
Phylogeny and Fatty Acid Profiles of New Pinnularia (Bacillariophyta) Species from Soils of Vietnam


Abstract
:1. Introduction
2. Materials and Methods
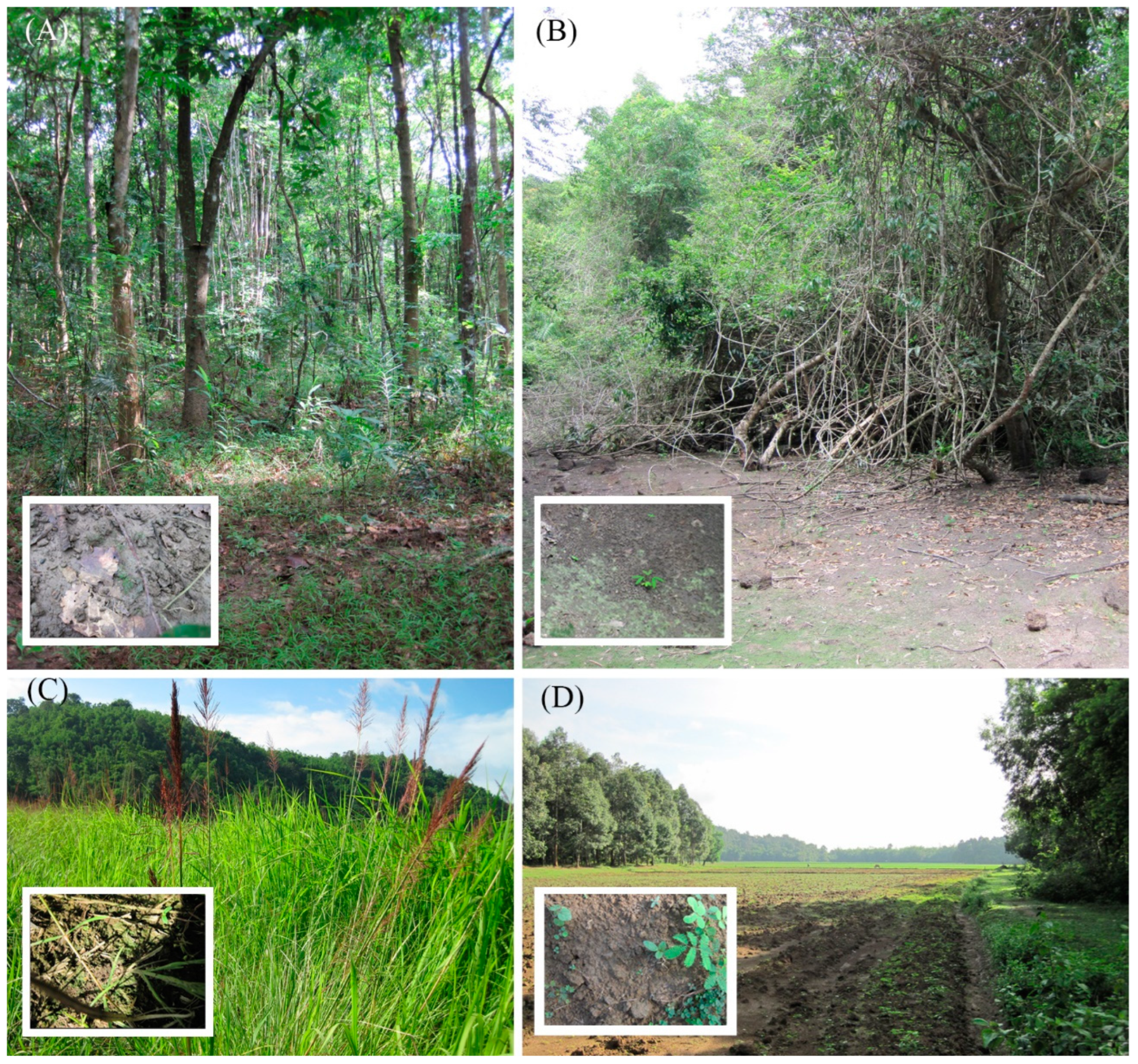
2.1. Sample Collection Procedure
2.2. Culturing
2.3. Microscopy
2.4. Molecular Study
2.5. Fatty Acid Analysis
3. Results and Discussion
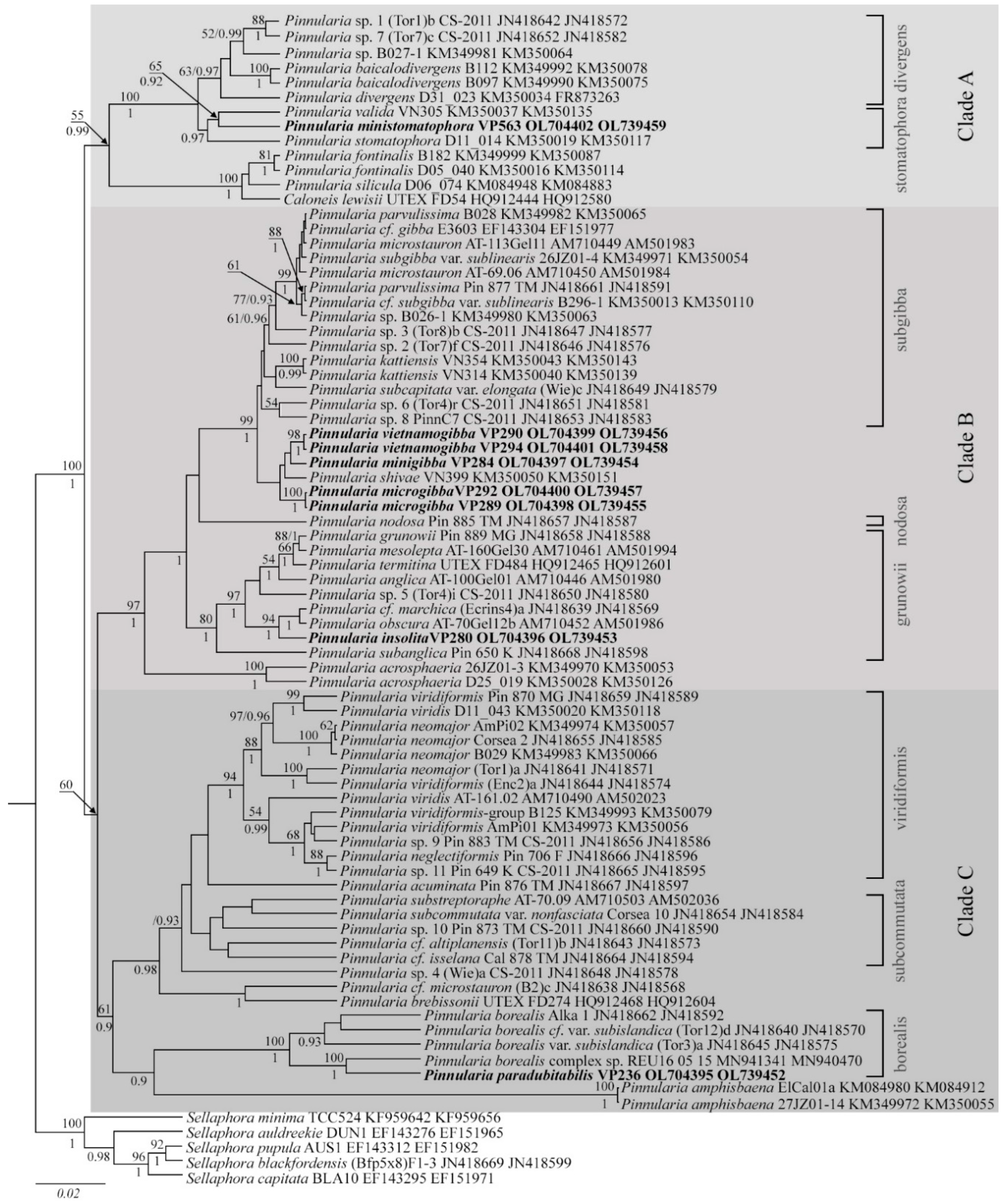
3.1. Molecular Phylogeny
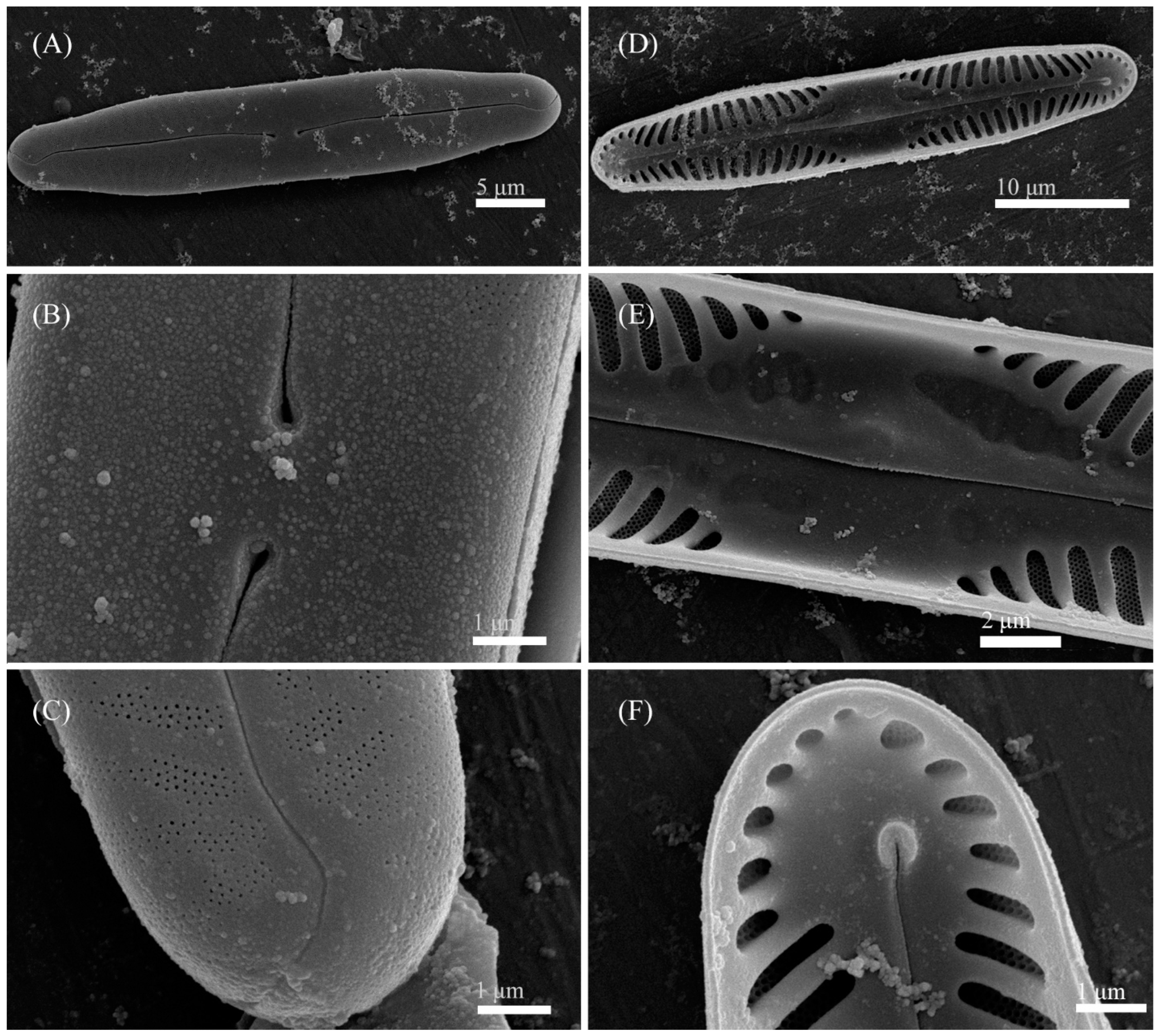
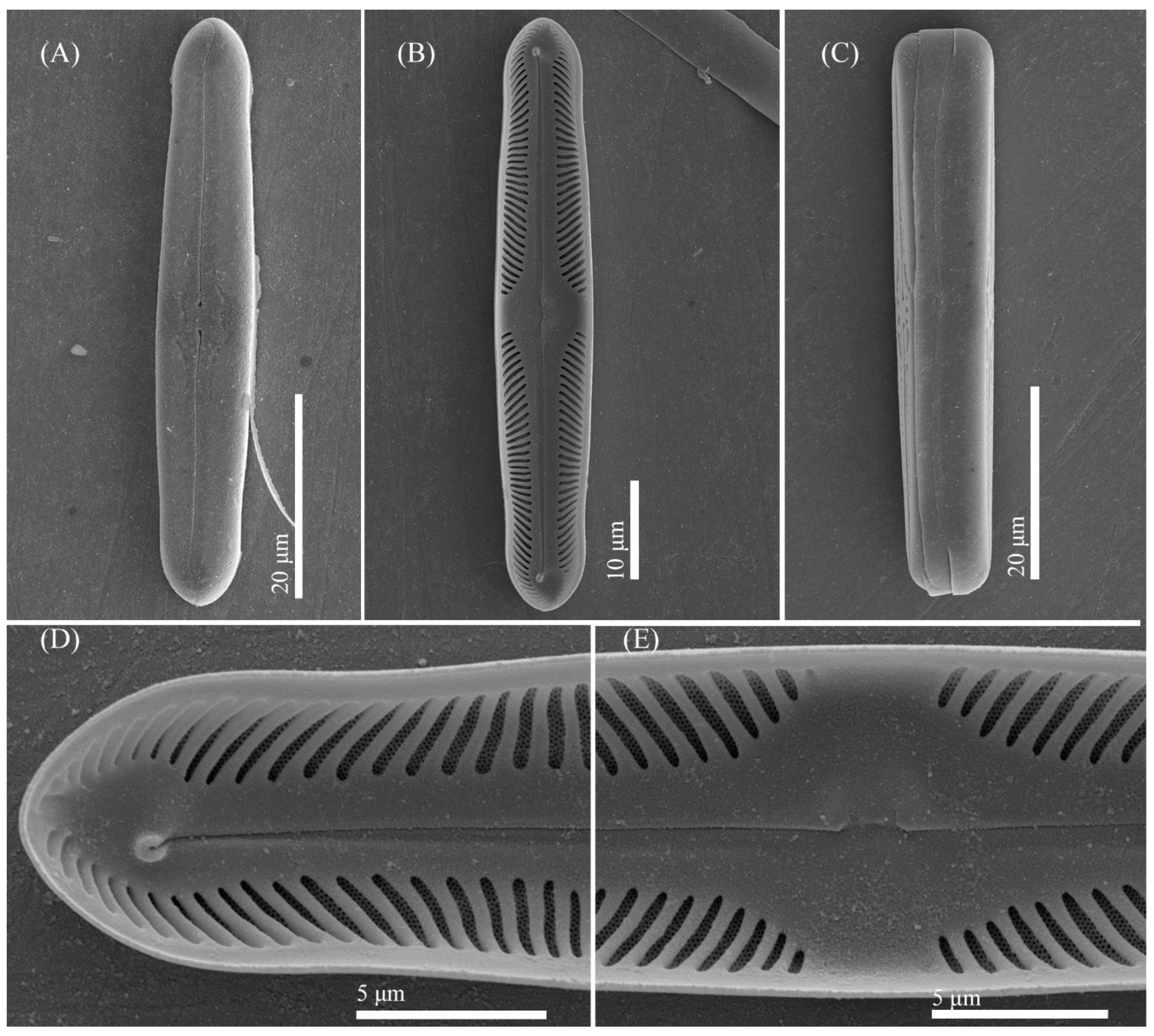
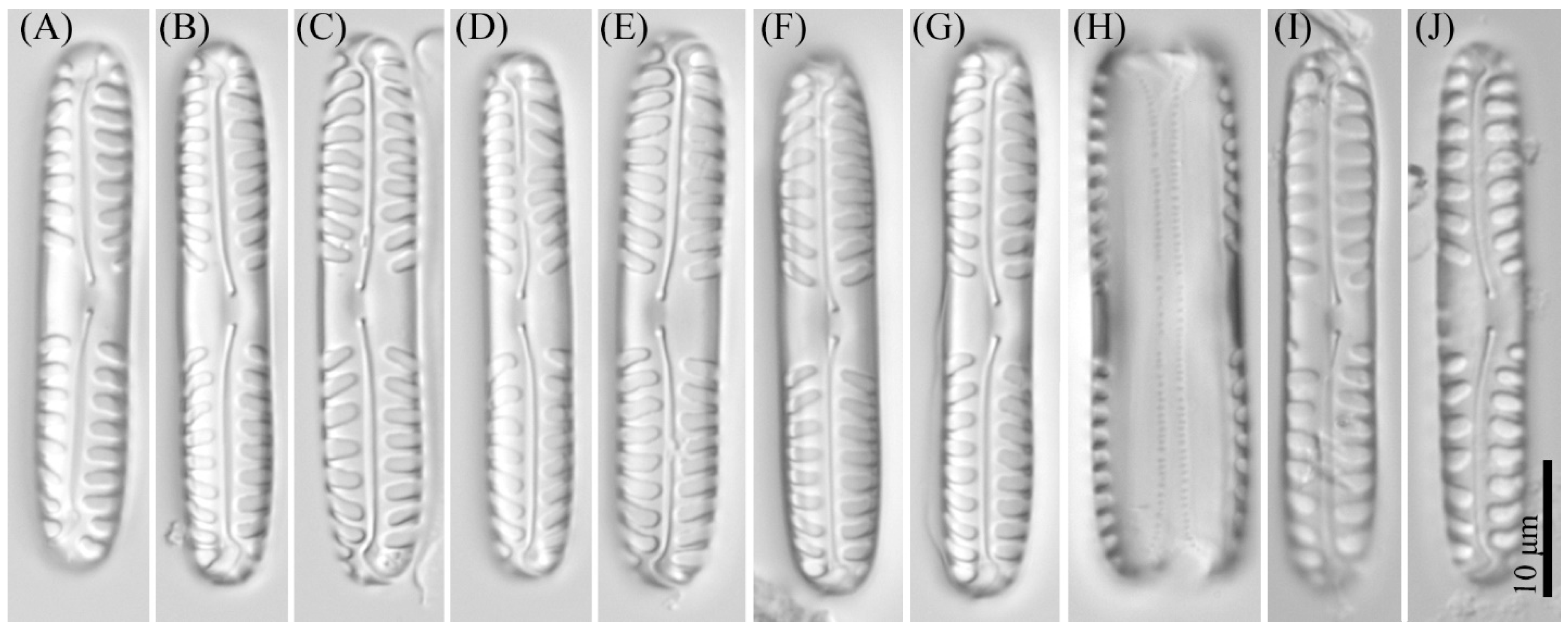
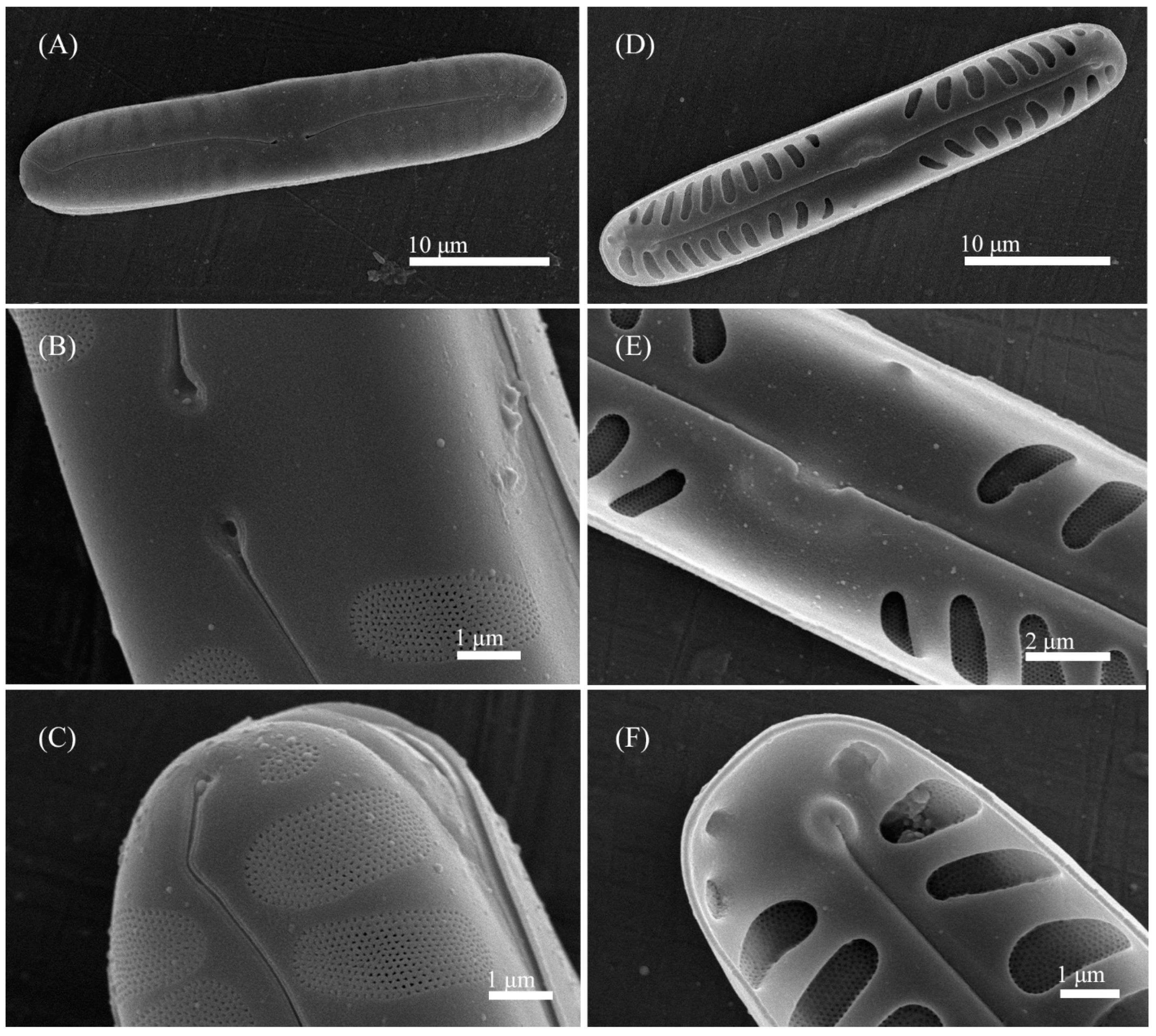
3.2. Comparative Morphology
3.3. Fatty Acid Profiles
3.4. Description of New Species
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Pinseel, E.; Kulichová, J.; Scharfen, V.; Urbánková, P.; Van de Vijver, B.; Vyverman, W. Extensive cryptic diversity in the terrestrial diatom Pinnularia borealis (Bacillariophyceae). Protist 2019, 170, 121–140. [Google Scholar] [CrossRef] [PubMed]
- Verma, K. Role of diatoms in the world of forensic science. J. Forensic Res. 2013, 4, 1000181. [Google Scholar] [CrossRef]
- Kuppusamy, P.; Soundharrajan, I.; Srigopalram, S.; Yusoff, M.M.; Maniam, G.P.; Govindan, N.; Choi, K.C. Potential pharmaceutical and biomedical applications of diatoms microalgae-An overview. J. Mar. Sci. 2017, 46, 663–667. [Google Scholar]
