
Cannabinoids and PPAR Ligands: The Future in Treatment of Polycystic Ovary Syndrome Women with Obesity and Reduced Fertility



Abstract
:
1. Introduction
2. PCOS
2.1. PCOS and Hormone Imbalance
2.2. PCOS and Obesity, Overweight and Other Metabolic Disturbances
2.3. PCOS and Fertility
3. Cannabinoids and Cannabinoid Receptors
3.1. Cannabinoid Synthesis and Classification
3.1.1. Endocannabinoids
3.1.2. Plant Cannabinoids
3.1.3. Synthetic Cannabinoids
3.2. Structure and Function of Cannabinoid Receptors
3.2.1. Plasma Membrane Receptors
3.2.2. Intracellular Receptors
3.3. Physiological Roles of Cannabinoids
3.3.1. Role of CBs in Carbohydrate Metabolism, Lipid Metabolism, and Obesity
3.3.2. Role of CBs in Embryo Implantation and Female Fertility
3.3.3. Role of CBs in PCOS
4. PPARs
4.1. PPARs and Their Ligands
4.1.1. PPARα
4.1.2. PPARβ/δ
4.1.3. PPARγ
4.2. Physiological Role of PPARs
4.2.1. Role of PPARs in Energy Metabolism and Obesity
4.2.2. Role of PPARs in Embryo Implantation and Female Fertility
4.2.3. Role of PPARs in PCOS
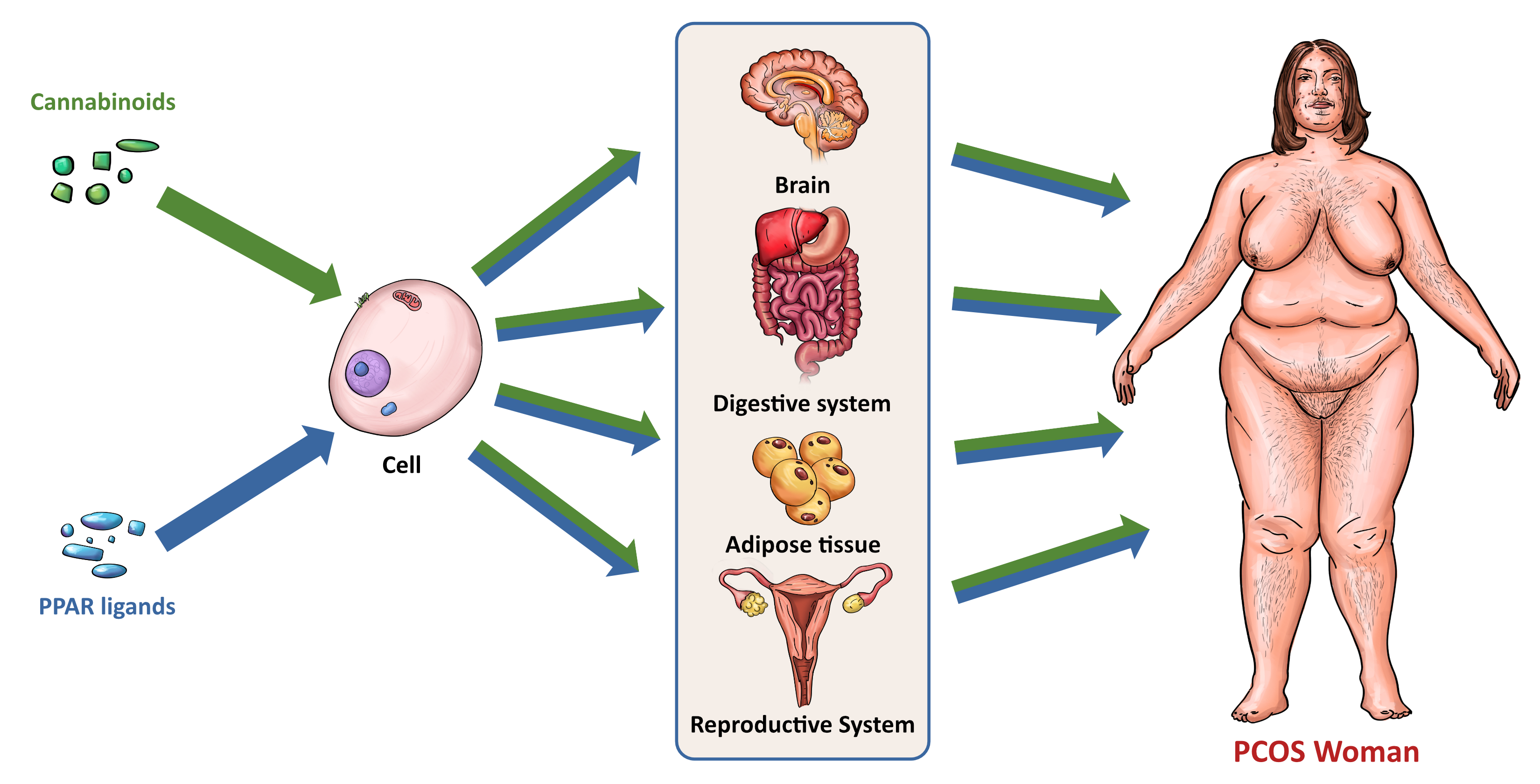
5. Interactions between PPARs and The ECS
5.1. Cannabinoids and Their Metabolites as PPAR Ligands

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Type of PPAR | Cannabinoid or Cannabinoid Metabolite | |
|---|---|---|
| PPARα | WIN 55212-2 [237] noladin ether [237] virodhamine [237] 2-AG [239] AEA [237] 15-HETE glycerol ester [239] CBDA [254] | CBM [256] CBGA [254] PEA [241] OEA [237,242] ODA [243] OLHHA [260,261] |
| PPARβ/δ | OEA [242] ODA [243] | |
| PPARγ | ∆9-THC [246,247] CBC [247] CBD [245,247] CBG [247] 2-AG [238] AEA [240] 15d-PGJ2 glycerol ester [244] WIN 55212-2 [249] ∆9-THCA [248] | ODA [243] CBDA [254] CBM [256] CBGA [254] AJA [252,253] VCE-003 [247] VCE–004.3 [250] VCE–004.8 [251] chromenopyrazole 4 [259] |
5.2. Link of Cannabinoid-Metabolizing Enzymes to PPARs
5.3. Link between Cannabinoids Receptors and PPARs
5.4. Non-Cannabinoid Dual Ligands of CBRs and PPARs
6. Concluding Remarks and Future Directions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Women’s Reproductive Health|CDC. Available online: https://www.cdc.gov/reproductivehealth/womensrh/index.htm (accessed on 23 July 2022).
- Metwally, M.; Ong, K.J.; Ledger, W.L.; Li, T.C. Does High Body Mass Index Increase the Risk of Miscarriage after Spontaneous and Assisted Conception? A Meta-Analysis of the Evidence. Fertil. Steril. 2008, 90, 714–726. [Google Scholar] [CrossRef] [PubMed]
- Carmina, E.; Bucchieri, S.; Esposito, A.; Del Puente, A.; Mansueto, P.; Orio, F.; Di Fede, G.; Rini, G. Abdominal Fat Quantity and Distribution in Women with Polycystic Ovary Syndrome and Extent of Its Relation to Insulin Resistance. J. Clin. Endocrinol. Metab. 2007, 92, 2500–2505. [Google Scholar] [CrossRef]
- Panidis, D.; Macut, D.; Tziomalos, K.; Papadakis, E.; Mikhailidis, K.; Kandaraki, E.A.; Tsourdi, E.A.; Tantanasis, T.; Mavromatidis, G.; Katsikis, I. Prevalence of Metabolic Syndrome in Women with Polycystic Ovary Syndrome. Clin. Endocrinol. 2013, 78, 586–592. [Google Scholar] [CrossRef]
- Yildiz, B.O.; Knochenhauer, E.S.; Azziz, R. Impact of Obesity on the Risk for Polycystic Ovary Syndrome. J. Clin. Endocrinol. Metab. 2008, 93, 162–168. [Google Scholar] [CrossRef] [PubMed]
- PCOS (Polycystic Ovary Syndrome) and Diabetes|CDC. Available online: https://www.cdc.gov/diabetes/basics/pcos.html (accessed on 23 July 2022).
- Rasquin Leon, L.I.; Anastasopoulou, C.; Mayrin, J.V. Polycystic Ovarian Disease; StatPearls: Treasure Island, FL, USA, 2022. [Google Scholar]
- WHO. Obesity: Preventing and Managing the Global Epidemic. Report of a WHO Consultation; WHO: Geneva, Switzerland, 2000; Volume 894. [Google Scholar]
- Obesity and Overweight. Available online: https://www.who.int/news-room/fact-sheets/detail/obesity-and-overweight (accessed on 23 July 2022).
- Salzet, M. Invertebrate Molecular Neuroimmune Processes. Brain Res. Brain Res. Rev. 2000, 34, 69–79. [Google Scholar] [CrossRef]
- Svizenska, I.; Dubovy, P.; Sulcova, A. Cannabinoid Receptors 1 and 2 (CB1 and CB2), Their Distribution, Ligands and Functional Involvement in Nervous System Structures—A Short Review. Pharmacol. Biochem. Behav. 2008, 90, 501–511. [Google Scholar] [CrossRef] [PubMed]
- Muniyappa, R.; Sable, S.; Ouwerkerk, R.; Mari, A.; Gharib, A.M.; Walter, M.; Courville, A.; Hall, G.; Chen, K.Y.; Volkow, N.D.; et al. Metabolic Effects of Chronic Cannabis Smoking. Diabetes Care 2013, 36, 2415–2422. [Google Scholar] [CrossRef]
- Herkenham, M.; Lynn, A.B.; Johnson, M.R.; Melvin, L.S.; de Costa, B.R.; Rice, K.C. Characterization and Localization of Cannabinoid Receptors in Rat Brain: A Quantitative in Vitro Autoradiographic Study. J. Neurosci. 1991, 11, 563–583. [Google Scholar] [CrossRef]
- Fonseca, B.M.; Rebelo, I. Cannabis and Cannabinoids in Reproduction and Fertility: Where We Stand. Reprod. Sci. 2021. [Google Scholar] [CrossRef]
- du Plessis, S.S.; Agarwal, A.; Syriac, A. Marijuana, Phytocannabinoids, the Endocannabinoid System, and Male Fertility. J. Assist. Reprod. Genet. 2015, 32, 1575–1588. [Google Scholar] [CrossRef]
- El-Talatini, M.R.; Taylor, A.H.; Elson, J.C.; Brown, L.; Davidson, A.C.; Konje, J.C. Localisation and Function of the Endocannabinoid System in the Human Ovary. PLoS ONE 2009, 4, e4579. [Google Scholar] [CrossRef] [PubMed]
- Horne, A.W.; Phillips, J.A., 3rd; Kane, N.; Lourenco, P.C.; McDonald, S.E.; Williams, A.R.W.; Simon, C.; Dey, S.K.; Critchley, H.O.D. CB1 Expression Is Attenuated in Fallopian Tube and Decidua of Women with Ectopic Pregnancy. PLoS ONE 2008, 3, e3969. [Google Scholar] [CrossRef] [PubMed]
- Taylor, A.H.; Abbas, M.S.; Habiba, M.A.; Konje, J.C. Histomorphometric Evaluation of Cannabinoid Receptor and Anandamide Modulating Enzyme Expression in the Human Endometrium through the Menstrual Cycle. Histochem. Cell Biol. 2010, 133, 557–565. [Google Scholar] [CrossRef] [PubMed]
- Brighton, P.J.; Marczylo, T.H.; Rana, S.; Konje, J.C.; Willets, J.M. Characterization of the Endocannabinoid System, CB(1) Receptor Signalling and Desensitization in Human Myometrium. Br. J. Pharmacol. 2011, 164, 1479–1494. [Google Scholar] [CrossRef]
- Taylor, A.H.; Ang, C.; Bell, S.C.; Konje, J.C. The Role of the Endocannabinoid System in Gametogenesis, Implantation and Early Pregnancy. Hum. Reprod. Update 2007, 13, 501–513. [Google Scholar] [CrossRef]
- Battista, N.; Bari, M.; Rapino, C.; Trasatti, F.; D’Agostino, A.; Maccarrone, M. Regulation of Female Fertility by the Endocannabinoid System. Hum. Fertil. 2007, 10, 207–216. [Google Scholar] [CrossRef]
- Battista, N.; Pasquariello, N.; Di Tommaso, M.; Maccarrone, M. Interplay between Endocannabinoids, Steroids and Cytokines in the Control of Human Reproduction. J. Neuroendocrinol. 2008, 20 (Suppl. 1), 82–89. [Google Scholar] [CrossRef]
- Yousefnia, S.; Momenzadeh, S.; Seyed Forootan, F.; Ghaedi, K.; Nasr Esfahani, M.H. The Influence of Peroxisome Proliferator-Activated Receptor γ (PPARγ) Ligands on Cancer Cell Tumorigenicity. Gene 2018, 649, 14–22. [Google Scholar] [CrossRef]
- Polvani, S.; Tarocchi, M.; Tempesti, S.; Bencini, L.; Galli, A. Peroxisome Proliferator Activated Receptors at the Crossroad of Obesity, Diabetes, and Pancreatic Cancer. World J. Gastroenterol. 2016, 22, 2441–2459. [Google Scholar] [CrossRef]
- Corrales, P.; Vidal-Puig, A.; Medina-Gomez, G. PPARs and Metabolic Disorders Associated with Challenged Adipose Tissue Plasticity. Int. J. Mol. Sci. 2018, 19, 2124. [Google Scholar] [CrossRef]
- Houri, M.; Pratley, R. Rimonabant: A Novel Treatment for Obesity and the Metabolic Syndrome. Curr. Diabetes Rep. 2005, 5, 43–44. [Google Scholar] [CrossRef]
- Rotterdam ESHRE/ASRM-Sponsored PCOS Consensus Workshop Group. Revised 2003 Consensus on Diagnostic Criteria and Long-Term Health Risks Related to Polycystic Ovary Syndrome. Fertil. Steril. 2004, 81, 19–25. [Google Scholar] [CrossRef]
- Teede, H.J.; Misso, M.L.; Costello, M.F.; Dokras, A.; Laven, J.; Moran, L.; Piltonen, T.; Norman, R.J. Recommendations from the International Evidence-Based Guideline for the Assessment and Management of Polycystic Ovary Syndrome. Fertil. Steril. 2018, 110, 364–379. [Google Scholar] [CrossRef]
- National Institutes of Health. Evidence-Based Methodology Workshop on Polycystic Ovary Syndrome. December 3–5, 2012. Executive Summary. Final Report; National Institutes of Health: Bethesda, MD, USA, 2012. [Google Scholar]
- Sachdeva, G.; Gainder, S.; Suri, V.; Sachdeva, N.; Chopra, S. Comparison of the Different PCOS Phenotypes Based on Clinical Metabolic, and Hormonal Profile, and Their Response to Clomiphene. Indian J. Endocrinol. Metab. 2019, 23, 326–331. [Google Scholar] [CrossRef] [PubMed]
- Livadas, S.; Pappas, C.; Karachalios, A.; Marinakis, E.; Tolia, N.; Drakou, M.; Kaldrymides, P.; Panidis, D.; Diamanti-Kandarakis, E. Prevalence and Impact of Hyperandrogenemia in 1218 Women with Polycystic Ovary Syndrome. Endocrine 2014, 47, 631–638. [Google Scholar] [CrossRef] [PubMed]
- Rosenfield, R.L.; Ehrmann, D.A. The Pathogenesis of Polycystic Ovary Syndrome (PCOS): The Hypothesis of PCOS as Functional Ovarian Hyperandrogenism Revisited. Endocr. Rev. 2016, 37, 467–520. [Google Scholar] [CrossRef] [PubMed]
- Ye, W.; Xie, T.; Song, Y.; Zhou, L. The Role of Androgen and Its Related Signals in PCOS. J. Cell. Mol. Med. 2021, 25, 1825–1837. [Google Scholar] [CrossRef]
- Taylor, A.E.; McCourt, B.; Martin, K.A.; Anderson, E.J.; Adams, J.M.; Schoenfeld, D.; Hall, J.E. Determinants of Abnormal Gonadotropin Secretion in Clinically Defined Women with Polycystic Ovary Syndrome. J. Clin. Endocrinol. Metab. 1997, 82, 2248–2256. [Google Scholar] [CrossRef]
- Scotf, J.R.; Gibbs, R.S.; Karlan, B.Y.; Haney, A.F. Danforth’s Obstetrics and Gynecology; Lippincott Williams & Wilkins: Philadelphia, PA, USA, 2003. [Google Scholar]
