows Attenuates LPS-Induced Inflammatory Responses in Mammary Epithelial Cells. Cells 2022, 11, 3144. https://doi.org/10.3390/cells11193144
Jiao P, Wang J, Yang J, Wang X, Luoreng Z. Bta-miR-223 Targeting the RHOB Gene in Dairy Cows Attenuates LPS-Induced Inflammatory Responses in Mammary Epithelial Cells. Cells. 2022; 11(19):3144. https://doi.org/10.3390/cells11193144
Chicago/Turabian StyleJiao, Peng, Jinpeng Wang, Jian Yang, Xingping Wang, and Zhuoma Luoreng. 2022. "Bta-miR-223 Targeting the RHOB Gene in Dairy Cows Attenuates LPS-Induced Inflammatory Responses in Mammary Epithelial Cells" Cells 11, no. 19: 3144. https://doi.org/10.3390/cells11193144