- Krishna, P.M.; Polisetti, V.; Damarla, K.; Mandal, S.K.; Kumar, A. Improved biorefinery pathways of marine diatoms using a water miscible ionic liquid and its colloidal solution: Efficient lipid extraction and in situ synthesis of fluorescent carbon dots for bio-imaging applications. RSC Adv. 2021, 11, 21207–21215. [Google Scholar] [CrossRef] [PubMed]
- Khan, M.J.; Bawra, N.; Verma, A.; Kumar, V.; Pugazhendhi, A.; Joshi, K.B.; Vinayak, V. Cultivation of diatom Pinnularia saprophila for lipid production: A comparison of methods for harvesting the lipid from the cells. Bioresource Technol. 2021, 319, 124–129. [Google Scholar] [CrossRef] [PubMed]
- Maltsev, Y.; Maltseva, K. Fatty acids of microalgae: Diversity and applications. Rev. Environ. Sci. Biotechnol. 2021, 20, 515–547. [Google Scholar] [CrossRef]
- Arao, T.; Yamada, M. Biosynthesis of polyunsaturated fatty acids in the marine diatom, Phaeodactylum tricornutum. Phytochemistry 1994, 35, 1177–1181. [Google Scholar] [CrossRef]
- Marella, T.K.; Tiwari, A. Marine diatom Thalassiosira weissflogii based biorefinery for co-production of eicosapentaenoic acid and fucoxanthin. Bioresour. Technol. 2020, 307, 123245. [Google Scholar] [CrossRef]