- Stener-Victorin, E.; Padmanabhan, V.; Walters, K.A.; Campbell, R.E.; Benrick, A.; Giacobini, P.; Dumesic, D.A.; Abbott, D.H. Animal Models to Understand the Etiology and Pathophysiology of Polycystic Ovary Syndrome. Endocr. Rev. 2020, 41, 538–576. [Google Scholar] [CrossRef]
- Paschou, S.A.; Palioura, E.; Ioannidis, D.; Anagnostis, P.; Panagiotakou, A.; Loi, V.; Karageorgos, G.; Goulis, D.G.; Vryonidou, A. Adrenal Hyperandrogenism Does Not Deteriorate Insulin Resistance and Lipid Profile in Women with PCOS. Endocr. Connect. 2017, 6, 601–606. [Google Scholar] [CrossRef]
- Legro, R.S.; Kunselman, A.R.; Dodson, W.C.; Dunaif, A. Prevalence and Predictors of Risk for Type 2 Diabetes Mellitus and Impaired Glucose Tolerance in Polycystic Ovary Syndrome: A Prospective, Controlled Study in 254 Affected Women. J. Clin. Endocrinol. Metab. 1999, 84, 165–169. [Google Scholar] [CrossRef] [PubMed]
- Broughton, D.E.; Moley, K.H. Obesity and Female Infertility: Potential Mediators of Obesity’s Impact. Fertil. Steril. 2017, 107, 840–847. [Google Scholar] [CrossRef] [PubMed]
- Moran, L.J.; Norman, R.J.; Teede, H.J. Metabolic Risk in PCOS: Phenotype and Adiposity Impact. Trends Endocrinol. Metab. 2015, 26, 136–143. [Google Scholar] [CrossRef] [PubMed]
- Diamanti-Kandarakis, E.; Dunaif, A. Insulin Resistance and the Polycystic Ovary Syndrome Revisited: An Update on Mechanisms and Implications. Endocr. Rev. 2012, 33, 981–1030. [Google Scholar] [CrossRef]
- Ehrmann, D.A.; Liljenquist, D.R.; Kasza, K.; Azziz, R.; Legro, R.S.; Ghazzi, M.N. Prevalence and Predictors of the Metabolic Syndrome in Women with Polycystic Ovary Syndrome. J. Clin. Endocrinol. Metab. 2006, 91, 48–53. [Google Scholar] [CrossRef] [PubMed]
- Robinson, S.; Kiddy, D.; Gelding, S.V.; Willis, D.; Niththyananthan, R.; Bush, A.; Johnston, D.G.; Franks, S. The Relationship of Insulin Insensitivity to Menstrual Pattern in Women with Hyperandrogenism and Polycystic Ovaries. Clin. Endocrinol. 1993, 39, 351–355. [Google Scholar] [CrossRef]
- Merhi, Z. Advanced Glycation End Products and Their Relevance in Female Reproduction. Hum. Reprod. 2014, 29, 135–145. [Google Scholar] [CrossRef]
- Legro, R.S.; Kunselman, A.R.; Dunaif, A. Prevalence and Predictors of Dyslipidemia in Women with Polycystic Ovary Syndrome. Am. J. Med. 2001, 111, 607–613. [Google Scholar] [CrossRef]
- Cerda, C.; Pérez-Ayuso, R.M.; Riquelme, A.; Soza, A.; Villaseca, P.; Sir-Petermann, T.; Espinoza, M.; Pizarro, M.; Solis, N.; Miquel, J.F.; et al. Nonalcoholic Fatty Liver Disease in Women with Polycystic Ovary Syndrome. J. Hepatol. 2007, 47, 412–417. [Google Scholar] [CrossRef]
- Pigny, P.; Jonard, S.; Robert, Y.; Dewailly, D. Serum Anti-Mullerian Hormone as a Surrogate for Antral Follicle Count for Definition of the Polycystic Ovary Syndrome. J. Clin. Endocrinol. Metab. 2006, 91, 941–945. [Google Scholar] [CrossRef]
- Strowitzki, T.; Capp, E.; von Eye Corleta, H. The Degree of Cycle Irregularity Correlates with the Grade of Endocrine and Metabolic Disorders in PCOS Patients. Eur. J. Obstet. Gynecol. Reprod. Biol. 2010, 149, 178–181. [Google Scholar] [CrossRef] [PubMed]
- NCD Risk Factor Collaboration (NCD-RisC). Trends in Adult Body-Mass Index in 200 Countries from 1975 to 2014: A Pooled Analysis of 1698 Population-Based Measurement Studies with 19.2 Million Participants. Lancet 2016, 387, 1377–1396. [Google Scholar] [CrossRef]
- Michalakis, K.; Mintziori, G.; Kaprara, A.; Tarlatzis, B.C.; Goulis, D.G. The Complex Interaction between Obesity, Metabolic Syndrome and Reproductive Axis: A Narrative Review. Metabolism. 2013, 62, 457–478. [Google Scholar] [CrossRef] [PubMed]
- Quinkler, M.; Sinha, B.; Tomlinson, J.W.; Bujalska, I.J.; Stewart, P.M.; Arlt, W. Androgen Generation in Adipose Tissue in Women with Simple Obesity—A Site-Specific Role for 17beta-Hydroxysteroid Dehydrogenase Type 5. J. Endocrinol. 2004, 183, 331–342. [Google Scholar] [CrossRef] [PubMed]
- Santoro, N.; Torrens, J.; Crawford, S.; Allsworth, J.E.; Finkelstein, J.S.; Gold, E.B.; Korenman, S.; Lasley, W.L.; Luborsky, J.L.; McConnell, D.; et al. Correlates of Circulating Androgens in Mid-Life Women: The Study of Women’s Health across the Nation. J. Clin. Endocrinol. Metab. 2005, 90, 4836–4845. [Google Scholar] [CrossRef] [PubMed]
- Freeman, A.M.; Pennings, N. Insulin Resistance; StatPearls: Treasure Island, FL, USA, 2022. [Google Scholar]
- Agarwal, S.K.; Vogel, K.; Weitsman, S.R.; Magoffin, D.A. Leptin Antagonizes the Insulin-like Growth Factor-I Augmentation of Steroidogenesis in Granulosa and Theca Cells of the Human Ovary. J. Clin. Endocrinol. Metab. 1999, 84, 1072–1076. [Google Scholar] [CrossRef] [PubMed]
- Mulder, C.L.; Lassi, Z.S.; Grieger, J.A.; Ali, A.; Jankovic-Karasoulos, T.; Roberts, C.T.; Andraweera, P.H. Cardio-Metabolic Risk Factors among Young Infertile Women: A Systematic Review and Meta-Analysis. BJOG 2020, 127, 930–939. [Google Scholar] [CrossRef]
- Palomba, S. Is Fertility Reduced in Ovulatory Women with Polycystic Ovary Syndrome? An Opinion Paper. Hum. Reprod. 2021, 36, 2421–2428. [Google Scholar] [CrossRef]
- Palomba, S.; Russo, T.; Falbo, A.; Di Cello, A.; Tolino, A.; Tucci, L.; La Sala, G.B.; Zullo, F. Macroscopic and Microscopic Findings of the Placenta in Women with Polycystic Ovary Syndrome. Hum. Reprod. 2013, 28, 2838–2847. [Google Scholar] [CrossRef]
- Palomba, S.; Daolio, J.; La Sala, G.B. Oocyte Competence in Women with Polycystic Ovary Syndrome. Trends Endocrinol. Metab. 2017, 28, 186–198. [Google Scholar] [CrossRef]
- Palomba, S.; Piltonen, T.T.; Giudice, L.C. Endometrial Function in Women with Polycystic Ovary Syndrome: A Comprehensive Review. Hum. Reprod. Update 2021, 27, 584–618. [Google Scholar] [CrossRef] [PubMed]
- Xue, Z.; Li, J.; Feng, J.; Han, H.; Zhao, J.; Zhang, J.; Han, Y.; Wu, X.; Zhang, Y. Research Progress on the Mechanism between Polycystic Ovary Syndrome and Abnormal Endometrium. Front. Physiol. 2021, 12, 788772. [Google Scholar] [CrossRef] [PubMed]
- Palomba, S.; de Wilde, M.A.; Falbo, A.; Koster, M.P.H.; La Sala, G.B.; Fauser, B.C.J.M. Pregnancy Complications in Women with Polycystic Ovary Syndrome. Hum. Reprod. Update 2015, 21, 575–592. [Google Scholar] [CrossRef]
- Palomba, S.; Falbo, A.; Zullo, F.; Orio, F.J. Evidence-Based and Potential Benefits of Metformin in the Polycystic Ovary Syndrome: A Comprehensive Review. Endocr. Rev. 2009, 30, 1–50. [Google Scholar] [CrossRef] [PubMed]
- Palomba, S.; Falbo, A.; Russo, T.; Tolino, A.; Orio, F.; Zullo, F. Pregnancy in Women with Polycystic Ovary Syndrome: The Effect of Different Phenotypes and Features on Obstetric and Neonatal Outcomes. Fertil. Steril. 2010, 94, 1805–1811. [Google Scholar] [CrossRef] [PubMed]
- Devane, W.A.; Hanus, L.; Breuer, A.; Pertwee, R.G.; Stevenson, L.A.; Griffin, G.; Gibson, D.; Mandelbaum, A.; Etinger, A.; Mechoulam, R. Isolation and Structure of a Brain Constituent That Binds to the Cannabinoid Receptor. Science 1992, 258, 1946–1949. [Google Scholar] [CrossRef]
- Sugiura, T.; Kondo, S.; Sukagawa, A.; Nakane, S.; Shinoda, A.; Itoh, K.; Yamashita, A.; Waku, K. 2-Arachidonoylglycerol: A Possible Endogenous Cannabinoid Receptor Ligand in Brain. Biochem. Biophys. Res. Commun. 1995, 215, 89–97. [Google Scholar] [CrossRef]
- Fonseca, B.M.; Costa, M.A.; Almada, M.; Correia-da-Silva, G.; Teixeira, N.A. Endogenous Cannabinoids Revisited: A Biochemistry Perspective. Prostaglandins Other Lipid Mediat. 2013, 102–103, 13–30. [Google Scholar] [CrossRef]
- Tagliamonte, S.; Gill, C.I.R.; Pourshahidi, L.K.; Slevin, M.M.; Price, R.K.; Ferracane, R.; Lawther, R.; O’Connor, G.; Vitaglione, P. Endocannabinoids, Endocannabinoid-like Molecules and Their Precursors in Human Small Intestinal Lumen and Plasma: Does Diet Affect Them? Eur. J. Nutr. 2021, 60, 2203–2215. [Google Scholar] [CrossRef]
- Liu, J.; Wang, L.; Harvey-White, J.; Huang, B.X.; Kim, H.-Y.; Luquet, S.; Palmiter, R.D.; Krystal, G.; Rai, R.; Mahadevan, A.; et al. Multiple Pathways Involved in the Biosynthesis of Anandamide. Neuropharmacology 2008, 54, 1–7. [Google Scholar] [CrossRef]
- Okamoto, Y.; Morishita, J.; Tsuboi, K.; Tonai, T.; Ueda, N. Molecular Characterization of a Phospholipase D Generating Anandamide and Its Congeners. J. Biol. Chem. 2004, 279, 5298–5305. [Google Scholar] [CrossRef] [PubMed]
- Hansen, H.S.; Moesgaard, B.; Hansen, H.H.; Schousboe, A.; Petersen, G. Formation of N-Acyl-Phosphatidylethanolamine and N-Acylethanolamine (Including Anandamide) during Glutamate-Induced Neurotoxicity. Lipids 1999, 34, S327–S330. [Google Scholar] [CrossRef] [PubMed]
- Simon, G.M.; Cravatt, B.F. Anandamide Biosynthesis Catalyzed by the Phosphodiesterase GDE1 and Detection of Glycerophospho-N-Acyl Ethanolamine Precursors in Mouse Brain. J. Biol. Chem. 2008, 283, 9341–9349. [Google Scholar] [CrossRef] [PubMed]
- Deutsch, D.G.; Chin, S.A. Enzymatic Synthesis and Degradation of Anandamide, a Cannabinoid Receptor Agonist. Biochem. Pharmacol. 1993, 46, 791–796. [Google Scholar] [CrossRef]
- Rouzer, C.A.; Marnett, L.J. Endocannabinoid Oxygenation by Cyclooxygenases, Lipoxygenases, and Cytochromes P450: Cross-Talk between the Eicosanoid and Endocannabinoid Signaling Pathways. Chem. Rev. 2011, 111, 5899–5921. [Google Scholar] [CrossRef]
- Smith, W.L.; Urade, Y.; Jakobsson, P.-J. Enzymes of the Cyclooxygenase Pathways of Prostanoid Biosynthesis. Chem. Rev. 2011, 111, 5821–5865. [Google Scholar] [CrossRef]
- Sugiura, T.; Kishimoto, S.; Oka, S.; Gokoh, M. Biochemistry, Pharmacology and Physiology of 2-Arachidonoylglycerol, an Endogenous Cannabinoid Receptor Ligand. Prog. Lipid Res. 2006, 45, 405–446. [Google Scholar] [CrossRef]
- Carrier, E.J.; Kearn, C.S.; Barkmeier, A.J.; Breese, N.M.; Yang, W.; Nithipatikom, K.; Pfister, S.L.; Campbell, W.B.; Hillard, C.J. Cultured Rat Microglial Cells Synthesize the Endocannabinoid 2-Arachidonylglycerol, Which Increases Proliferation via a CB2 Receptor-Dependent Mechanism. Mol. Pharmacol. 2004, 65, 999–1007. [Google Scholar] [CrossRef]
