
Exosomes Derived from Adipose Mesenchymal Stem Cells Promote Diabetic Chronic Wound Healing through SIRT3/SOD2

Abstract
:1. Introduction
2. Materials and Methods
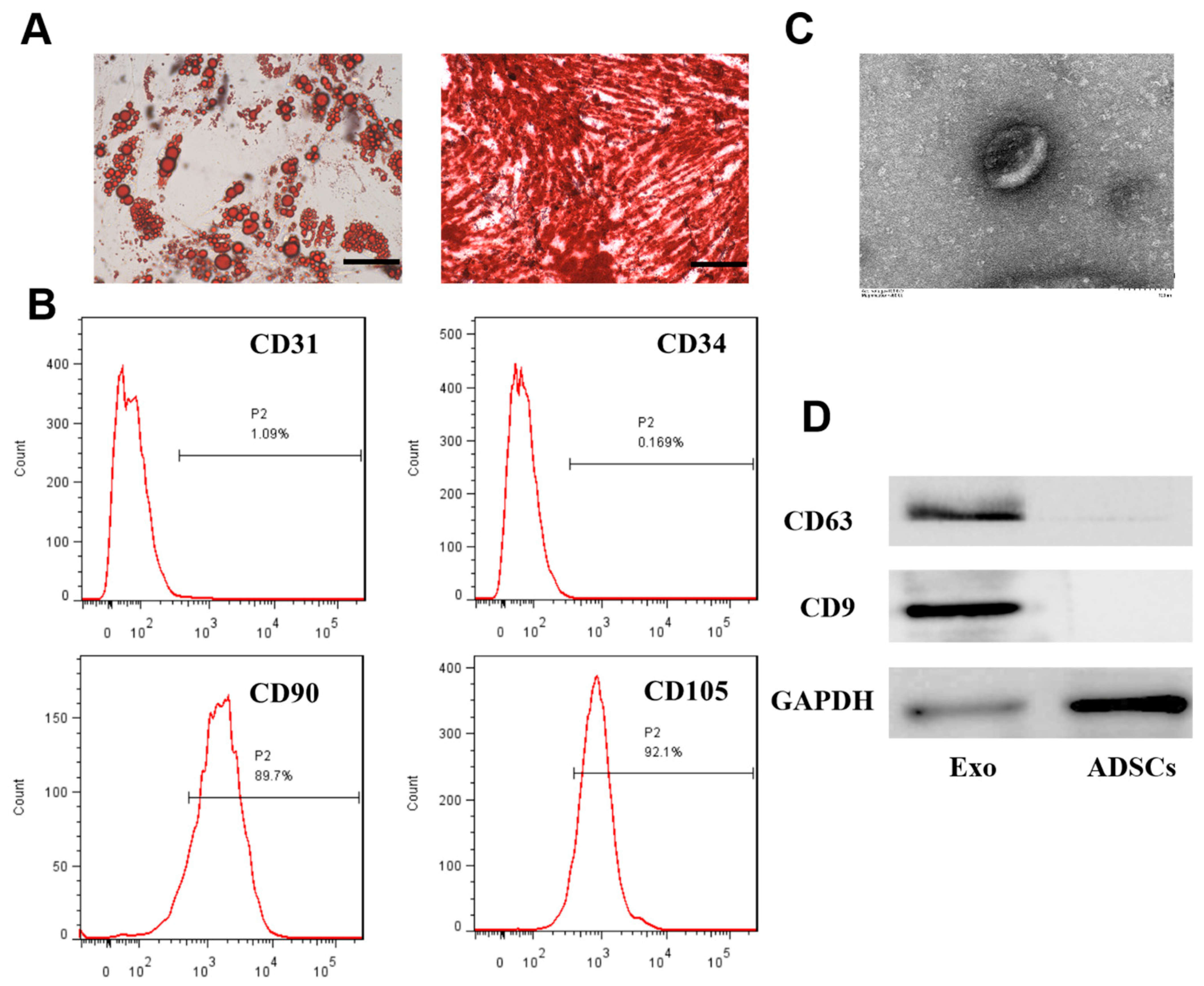
2.1. Isolation, Culture, and Identification of ADSCs
2.2. Exosome Isolation and Identification
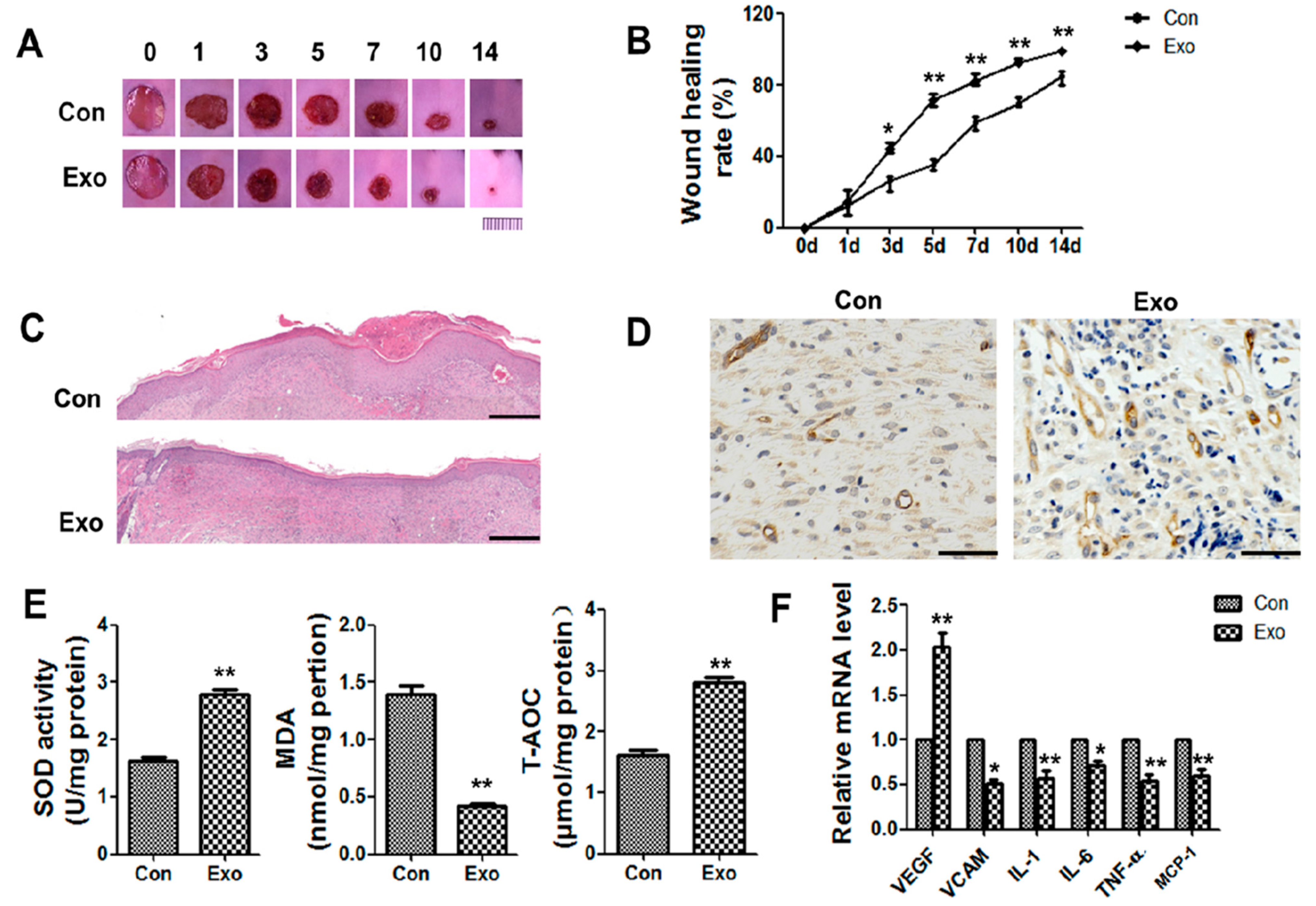
2.3. Animal Models
2.4. Haematoxylin and Eosin (HE) and CD34 Staining
2.5. Measurement of Oxidative Stress in Skin Tissue
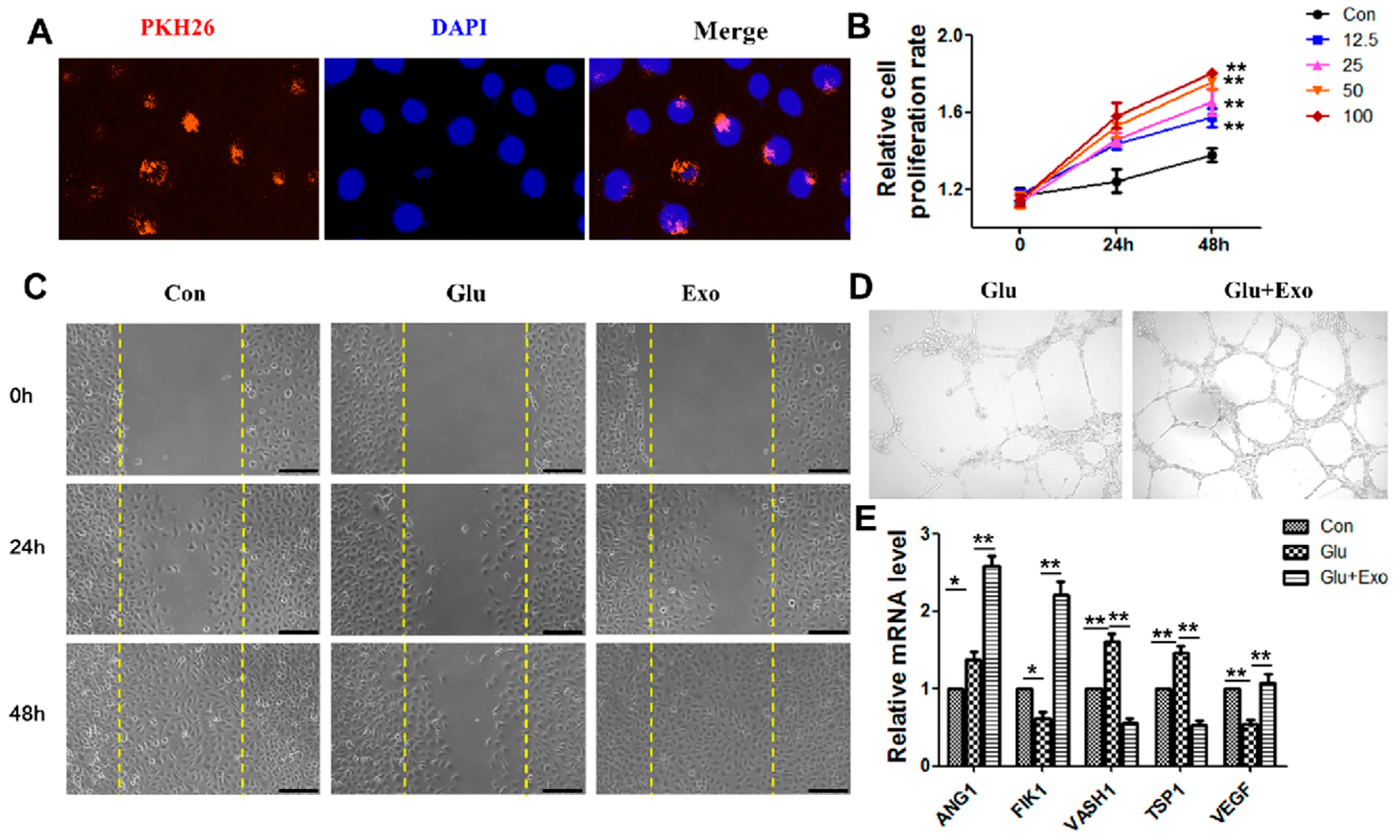
2.6. Cell Culture, Treatment, and Transfection
2.7. HUVECs Proliferation, Migration, and Tube Formation Assays
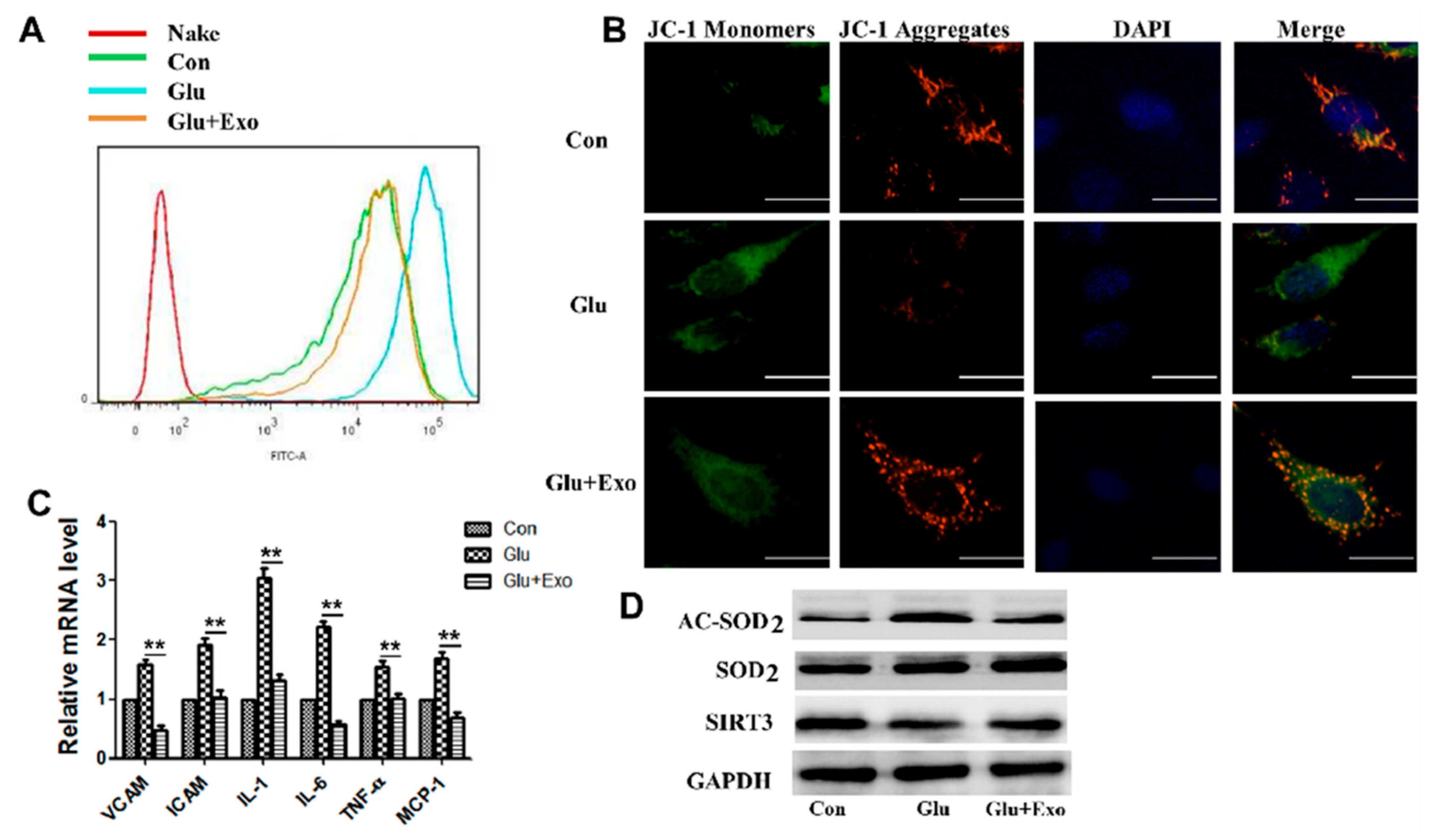
2.8. Measurement of Intracellular ROS
2.9. MMP Assay
2.10. Quantitative RT-PCR
2.11. Western Blot Analysis
2.12. Statistical Analysis
3. Results
3.1. Extraction and Identification of ADSCs and Exosomes
3.2. ADSC-Exos Promote Angiogenesis and Accelerate Wound Closure
3.3. ADSC-Exos Promote the Proliferation, Migration, and Angiogenesis of HUVECs
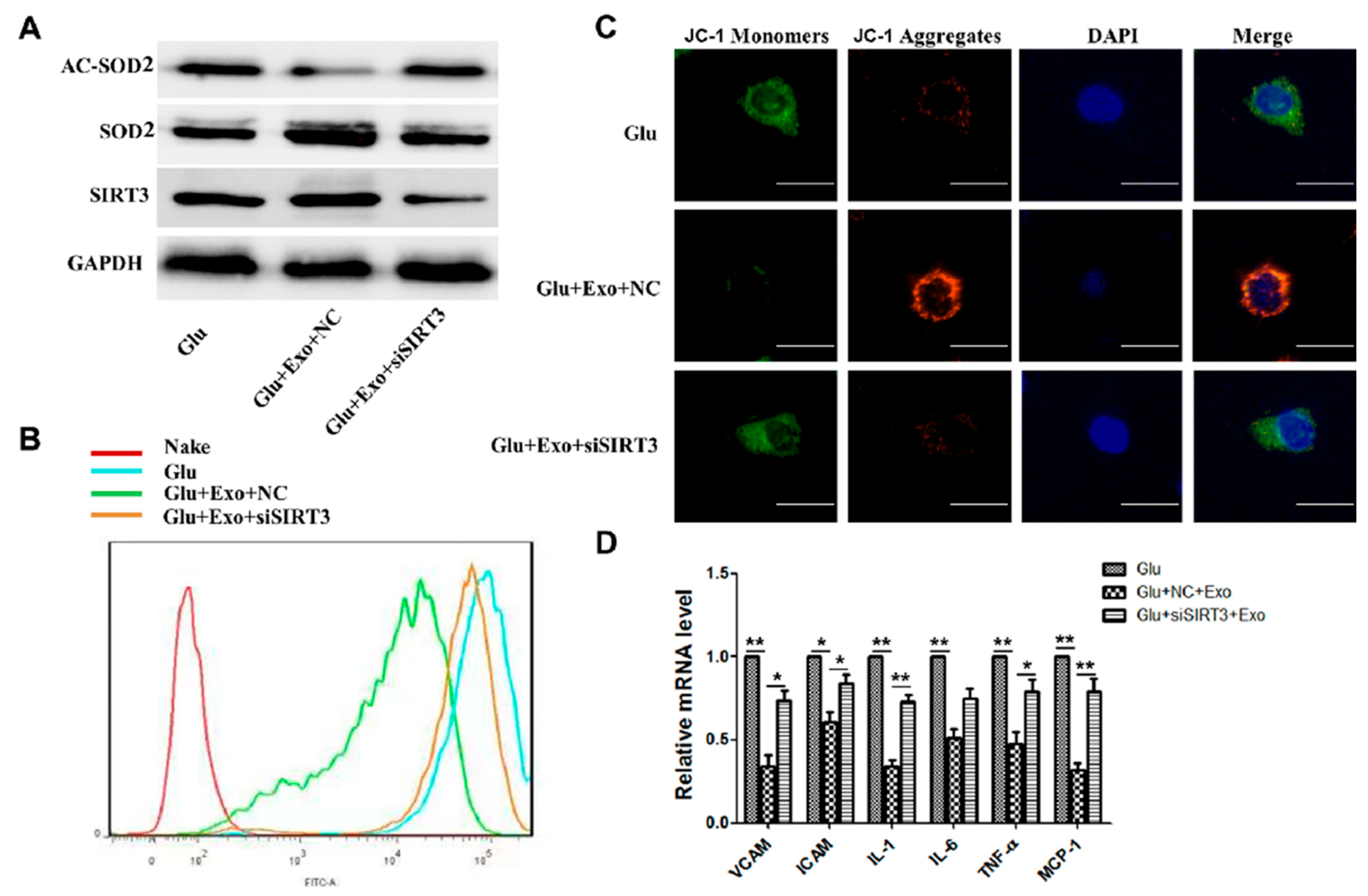
3.4. ADSC-Exos Reduce ROS Production, Protect Mitochondrial Function, and Promote SIRT3 Expression
3.5. ADSC-Exos Reduce ROS Production and Protect Mitochondrial Function through SIRT3
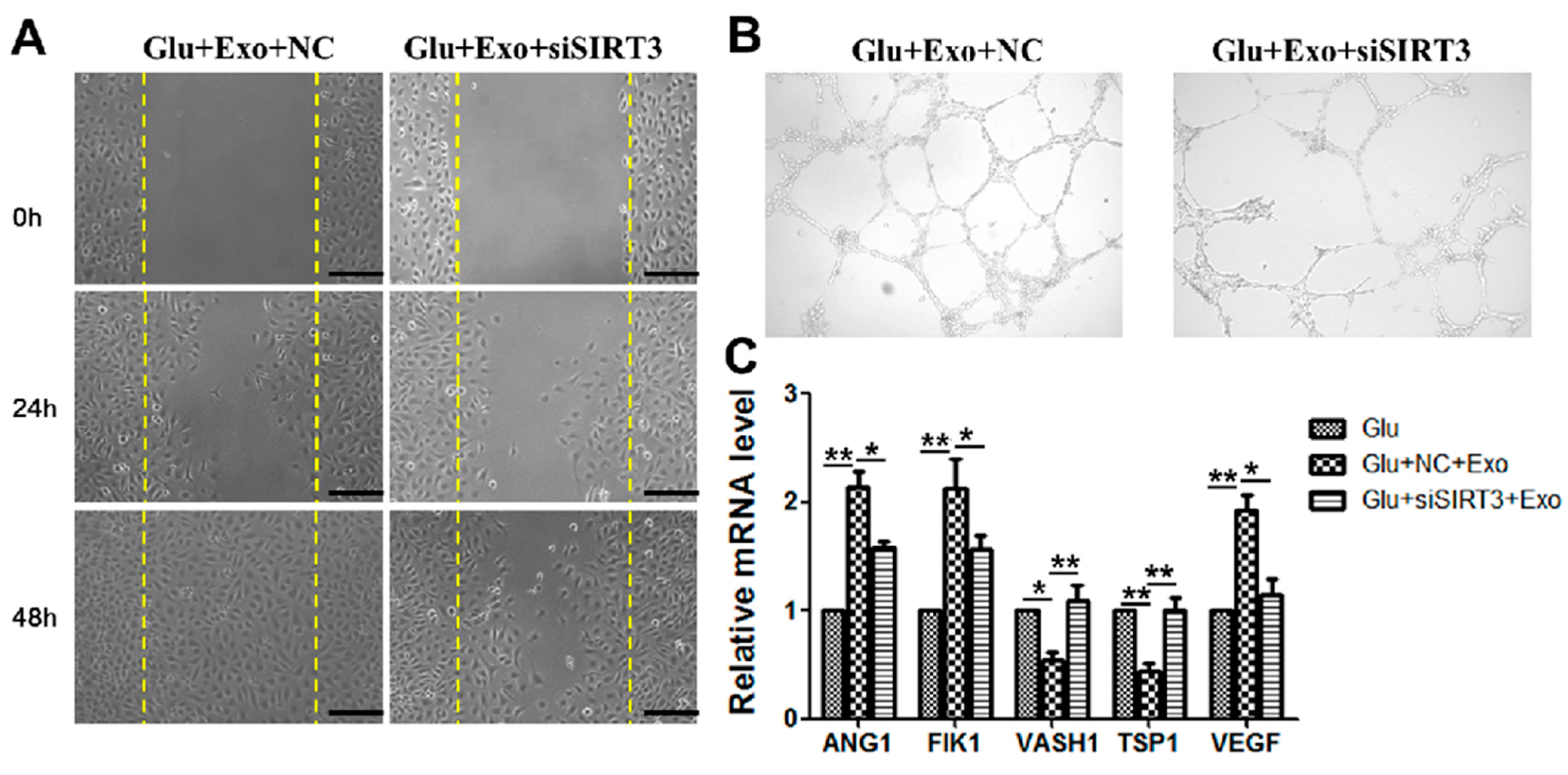
3.6. The Ability of ADSC-Exos Is Weakened When SIRT3 Is Silenced
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Chen, A.; Wang, H.; Su, Y.; Zhang, C.; Qiu, Y.; Zhou, Y.; Wan, Y.; Hu, B.; Li, Y. Exosomes: Biomarkers and Therapeutic Targets of Diabetic Vascular Complications. Front. Endocrinol. 2021, 12, 720466. [Google Scholar] [CrossRef] [PubMed]