- Kulikovskiy, M.; Glushchenko, A.; Genkal, S.I.; Kuznetsova, I. Identification Book of Diatoms from Russia; Filigran: Yaroslavl, Russia, 2016; 803p. [Google Scholar]
- Guiry, M.D.; Guiry, G.M. AlgaeBase. World-Wide Electronic Publication, National University of Ireland, Galway. Available online: http://www.algaebase.org (accessed on 11 July 2022).
- Krammer, K. The Genus Pinnularia; ARG Ganter Verlag KG: Ruggell, Liechtenstein, 2000; 703p. [Google Scholar]
- Metzeltin, D.; Lange-Bertalot, H. Tropical Diatoms of South America II. Special Remarks on Biogeography Disjunction. Iconographia Diatomologica 18; ARG Gantner: Ruggell, Liechtenstein, 2007; 877p. [Google Scholar]
- Barragán, C.; Wetzel, C.; Ector, L. A standard method for the routine sampling of terrestrial diatom communities for soil quality assessment. J. Appl. Phycol. 2018, 30, 1095–1113. [Google Scholar] [CrossRef]
- Foets, J.; Stanek-Tarkowska, J.; Teuling, A.; Van de Vijver, B.; Wetzel, C.; Pfister, L. Autecology of terrestrial diatoms under anthropic disturbance and across climate zones. Ecol. Indic. 2021, 122, 107248. [Google Scholar] [CrossRef]
- Novakovskaya, I.V.; Patova, E.N. Soil Algae of Spurce Forests and Their Change under Conditions of Aero-Technogenic Pollution; Biologia: Syktyvkar, Russian, 2011; 128p. [Google Scholar]