- Bisogno, T.; Melck, D.; De Petrocellis, L.; Di Marzo, V. Phosphatidic Acid as the Biosynthetic Precursor of the Endocannabinoid 2-Arachidonoylglycerol in Intact Mouse Neuroblastoma Cells Stimulated with Ionomycin. J. Neurochem. 1999, 72, 2113–2119. [Google Scholar] [CrossRef]
- Oka, S.; Yanagimoto, S.; Ikeda, S.; Gokoh, M.; Kishimoto, S.; Waku, K.; Ishima, Y.; Sugiura, T. Evidence for the Involvement of the Cannabinoid CB2 Receptor and Its Endogenous Ligand 2-Arachidonoylglycerol in 12-O-Tetradecanoylphorbol-13-Acetate-Induced Acute Inflammation in Mouse Ear. J. Biol. Chem. 2005, 280, 18488–18497. [Google Scholar] [CrossRef]
- Bisogno, T.; Howell, F.; Williams, G.; Minassi, A.; Cascio, M.G.; Ligresti, A.; Matias, I.; Schiano-Moriello, A.; Paul, P.; Williams, E.-J.; et al. Cloning of the First Sn1-DAG Lipases Points to the Spatial and Temporal Regulation of Endocannabinoid Signaling in the Brain. J. Cell Biol. 2003, 163, 463–468. [Google Scholar] [CrossRef] [PubMed]
- Tsuboi, K.; Uyama, T.; Okamoto, Y.; Ueda, N. Endocannabinoids and Related N-Acylethanolamines: Biological Activities and Metabolism. Inflamm. Regen. 2018, 38, 28. [Google Scholar] [CrossRef] [PubMed]
- Maccarrone, M. Metabolism of the Endocannabinoid Anandamide: Open Questions after 25 Years. Front. Mol. Neurosci. 2017, 10, 166. [Google Scholar] [CrossRef]
- Mackie, K. Cannabinoid Receptors: Where They Are and What They Do. J. Neuroendocrinol. 2008, 20 (Suppl. 1), 10–14. [Google Scholar] [CrossRef] [PubMed]
- Thomas, B.; ElSohly, M. The Botany of Cannabis Sativa L. In The Analytical Chemistry of Cannabis; Elsevier: Amsterdam, The Netherlands, 2016; pp. 1–26. ISBN 9780128046463. [Google Scholar]
- Monti, M.C.; Frei, P.; Weber, S.; Scheurer, E.; Mercer-Chalmers-Bender, K. Beyond Δ9-Tetrahydrocannabinol and Cannabidiol: Chemical Differentiation of Cannabis Varieties Applying Targeted and Untargeted Analysis. Anal. Bioanal. Chem. 2022, 414, 3847–3862. [Google Scholar] [CrossRef] [PubMed]
- Ng, T.; Gupta, V. Tetrahydrocannabinol (THC); StatPearls: Treasure Island, FL, USA, 2022. [Google Scholar]
- World Drug Report 2021. Available online: https://www.unodc.org/unodc/en/data-and-analysis/wdr2021.html (accessed on 23 July 2022).
- Meissner, H.; Cascella, M. Cannabidiol (CBD); StatPearls: Treasure Island, FL, USA, 2022. [Google Scholar]
- Alves, V.L.; Gonçalves, J.L.; Aguiar, J.; Teixeira, H.M.; Câmara, J.S. The Synthetic Cannabinoids Phenomenon: From Structure to Toxicological Properties: A Review. Crit. Rev. Toxicol. 2020, 50, 359–382. [Google Scholar] [CrossRef]
- Hoehe, M.R.; Caenazzo, L.; Martinez, M.M.; Hsieh, W.T.; Modi, W.S.; Gershon, E.S.; Bonner, T.I. Genetic and Physical Mapping of the Human Cannabinoid Receptor Gene to Chromosome 6q14-Q15. New Biol. 1991, 3, 880–885. [Google Scholar]
- CNR1 Cannabinoid Receptor 1 [Homo Sapiens (Human)]—Gene—NCBI. Available online: https://www.ncbi.nlm.nih.gov/gene/1268 (accessed on 23 July 2022).
- González-Mariscal, I.; Krzysik-Walker, S.M.; Doyle, M.E.; Liu, Q.-R.; Cimbro, R.; Santa-Cruz Calvo, S.; Ghosh, S.; Cieśla, Ł.; Moaddel, R.; Carlson, O.D.; et al. Human CB1 Receptor Isoforms, Present in Hepatocytes and β-Cells, Are Involved in Regulating Metabolism. Sci. Rep. 2016, 6, 33302. [Google Scholar] [CrossRef]
- Munro, S.; Thomas, K.L.; Abu-Shaar, M. Molecular Characterization of a Peripheral Receptor for Cannabinoids. Nature 1993, 365, 61–65. [Google Scholar] [CrossRef]
- CNR2 Cannabinoid Receptor 2 [Homo Sapiens (Human)]—Gene—NCBI. Available online: https://www.ncbi.nlm.nih.gov/gene/1269 (accessed on 23 July 2022).
- Leo, L.M.; Abood, M.E. CB1 Cannabinoid Receptor Signaling and Biased Signaling. Molecules 2021, 26, 5413. [Google Scholar] [CrossRef]
- Howlett, A.C.; Barth, F.; Bonner, T.I.; Cabral, G.; Casellas, P.; Devane, W.A.; Felder, C.C.; Herkenham, M.; Mackie, K.; Martin, B.R.; et al. International Union of Pharmacology. XXVII. Classification of Cannabinoid Receptors. Pharmacol. Rev. 2002, 54, 161–202. [Google Scholar] [CrossRef] [PubMed]
- Wilson, R.I.; Nicoll, R.A. Endocannabinoid Signaling in the Brain. Science 2002, 296, 678–682. [Google Scholar] [CrossRef] [PubMed]
- Dietel, E.; Brobeil, A.; Gattenlohner, S.; Wimmer, M. The Importance of the Right Framework: Mitogen-Activated Protein Kinase Pathway and the Scaffolding Protein PTPIP51. Int. J. Mol. Sci. 2018, 19, 3282. [Google Scholar] [CrossRef]
- Ye, L.; Cao, Z.; Wang, W.; Zhou, N. New Insights in Cannabinoid Receptor Structure and Signaling. Curr. Mol. Pharmacol. 2019, 12, 239–248. [Google Scholar] [CrossRef]
- Stasiłowicz, A.; Tomala, A.; Podolak, I.; Cielecka-Piontek, J. Cannabis Sativa L. as a Natural Drug Meeting the Criteria of a Multitarget Approach to Treatment. Int. J. Mol. Sci. 2021, 22, 778. [Google Scholar] [CrossRef] [PubMed]
- Rozenfeld, R.; Devi, L.A. Regulation of CB1 Cannabinoid Receptor Trafficking by the Adaptor Protein AP-3. FASEB J. Off. Publ. Fed. Am. Soc. Exp. Biol. 2008, 22, 2311–2322. [Google Scholar] [CrossRef]
- Grimsey, N.L.; Graham, E.S.; Dragunow, M.; Glass, M. Cannabinoid Receptor 1 Trafficking and the Role of the Intracellular Pool: Implications for Therapeutics. Biochem. Pharmacol. 2010, 80, 1050–1062. [Google Scholar] [CrossRef]
- Leterrier, C.; Bonnard, D.; Carrel, D.; Rossier, J.; Lenkei, Z. Constitutive Endocytic Cycle of the CB1 Cannabinoid Receptor. J. Biol. Chem. 2004, 279, 36013–36021. [Google Scholar] [CrossRef]
- Mendizabal-Zubiaga, J.; Melser, S.; Bénard, G.; Ramos, A.; Reguero, L.; Arrabal, S.; Elezgarai, I.; Gerrikagoitia, I.; Suarez, J.; Rodríguez De Fonseca, F.; et al. Cannabinoid CB(1) Receptors Are Localized in Striated Muscle Mitochondria and Regulate Mitochondrial Respiration. Front. Physiol. 2016, 7, 476. [Google Scholar] [CrossRef]
- Bénard, G.; Massa, F.; Puente, N.; Lourenço, J.; Bellocchio, L.; Soria-Gómez, E.; Matias, I.; Delamarre, A.; Metna-Laurent, M.; Cannich, A.; et al. Mitochondrial CB1 Receptors Regulate Neuronal Energy Metabolism. Nat. Neurosci. 2012, 15, 558–564. [Google Scholar] [CrossRef]
- Gutiérrez-Rodríguez, A.; Bonilla-Del Río, I.; Puente, N.; Gómez-Urquijo, S.M.; Fontaine, C.J.; Egaña-Huguet, J.; Elezgarai, I.; Ruehle, S.; Lutz, B.; Robin, L.M.; et al. Localization of the Cannabinoid Type-1 Receptor in Subcellular Astrocyte Compartments of Mutant Mouse Hippocampus. Glia 2018, 66, 1417–1431. [Google Scholar] [CrossRef] [PubMed]
- Kamnate, A.; Sirisin, J.; Watanabe, M.; Kondo, H.; Hipkaeo, W.; Chomphoo, S. Mitochondrial Localization of CB1 in Progesterone-Producing Cells of Ovarian Interstitial Glands of Adult Mice. J. Histochem. Cytochem. Off. J. Histochem. Soc. 2022, 70, 251–257. [Google Scholar] [CrossRef] [PubMed]
- Jimenez-Blasco, D.; Busquets-Garcia, A.; Hebert-Chatelain, E.; Serrat, R.; Vicente-Gutierrez, C.; Ioannidou, C.; Gómez-Sotres, P.; Lopez-Fabuel, I.; Resch-Beusher, M.; Resel, E.; et al. Glucose Metabolism Links Astroglial Mitochondria to Cannabinoid Effects. Nature 2020, 583, 603–608. [Google Scholar] [CrossRef] [PubMed]
- Glass, M.; Faull, R.L.M.; Dragunow, M. Cannabinoid Receptors in the Human Brain: A Detailed Anatomical and Quantitative Autoradiographic Study in the Fetal, Neonatal and Adult Human Brain. Neuroscience 1997, 77, 299–318. [Google Scholar] [CrossRef]
- Herkenham, M.; Lynn, A.B.; Little, M.D.; Johnson, M.R.; Melvin, L.S.; de Costa, B.R.; Rice, K.C. Cannabinoid Receptor Localization in Brain. Proc. Natl. Acad. Sci. USA 1990, 87, 1932–1936. [Google Scholar] [CrossRef]
- Bellocchio, L.; Cervino, C.; Vicennati, V.; Pasquali, R.; Pagotto, U. Cannabinoid Type 1 Receptor: Another Arrow in the Adipocytes’ Bow. J. Neuroendocrinol. 2008, 20 (Suppl. 1), 130–138. [Google Scholar] [CrossRef]
- Di Marzo, V.; Melck, D.; Orlando, P.; Bisogno, T.; Zagoory, O.; Bifulco, M.; Vogel, Z.; Petrocellis, L. De Palmitoylethanolamide Inhibits the Expression of Fatty Acid Amide Hydrolase and Enhances the Anti-Proliferative Effect of Anandamide in Human Breast Cancer Cells. Biochem. J. 2001, 358, 249–255. [Google Scholar] [CrossRef]
- Silvestri, C.; Di Marzo, V. The Endocannabinoid System in Energy Homeostasis and the Etiopathology of Metabolic Disorders. Cell Metab. 2013, 17, 475–490. [Google Scholar] [CrossRef]
- Hirsch, S.; Tam, J. Cannabis: From a Plant That Modulates Feeding Behaviors toward Developing Selective Inhibitors of the Peripheral Endocannabinoid System for the Treatment of Obesity and Metabolic Syndrome. Toxins 2019, 11, 275. [Google Scholar] [CrossRef]
- Foltin, R.W.; Brady, J.V.; Fischman, M.W. Behavioral Analysis of Marijuana Effects on Food Intake in Humans. Pharmacol. Biochem. Behav. 1986, 25, 577–582. [Google Scholar] [CrossRef]
- Rajavashisth, T.B.; Shaheen, M.; Norris, K.C.; Pan, D.; Sinha, S.K.; Ortega, J.; Friedman, T.C. Decreased Prevalence of Diabetes in Marijuana Users: Cross-Sectional Data from the National Health and Nutrition Examination Survey (NHANES) III. BMJ Open 2012, 2, e000494. [Google Scholar] [CrossRef] [PubMed]
- Mahler, S.V.; Smith, K.S.; Berridge, K.C. Endocannabinoid Hedonic Hotspot for Sensory Pleasure: Anandamide in Nucleus Accumbens Shell Enhances “liking” of a Sweet Reward. Neuropsychopharmacol. Off. Publ. Am. Coll. Neuropsychopharmacol. 2007, 32, 2267–2278. [Google Scholar] [CrossRef] [PubMed]
- De Luca, M.A.; Solinas, M.; Bimpisidis, Z.; Goldberg, S.R.; Di Chiara, G. Cannabinoid Facilitation of Behavioral and Biochemical Hedonic Taste Responses. Neuropharmacology 2012, 63, 161–168. [Google Scholar] [CrossRef]
- Gong, J.-P.; Onaivi, E.S.; Ishiguro, H.; Liu, Q.-R.; Tagliaferro, P.A.; Brusco, A.; Uhl, G.R. Cannabinoid CB2 Receptors: Immunohistochemical Localization in Rat Brain. Brain Res. 2006, 1071, 10–23. [Google Scholar] [CrossRef] [PubMed]