- Acosta, J.B.; del Barco, D.G.; Vera, D.C.; Savigne, W.; Lopez-Saura, P.; Nieto, G.G.; Schultz, G.S. The pro-inflammatory environment in recalcitrant diabetic foot wounds. Int. Wound J. 2008, 5, 530–539. [Google Scholar] [CrossRef] [PubMed]
- Rovira-Llopis, S.; Bañuls, C.; Diaz-Morales, N.; Hernandez-Mijares, A.; Rocha, M.; Victor, V.M. Mitochondrial dynamics in type 2 diabetes: Pathophysiological implications. Redox Biol. 2017, 11, 637–645. [Google Scholar] [CrossRef] [PubMed]
- Shi, Y.; Vanhoutte, P.M. Macro- and microvascular endothelial dysfunction in diabetes. J. Diabetes 2017, 9, 434–449. [Google Scholar] [CrossRef] [PubMed]
- Deng, L.; Du, C.; Song, P.; Chen, T.; Rui, S.; Armstrong, D.G.; Deng, W. The Role of Oxidative Stress and Antioxidants in Diabetic Wound Healing. Oxidative Med. Cell. Longev. 2021, 2021, 8852759. [Google Scholar] [CrossRef]
- Xian, D.; Song, J.; Yang, L.; Xiong, X.; Lai, R.; Zhong, J. Emerging Roles of Redox-Mediated Angiogenesis and Oxidative Stress in Dermatoses. Oxidative Med. Cell. Longev. 2019, 2019, 2304018. [Google Scholar] [CrossRef]
- Kalluri, R.; LeBleu, V.S. The biology, function, and biomedical applications of exosomes. Science 2020, 367, eauu6977. [Google Scholar] [CrossRef]
- Li, X.; Xie, X.; Lian, W.; Shi, R.; Han, S.; Zhang, H.; Lu, L.; Li, M. Exosomes from adipose-derived stem cells overexpressing Nrf2 accelerate cutaneous wound healing by promoting vascularization in a diabetic foot ulcer rat model. Exp. Mol. Med. 2018, 50, 1–14. [Google Scholar] [CrossRef]