- Fazlutdinova, A.; Gabidullin, Y.; Allaguvatova, R.; Gaysina, L. Diatoms in Volcanic Soils of Mutnovsky and Gorely Volcanoes (Kamchatka Peninsula, Russia). Microorganisms 2021, 9, 1851. [Google Scholar] [CrossRef] [PubMed]
- Stanek-Tarkowska, J.; Czyz, E.A.; Kaniuczak, J.; Poradowska, A. Physicochemical properties of silt loamy soil and diversity of diatom species under winter wheat and oats. J. Ecol. Eng. 2017, 18, 142–151. [Google Scholar] [CrossRef]
- Stanek-Tarkowska, J.; Szostek, M.; Rybak, M. Effect of different doses of ash from biomass combustion on the development of diatom assemblages on podzolic soil under oilseed rape cultivation. Agronomy 2021, 11, 2422. [Google Scholar] [CrossRef]
- Antonelli, M.; Wetzel, C.; Ector, L.; Teuling, A.J.; Pfister, A. On the potential for terrestrial diatom communities and diatom indices to identify anthropic disturbance in soils. Ecol. Indic. 2017, 75, 73–81. [Google Scholar] [CrossRef]
- Johansen, J.R.; Rushforth, S.R.; Brotherson, J.D. Subaerial algae of Navajo National Monument, Arizona. Great Basin Nat. 1981, 41, 8. Available online: https://scholarsarchive.byu.edu/gbn/vol41/iss4/8 (accessed on 11 July 2022).