- Rossi, F.; Punzo, F.; Umano, G.R.; Argenziano, M.; Miraglia Del Giudice, E. Role of Cannabinoids in Obesity. Int. J. Mol. Sci. 2018, 19, 2690. [Google Scholar] [CrossRef] [PubMed]
- Di Marzo, V.; Goparaju, S.K.; Wang, L.; Liu, J.; Batkai, S.; Jarai, Z.; Fezza, F.; Miura, G.I.; Palmiter, R.D.; Sugiura, T.; et al. Leptin-Regulated Endocannabinoids Are Involved in Maintaining Food Intake. Nature 2001, 410, 822–825. [Google Scholar] [CrossRef] [PubMed]
- Pagotto, U.; Marsicano, G.; Cota, D.; Lutz, B.; Pasquali, R. The Emerging Role of the Endocannabinoid System in Endocrine Regulation and Energy Balance. Endocr. Rev. 2006, 27, 73–100. [Google Scholar] [CrossRef] [PubMed]
- Rinaldi-Carmona, M.; Barth, F.; Heaulme, M.; Alonso, R.; Shire, D.; Congy, C.; Soubrie, P.; Breliere, J.C.; Le Fur, G. Biochemical and Pharmacological Characterisation of SR141716A, the First Potent and Selective Brain Cannabinoid Receptor Antagonist. Life Sci. 1995, 56, 1941–1947. [Google Scholar] [CrossRef]
- Colombo, G.; Agabio, R.; Diaz, G.; Lobina, C.; Reali, R.; Gessa, G.L. Appetite Suppression and Weight Loss after the Cannabinoid Antagonist SR 141716. Life Sci. 1998, 63, PL113–PL117. [Google Scholar] [CrossRef]
- Bensaid, M.; Gary-Bobo, M.; Esclangon, A.; Maffrand, J.P.; Le Fur, G.; Oury-Donat, F.; Soubrie, P. The Cannabinoid CB1 Receptor Antagonist SR141716 Increases Acrp30 MRNA Expression in Adipose Tissue of Obese Fa/Fa Rats and in Cultured Adipocyte Cells. Mol. Pharmacol. 2003, 63, 908–914. [Google Scholar] [CrossRef]
- Carai, M.A.M.; Colombo, G.; Gessa, G.L. Rimonabant: The First Therapeutically Relevant Cannabinoid Antagonist. Life Sci. 2005, 77, 2339–2350. [Google Scholar] [CrossRef] [PubMed]
- Poirier, B.; Bidouard, J.-P.; Cadrouvele, C.; Marniquet, X.; Staels, B.; O’Connor, S.E.; Janiak, P.; Herbert, J.-M. The Anti-Obesity Effect of Rimonabant Is Associated with an Improved Serum Lipid Profile. Diabetes Obes. Metab. 2005, 7, 65–72. [Google Scholar] [CrossRef] [PubMed]
- Ravinet Trillou, C.; Arnone, M.; Delgorge, C.; Gonalons, N.; Keane, P.; Maffrand, J.-P.; Soubrie, P. Anti-Obesity Effect of SR141716, a CB1 Receptor Antagonist, in Diet-Induced Obese Mice. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2003, 284, R345–R353. [Google Scholar] [CrossRef] [PubMed]
- Moreira, F.A.; Crippa, J.A.S. The Psychiatric Side-Effects of Rimonabant. Rev. Bras. Psiquiatr. 2009, 31, 145–153. [Google Scholar] [CrossRef]
- Pagano, C.; Pilon, C.; Calcagno, A.; Urbanet, R.; Rossato, M.; Milan, G.; Bianchi, K.; Rizzuto, R.; Bernante, P.; Federspil, G.; et al. The Endogenous Cannabinoid System Stimulates Glucose Uptake in Human Fat Cells via Phosphatidylinositol 3-Kinase and Calcium-Dependent Mechanisms. J. Clin. Endocrinol. Metab. 2007, 92, 4810–4819. [Google Scholar] [CrossRef]
- Cavuoto, P.; McAinch, A.J.; Hatzinikolas, G.; Janovska, A.; Game, P.; Wittert, G.A. The Expression of Receptors for Endocannabinoids in Human and Rodent Skeletal Muscle. Biochem. Biophys. Res. Commun. 2007, 364, 105–110. [Google Scholar] [CrossRef]
- Bermúdez-Silva, F.J.; Suárez, J.; Baixeras, E.; Cobo, N.; Bautista, D.; Cuesta-Muñoz, A.L.; Fuentes, E.; Juan-Pico, P.; Castro, M.J.; Milman, G.; et al. Presence of Functional Cannabinoid Receptors in Human Endocrine Pancreas. Diabetologia 2008, 51, 476–487. [Google Scholar] [CrossRef]
- Manara, L.; Croci, T.; Guagnini, F.; Rinaldi-Carmona, M.; Maffrand, J.-P.; Le Fur, G.; Mukenge, S.; Ferla, G. Functional Assessment of Neuronal Cannabinoid Receptors in the Muscular Layers of Human Ileum and Colon. Dig. Liver Dis. 2002, 34, 262–269. [Google Scholar] [CrossRef]
- Mukhopadhyay, B.; Liu, J.; Osei-Hyiaman, D.; Godlewski, G.; Mukhopadhyay, P.; Wang, L.; Jeong, W.-I.; Gao, B.; Duester, G.; Mackie, K.; et al. Transcriptional Regulation of Cannabinoid Receptor-1 Expression in the Liver by Retinoic Acid Acting via Retinoic Acid Receptor-Gamma. J. Biol. Chem. 2010, 285, 19002–19011. [Google Scholar] [CrossRef]
- Gasperi, V.; Fezza, F.; Pasquariello, N.; Bari, M.; Oddi, S.; Agro, A.F.; Maccarrone, M. Endocannabinoids in Adipocytes during Differentiation and Their Role in Glucose Uptake. Cell. Mol. Life Sci. 2007, 64, 219–229. [Google Scholar] [CrossRef]
- Roche, R.; Hoareau, L.; Bes-Houtmann, S.; Gonthier, M.-P.; Laborde, C.; Baron, J.-F.; Haffaf, Y.; Cesari, M.; Festy, F. Presence of the Cannabinoid Receptors, CB1 and CB2, in Human Omental and Subcutaneous Adipocytes. Histochem. Cell Biol. 2006, 126, 177–187. [Google Scholar] [CrossRef] [PubMed]
- Juan-Picó, P.; Fuentes, E.; Bermúdez-Silva, F.J.; Díaz-Molina, F.J.; Ripoll, C.; de Fonseca, F.R.; Nadal, A. Cannabinoid Receptors Regulate Ca(2+) Signals and Insulin Secretion in Pancreatic Beta-Cell. Cell Calcium 2006, 39, 155–162. [Google Scholar] [CrossRef] [PubMed]
- Horn, H.; Böhme, B.; Dietrich, L.; Koch, M. Endocannabinoids in Body Weight Control. Pharmaceuticals 2018, 11, 55. [Google Scholar] [CrossRef] [PubMed]
- Izzo, A.A.; Sharkey, K.A. Cannabinoids and the Gut: New Developments and Emerging Concepts. Pharmacol. Ther. 2010, 126, 21–38. [Google Scholar] [CrossRef] [PubMed]
- Tam, J. Role of the Endocannabinoid System in Hepatic Lipogenesis. In Hepatic De Novo Lipogenesis and Regulation of Metabolism; Springer: Berlin/Heidelberg, Germany, 2016; pp. 131–142. ISBN 978-3-319-25063-2. [Google Scholar]
- Osei-Hyiaman, D.; DePetrillo, M.; Pacher, P.; Liu, J.; Radaeva, S.; Bátkai, S.; Harvey-White, J.; Mackie, K.; Offertáler, L.; Wang, L.; et al. Endocannabinoid Activation at Hepatic CB1 Receptors Stimulates Fatty Acid Synthesis and Contributes to Diet-Induced Obesity. J. Clin. Investig. 2005, 115, 1298–1305. [Google Scholar] [CrossRef]
- Adejumo, A.C.; Alliu, S.; Ajayi, T.O.; Adejumo, K.L.; Adegbala, O.M.; Onyeakusi, N.E.; Akinjero, A.M.; Durojaiye, M.; Bukong, T.N. Cannabis Use Is Associated with Reduced Prevalence of Non-Alcoholic Fatty Liver Disease: A Cross-Sectional Study. PLoS ONE 2017, 12, e0176416. [Google Scholar] [CrossRef]
- Mumford, S.L.; Flannagan, K.S.; Radoc, J.G.; Sjaarda, L.A.; Zolton, J.R.; Metz, T.D.; Plowden, T.C.; Perkins, N.J.; DeVilbiss, E.A.; Andriessen, V.C.; et al. Cannabis Use While Trying to Conceive: A Prospective Cohort Study Evaluating Associations with Fecundability, Live Birth and Pregnancy Loss. Hum. Reprod. 2021, 36, 1405–1415. [Google Scholar] [CrossRef]
- Carroll, K.; Pottinger, A.M.; Wynter, S.; DaCosta, V. Marijuana Use and Its Influence on Sperm Morphology and Motility: Identified Risk for Fertility among Jamaican Men. Andrology 2020, 8, 136–142. [Google Scholar] [CrossRef]
- El-Talatini, M.R.; Taylor, A.H.; Konje, J.C. The Relationship between Plasma Levels of the Endocannabinoid, Anandamide, Sex Steroids, and Gonadotrophins during the Menstrual Cycle. Fertil. Steril. 2010, 93, 1989–1996. [Google Scholar] [CrossRef]
- MacCarrone, M.; De Felici, M.; Bari, M.; Klinger, F.; Siracusa, G.; Finazzi-Agrò, A. Down-Regulation of Anandamide Hydrolase in Mouse Uterus by Sex Hormones. Eur. J. Biochem. 2000, 267, 2991–2997. [Google Scholar] [CrossRef]
- Schuel, H.; Burkman, L.J.; Lippes, J.; Crickard, K.; Forester, E.; Piomelli, D.; Giuffrida, A. N-Acylethanolamines in Human Reproductive Fluids. Chem. Phys. Lipids 2002, 121, 211–227. [Google Scholar] [CrossRef]
- Schmid, P.C.; Paria, B.C.; Krebsbach, R.J.; Schmid, H.H.; Dey, S.K. Changes in Anandamide Levels in Mouse Uterus Are Associated with Uterine Receptivity for Embryo Implantation. Proc. Natl. Acad. Sci. USA 1997, 94, 4188–4192. [Google Scholar] [CrossRef] [PubMed]
- Paria, B.C.; Das, S.K.; Dey, S.K. The Preimplantation Mouse Embryo Is a Target for Cannabinoid Ligand-Receptor Signaling. Proc. Natl. Acad. Sci. USA 1995, 92, 9460–9464. [Google Scholar] [CrossRef]
- Paria, B.C.; Song, H.; Wang, X.; Schmid, P.C.; Krebsbach, R.J.; Schmid, H.H.; Bonner, T.I.; Zimmer, A.; Dey, S.K. Dysregulated Cannabinoid Signaling Disrupts Uterine Receptivity for Embryo Implantation. J. Biol. Chem. 2001, 276, 20523–20528. [Google Scholar] [CrossRef] [PubMed]
- Paria, B.C.; Ma, W.; Andrenyak, D.M.; Schmid, P.C.; Schmid, H.H.O.; Moody, D.E.; Deng, H.; Makriyannis, A.; Dey, S.K. Effects of Cannabinoids on Preimplantation Mouse Embryo Development and Implantation Are Mediated by Brain-Type Cannabinoid Receptors1. Biol. Reprod. 1998, 58, 1490–1495. [Google Scholar] [CrossRef]
- Wang, H.; Guo, Y.; Wang, D.; Kingsley, P.J.; Marnett, L.J.; Das, S.K.; DuBois, R.N.; Dey, S.K. Aberrant Cannabinoid Signaling Impairs Oviductal Transport of Embryos. Nat. Med. 2004, 10, 1074–1080. [Google Scholar] [CrossRef]
- Habayeb, O.M.H.; Taylor, A.H.; Evans, M.D.; Cooke, M.S.; Taylor, D.J.; Bell, S.C.; Konje, J.C. Plasma Levels of the Endocannabinoid Anandamide in Women—A Potential Role in Pregnancy Maintenance and Labor? J. Clin. Endocrinol. Metab. 2004, 89, 5482–5487. [Google Scholar] [CrossRef]
- Wang, H.; Xie, H.; Sun, X.; Kingsley, P.J.; Marnett, L.J.; Cravatt, B.F.; Dey, S.K. Differential Regulation of Endocannabinoid Synthesis and Degradation in the Uterus during Embryo Implantation. Prostaglandins Other Lipid Mediat. 2007, 83, 62–74. [Google Scholar] [CrossRef]
- Cui, N.; Yang, Y.; Xu, Y.; Zhang, J.; Jiang, L.; Hao, G. Decreased Expression of Fatty Acid Amide Hydrolase in Women with Polycystic Ovary Syndrome. Gynecol. Endocrinol. Off. J. Int. Soc. Gynecol. Endocrinol. 2017, 33, 368–372. [Google Scholar] [CrossRef]
- Juan, C.-C.; Chen, K.-H.; Wang, P.-H.; Hwang, J.-L.; Seow, K.-M. Endocannabinoid System Activation May Be Associated with Insulin Resistance in Women with Polycystic Ovary Syndrome. Fertil. Steril. 2015, 104, 200–206. [Google Scholar] [CrossRef]
- Cui, N.; Feng, X.; Zhao, Z.; Zhang, J.; Xu, Y.; Wang, L.; Hao, G. Restored Plasma Anandamide and Endometrial Expression of Fatty Acid Amide Hydrolase in Women With Polycystic Ovary Syndrome by the Combination Use of Diane-35 and Metformin. Clin. Ther. 2017, 39, 751–758. [Google Scholar] [CrossRef] [PubMed]