- Gao, W.; Liu, H.; Yuan, J.; Wu, C.; Huang, D.; Ma, Y.; Zhu, J.; Ma, L.; Guo, J.; Shi, H.; et al. Exosomes derived from mature dendritic cells increase endothelial inflammation and atherosclerosis via membrane TNF-alpha mediated NF-kappaB pathway. J. Cell. Mol. Med. 2016, 20, 2318–2327. [Google Scholar] [CrossRef]
- Zhang, J.; Xiang, H.; Liu, J.; Chen, Y.; He, R.-R.; Liu, B. Mitochondrial Sirtuin 3: New emerging biological function and therapeutic target. Theranostics 2020, 10, 8315–8342. [Google Scholar] [CrossRef]
- Yang, S.; Xu, M.; Meng, G.; Lu, Y. SIRT3 deficiency delays diabetic skin wound healing via oxidative stress and necroptosis enhancement. J. Cell. Mol. Med. 2020, 24, 4415–4427. [Google Scholar] [CrossRef] [PubMed]
- Dikalova, A.E.; Pandey, A.; Xiao, L.; Arslanbaeva, L.; Sidorova, T.; Lopez, M.G.; Billings, F.T.; Verdin, E.; Auwerx, J.; Harrison, D.G.; et al. DikalovMitochondrial Deacetylase Sirt3 Reduces Vascular Dysfunction and Hypertension While Sirt3 Depletion in Essential Hypertension Is Linked to Vascular Inflammation and Oxidative Stress. Circ. Res. 2020, 126, 439–452. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.Z.; Jia, J.; Chen, C.H. lncRNA-KCNQ1OT1: A Potential Target in Exosomes Derived from Adipose-Derived Stem Cells for the Treatment of Osteoporosis. Stem Cells Int. 2021, 2021, 7690006. [Google Scholar] [CrossRef] [PubMed]