- Ohtsuka, T.; Fujita, Y. The diatom flora and its seasonal changes in a paddy field in Central Japan. Nova Hedwig. 2001, 73, 97–128. [Google Scholar] [CrossRef]
- Van de Vijver, B.; Ledeganck, P.; Beyens, L. Soil diatom communities from Ile de la Possession (Crozet, sub-Antarctica). Polar. Biol. 2002, 25, 721–729. [Google Scholar] [CrossRef]
- Rybak, M.; Noga, T.; Zubel, R. The aerophytic diatom assemblages developed on mosses covering the bark of Populus alba L. J. Ecol. Eng. 2018, 19, 113–123. [Google Scholar] [CrossRef]
- Bruder, K.; Medlin, L.K. Morphological and molecular investigations of naviculoid diatoms. II. Selected genera and families. Diatom Res. 2008, 23, 283–329. [Google Scholar] [CrossRef]
- Souffreau, C.; Verbruggen, H.; Wolfe, A.P.; Vanormelingen, P.; Siver, P.A.; Cox, E.J.; Mann, D.G.; Van de Vijver, B.; Sabbe, K.; Vyverman, W. A time-calibrated multi-gene phylogeny of the diatom genus Pinnularia. Mol. Phylogen. Evol. 2011, 61, 866–879. [Google Scholar] [CrossRef]
- Kollár, J.; Pinseel, E.; Vanormelingen, P.; Poulíčková, A.; Souffreau, C.; Dvořák, P.; Vyverman, W. A polyphasic approach to the delimitation of diatom species: A case study for the genus Pinnularia (Bacillariophyta). J. Phycol. 2019, 55, 365–379. [Google Scholar] [CrossRef]
- Kollár, J.; Pinseel, E.; Vyverman, W.; Poulíčková, A. A time-calibrated multi-gene phylogeny provides insights into the evolution, taxonomy and DNA barcoding of the Pinnularia gibba group (Bacillariophyta). Fottea 2021, 21, 62–72. [Google Scholar] [CrossRef]
- Souffreau, C.; Vanormelingen, P.; Van de Vijver, B.; Isheva, T.; Verleyen, E.; Sabbe, K.; Vyverman, W. Molecular evidence for distinct Antarctic lineages in the cosmopolitan terrestrial diatoms Pinnularia borealis and Hantzschia amphioxys. Protist 2013, 164, 101–115. [Google Scholar] [CrossRef] [PubMed]
- Pinseel, E.; Janssens, S.B.; Verleyen, E.; Vanormelingen, P.; Kohler, T.J.; Biersma, E.M.; Sabbe, K.; Van de Vijver, B.; Vyverman, W. Global radiation in a rare biosphere soil diatom. Nat. Commun. 2020, 11, 2382. [Google Scholar] [CrossRef]
- Mann, D.G.; McDonald, S.M.; Bayer, M.M.; Droop, S.J.M.; Chepurnov, V.A.; Loke, R.E.; Ciobanu, A.; Du Buf, J.M.H. The Sellaphora pupula species complex (Bacillariophyceae): Morphometric analysis, ultrastructure and mating data provide evidence for five new species. Phycologia 2004, 43, 459–482. [Google Scholar] [CrossRef] [Green Version]
- Trobajo, R.; Clavero, E.; Chepurnov, V.A.; Sabbe, K.; Mann, D.G.; Ishihara, S.; Cox, E.J. Morphological, genetic, and mating diversity within the widespread bioindicator Nitzschia palea (Bacillariophyceae). Phycologia 2009, 48, 443–459. [Google Scholar] [CrossRef]
- Maltsev, Y.; Maltseva, S.; Kociolek, J.P.; Jahn, R.; Kulikovskiy, M. Biogeography of the cosmopolitan terrestrial diatom Hantzschia amphioxys sensu lato based on molecular and morphological data. Sci. Rep. 2021, 11, 4266. [Google Scholar] [CrossRef]
- Hodač, L. Green Algae in Soil: Assessing Their Biodiversity and Biogeography with Molecular-Phylogenetic Methods Based on Cultures. Ph.D. Thesis, Georg August University of Göttingen, Göttingen, Germany, 2016; 188p. [Google Scholar]
- Rybak, M.; Kochman-Kędziora, N.; Peszek, L. Description of four new terrestrial diatom species from Luticola and Microcostatus genera from South Africa. PhytoKeys 2021, 182, 1–26. [Google Scholar] [CrossRef]
- Hameed, M.S.A. Survey of soil algal flora of some cultivated soils in Beni Suef, Egypt. Egypt. J. Phycol. 2006, 7, 1–15. [Google Scholar] [CrossRef]
- Fujita, Y.; Ohtsuka, T. Diatoms from paddy fields in northern Laos. Diatom 2005, 21, 71–89. [Google Scholar] [CrossRef]
- Vijayan, D.; Ray, J.G. Green algae of a unique tropical wetland Kuttanadu, Kerala, India in relation to soil regions, seasons and paddy growth stages. Int. J. Sci. Environ. Technol. 2015, 4, 770–803. [Google Scholar]
- Hodač, L.; Hallmann, C.; Rosenkranz, H.; Faßhauer, F.; Friedl, T. Molecular evidence for the wide distribution of two lineages of terrestrial green algae (Chlorophyta) over tropics to temperate zone. ISRN Ecol. 2012, 2012, 795924. [Google Scholar] [CrossRef] [Green Version]
- Kezlya, E.; Glushchenko, A.; Kociolek, J.P.; Maltsev, Y.; Martynenko, N.; Genkal, S.; Kulikovskiy, M. Mayamaea vietnamica sp. nov.: A new, terrestrial diatom (Bacillariophyceae) species from Vietnam. Algae 2020, 35, 325–335. [Google Scholar] [CrossRef]
- Kezlya, E.M.; Glushchenko, A.M.; Maltsev, Y.I.; Gusev, E.S.; Genkal, S.I.; Kuznetsov, A.N.; Kociolek, J.P.; Kulikovskiy, M.S. Placoneis cattiensis sp. nov.—A new, diatom (Bacillariophyceae: Cymbellales) soil species from Cát Tiên National Park (Vietnam). Phytotaxa 2020, 460, 237–248. [Google Scholar] [CrossRef]
- Blanc, L.; Maury-Lechon, G.; Pascal, J.-P. Structure, floristic composition and natural regeneration in the forests of Cat Tien National Park, Vietnam: An analysis of the successional trends. J. Biogeogr. 2001, 27, 141–157. [Google Scholar] [CrossRef]
- Vadjunina, A.F.; Korchagina, Z.A. Methods of Studying the Physical Properties of Soils; Agropromisdat: Moscow, Russian, 1986; 416p. [Google Scholar]
- Arinushkina, E.W. Handbook for Chemical Soil Analysis; Moscow State University: Moscow, Russian, 1970; 488p. [Google Scholar]
- McFadden, G.I.; Melkonian, M. Use of Hepes buffer for microalgal culture media and fixation for electron microscopy. Phycologia 1986, 25, 551–557. [Google Scholar] [CrossRef]
- Zimmermann, J.; Jahn, R.; Gemeinholzer, B. Barcoding diatoms: Evaluation of the V4 subregion on the 18S rRNA gene, including new primers and protocols. Org. Divers. Evol. 2011, 11, 173–192. [Google Scholar] [CrossRef]
- Ruck, E.C.; Theriot, E.C. Origin and evolution of the canal raphe system in diatoms. Protist 2011, 162, 723–737. [Google Scholar] [CrossRef] [PubMed]
- Alverson, A.J.; Jansen, R.K.; Theriot, E.C. Bridging the Rubicon: Phylogenetic analysis reveals repeated colonizations of marine and fresh waters by thalassiosiroid diatoms. Mol. Phylogen. Evol. 2007, 45, 193–210. [Google Scholar] [CrossRef] [PubMed]
- Kumar, S.; Stecher, G.; Tamura, K. MEGA7: Molecular evolutionary genetics analysis version 7.0 for bigger datasets. Mol. Biol. Evol. 2016, 33, 1870–1874. [Google Scholar] [CrossRef] [Green Version]
- Katoh, K.; Toh, H. Parallelization of the MAFFT multiple sequence alignment program. Bioinformatics 2010, 26, 1899–1900. [Google Scholar] [CrossRef] [PubMed]
- Drummond, A.J.; Rambaut, A. BEAST: Bayesian evolutionary analysis by sampling trees. BMC Evol. Biol. 2007, 7, 214. [Google Scholar] [CrossRef] [Green Version]
- Darriba, D.; Taboada, G.L.; Doallo, R.; Posada, D. jModelTest 2: More models, new heuristics and parallel computing. Nat. Methods 2012, 9, 772. [Google Scholar] [CrossRef] [Green Version]
- Stamatakis, A.; Hoover, P.; Rougemont, J. A rapid bootstrap algorithm for the RAxML web–servers. Syst. Biol. 2008, 57, 758–771. [Google Scholar] [CrossRef]
- Maltsev, Y.; Krivova, Z.; Maltseva, S.; Maltseva, K.; Gorshkova, E.; Kulikovskiy, M. Lipid accumulation by Coelastrella multistriata (Scenedesmaceae, Sphaeropleales) during nitrogen and phosphorus starvation. Sci. Rep. 2021, 11, 19818. [Google Scholar] [CrossRef] [PubMed]
- Cox, E.J. Variation in patterns of valve morphogenesis between representatives of six biraphid diatom genera. J. Phycol. 1999, 35, 1297–1312. [Google Scholar] [CrossRef]
- Van de Vijver, B.; Chattová, B.; Metzeltin, D.; Lebouvier, M. The genus Pinnularia (Bacillariophyta) on Île Amsterdam (TAAF, Southern Indian Ocean). Nova Hedwig. 2012, 141, 201–236. [Google Scholar]
- Jahn, R.; Kusber, W.-H. AlgaTerra Information System. Botanic Garden and Botanical Museum Berlin-Dahlem. Freie Universität Berlin. Available online: https://www.algaterra.org (accessed on 11 July 2022).