- Kabakchieva, P.; Gateva, A.; Hristova, J.; Georgiev, T.; Kamenov, Z. Analysis of 2-Arachidonoylglycerol Levels in Polycystic Ovary Syndrome in the Context of Hormonal and Metabolic Alterations and Across the Classical Phenotypes. Cannabis Cannabinoid Res. 2022. [Google Scholar] [CrossRef] [PubMed]
- Kabakchieva, P.; Gateva, A.; Velikova, T.; Georgiev, T.; Kamenov, Z. Anandamide Is Associated with Waist-to-Hip Ratio but Not with Body Mass Index in Women with Polycystic Ovary Syndrome. Minerva Endocrinol. 2022; ahead of print. [Google Scholar] [CrossRef]
- Pagotto, U.; Marsicano, G.; Fezza, F.; Theodoropoulou, M.; Grübler, Y.; Stalla, J.; Arzberger, T.; Milone, A.; Losa, M.; Di Marzo, V.; et al. Normal Human Pituitary Gland and Pituitary Adenomas Express Cannabinoid Receptor Type 1 and Synthesize Endogenous Cannabinoids: First Evidence for a Direct Role of Cannabinoids on Hormone Modulation at the Human Pituitary Level. J. Clin. Endocrinol. Metab. 2001, 86, 2687–2696. [Google Scholar] [CrossRef] [PubMed]
- Ziegler, C.G.; Mohn, C.; Lamounier-Zepter, V.; Rettori, V.; Bornstein, S.R.; Krug, A.W.; Ehrhart-Bornstein, M. Expression and Function of Endocannabinoid Receptors in the Human Adrenal Cortex. Horm. Metab. 2010, 42, 88–92. [Google Scholar] [CrossRef]
- Wang, J.; Wu, X. The Effects of Mitochondrial Dysfunction on Energy Metabolism Switch by HIF-1α Signalling in Granulosa Cells of Polycystic Ovary Syndrome. Endokrynol. Pol. 2020, 71, 134–145. [Google Scholar] [CrossRef]
- Montgomery, M.K.; Turner, N. Mitochondrial Dysfunction and Insulin Resistance: An Update. Endocr. Connect. 2015, 4, R1–R15. [Google Scholar] [CrossRef]
- Han, L.; Shen, W.-J.; Bittner, S.; Kraemer, F.B.; Azhar, S. PPARs: Regulators of Metabolism and as Therapeutic Targets in Cardiovascular Disease. Part II: PPAR-β/δ and PPAR-γ. Future Cardiol. 2017, 13, 279–296. [Google Scholar] [CrossRef]
- Chiazza, F.; Collino, M. Chapter 9—Peroxisome Proliferator-Activated Receptors (PPARs) in Glucose Control. In Molecular Nutrition and Diabetes; Mauricio, D., Ed.; Academic Press: San Diego, CA, USA, 2016; pp. 105–114. ISBN 978-0-12-801585-8. [Google Scholar]
- Berger, J.; Moller, D.E. The Mechanisms of Action of PPARs. Annu. Rev. Med. 2002, 53, 409–435. [Google Scholar] [CrossRef]
- Luquet, S.; Lopez-Soriano, J.; Holst, D.; Gaudel, C.; Jehl-Pietri, C.; Fredenrich, A.; Grimaldi, P.A. Roles of Peroxisome Proliferator-Activated Receptor Delta (PPARdelta) in the Control of Fatty Acid Catabolism. A New Target for the Treatment of Metabolic Syndrome. Biochimie 2004, 86, 833–837. [Google Scholar] [CrossRef]
- PPARA Peroxisome Proliferator Activated Receptor Alpha [Homo Sapiens (Human)]—Gene—NCBI. Available online: https://www.ncbi.nlm.nih.gov/gene?Db=gene&Cmd=DetailsSearch&Term=5465 (accessed on 23 July 2022).
- PPARD Peroxisome Proliferator Activated Receptor Delta [Homo Sapiens (Human)]—Gene—NCBI. Available online: https://www.ncbi.nlm.nih.gov/gene/5467 (accessed on 23 July 2022).
- PPARG Peroxisome Proliferator Activated Receptor Gamma [Homo Sapiens (Human)]—Gene—NCBI. Available online: https://www.ncbi.nlm.nih.gov/gene?Db=gene&Cmd=DetailsSearch&Term=5468 (accessed on 23 July 2022).
- Brunmeir, R.; Xu, F. Functional Regulation of PPARs through Post-Translational Modifications. Int. J. Mol. Sci. 2018, 19, 1738. [Google Scholar] [CrossRef]
- Wang, L.; Waltenberger, B.; Pferschy-Wenzig, E.-M.; Blunder, M.; Liu, X.; Malainer, C.; Blazevic, T.; Schwaiger, S.; Rollinger, J.M.; Heiss, E.H.; et al. Natural Product Agonists of Peroxisome Proliferator-Activated Receptor Gamma (PPARγ): A Review. Biochem. Pharmacol. 2014, 92, 73–89. [Google Scholar] [CrossRef] [PubMed]
- Sinha, R.A.; Rajak, S.; Singh, B.K.; Yen, P.M. Hepatic Lipid Catabolism via PPARα-Lysosomal Crosstalk. Int. J. Mol. Sci. 2020, 21, 2391. [Google Scholar] [CrossRef] [PubMed]
- Sher, T.; Yi, H.F.; McBride, O.W.; Gonzalez, F.J. CDNA Cloning, Chromosomal Mapping, and Functional Characterization of the Human Peroxisome Proliferator Activated Receptor. Biochemistry 1993, 32, 5598–5604. [Google Scholar] [CrossRef] [PubMed]
- Desvergne, B.; Wahli, W. Peroxisome Proliferator-Activated Receptors: Nuclear Control of Metabolism. Endocr. Rev. 1999, 20, 649–688. [Google Scholar] [CrossRef]
- Kliewer, S.A.; Sundseth, S.S.; Jones, S.A.; Brown, P.J.; Wisely, G.B.; Koble, C.S.; Devchand, P.; Wahli, W.; Willson, T.M.; Lenhard, J.M.; et al. Fatty Acids and Eicosanoids Regulate Gene Expression through Direct Interactions with Peroxisome Proliferator-Activated Receptors Alpha and Gamma. Proc. Natl. Acad. Sci. USA 1997, 94, 4318–4323. [Google Scholar] [CrossRef]
- Gross, B.; Pawlak, M.; Lefebvre, P.; Staels, B. PPARs in Obesity-Induced T2DM, Dyslipidaemia and NAFLD. Nat. Rev. Endocrinol. 2017, 13, 36–49. [Google Scholar] [CrossRef]
- Singh, G.; Correa, R. Fibrate Medications; StatPearls: Treasure Island, FL, USA, 2022. [Google Scholar]
- Skogsberg, J.; Kannisto, K.; Roshani, L.; Gagne, E.; Hamsten, A.; Larsson, C.; Ehrenborg, E. Characterization of the Human Peroxisome Proliferator Activated Receptor Delta Gene and Its Expression. Int. J. Mol. Med. 2000, 6, 73–81. [Google Scholar] [CrossRef]
- Fan, W.; Waizenegger, W.; Lin, C.S.; Sorrentino, V.; He, M.-X.; Wall, C.E.; Li, H.; Liddle, C.; Yu, R.T.; Atkins, A.R.; et al. PPARδ Promotes Running Endurance by Preserving Glucose. Cell Metab. 2017, 25, 1186–1193.e4. [Google Scholar] [CrossRef]
- Liu, Y.; Colby, J.K.; Zuo, X.; Jaoude, J.; Wei, D.; Shureiqi, I. The Role of PPAR-δ in Metabolism, Inflammation, and Cancer: Many Characters of a Critical Transcription Factor. Int. J. Mol. Sci. 2018, 19, 3339. [Google Scholar] [CrossRef]
- Fajas, L.; Auboeuf, D.; Raspé, E.; Schoonjans, K.; Lefebvre, A.-M.; Saladin, R.; Najib, J.; Laville, M.; Fruchart, J.-C.; Deeb, S.; et al. The Organization, Promoter Analysis, and Expression of the Human PPARγ Gene*. J. Biol. Chem. 1997, 272, 18779–18789. [Google Scholar] [CrossRef]
- McIntyre, T.M.; Pontsler, A.V.; Silva, A.R.; St Hilaire, A.; Xu, Y.; Hinshaw, J.C.; Zimmerman, G.A.; Hama, K.; Aoki, J.; Arai, H.; et al. Identification of an Intracellular Receptor for Lysophosphatidic Acid (LPA): LPA Is a Transcellular PPARgamma Agonist. Proc. Natl. Acad. Sci. USA 2003, 100, 131–136. [Google Scholar] [CrossRef] [PubMed]
- Nanjan, M.J.; Mohammed, M.; Kumar, B.R.P.; Chandrasekar, M.J.N. Thiazolidinediones as Antidiabetic Agents: A Critical Review. Bioorg. Chem. 2018, 77, 548–567. [Google Scholar] [CrossRef] [PubMed]
- Tenenbaum, A.; Fisman, E.Z. Fibrates Are an Essential Part of Modern Anti-Dyslipidemic Arsenal: Spotlight on Atherogenic Dyslipidemia and Residual Risk Reduction. Cardiovasc. Diabetol. 2012, 11, 125. [Google Scholar] [CrossRef] [PubMed]
- Tenenbaum, A.; Motro, M.; Fisman, E.Z. Dual and Pan-Peroxisome Proliferator-Activated Receptors (PPAR) Co-Agonism: The Bezafibrate Lessons. Cardiovasc. Diabetol. 2005, 4, 14. [Google Scholar] [CrossRef]
- Teramoto, T.; Shirai, K.; Daida, H.; Yamada, N. Effects of Bezafibrate on Lipid and Glucose Metabolism in Dyslipidemic Patients with Diabetes: The J-BENEFIT Study. Cardiovasc. Diabetol. 2012, 11, 29. [Google Scholar] [CrossRef]
- Tenenbaum, A.; Fisman, E.Z.; Boyko, V.; Benderly, M.; Tanne, D.; Haim, M.; Matas, Z.; Motro, M.; Behar, S. Attenuation of Progression of Insulin Resistance in Patients with Coronary Artery Disease by Bezafibrate. Arch. Intern. Med. 2006, 166, 737–741. [Google Scholar] [CrossRef]
- Ogawa, S.; Takeuchi, K.; Sugimura, K.; Fukuda, M.; Lee, R.; Ito, S.; Sato, T. Bezafibrate Reduces Blood Glucose in Type 2 Diabetes Mellitus. Metabolism 2000, 49, 331–334. [Google Scholar] [CrossRef]
- Tenenbaum, H.; Behar, S.; Boyko, V.; Adler, Y.; Fisman, E.Z.; Tanne, D.; Lapidot, M.; Schwammenthal, E.; Feinberg, M.; Matas, Z.; et al. Long-Term Effect of Bezafibrate on Pancreatic Beta-Cell Function and Insulin Resistance in Patients with Diabetes. Atherosclerosis 2007, 194, 265–271. [Google Scholar] [CrossRef]
- Hiuge, A.; Tenenbaum, A.; Maeda, N.; Benderly, M.; Kumada, M.; Fisman, E.Z.; Tanne, D.; Matas, Z.; Hibuse, T.; Fujita, K.; et al. Effects of Peroxisome Proliferator-Activated Receptor Ligands, Bezafibrate and Fenofibrate, on Adiponectin Level. Arterioscler. Thromb. Vasc. Biol. 2007, 27, 635–641. [Google Scholar] [CrossRef]
- Sun, N.; Shen, C.; Zhang, L.; Wu, X.; Yu, Y.; Yang, X.; Yang, C.; Zhong, C.; Gao, Z.; Miao, W.; et al. Hepatic Krüppel-like Factor 16 (KLF16) Targets PPARα to Improve Steatohepatitis and Insulin Resistance. Gut 2021, 70, 2183–2195. [Google Scholar] [CrossRef]
- Wang, Y.-X.; Lee, C.-H.; Tiep, S.; Yu, R.T.; Ham, J.; Kang, H.; Evans, R.M. Peroxisome-Proliferator-Activated Receptor Delta Activates Fat Metabolism to Prevent Obesity. Cell 2003, 113, 159–170. [Google Scholar] [CrossRef]
- Ravnskjaer, K.; Frigerio, F.; Boergesen, M.; Nielsen, T.; Maechler, P.; Mandrup, S. PPARδ Is a Fatty Acid Sensor That Enhances Mitochondrial Oxidation in Insulin-Secreting Cells and Protects against Fatty Acid-Induced Dysfunction. J. Lipid Res. 2010, 51, 1370–1379. [Google Scholar] [CrossRef] [PubMed]
- Doktorova, M.; Zwarts, I.; van Zutphen, T.; van Dijk, T.H.; Bloks, V.W.; Harkema, L.; de Bruin, A.; Downes, M.; Evans, R.M.; Verkade, H.J.; et al. Intestinal PPARδ Protects against Diet-Induced Obesity, Insulin Resistance and Dyslipidemia. Sci. Rep. 2017, 7, 846. [Google Scholar] [CrossRef] [PubMed]
- Kintz, P.; Ameline, A.; Gheddar, L.; Raul, J.-S. Testing for GW501516 (Cardarine) in Human Hair Using LC/MS-MS and Confirmation by LC/HRMS. Drug Test. Anal. 2020, 12, 980–986. [Google Scholar] [CrossRef] [PubMed]
- Sahebkar, A.; Chew, G.T.; Watts, G.F. New Peroxisome Proliferator-Activated Receptor Agonists: Potential Treatments for Atherogenic Dyslipidemia and Non-Alcoholic Fatty Liver Disease. Expert Opin. Pharmacother. 2014, 15, 493–503. [Google Scholar] [CrossRef] [PubMed]