- Shen, K.; Jia, Y.; Wang, X.; Zhang, J.; Liu, K.; Wang, J.; Cai, W.; Li, J.; Li, S.; Zhao, M.; et al. Exosomes from adipose-derived stem cells alleviate the inflammation and oxidative stress via regulating Nrf2/HO-1 axis in macrophages. Free Radic. Biol. Med. 2021, 165, 54–66. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Liu, J.; Liu, Y.; Hu, X.; Dong, M.; Wang, H.; Hu, D. Negative pressure wound therapy accelerates rats diabetic wound by promoting agenesis. Int. J. Clin. Exp. Med. 2015, 8, 3506–3513. [Google Scholar]
- Zhang, X.; Jiang, Y.; Huang, Q.; Wu, Z.; Pu, H.; Xu, Z.; Li, B.; Lu, X.; Yang, X.; Qin, J.; et al. Exosomes derived from adipose-derived stem cells overexpressing glyoxalase-1 protect endothelial cells and enhance angiogenesis in type 2 diabetic mice with limb ischemia. Stem. Cell. Res. Ther. 2021, 12, 403. [Google Scholar] [CrossRef]
- Saeedi, P.; Petersohn, I.; Salpea, P.; Malanda, B.; Karuranga, S.; Unwin, N.; Colagiuri, S.; Guariguata, L.; Motala, A.A.; Ogurtsova, K.; et al. Global and regional diabetes prevalence estimates for 2019 and projections for 2030 and 2045: Results from the International Diabetes Federation Diabetes Atlas, 9(th) edition. Diabetes Res. Clin. Pract. 2019, 157, 107843. [Google Scholar] [CrossRef]
- Standl, E.; Khunti, K.; Hansen, T.B.; Schnell, O. The global epidemics of diabetes in the 21st century: Current situation and perspectives. Eur. J. Prev. Cardiol. 2019, 26 (Suppl. S2), 7–14. [Google Scholar] [CrossRef]
- Naderi, N.; Combellack, E.J.; Griffin, M.; Sedaghati, T.; Javed, M.; Findlay, M.W.; Wallace, C.G.; Mosahebi, A.; Butler, P.E.; Seifalian, A.M.; et al. The regenerative role of adipose-derived stem cells (ADSC) in plastic and reconstructive surgery. Int. Wound J. 2017, 14, 112–124. [Google Scholar] [CrossRef]