- Metzeltin, D.; Lange-Bertalot, H.; García-Rodrígues, F. Diatoms of Uruguay. Compared with Other Taxa from South America and Elsewhere. Iconographia Diatomologica 15; ARG Gantner: Ruggell, Liechtenstein, 2005; 736p. [Google Scholar]
- Manguin, E. Contribution à la connaissance des Diatomées des Andes du Pérou. Mémoires du Museum National d’Histoire Naturelle, nouvelle série. Série B Bot. 1964, 12. [Google Scholar]
- Rumrich, U.; Lange-Bertalot, H.; Rumrich, M. Diatoms of the Andes. From Venezuela to Patagonia/Tierra del Fuego and Two Additional Contributions. Iconographia Diatomologica, 9; Koeltz Scientific Books: Königstein, Germany, 2000; 673p. [Google Scholar]
- Hustedt, F. Süsswasser-Diatomeen aus dem Albert-Nationalpark in Belgisch-Kongo. Exploration du Parc National Albert, Mission H. Damas (1935-1936); Institut des Parcs Nationaux du Congo Belge: Bruxelles, Belgium, 1949; 199p. [Google Scholar]
- Noga, T.; Kochman, N.; Peszek, Ł.; Stanek-Tarkowska, J.; Pajączek, A. Diatoms (Bacillariophyceae) in rivers and streams and on cultivated soil of the Podkarpacie region in the years 2007–2011. J. Ecol. Eng. 2014, 15, 6–25. [Google Scholar] [CrossRef]
- Lange-Bertalot, H.; Cavacini, P.; Tagliaventi, N.; Alfinito, S. Diatoms of Sardinia. Rare and 76 New Species in Rock Pools and Other Ephemeral Waters. Iconographia Diatomologica 12; ARG Gantner: Ruggell, Liechtenstein, 2003; 438p. [Google Scholar]
- Krammer, K. Pinnularia eine Monographie der Europäischen Taxa. Bibliotheca Diatomologica, 26; Stuttgart Cramer in der Gebr-Borntraeger-Verl-Buchh: Berlin, Germany, 1992; 353p. [Google Scholar]
- Siver, P.; Hamilton, P. Diatoms of North America. The Freshwater Flora of Waterbodies on the Atlantic Coastal Plain. Iconographia Diatomologica, 22; ARG Gantner: Ruggell, Liechtenstein, 2011; 916p. [Google Scholar]
- Karthick, B.; Hamilton, P.B.; Kociolek, J.P. An Illustrated Guide to Common Diatoms of Peninsular India; Gubbi Labs: Karnataka, India, 2013; 208p. [Google Scholar]
- Liu, Y.; Kociolec, J.P.; Wang, Q.X.; Fan, Y.W. The Diatom Genus Pinnularia from Great Xing’an Mountains, China. Bibliotheca Diatomologica, 65; Cramer in Borntraeger Science Publishers: Stuttgart, Germany, 2018; 298p. [Google Scholar]
- Krammer, K.; Lange-Bertalot, H. 1988. Bacillariophyceae. 2. Teil: Bacillariaceae, Epithemiaceae, Surirellaceae. In Susswasserflora von Mitteleuropa, Band 2/2; Ettl, H., Gerloff, J., Heynig, H., Mollenhauer, D., Eds.; Gustav Fisher Verlag: Jena, Germany, 1988; 596p. [Google Scholar]
- Moreno, R.R.; Aita, G.M.; Madsen, L.; Gutierrez, D.L.; Yao, S.; Hurlburt, B.; Brashear, S. Identification of naturally isolated Southern Louisiana’s algal strains and the effect of higher CO2 content on fatty acid profiles for biodiesel production. J. Chem. Technol. Biotechnol. 2013, 88, 948–957. [Google Scholar] [CrossRef]
- Bedoshvili, Y.; Podunay, Y.; Nikonova, A.; Marchenkov, A.; Bairamova, E.; Davidovich, N.; Likhoshway, Y. Lipid and fatty acids accumulation features of Entomoneis cf. paludosa during exponential and stationary growth phases in laboratory culture. Diversity 2021, 13, 459. [Google Scholar] [CrossRef]
- Lang, I.; Hodac, L.; Friedl, T.; Feussner, I. Fatty acid profiles and their distribution patterns in microalgae: A comprehensive analysis of more than 2000 strains from the SAG culture collection. BMC Plant Biol. 2011, 11, 124. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Knothe, G. A comprehensive evaluation of the cetane numbers of fatty acid methyl esters. Fuel 2014, 119, 6–13. [Google Scholar] [CrossRef]
- Foged, N. Diatoms in Thailand. Nova Hedwigia 1971, 22, 267–369. [Google Scholar]















{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sample ID | Collection Date | Sample Locality | Habitat Type | Substratum | pH | Absolute Humidity, % | Species, Strains |
|---|---|---|---|---|---|---|---|
| KT19 | 5 June 2019 | N11°26′8.13″ E107°26′42.9″ Cát Tiên National Park, Vietnam | forest | soil | 5.0 | 21.32 | P. paradubitabilis |
| KT53 | 16 June 2019 | N11°26′9.75″ E107°21′46.2″ Cát Tiên National Park, Vietnam | forest | basalt | n.a. | n.a. | P. paradubitabilis, VP236 |
| KT68 | 19 March 2020 | N11°26′63.1″ E107°23′52.2″ Cát Tiên National Park, Vietnam | forest | soil | 5.4 | n.a. | P. paradubitabilis |
| 965 975 | 23 November 2011 | N18°36′8.08″ E102°24′6.05″ Vientiane province, Vang Vieng region, Nam Lik village, Laos | Nam Lik river | benthos periphyton | n.a. | n.a. | P. paradubitabilis |
| KT80 | 19 March 2020 | N11°27′48.3″ E107°20′8.27″ Cát Tiên National Park, Vietnam | forest | soil | n.a. | n.a. | P. microgibba |
| KT39 | 7 June 2019 | N11°26′7.52″ E107°23′17.2″ Cát Tiên National Park, Vietnam | forest, the bottom of a dry reservoir | soil | 5.19 | 36.79 | P. microgibba P. ministomatophora, VP563 |
| KT61 | 25 June 2019 | N11°24′24.7″ E107°23′6.61″ Cát Tiên National Park, Vietnam | dry swamp | soil | 5.11 | 49.02 | P. microgibba, VP289, VP292 P. vietnamogibba, VP290, VP294 |
| KT70 | 19 March 2020 | N11°26′6.60″ E107°23′7.39″ Cát Tiên National Park, Vietnam | forest, the bottom of a dry reservoir | soil | 4.7 | n.a. | P. ministomatophora |
| KT54 | 25 June 2019 | N11°23′35.8″ E107°21′9.48″ Cát Tiên National Park, Vietnam | agricultural field | soil | 4.91 | 24.68 | P. minigibba, VP284 |
| KT55 | 25 June 2019 | N11°23′9.17″ E107°22′52.3″ Cát Tiên National Park, Vietnam | forest | soil | 5.55 | 44.8 | P. insolita, VP280 |
| Species, Strains | Outline, Apices | Valve Length, μm | Valve Width, μm | Number of Striae in 10 μm | Axial Area | Central Area | Markings on The Central Area | References |
|---|---|---|---|---|---|---|---|---|