- Adams, M.; Montague, C.T.; Prins, J.B.; Holder, J.C.; Smith, S.A.; Sanders, L.; Digby, J.E.; Sewter, C.P.; Lazar, M.A.; Chatterjee, V.K.; et al. Activators of Peroxisome Proliferator-Activated Receptor Gamma Have Depot-Specific Effects on Human Preadipocyte Differentiation. J. Clin. Investig. 1997, 100, 3149–3153. [Google Scholar] [CrossRef]
- Okuno, A.; Tamemoto, H.; Tobe, K.; Ueki, K.; Mori, Y.; Iwamoto, K.; Umesono, K.; Akanuma, Y.; Fujiwara, T.; Horikoshi, H.; et al. Troglitazone Increases the Number of Small Adipocytes without the Change of White Adipose Tissue Mass in Obese Zucker Rats. J. Clin. Investig. 1998, 101, 1354–1361. [Google Scholar] [CrossRef]
- Tontonoz, P.; Hu, E.; Spiegelman, B.M. Stimulation of Adipogenesis in Fibroblasts by PPARγ2, a Lipid-Activated Transcription Factor. Cell 1994, 79, 1147–1156. [Google Scholar] [CrossRef]
- Yamanouchi, T. Concomitant Therapy with Pioglitazone and Insulin for the Treatment of Type 2 Diabetes. Vasc. Health Risk Manag. 2010, 6, 189–197. [Google Scholar] [CrossRef]
- National Institutes of Health. Antidiabetic Agents. In LiverTox: Clinical and Research Information on Drug-Induced Liver Injury; National Institutes of Health: Bethesda, MD, USA, 2017. [Google Scholar]
- Gor, D.; Gerber, B.S.; Walton, S.M.; Lee, T.A.; Nutescu, E.A.; Touchette, D.R. Antidiabetic Drug Use Trends in Patients with Type 2 Diabetes Mellitus and Chronic Kidney Disease: A Cross-Sectional Analysis of the National Health and Nutrition Examination Survey. J. Diabetes 2020, 12, 385–395. [Google Scholar] [CrossRef]
- Vieira, R.; Souto, S.B.; Sánchez-López, E.; Machado, A.L.; Severino, P.; Jose, S.; Santini, A.; Fortuna, A.; García, M.L.; Silva, A.M.; et al. Sugar-Lowering Drugs for Type 2 Diabetes Mellitus and Metabolic Syndrome-Review of Classical and New Compounds: Part-I. Pharmaceuticals 2019, 12, 152. [Google Scholar] [CrossRef] [PubMed]
- Yau, H.; Rivera, K.; Lomonaco, R.; Cusi, K. The Future of Thiazolidinedione Therapy in the Management of Type 2 Diabetes Mellitus. Curr. Diabetes Rep. 2013, 13, 329–341. [Google Scholar] [CrossRef] [PubMed]
- Choi, S.-S.; Park, J.; Choi, J.H. Revisiting PPARγ as a Target for the Treatment of Metabolic Disorders. BMB Rep. 2014, 47, 599–608. [Google Scholar] [CrossRef] [PubMed]
- Tyagi, S.; Gupta, P.; Saini, A.S.; Kaushal, C.; Sharma, S. The Peroxisome Proliferator-Activated Receptor: A Family of Nuclear Receptors Role in Various Diseases. J. Adv. Pharm. Technol. Res. 2011, 2, 236–240. [Google Scholar] [CrossRef] [PubMed]
- Lebovitz, H.E. Thiazolidinediones: The Forgotten Diabetes Medications. Curr. Diabetes Rep. 2019, 19, 151. [Google Scholar] [CrossRef] [PubMed]
- Schernthaner, G.; Matthews, D.R.; Charbonnel, B.; Hanefeld, M.; Brunetti, P. Efficacy and Safety of Pioglitazone versus Metformin in Patients with Type 2 Diabetes Mellitus: A Double-Blind, Randomized Trial. J. Clin. Endocrinol. Metab. 2004, 89, 6068–6076. [Google Scholar] [CrossRef]
- Komar, C.M. Peroxisome Proliferator-Activated Receptors (PPARs) and Ovarian Function—Implications for Regulating Steroidogenesis, Differentiation, and Tissue Remodeling. Reprod. Biol. Endocrinol. 2005, 3, 41. [Google Scholar] [CrossRef]
- Komar, C.M.; Curry, T.E.J. Localization and Expression of Messenger RNAs for the Peroxisome Proliferator-Activated Receptors in Ovarian Tissue from Naturally Cycling and Pseudopregnant Rats. Biol. Reprod. 2002, 66, 1531–1539. [Google Scholar] [CrossRef]
- Komar, C. Initiation of Peroxisome Proliferator-Activated Receptor Gamma (PPAR Gamma) Expression in the Neonatal Rat Ovary. In Proceedings of the Society for the Study of Reproduction 38th Annual Meeting, Quebec City, QC, Canada, 24–27 July 2005. [Google Scholar]
- Froment, P.; Fabre, S.; Dupont, J.; Pisselet, C.; Chesneau, D.; Staels, B.; Monget, P. Expression and Functional Role of Peroxisome Proliferator-Activated Receptor-Gamma in Ovarian Folliculogenesis in the Sheep. Biol. Reprod. 2003, 69, 1665–1674. [Google Scholar] [CrossRef]
- Komar, C.M.; Braissant, O.; Wahli, W.; Curry, T.E.J. Expression and Localization of PPARs in the Rat Ovary during Follicular Development and the Periovulatory Period. Endocrinology 2001, 142, 4831–4838. [Google Scholar] [CrossRef]
- Cammas, L.; Reinaud, P.; Bordas, N.; Dubois, O.; Germain, G.; Charpigny, G. Developmental Regulation of Prostacyclin Synthase and Prostacyclin Receptors in the Ovine Uterus and Conceptus during the Peri-Implantation Period. Reproduction 2006, 131, 917–927. [Google Scholar] [CrossRef] [PubMed]
- Peng, L.; Yang, H.; Ye, Y.; Ma, Z.; Kuhn, C.; Rahmeh, M.; Mahner, S.; Makrigiannakis, A.; Jeschke, U.; von Schönfeldt, V. Role of Peroxisome Proliferator-Activated Receptors (PPARs) in Trophoblast Functions. Int. J. Mol. Sci. 2021, 22, 433. [Google Scholar] [CrossRef] [PubMed]
- Wang, Q.; Fujii, H.; Knipp, G.T. Expression of PPAR and RXR Isoforms in the Developing Rat and Human Term Placentas. Placenta 2002, 23, 661–671. [Google Scholar] [CrossRef] [PubMed]
- Barak, Y.; Nelson, M.C.; Ong, E.S.; Jones, Y.Z.; Ruiz-Lozano, P.; Chien, K.R.; Koder, A.; Evans, R.M. PPAR Gamma Is Required for Placental, Cardiac, and Adipose Tissue Development. Mol. Cell 1999, 4, 585–595. [Google Scholar] [CrossRef]
- Tarrade, A.; Schoonjans, K.; Guibourdenche, J.; Bidart, J.M.; Vidaud, M.; Auwerx, J.; Rochette-Egly, C.; Evain-Brion, D. PPARγ/RXRα Heterodimers Are Involved in Human CGβ Synthesis and Human Trophoblast Differentiation. Endocrinology 2001, 142, 4504–4514. [Google Scholar] [CrossRef] [PubMed]
- Mayama, R.; Izawa, T.; Sakai, K.; Suciu, N.; Iwashita, M. Improvement of Insulin Sensitivity Promotes Extravillous Trophoblast Cell Migration Stimulated by Insulin-like Growth Factor-I. Endocr. J. 2013, 60, 359–368. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Huo, Y.; He, W.; Liu, S.; Li, H.; Li, L. Visfatin Is Regulated by Interleukin-6 and Affected by the PPAR-γ Pathway in BeWo Cells. Mol. Med. Rep. 2019, 19, 400–406. [Google Scholar] [CrossRef]
- Aye, I.L.M.H.; Gao, X.; Weintraub, S.T.; Jansson, T.; Powell, T.L. Adiponectin Inhibits Insulin Function in Primary Trophoblasts by PPARα-Mediated Ceramide Synthesis. Mol. Endocrinol. 2014, 28, 512–524. [Google Scholar] [CrossRef]
- Ding, N.-Z.; Ma, X.-H.; Diao, H.-L.; Xu, L.-B.; Yang, Z.-M. Differential Expression of Peroxisome Proliferator-Activated Receptor Delta at Implantation Sites and in Decidual Cells of Rat Uterus. Reproduction 2003, 125, 817–825. [Google Scholar] [CrossRef]
- Toda, K.; Okada, T.; Miyaura, C.; Saibara, T. Fenofibrate, a Ligand for PPARα, Inhibits Aromatase Cytochrome P450 Expression in the Ovary of Mouse. J. Lipid Res. 2003, 44, 265–270. [Google Scholar] [CrossRef]
- Kang, H.J.; Hwang, S.J.; Yoon, J.A.; Jun, J.H.; Lim, H.J.; Yoon, T.K.; Song, H. Activation of Peroxisome Proliferators-Activated Receptor δ (PPARδ) Promotes Blastocyst Hatching in Mice. Mol. Hum. Reprod. 2011, 17, 653–660. [Google Scholar] [CrossRef] [PubMed]
- Rak-Mardyła, A.; Karpeta, A. Rosiglitazone Stimulates Peroxisome Proliferator-Activated Receptor Gamma Expression and Directly Affects in Vitro Steroidogenesis in Porcine Ovarian Follicles. Theriogenology 2014, 82, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Veldhuis, J.D.; Zhang, G.; Garmey, J.C. Troglitazone, an Insulin-Sensitizing Thiazolidinedione, Represses Combined Stimulation by LH and Insulin of de Novo Androgen Biosynthesis by Thecal Cells in Vitro. J. Clin. Endocrinol. Metab. 2002, 87, 1129–1133. [Google Scholar] [CrossRef] [PubMed]
- Du, Q.; Yang, S.; Wang, Y.-J.; Wu, B.; Zhao, Y.-Y.; Fan, B. Effects of Thiazolidinediones on Polycystic Ovary Syndrome: A Meta-Analysis of Randomized Placebo-Controlled Trials. Adv. Ther. 2012, 29, 763–774. [Google Scholar] [CrossRef]
- Zhao, H.; Xing, C.; Zhang, J.; He, B. Comparative Efficacy of Oral Insulin Sensitizers Metformin, Thiazolidinediones, Inositol, and Berberine in Improving Endocrine and Metabolic Profiles in Women with PCOS: A Network Meta-Analysis. Reprod. Health 2021, 18, 171. [Google Scholar] [CrossRef]
- Cataldo, N.A.; Abbasi, F.; McLaughlin, T.L.; Basina, M.; Fechner, P.Y.; Giudice, L.C.; Reaven, G.M. Metabolic and Ovarian Effects of Rosiglitazone Treatment for 12 Weeks in Insulin-Resistant Women with Polycystic Ovary Syndrome. Hum. Reprod. 2006, 21, 109–120. [Google Scholar] [CrossRef]
- Rashid, R.; Mir, S.A.; Kareem, O.; Ali, T.; Ara, R.; Malik, A.; Amin, F.; Bader, G.N. Polycystic Ovarian Syndrome-Current Pharmacotherapy and Clinical Implications. Taiwan J. Obstet. Gynecol. 2022, 61, 40–50. [Google Scholar] [CrossRef]
- Eggleton, J.S.; Jialal, I. Thiazolidinediones; StatPearls: Treasure Island, FL, USA, 2022. [Google Scholar]
- Uruno, A.; Matsuda, K.; Noguchi, N.; Yoshikawa, T.; Kudo, M.; Satoh, F.; Rainey, W.E.; Hui, X.-G.; Akahira, J.; Nakamura, Y.; et al. Peroxisome Proliferator-Activated Receptor-{gamma} Suppresses CYP11B2 Expression and Aldosterone Production. J. Mol. Endocrinol. 2011, 46, 37–49. [Google Scholar] [CrossRef]
- Ventura, R.D.; Chaves, A.S.; Magalhães, N.S.; Gonzalez, F.B.; Pacini, M.F.; Pérez, A.R.; Silva, P.M.R.; Martins, M.A.; Carvalho, V.F. Activation of PPARγ Reduces N-Acetyl-Cysteine -Induced Hypercorticoidism by down-Regulating MC2R Expression into Adrenal Glands. Free Radic. Biol. Med. 2020, 156, 137–143. [Google Scholar] [CrossRef]
- O’Sullivan, S.E.; Kendall, D.A. Cannabinoid Activation of Peroxisome Proliferator-Activated Receptors: Potential for Modulation of Inflammatory Disease. Immunobiology 2010, 215, 611–616. [Google Scholar] [CrossRef]
- O’Sullivan, S.E. Cannabinoids Go Nuclear: Evidence for Activation of Peroxisome Proliferator-Activated Receptors. Br. J. Pharmacol. 2007, 152, 576–582. [Google Scholar] [CrossRef] [PubMed]