- Dung, T.N.; Han, V.D.; Tien, G.N.; Lam, H.Q. Autologous Adipose-Derived Stem Cell (Adsc) Transplantation In The Management Of Chronic Wounds. Ann Burn. Fire Disasters 2021, 34, 343–350. [Google Scholar] [CrossRef]
- Lin, J.; Wang, Z.; Huang, J.; Tang, S.; Saiding, Q.; Zhu, Q.; Cui, W. Microenvironment-Protected Exosome-Hydrogel for Facilitating Endometrial Regeneration, Fertility Restoration, and Live Birth of Offspring. Small 2021, 17, e2007235. [Google Scholar] [CrossRef] [PubMed]
- Qiu, X.; Liu, J.; Zheng, C.; Su, Y.; Bao, L.; Zhu, B.; Liu, S.; Wang, L.; Wang, X.; Wang, Y.; et al. Exosomes released from educated mesenchymal stem cells accelerate cutaneous wound healing via promoting angiogenesis. Cell. Prolif. 2020, 53, e12830. [Google Scholar] [CrossRef] [PubMed]
- Qi, H.; Wang, Y.; Fa, S.; Yuan, C.; Yang, L. Extracellular Vesicles as Natural Delivery Carriers Regulate Oxidative Stress Under Pathological Conditions. Front. Bioeng. Biotechnol. 2021, 9, 752019. [Google Scholar] [CrossRef] [PubMed]
- Burgess, J.L.; Wyant, W.A.; Abujamra, B.A.; Kirsner, R.S.; Jozic, I. Diabetic Wound-Healing Science. Medicina 2021, 57, 1072. [Google Scholar] [CrossRef]
- Sangiorgi, S.; Manelli, A.; Reguzzoni, M.; Ronga, M.; Protasoni, M.; Dell’Orbo, C. The cutaneous microvascular architecture of human diabetic toe studied by corrosion casting and scanning electron microscopy analysis. Anat. Rec. 2010, 293, 1639–1645. [Google Scholar] [CrossRef]
- Choke, E.; Tang, T.Y.; Cheng, S.C.; Tay, J.S. Treatment of lower limb ischaemia in patients with diabetes. Diabetes Metab. Res. Rev. 2020, 36 (Suppl. S1), e3262. [Google Scholar] [CrossRef]