| P. minigibba Kezlya, Maltsev, Krivova et Kulikovskiy, VP284 | linear, slightly concave, apices subcapitate | 40–43 | 7–8 | 9–10 | narrow, linear, to 1/4 the breadth of valve | large, rhombic with a broad slightly asymmetric fascia | internally: ghost striae irregular in the shape | this study |
| P. vietnamogibba Kezlya, Maltsev, Krivova et Kulikovskiy, VP290, VP294 | linear, linear-elliptical, slightly convex with broadly rounded apices | 34–54 | 7–8.5 | 10–11 | moderately broad about 1/4 the breadth of valve | large, rhombic with a broad slightly asymmetric fascia | internally: ghost striae irregular in the shape | this study |
| P. microgibba Kezlya, Maltsev, Krivova et Kulikovskiy, VP289, VP292 | narrow-linear, slightly concave, apices subcapitate | 35–40 | 5.5–6.0 | 11–12 | narrow, linear | large, rhombic with a broad slightly asymmetric fascia | internally: ghost striae irregular in the shape | this study |
| P. insolita Kezlya, Maltsev, Krivova et Kulikovskiy, VP280 | linear, slightly concave, apices rostrate | 50–52 | 7–7.5 | 11–12 | moderately broad, to 1/3 the breadth of valve | very large with a broad slightly asymmetric fascia widening towards the valve margin | no | this study |
| P. ministomatophora Kezlya, Maltsev, Krivova et Kulikovskiy, VP563 | linear, slightly convex, apices subcapitate | 44–57 | 7–9.5 | 10–11 | narrow, up to 1/4 the breadth of valve | large with a broad slightly asymmetric fascia widening towards the valve margin | externally: hollows in the valve surface irregular in the shape | this study |
| P. paradubitabilis Kezlya, Maltsev, Krivova et Kulikovskiy, VP236 | linear with parallel or slightly concave margins, apices obtusely rounded | 39–44 | 6–7 | 5–6 | narrow, linear | large with a broad slightly asymmetric fascia widening towards the valve margin | no | this study |
| P. australogibba var. subcapitata Van de Vijver, Chattová et Metzeltin | lanceolate to narrowly lanceolate with subcapitate apices | 22–45 | 5.3–7.3 | 11–12 | moderately broad, lanceolate, | large, rhombic–rounded, forming a broad fascia | internally: ghost striae | [55] |
| P. parvulissima Krammer | linear, slightly convex, apices very broadly rostrate to subcapitate and broadly rounded | 34–70 | 10–12 | 8–10 | 1/4–1/3 the valve breadth | with a moderately broad slightly asymmetric fascia | internally: ghost striae, four large markings, larger in the ventral side | [11] |
| P. microstauron var. angusta Krammer | linear, apices always distinctly offset and much smaller than the valve width, waged-shaped | 25–47 | 6.5–8.0 | 10–12 | narrow, linear | with a broad slightly asymmetric fascia | no date, not visible on LM photo | [11,57,61] |
| P. gibba var. subsancta Manguin | linear-lanceolate with hardly protracted apices | 37.5 | 7.5–8.0 | 13–15 | very broad, lanceolate | rectangular with a fascia | internally: ghost striae | [11,58,59] |
| P. australogibba Van de Vijver, Chattová et Metzeltin | lanceolate to narrowly lanceolate, weakly protracted, rostrate, broadly rounded apices | 45–60 | 7.8–9.4 | 12–13 | moderately broad, lanceolate | large, rhombic–rounded, forming a broad fascia | internally: ghost striae | [55] |
| P. tagliaventiae Lange-Bertalot et Metzeltin | strictly linear, slightly triangulate, apices broadly protracted, rounded to weakly cuneate | 40–70 | 7–10 | 10–11 | broad and widened deltoid towards the central area | with a broad fascia over the valve face and mantle | no date, not visible on LM photo | [62] |
| Pinnularia sp. Tor4r | narrow-linear, slightly concave, apices subcapitate | 42.6 ± 0.4 | 5.8 ± 0.3 | 12.4 ± 0.5 | narrow, linear * | large with a broad slightly asymmetric fascia widening towards the valve margin * | no date, not visible on LM photo | [25] |
| Pinnularia sp. Tor8b | narrow-linear, slightly concave, apices subcapitate | 40 ± 0.5 | 5.9 ± 0.3 | 11.8 ± 0.3 | narrow, linear * | large with a broad slightly asymmetric fascia widening towards the valve margin * | internally: ghost striae * | [25] |
| P. sinistra Krammer | linear, slightly convex or concave, apices indistinctly differentiated, broadly protracted | 17–52 | 4–6.5 | 11–14 | linear, in large individuals lanceolate | slightly asymmetric fascia | no | [11,55,63] |
| P. subcapitata W.Gregory (given as P. hilseana Janisch) | linear to weakly linear-elliptical, apices distinctly offset capitate | 17–57 | 4–6.8 | 10–14 | linear to narrowly lanceolate, expanding into a fascia | fascia | no date, not visible on LM photo | [64] |
| P. saprophila Lange-Bertalot, Kobayasi et Krammer | linear, sides weakly convex, apices distinctly offset, capitate, in small individuals subcapitate or broadly protracted | 21–45 | 5.7–7.5 | 9.5–11 | lanceolate | large, rhombic with a broad fascia | no date, not visible on LM photo | [11] |
| P. pisciculus Ehrenberg | linear, sides straight to very weakly convex or concave to triangulate, apices capitate | 22–50 | 6.0–8.3 | 10.5–12 | narrow or lanceolate | relatively large, rhombic, widened into fascia | no date, not visible on LM photo | [11,65] |
| P. similiformis var. koreana Metzeltin et Krammer | linear, linear-lanceolate to rhombic-lanceolate, apices not offset, obtusely cuneate-rounded | 40–60 | 7.7–8.0 | 10–12 | very narrow, to 1/5 of valve width, linear to slightly lanceolate | rhombic, widened into broad fascia | no date, not visible on LM photo | [11] |
| P. marchica I. Schönfelder | linear to elliptic-lanceolate, in larger specimens slightly concave, apices relatively long rostrate or subcapitate | 22–37 | 4.7–6.3 | 11–14 | narrow, linear | broad rhombic fascia | no date, not visible on LM photo | [11] |
| P. obscura Krasske | linear-elliptical with straight to weakly convex or concave sides, apices weakly rostrate or cuneiform and not offset and broadly rounded | 12–34 | 3–5.4 | 10–13 | very narrow, linear | large, widened into fascia | no | [11,56,66] |