- Pistis, M.; O’Sullivan, S.E. The Role of Nuclear Hormone Receptors in Cannabinoid Function. Adv. Pharmacol. 2017, 80, 291–328. [Google Scholar] [CrossRef] [PubMed]
- Sun, Y.; Alexander, S.P.H.; Garle, M.J.; Gibson, C.L.; Hewitt, K.; Murphy, S.P.; Kendall, D.A.; Bennett, A.J. Cannabinoid Activation of PPAR Alpha; a Novel Neuroprotective Mechanism. Br. J. Pharmacol. 2007, 152, 734–743. [Google Scholar] [CrossRef] [PubMed]
- Rockwell, C.E.; Snider, N.T.; Thompson, J.T.; Vanden Heuvel, J.P.; Kaminski, N.E. Interleukin-2 Suppression by 2-Arachidonyl Glycerol Is Mediated through Peroxisome Proliferator-Activated Receptor Gamma Independently of Cannabinoid Receptors 1 and 2. Mol. Pharmacol. 2006, 70, 101–111. [Google Scholar] [CrossRef]
- Kozak, K.R.; Gupta, R.A.; Moody, J.S.; Ji, C.; Boeglin, W.E.; DuBois, R.N.; Brash, A.R.; Marnett, L.J. 15-Lipoxygenase Metabolism of 2-Arachidonylglycerol. Generation of a Peroxisome Proliferator-Activated Receptor Alpha Agonist. J. Biol. Chem. 2002, 277, 23278–23286. [Google Scholar] [CrossRef]
- Bouaboula, M.; Hilairet, S.; Marchand, J.; Fajas, L.; Le Fur, G.; Casellas, P. Anandamide Induced PPARγ Transcriptional Activation and 3T3-L1 Preadipocyte Differentiation. Eur. J. Pharmacol. 2005, 517, 174–181. [Google Scholar] [CrossRef]
- LoVerme, J.; La Rana, G.; Russo, R.; Calignano, A.; Piomelli, D. The Search for the Palmitoylethanolamide Receptor. Life Sci. 2005, 77, 1685–1698. [Google Scholar] [CrossRef]
- Fu, J.; Gaetani, S.; Oveisi, F.; Lo Verme, J.; Serrano, A.; Rodriguez De Fonseca, F.; Rosengarth, A.; Luecke, H.; Di Giacomo, B.; Tarzia, G.; et al. Oleylethanolamide Regulates Feeding and Body Weight through Activation of the Nuclear Receptor PPAR-Alpha. Nature 2003, 425, 90–93. [Google Scholar] [CrossRef]
- Dionisi, M.; Alexander, S.P.H.; Bennett, A.J. Oleamide Activates Peroxisome Proliferator-Activated Receptor Gamma (PPARγ) in Vitro. Lipids Health Dis. 2012, 11, 51. [Google Scholar] [CrossRef]
- Raman, P.; Kaplan, B.L.F.; Thompson, J.T.; Heuvel, J.P.V.; Kaminski, N.E. 15-Deoxy-Δ12,14-Prostaglandin J2-Glycerol Ester, a Putative Metabolite of 2-Arachidonyl Glycerol, Activates Peroxisome Proliferator Activated Receptor γ. Mol. Pharmacol. 2011, 80, 201–209. [Google Scholar] [CrossRef]
- Hegde, V.L.; Singh, U.P.; Nagarkatti, P.S.; Nagarkatti, M. Critical Role of Mast Cells and Peroxisome Proliferator-Activated Receptor Gamma in the Induction of Myeloid-Derived Suppressor Cells by Marijuana Cannabidiol In Vivo. J. Immunol. 2015, 194, 5211–5222. [Google Scholar] [CrossRef] [PubMed]
- O’Sullivan, S.E.; Tarling, E.J.; Bennett, A.J.; Kendall, D.A.; Randall, M.D. Novel Time-Dependent Vascular Actions of Δ9-Tetrahydrocannabinol Mediated by Peroxisome Proliferator-Activated Receptor Gamma. Biochem. Biophys. Res. Commun. 2005, 337, 824–831. [Google Scholar] [CrossRef]
- Granja, A.G.; Carrillo-Salinas, F.; Pagani, A.; Gomez-Canas, M.; Negri, R.; Navarrete, C.; Mecha, M.; Mestre, L.; Fiebich, B.L.; Cantarero, I.; et al. A Cannabigerol Quinone Alleviates Neuroinflammation in a Chronic Model of Multiple Sclerosis. J. Neuroimmune Pharmacol. 2012, 7, 1002–1016. [Google Scholar] [CrossRef] [PubMed]
- Nadal, X.; del Río, C.; Casano, S.; Palomares, B.; Ferreiro-Vera, C.; Navarrete, C.; Sánchez-Carnerero, C.; Cantarero, I.; Bellido, M.L.; Meyer, S.; et al. Tetrahydrocannabinolic Acid Is a Potent PPARγ Agonist with Neuroprotective Activity. Br. J. Pharmacol. 2017, 174, 4263–4276. [Google Scholar] [CrossRef] [PubMed]
- Fakhfouri, G.; Ahmadiani, A.; Rahimian, R.; Grolla, A.A.; Moradi, F.; Haeri, A. WIN55212-2 Attenuates Amyloid-Beta-Induced Neuroinflammation in Rats through Activation of Cannabinoid Receptors and PPAR-γ Pathway. Neuropharmacology 2012, 63, 653–666. [Google Scholar] [CrossRef] [PubMed]
- del Rio, C.; Cantarero, I.; Palomares, B.; Gómez-Cañas, M.; Fernández-Ruiz, J.; Pavicic, C.; García-Martín, A.; Luz Bellido, M.; Ortega-Castro, R.; Pérez-Sánchez, C.; et al. VCE-004.3, a Cannabidiol Aminoquinone Derivative, Prevents Bleomycin-Induced Skin Fibrosis and Inflammation through PPARγ- and CB2 Receptor-Dependent Pathways. Br. J. Pharmacol. 2018, 175, 3813–3831. [Google Scholar] [CrossRef] [PubMed]
- García-Martín, A.; Garrido-Rodríguez, M.; Navarrete, C.; Caprioglio, D.; Palomares, B.; DeMesa, J.; Rollland, A.; Appendino, G.; Muñoz, E. Cannabinoid Derivatives Acting as Dual PPARγ/CB2 Agonists as Therapeutic Agents for Systemic Sclerosis. Biochem. Pharmacol. 2019, 163, 321–334. [Google Scholar] [CrossRef]
- Ambrosio, A.L.B.; Dias, S.M.G.; Polikarpov, I.; Zurier, R.B.; Burstein, S.H.; Garratt, R.C. Ajulemic Acid, a Synthetic Nonpsychoactive Cannabinoid Acid, Bound to the Ligand Binding Domain of the Human Peroxisome Proliferator-Activated Receptor Gamma. J. Biol. Chem. 2007, 282, 18625–18633. [Google Scholar] [CrossRef]
- Tepper, M.A.; Zurier, R.B.; Burstein, S.H. Ultrapure Ajulemic Acid Has Improved CB2 Selectivity with Reduced CB1 Activity. Bioorganic Med. Chem. 2014, 22, 3245–3251. [Google Scholar] [CrossRef]
- D’Aniello, E.; Fellous, T.; Iannotti, F.A.; Gentile, A.; Allarà, M.; Balestrieri, F.; Gray, R.; Amodeo, P.; Vitale, R.M.; Di Marzo, V. Identification and Characterization of Phytocannabinoids as Novel Dual PPARα/γ Agonists by a Computational and in Vitro Experimental Approach. Biochim. Biophys. Acta—Gen. Subj. 2019, 1863, 586–597. [Google Scholar] [CrossRef]
- Zagzoog, A.; Mohamed, K.A.; Kim, H.J.J.; Kim, E.D.; Frank, C.S.; Black, T.; Jadhav, P.D.; Holbrook, L.A.; Laprairie, R.B. In Vitro and in Vivo Pharmacological Activity of Minor Cannabinoids Isolated from Cannabis Sativa. Sci. Rep. 2020, 10, 20405. [Google Scholar] [CrossRef] [PubMed]
- Iannotti, F.A.; De Maio, F.; Panza, E.; Appendino, G.; Taglialatela-Scafati, O.; De Petrocellis, L.; Amodeo, P.; Vitale, R.M. Identification and Characterization of Cannabimovone, a Cannabinoid from Cannabis Sativa, as a Novel PPARγ Agonist via a Combined Computational and Functional Study. Molecules 2020, 25, 1119. [Google Scholar] [CrossRef]
- Taglialatela-Scafati, O.; Pagani, A.; Scala, F.; De Petrocellis, L.; Di Marzo, V.; Grassi, G.; Appendino, G. Cannabimovone, a Cannabinoid with a Rearranged Terpenoid Skeleton from Hemp. Eur. J. Org. Chem. 2010, 2010, 2067–2072. [Google Scholar] [CrossRef]
- Navarro, G.; Varani, K.; Lillo, A.; Vincenzi, F.; Rivas-Santisteban, R.; Raïch, I.; Reyes-Resina, I.; Ferreiro-Vera, C.; Borea, P.A.; Sánchez de Medina, V.; et al. Pharmacological Data of Cannabidiol- and Cannabigerol-Type Phytocannabinoids Acting on Cannabinoid CB1, CB2 and CB1/CB2 Heteromer Receptors. Pharmacol. Res. 2020, 159, 104940. [Google Scholar] [CrossRef] [PubMed]
- Morales, P.; Vara, D.; Goméz-Cañas, M.; Zúñiga, M.C.; Olea-Azar, C.; Goya, P.; Fernández-Ruiz, J.; Díaz-Laviada, I.; Jagerovic, N. Synthetic Cannabinoid Quinones: Preparation, in Vitro Antiproliferative Effects and in Vivo Prostate Antitumor Activity. Eur. J. Med. Chem. 2013, 70, 111–119. [Google Scholar] [CrossRef]
- Decara, J.M.; Pavón, F.J.; Suárez, J.; Romero-Cuevas, M.; Baixeras, E.; Vázquez, M.; Rivera, P.; Gavito, A.L.; Almeida, B.; Joglar, J.; et al. Treatment with a Novel Oleic-Acid-Dihydroxyamphetamine Conjugation Ameliorates Non-Alcoholic Fatty Liver Disease in Obese Zucker Rats. Dis. Model. Mech. 2015, 8, 1213–1225. [Google Scholar] [CrossRef]
- Alen, F.; Decara, J.; Brunori, G.; You, Z.-B.; Bühler, K.-M.; López-Moreno, J.A.; Cippitelli, A.; Pavon, F.J.; Suárez, J.; Gardner, E.L.; et al. PPARα/CB1 Receptor Dual Ligands as a Novel Therapy for Alcohol Use Disorder: Evaluation of a Novel Oleic Acid Conjugate in Preclinical Rat Models. Biochem. Pharmacol. 2018, 157, 235–243. [Google Scholar] [CrossRef]
- Mazzola, C.; Medalie, J.; Scherma, M.; Panlilio, L.V.; Solinas, M.; Tanda, G.; Drago, F.; Cadet, J.L.; Goldberg, S.R.; Yasar, S. Fatty Acid Amide Hydrolase (FAAH) Inhibition Enhances Memory Acquisition through Activation of PPAR-Alpha Nuclear Receptors. Learn. Mem. 2009, 16, 332–337. [Google Scholar] [CrossRef]
- Han, S.; Inoue, H.; Flowers, L.C.; Sidell, N. Control of COX-2 Gene Expression through Peroxisome Proliferator-Activated Receptor Gamma in Human Cervical Cancer Cells. Clin. Cancer Res. Off. J. Am. Assoc. Cancer Res. 2003, 9, 4627–4635. [Google Scholar]
- Brunetti, L.; Carrieri, A.; Piemontese, L.; Tortorella, P.; Loiodice, F.; Laghezza, A. Beyond the Canonical Endocannabinoid System. A Screening of PPAR Ligands as FAAH Inhibitors. Int. J. Mol. Sci. 2020, 21, 7023. [Google Scholar] [CrossRef]
- Azar, S.; Udi, S.; Drori, A.; Hadar, R.; Nemirovski, A.; Vemuri, K.V.; Miller, M.; Sherill-Rofe, D.; Arad, Y.; Gur-Wahnon, D.; et al. Reversal of Diet-Induced Hepatic Steatosis by Peripheral CB1 Receptor Blockade in Mice Is P53/MiRNA-22/SIRT1/PPARα Dependent. Mol. Metab. 2020, 42, 101087. [Google Scholar] [CrossRef] [PubMed]
- Matias, I.; Gonthier, M.-P.; Orlando, P.; Martiadis, V.; De Petrocellis, L.; Cervino, C.; Petrosino, S.; Hoareau, L.; Festy, F.; Pasquali, R.; et al. Regulation, Function, and Dysregulation of Endocannabinoids in Models of Adipose and Beta-Pancreatic Cells and in Obesity and Hyperglycemia. J. Clin. Endocrinol. Metab. 2006, 91, 3171–3180. [Google Scholar] [CrossRef] [PubMed]
- Guida, F.; Luongo, L.; Boccella, S.; Giordano, M.E.; Romano, R.; Bellini, G.; Manzo, I.; Furiano, A.; Rizzo, A.; Imperatore, R.; et al. Palmitoylethanolamide Induces Microglia Changes Associated with Increased Migration and Phagocytic Activity: Involvement of the CB2 Receptor. Sci. Rep. 2017, 7, 375. [Google Scholar] [CrossRef] [PubMed]
- Ambrosino, P.; Soldovieri, M.V.; De Maria, M.; Russo, C.; Taglialatela, M. Functional and Biochemical Interaction between PPARα Receptors and TRPV1 Channels: Potential Role in PPARα Agonists-Mediated Analgesia. Pharmacol. Res. 2014, 87, 113–122. [Google Scholar] [CrossRef] [PubMed]