- Li, X.; Wu, G.; Han, F.; Wang, K.; Bai, X.; Jia, Y.; Li, Z.; Cai, W.; Zhang, W.; Su, L.; et al. SIRT1 activation promotes angiogenesis in diabetic wounds by protecting endothelial cells against oxidative stress. Arch. Biochem. Biophys. 2019, 661, 117–124. [Google Scholar] [CrossRef]
- Johansen, J.S.; Harris, A.K.; Rychly, D.J.; Ergul, A. Oxidative stress and the use of antioxidants in diabetes: Linking basic science to clinical practice. Cardiovasc. Diabetol. 2005, 4, 5. [Google Scholar] [CrossRef]
- Mittal, M.; Siddiqui, M.R.; Tran, K.; Reddy, S.P.; Malik, A.B. Reactive oxygen species in inflammation and tissue injury. Antioxid. Redox Signal. 2014, 20, 1126–1167. [Google Scholar] [CrossRef]
- El Assar, M.; Angulo, J.; Rodríguez-Mañas, L. Oxidative stress and vascular inflammation in aging. Free Radic. Biol. Med. 2013, 65, 380–401. [Google Scholar] [CrossRef]
- Wolff, P.S.; Jiang, Z.Y.; Hunt, J.V. Protein glycation and oxidative stress in diabetes mellitus and ageing. Free Radic. Biol. Med. 1991, 10, 339–352. [Google Scholar] [CrossRef]
- Zelko, N.I.; Mariani, T.J.; Folz, R.J. Superoxide dismutase multigene family: A comparison of the CuZn-SOD (SOD1), Mn-SOD (SOD2), and EC-SOD (SOD3) gene structures, evolution, and expression. Free Radic. Biol. Med. 2002, 33, 337–349. [Google Scholar] [CrossRef]
- Wang, Y.; Branicky, R.; Noe, A.; Hekimi, S. Superoxide dismutases: Dual roles in controlling ROS damage and regulating ROS signaling. J. Cell. Biol. 2018, 217, 1915–1928. [Google Scholar] [CrossRef] [PubMed]