| P. brebissonii var. bicuneata Grunow | linear, sides almost straight and parallel, apices distinctly obtuse or acutely wedge-shaped | 14–60 | 8–11 | 9–13 | narrow, linear | broadly rhombic fascia | no date, not visible on LM photo | [11] |
| P. cavancinii Lange-Bertalot et Metzeltin | rhombic-lanceolate to elliptic-lanceolate, apices gently but distinctly protracted to a wadge, finally obtusely rounded | 32–48 | 7.5–9.0 | 12–13 | lanceolate | fascia, which is rather broad and extended over the mantle | no date, not visible on LM photo | [57] |
| P. stomatophora var. irregularis Krammer | linear, apices obtusely rounded | 40–70 | 10–11 | 12–13 | moderately broad, 1/4–1/3 the breadth of the valve, linear to linear-lanceolate | rectangular with a small fascia | externally, diverse structured flecks on both sides | [11] |
| P. graciloides var. triundulata (Fontell) Krammer | linear, sides slightly undulate, undulates in small valves nearly absent, apices broadly rounded | 82–105 | 11–13 | 10–12 | linear, 1/4–1/3 the breadth of the valve | rhombic, with broad fascia | externally: irregular markings, often difficult to see | [11] |
| P. subgibba var. undulata Krammer | linear, sides weakly undulate, apices slightly capitate | 52–84 | 8–10 | 9–10 | broader, almost 1/4–1/2 breadth of the valve | with broad fascia | internally: ghost striae irregular in the shape on both sides and larger on the ventral side | [11] |
| P. borealis Ehrenberg | linear, linear-elliptical, margins parallel to weakly convex, apices rounded | 24–42 | 8.5–10 | 5–6 | narrow | large, rounded, 1–2 central striae often absent | no | [1,11,28] |
| P. angustiborealis Krammer et Lange-Bertalot | linear, margins moderately convex, apices broadly subrostrate | 34–45 | 7.4–8.0 | 5–6 (7 *) | moderately narrow, widening | transverse fascia | no date, not visible on LM photo | [11] |
| P. dubitabilis Hustedt | rectangular, linear, margins parallel, apices bluntly rounded | 23–40 | 6–7 | 3–5 | wide | absent or fascia | no date, not visible on LM photo | [11] |
| P. intermedia (Lagerstedt) Cleve | linear with straight to weakly convex sides, apices capitate or not, and broadly to obtusely rounded | (15)18–40 | 4.8–7 | 7–10 | very narrow | moderately broad, widened into a fascia | no date, not visible on LM photo | [11] |
| P. angulosa Krammer | rectangular, linear, margins parallel, apices broadly rounded | 42–53 | 9.7–10.3 | 3–4 | wide | nearly absent | no date, not visible on LM photo | [11] |
| Fatty Acid | Pinnularia ministomatophora | Pinnularia vietnamogibba | Pinnularia minigibba | Pinnularia microgibba | Pinnularia insolita | Pinnularia paradubitabilis | ||
|---|---|---|---|---|---|---|---|---|
| VP563 | VP290 | VP294 | VP284 | VP292 | VP289 | VP280 | VP236 | |
| anteiso-15:0 Sarcinic acid | 0.7 ± 0.02 | |||||||
| 10:0 Capric acid | 0.3 ± 0.01 | |||||||
| 12:0 Lauric acid | 0.3 ± 0.01 | 0.2 ± 0.01 | 0.2 ± 0.02 | 0.2 ± 0.01 | 0.4 ± 0.01 | 0.2 ± 0.02 | 0.2 ± 0.01 | |
| 14:0 Myristic acid | 4.6 ± 0.1 | 3.1 ± 0.04 | 4.0 ± 0.1 | 3.0 ± 0.1 | 6.3 ± 0.1 | 2.1 ± 0.1 | 5.1 ± 0.1 | 1.7 ± 0.04 |
| 16:0 Palmitic acid | 30.4 ± 0.8 | 26.7 ± 0.6 | 26.8 ± 0.7 | 24.6 ± 0.7 | 30.3 ± 0.8 | 20.1 ± 0.52 | 23.4 ± 0.7 | 25.4 ± 0.5 |
| 18:0 Stearic acid | 45.7 ± 1.3 | 58.2 ± 1.7 | 51.3 ± 1.7 | 64.4 ± 1.7 | 36.0 ± 0.7 | 61.1 ± 1.9 | 48.9 ± 1.8 | 50.2 ± 1.5 |
| 20:0 Arachidic acid | 0.2 ± 0.01 | 0.3 ± 0.01 | 0.3 ± 0.01 | 0.3 ± 0.01 | 0.1 ± 0.01 | 0.4 ± 0.02 | 0.2 ± 0.01 | 0.2 ± 0.01 |
| 22:0 Behenic acid | 0.1 ± 0.01 | 0.2 ± 0.01 | 0.1 ± 0.01 | |||||
| 16:1n-7 cis-9-Palmitoleic acid | 15.2 ± 0.7 | 9.7 ± 0.8 | 15.5 ± 0.5 | 5.5 ± 0.4 | 20.8 ± 0.6 | 2.7 ± 0.1 | 17.6 ± 0.8 | 15.3 ± 0.6 |
| 16:1n-5 cis-11-Palmitovaccenic acid | 0.2 ± 0.01 | |||||||
| 18:1n-11 cis-7-Vaccenic acid | 1.3 ± 0.1 | |||||||
| 18:1n-9 cis-9-Oleic acid | 1.5 ± 0.04 | 1.7 ± 0.04 | 0.4 ± 0.02 | 1.3 ± 0.04 | 1.6 ± 0.1 | 6.4 ± 0.2 | 1.3 ± 0.02 | 1.9 ± 0.1 |
| 18:1n-7 cis-11-Vaccenic acid | 0.7 ± 0.04 | |||||||
| 16:2n-6 cis-7,10-Hexadecadienoic acid | 0.3 ± 0.02 | |||||||
| 16:2n-4 cis-9,12-Hexadecadienoic acid | 2.4 ± 0.1 | 0.7 ± 0.02 | ||||||
| 18:2n-6 cis-9,12-Linoleic acid | 5.5 ± 0.2 | 0.2 ± 0.02 | 1.1 ± 0.03 | |||||
| 16:3n-4 cis-6,9,12-Hexadecatrienoic acid | 1.1 ± 0.03 | 1.4 ± 0.1 | 0.9 ± 0.03 | |||||
| 18:3n-6 cis-6,9,12-gamma-Linolenic acid | 0.4 ± 0.02 | 0.8 ± 0.02 | ||||||
| 20:4n-6 cis-5,8,11,14-Arachidonic acid | 1.0 ± 0.03 | 0.5 ± 0.02 | 2.3 ± 0.1 | 0.6 ± 0.02 | 0.6 ± 0.02 | |||
| 20:5n-3 cis-5,8,11,14,17-Eicosapentaenoic acid | 1.1 ± 0.03 | |||||||
| total SFAs | 81.2 ± 2.3 | 88.6 ± 2.3 | 82.7 ± 2.5 | 92.7 ± 2.5 | 72.9 ± 1.6 | 84.8 ± 2.4 | 77.8 ± 2.6 | 77.8 ± 2.1 |
| total MUFAs | 16.7 ± 0.8 | 11.4 ± 0.8 | 16.6 ± 0.5 | 6.8 ± 0.4 | 22.4 ± 0.5 | 9.1 ± 0.3 | 18.9 ± 0.8 | 18.7 ± 0.8 |
| total PUFAs | 2.1 ± 0.1 | 0.7 ± 0.03 | 0.5 ± 0.02 | 4.7 ± 0.2 | 6.1 ± 0.2 | 3.3 ± 0.1 | 3.5 ± 0.1 | |
| total fatty acids, mg g−1 dry biomass | 43.9 ± 2.3 | 44.3 ± 2.1 | 42.7 ± 1.9 | 47.8 ± 2.4 | 40.1 ± 1.8 | 39.5 ± 2.2 | 33.7 ± 1.8 | 27.8 ± 1.5 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kezlya, E.; Maltsev, Y.; Genkal, S.; Krivova, Z.; Kulikovskiy, M. Phylogeny and Fatty Acid Profiles of New Pinnularia (Bacillariophyta) Species from Soils of Vietnam. Cells 2022, 11, 2446. https://doi.org/10.3390/cells11152446
Kezlya E, Maltsev Y, Genkal S, Krivova Z, Kulikovskiy M. Phylogeny and Fatty Acid Profiles of New Pinnularia (Bacillariophyta) Species from Soils of Vietnam. Cells. 2022; 11(15):2446. https://doi.org/10.3390/cells11152446
Chicago/Turabian StyleKezlya, Elena, Yevhen Maltsev, Sergei Genkal, Zinaida Krivova, and Maxim Kulikovskiy. 2022. "Phylogeny and Fatty Acid Profiles of New Pinnularia (Bacillariophyta) Species from Soils of Vietnam" Cells 11, no. 15: 2446. https://doi.org/10.3390/cells11152446