- Han, Y.; Liu, J.; Ahn, S.; An, S.; Ko, H.; Shin, J.C.; Jin, S.H.; Ki, M.W.; Lee, S.H.; Lee, K.H.; et al. Diallyl Biphenyl-Type Neolignans Have a Pharmacophore of PPARα/γ Dual Modulators. Biomol. Ther. 2020, 28, 397–404. [Google Scholar] [CrossRef]
- Rempel, V.; Fuchs, A.; Hinz, S.; Karcz, T.; Lehr, M.; Koetter, U.; Müller, C.E. Magnolia Extract, Magnolol, and Metabolites: Activation of Cannabinoid CB2 Receptors and Blockade of the Related GPR55. ACS Med. Chem. Lett. 2013, 4, 41–45. [Google Scholar] [CrossRef]
- Wu, C.; Jia, Y.; Lee, J.H.; Jun, H.; Lee, H.-S.; Hwang, K.-Y.; Lee, S.-J. Trans-Caryophyllene Is a Natural Agonistic Ligand for Peroxisome Proliferator-Activated Receptor-α. Bioorganic Med. Chem. Lett. 2014, 24, 3168–3174. [Google Scholar] [CrossRef]
- Priestley, R.S.; Nickolls, S.A.; Alexander, S.P.H.; Kendall, D.A. A Potential Role for Cannabinoid Receptors in the Therapeutic Action of Fenofibrate. FASEB J. 2015, 29, 1446–1455. [Google Scholar] [CrossRef]
- Pérez-Fernández, R.; Fresno, N.; Macías-González, M.; Elguero, J.; Decara, J.; Girón, R.; Rodríguez-Álvarez, A.; Martín, M.I.; Rodríguez de Fonseca, F.; Goya, P. Discovery of Potent Dual PPARα Agonists/CB1 Ligands. ACS Med. Chem. Lett. 2011, 2, 793–797. [Google Scholar] [CrossRef]




| Phenotype | Includes | Definition | Distribution (%) |
|---|---|---|---|
| A | HA + OD + PCO | Full-blown syndrome PCOS | 67.7% |
| B | HA + OD | Non-PCO PCOS | 11% |
| C | HA + PCO | Ovulatory PCOS | 17.7% |
| D | OD + PCO | Non-hyperandrogenic PCOS | 3.6% |
| Endocannabinoids | |||
 *AEA (Anandamide) (5Z,8Z,11Z,14Z)-N-(2-hydroxyethyl)icosa-5,8,11,14-tetraenamide C22H37NO2 |  *2-AG (2-Arachidonoylglycerol) 1,3-dihydroxypropan-2-yl (5Z,8Z,11Z,14Z)-icosa-5,8,11,14-tetraenoate C23H38O4 | ||
| Endogenous cannabimimetic molecules and derivatives | |||
 NAGLy (N-arachidonoylglycine) 2-[[(5Z,8Z,11Z,14Z)-icosa-5,8,11,14-tetraenoyl]amino]acetic acid C22H35NO3 |  *O-AEA (O-arachidonoyl ethanolamine = virodhamine) 2-aminoethyl (5Z,8Z,11Z,14Z)-icosa-5,8,11,14-tetraenoate C22H37NO2 |  *2-AGE (2-Arachidonyl glyceryl ether = noladin ether = noladin) 2-[(5Z,8Z,11Z,14Z)-icosa-5,8,11,14-tetraenoxy]propane-1,3-diol C23H40O3 | |
 NADA (N-arachidonoyldopamine) (5Z,8Z,11Z,14Z)-N-[2-(3,4-dihydroxyphenyl)ethyl]icosa-5,8,11,14-tetraenamide C28H41NO3 |  *ODA (Oleamide= Oleic acid amide) (Z)-octadec-9-enamide C18H35NO |  *15d-PGJ 2-glycerol ester (15-deoxy-delta12,14-prostaglandin J2-2-glycerol ester) 1,3-dihydroxypropan-2-yl (Z)-7-[(1S,5E)-5-[(Z)-oct-2-enylidene]-4-oxocyclopent-2-en-1-yl]hept-5-enoate C23H34O5 | |
 *15-HETE-glycerol ester (2-(15-Hydroxyeicosatetraenoyl)-sn-glycerol) 1,3-dihydroxypropan-2-yl (5Z,8Z,11Z,13E,15S)-15-hydroxyicosa-5,8,11,13-tetraenoate C23H38O5 |  *OLHHA N-[2-(3,4-dihydroxyphenyl)-1-methylethyl]-9Z-octadecenamide C27H45NO3 | ||
| Endocannabinoid-like compounds | |||
 ***OEA (Oleoylethanolamine) (Z)-N-(2-hydroxyethyl)octadec-9-enamide C20H39NO2 |  *PEA (Palmitoylethanolamide) N-(2-hydroxyethyl)hexadecanamide C18H37NO2 |  SEA (Stearoylethanolamide) N-(2-hydroxyethyl)octadecanamide C20H41NO2 |  LEA (Linoleoylethanolamide) (9Z,12Z)-N-(2-hydroxyethyl)octadeca-9,12-dienamide C20H37NO2 |
| Phytocannabinoids | |||
 *Δ9-THC (delta9-Tetrahydrocannabinol =Dronabinol) (6aR,10aR)-6,6,9-trimethyl-3-pentyl-6a,7,8,10a-tetrahydrobenzo[c]chromen-1-ol C21H30O2 |  *Δ9-THCA (delta(9)-Tetrahydrocannabinolic acid) (6aR,10aR)-1-hydroxy-6,6,9-trimethyl-3-pentyl-6a,7,8,10a-tetrahydrobenzo[c]chromene-2-carboxylic acid C22H30O4 |  *CBD (Cannabidiol) 2-[(1R,6R)-3-methyl-6-prop-1-en-2-ylcyclohex-2-en-1-yl]-5-pentylbenzene-1,3-diol C21H30O2 |  ***CBDA (Cannabidiolic acid) 2,4-dihydroxy-3-[(1R,6R)-3-methyl-6-prop-1-en-2-ylcyclohex-2-en-1-yl]-6-pentylbenzoic acid C22H30O4 |
 Δ9-THCV (delta(9)-Tetrahydrocannabivarin (6aR,10aR)-6,6,9-trimethyl-3-propyl-6a,7,8,10a-tetrahydrobenzo[c]chromen-1-ol C19H26O2 |  CBN (Cannabinol) 6,6,9-trimethyl-3-pentylbenzo[c]chromen-1-ol C21H26O2 |  CBDV (Cannabidivarin = GWP42006) 2-[(1R,6R)-3-methyl-6-prop-1-en-2-ylcyclohex-2-en-1-yl]-5-propylbenzene-1,3-diol C19H26O2 |  *CBC (Cannabichrome) 2-methyl-2-(4-methylpent-3-enyl)-7-pentylchromen-5-ol C21H30O2 |
 *CBG (Cannabigerol) 2-[(2E)-3,7-dimethylocta-2,6-dienyl]-5-pentylbenzene-1,3-diol C21H32O2 |  ***CBGA (Cannabigerolic acid) 3-[(2E)-3,7-dimethylocta-2,6-dienyl]-2,4-dihydroxy-6-pentylbenzoic acid C22H32O4 |  ***CBM (Cannabimovone) 1-[(1R,2R,3R,4R)-3-(2,6-dihydroxy-4-pentylphenyl)-2-hydroxy-4-prop-1-en-2-ylcyclopentyl]ethanone C21H30O4 | |
| Phytocannabinoid derivatives | |||
 *AJA (Ajulemic acid) (6aR,10aR)-1-hydroxy-6,6-dimethyl-3-(2-methyloctan-2-yl)-6a,7,10,10a-tetrahydrobenzo[c]chromene-9-carboxylic acid C25H36O4 |  *CBGQ =VCE003 (Cannabigeroquinone) 2-[(2E)-3,7-dimethylocta-2,6-dienyl]-3-hydroxy-5-pentylcyclohexa-2,5-diene-1,4-dione C21H30O3 |  *Chromenopyrazoledione 4 7-(1′,1′-dimethylheptyl)-1,4-dihydro-4,4-dimethylchromen [4,3-c]pyrazol-6,9-dione C21H28N2O3 | |
 *VCE-004.3 (1′R,6′R)-6-hydroxy-3′-methyl-4-pentyl-3-(pentylamino)-6′-(prop-1-en-2-yl)-[1,1′-bi(cyclohexane)]-2′,3,6-triene-2,5-dione C26H39NO3 |  *VCE-004.8 5-(benzylamino)-4-hydroxy-3-[(1R,6R)-3-methyl-6-prop-1-en-2-ylcyclohex-2-en-1-yl]-6-pentylcyclohexa-3,5-diene-1,2-dione C28H35NO3 | ||
| Synthetic cannabinoids | |||
 *WIN 55,212 [(11R)-2-methyl-11-(morpholin-4-ylmethyl)-9-oxa-1-azatricyclo [6.3.1.04,12]dodeca-2,4(12),5,7-tetraen-3-yl]-naphthalen-1-ylmethanone C27H26N2O3 |  JWH 018 naphthalen-1-yl-(1-pentylindol-3-yl)methanone C24H23NO | ||
 AM 2201 [1-(5-fluoropentyl)indol-3-yl]-naphthalen-1-ylmethanone C24H22FNO |  CP 55940 2-[(1R,2R,5R)-5-hydroxy-2-(3-hydroxypropyl)cyclohexyl]-5-(2-methyloctan-2-yl)phenol C24H40O3 |  HU 210 (6aR,10aR)-9-(hydroxymethyl)-6,6-dimethyl-3-(2-methyloctan-2-yl)-6a,7,10,10a-tetrahydrobenzo[c]chromen-1-ol C25H38O3 | |
| Endogenous PPAR ligands | |||
 AA (Arachidonic acid) (5Z,8Z,11Z,14Z)-icosa-5,8,11,14-tetraenoic acid C20H32O2 |  ALA (alpha-Linolenic acid) (9Z,12Z,15Z)-octadeca-9,12,15-trienoic acidC18H30O2 |  LA (Linolic acid = linoleic acid) (9Z,12Z)-octadeca-9,12-dienoic acid C18H32O2 | |
 9-HODE (9-Hydroxyoctadecadienoic acid,) (10E,12Z)-9-hydroxyoctadeca-10,12-dienoic acid C18H32O3 |  13-HODE (13-Hydroxyoctadecadienoic acid) (9E,11E)-13-hydroxyoctadeca-9,11-dienoic acid C18H32O3 |  8-HETE (8-Hydroxyeicosatetraenoic acid) (5Z,9E,11Z,14Z)-8-hydroxyicosa-5,9,11,14-tetraenoic acid C20H32O3 |  15-HETE (15-Hydroxyeicosatetraenoic acid) (5E,8Z,11Z,13Z)-15-hydroxyicosa-5,8,11,13-tetraenoic acid C20H32O3 |
 LTB4 (Leukotriene B4) (5S,6Z,8E,10E,12R,14Z)-5,12-dihydroxyicosa-6,8,10,14-tetraenoic acid C20H32O4 |  15d-PGJ 2 (15-deoxy-delta12,14-prostaglandin J2) (Z)-7-[(1S,5E)-5-[(E)-oct-2-enylidene]-4-oxocyclopent-2-en-1-yl]hept-5-enoic acid C20H28O3 |  PGI 2 (Prostaglandin I2 = Epoprostenol = Prostacyclin) (5Z)-5-[(3aR,4R,5R,6aS)-5-hydroxy-4-[(E,3S)-3-hydroxyoct-1-enyl]-3,3a,4,5,6,6a-hexahydrocyclopenta[b]furan-2-ylidene]pentanoic acid C20H32O5 | |
| Fibrates | |||
 Clofibrate ethyl 2-(4-chlorophenoxy)-2-methylpropanoate C12H15ClO3 |  Bezafibrate 2-[4-[2-[(4-chlorobenzoyl)amino]ethyl]phenoxy]-2-methylpropanoic acid C19H20ClNO4 |  *****Fenofibrate propan-2-yl 2-[4-(4-chlorobenzoyl)phenoxy]-2-methylpropanoate C20H21ClO4 |  Gemfibrozil 5-(2,5-dimethylphenoxy)-2,2-dimethylpentanoic acid C15H22O3 |
| Synthetic PPAR β/δ agonists | |||
 GW 0742 2-[4-[[2-[3-fluoro-4-(trifluoromethyl)phenyl]-4-methyl-1,3-thiazol-5-yl]methylsulfanyl]-2-methylphenoxy]acetic acid C21H17F4NO3S2 |  GW 501516 (Endurobol) 2-[2-methyl-4-[[4-methyl-2-[4-(trifluoromethyl)phenyl]-1,3-thiazol-5-yl]methylsulfanyl]phenoxy]acetic acid C21H18F3NO3S2 | ||
| Thiazolidinediones | |||
 Pioglitazone 5-[[4-[2-(5-ethylpyridin-2-yl)ethoxy]phenyl]methyl]-1,3-thiazolidine-2,4-dione C19H20N2O3S |  Troglitazone 5-[[4-[(6-hydroxy-2,5,7,8-tetramethyl-3,4-dihydrochromen-2-yl)methoxy]phenyl]methyl]-1,3-thiazolidine-2,4-dione C24H27NO5S |  Rosiglitazone 5-[[4-[2-[methyl(pyridin-2-yl)amino]ethoxy]phenyl]methyl]-1,3-thiazolidine-2,4-dione C18H19N3O3S |  Ciglitazone 5-[[4-[(1-methylcyclohexyl)methoxy]phenyl]methyl]-1,3-thiazolidine-2,4-dione C18H23NO3S |
| Other non-cannabinoid dual ligands of CBRs and PPARs | |||
 *****Magnolol 2-(2-hydroxy-5-prop-2-enylphenyl)-4-prop-2-enylphenol C18H18O2 |  *****Honokiol 2-(4-hydroxy-3-prop-2-enylphenyl)-4-prop-2-enylphenol C18H18O2 |  *****BCP (beta-Caryophyllene) (1R,4E,9S)-4,11,11-trimethyl-8-methylidenebicyclo [7.2.0]undec-4-eneC15H24 |  *****Rimonabant fibrate |
| Other | |||
 SR141716 (Rimonabant) 5-(4-chlorophenyl)-1-(2,4-dichlorophenyl)-4-methyl-N-piperidin-1-ylpyrazole-3-carboxamide C22H21Cl3N4O | |||
| Ligand | CBRs | PPARs | ||
|---|---|---|---|---|
| CB1R | CB2R | PPARα | PPARγ | |
| Magnolol [269,270] | (+) agonist | (+) agonist | (+) agonist | (+) agonist |
| Honokiol [269,270] | (+) agonist | (−) antagonist | (+) agonist | (+) agonist |
| BCP [271] | (+) agonist | (+) agonist | ||
| Fenofibrate [272] | (+) agonist | (+) agonist | (+) agonist | |
| Rimonabant fibrate 2 [273] | (−) antagonist | (+) agonist | ||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Przybycień, P.; Gąsior-Perczak, D.; Placha, W. Cannabinoids and PPAR Ligands: The Future in Treatment of Polycystic Ovary Syndrome Women with Obesity and Reduced Fertility. Cells 2022, 11, 2569. https://doi.org/10.3390/cells11162569
Przybycień P, Gąsior-Perczak D, Placha W. Cannabinoids and PPAR Ligands: The Future in Treatment of Polycystic Ovary Syndrome Women with Obesity and Reduced Fertility. Cells. 2022; 11(16):2569. https://doi.org/10.3390/cells11162569
Chicago/Turabian StylePrzybycień, Piotr, Danuta Gąsior-Perczak, and Wojciech Placha. 2022. "Cannabinoids and PPAR Ligands: The Future in Treatment of Polycystic Ovary Syndrome Women with Obesity and Reduced Fertility" Cells 11, no. 16: 2569. https://doi.org/10.3390/cells11162569