- Lewandowski, L.; Kepinska, M.; Milnerowicz, H. Alterations in Concentration/Activity of Superoxide Dismutases in Context of Obesity and Selected Single Nucleotide Polymorphisms in Genes: SOD1, SOD2, SOD3. Int. J. Mol. Sci. 2020, 21, 5069. [Google Scholar] [CrossRef] [PubMed]
- Chen, M.L.; Zhu, X.H.; Ran, L.; Lang, H.D.; Yi, L.; Mi, M.T. Trimethylamine-N-Oxide Induces Vascular Inflammation by Activating the NLRP3 Inflammasome Through the SIRT3-SOD2-mtROS Signaling Pathway. J. Am. Heart Assoc. 2017, 6, e006347. [Google Scholar] [CrossRef]
- Sun, W.; Liu, C.; Chen, Q.; Liu, N.; Yan, Y.; Liu, B. SIRT3: A New Regulator of Cardiovascular Diseases. Oxidative Med. Cell. Longev. 2018, 2018, 7293861. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Mouse GAPDH | F:5′-GTGTTCCTACCCCCAATGTG-3′ | R:5′-CATCGAAGGTGGAAGAGTGG-3′ |
| Mouse VEGF | F:5’-GTGGACATCTTCCAGGAGTA-3’ | R:5’-TCTGCATTCACATCTGCTGT-3’ |
| Mouse VCAM | F:5’-GTTCCAGCGAGGGTCTACC-3’ | R:5’-AACTCTTGGCAAACATTAGGTGT-3’ |
| Mouse IL-1 | F:5′-TCCTGTGTAATGAAAGACGGC-3′ | R:5′-TGCTTGTGAGGTGCTGATGTA-3′ |
| Mouse IL-6 | F:5′-GGGACTGATGCTGGTGACAA-3′ | R:5′-TCCACGATTTCCCAGAGAACA-3′ |
| Mouse TNF-α | F:5′-GAACTGGCAGAAGAGGCACT-3′ | R:5′-CATAGAACTGATGAGAGGGAGG-3′ |
| Mouse MCP-1 | F:5′-ACTGCACCCAAACCGAAGTC-3′ | R:5′-TGGGGACACCTTTTAGCATCTT-3′ |
| Human GAPDH | F:5′-ATGGGGAAGGTGAAGGTCG-3′ | R:5′-GGGGTCATTGATGGCAACAATA-3′ |
| Human ANG1 | F:5′-CAGACTGCAGAGCAGACCAGAA-3′ | R:5′-CTCTAGCTTGTAGGTGGATAATGAATTC-3′ |
| Human FLK1: | F:5’-GACTTCCTGACCTTGGAGCATCT-3’ | R:5’-GATTTTAACCACGTTCTTCTCCGA-3′ |
| Human VASH1 | F:5′-AACTACTTCCGCCACATCGT-3′ | R:5′-GGCGGCTTGTACATCAGGTC-3′ |
| Human TSP1 | F:5′- UCCUUCUAGGUGGCCUCAGAC-3′ | R:5′-CATTGGAGCAGGGCATGATGG-3′ |
| Human VEGF | F:5′-GCAGAATCATCACGAAGTGGTG-3′ | R:5′-TCTCGATTGGATGCAGTAGCT-3′ |
| Human VCAM | F:5′-GTAAAAGAATTGCAAGTCTACATATCAC-3′ | R:5′-GATGGATTCACAGAAATAACTGTATTC-3′ |
| Human ICAM | F:5′-AACCAGAGCCAGGAGACACTG -3′ | R:5′-GCGCCGGAAAGCTGTAGATG-3′ |
| Human IL-1 | F:5′-ATGATGGCTTATTACAGTGGCAA -3′ | R:5′-GTCGGAGATTCGTAGCTGGA-3′ |
| Human IL-6 | F:5′-ACTCACCTCTTCAGAACGAATTG-3′ | R:5′-CCATCTTTGGAAGGTTCAGGTTG-3′ |
| Human TNF-α | F:5′-CTATCTGGGAGGGGTCTTCC-3′ | R:5′-GGTTGAGGGTGTCTGAAGGA-3′ |
| Human MCP-1 | F:5′-CAGCCAGATGCAATCAATGCC-3′ | R:5′-TGGAATCCTGAACCCACTTCT-3′ |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, Y.; Bai, X.; Shen, K.; Luo, L.; Zhao, M.; Xu, C.; Jia, Y.; Xiao, D.; Li, Y.; Gao, X.; et al. Exosomes Derived from Adipose Mesenchymal Stem Cells Promote Diabetic Chronic Wound Healing through SIRT3/SOD2. Cells 2022, 11, 2568. https://doi.org/10.3390/cells11162568
Zhang Y, Bai X, Shen K, Luo L, Zhao M, Xu C, Jia Y, Xiao D, Li Y, Gao X, et al. Exosomes Derived from Adipose Mesenchymal Stem Cells Promote Diabetic Chronic Wound Healing through SIRT3/SOD2. Cells. 2022; 11(16):2568. https://doi.org/10.3390/cells11162568
Chicago/Turabian StyleZhang, Yue, Xiaozhi Bai, Kuo Shen, Liang Luo, Ming Zhao, Chaolei Xu, Yanhui Jia, Dan Xiao, Yan Li, Xiaowen Gao, and et al. 2022. "Exosomes Derived from Adipose Mesenchymal Stem Cells Promote Diabetic Chronic Wound Healing through SIRT3/SOD2" Cells 11, no. 16: 2568. https://doi.org/10.3390/cells11162568