
Duplication and Segregation of Centrosomes during Cell Division


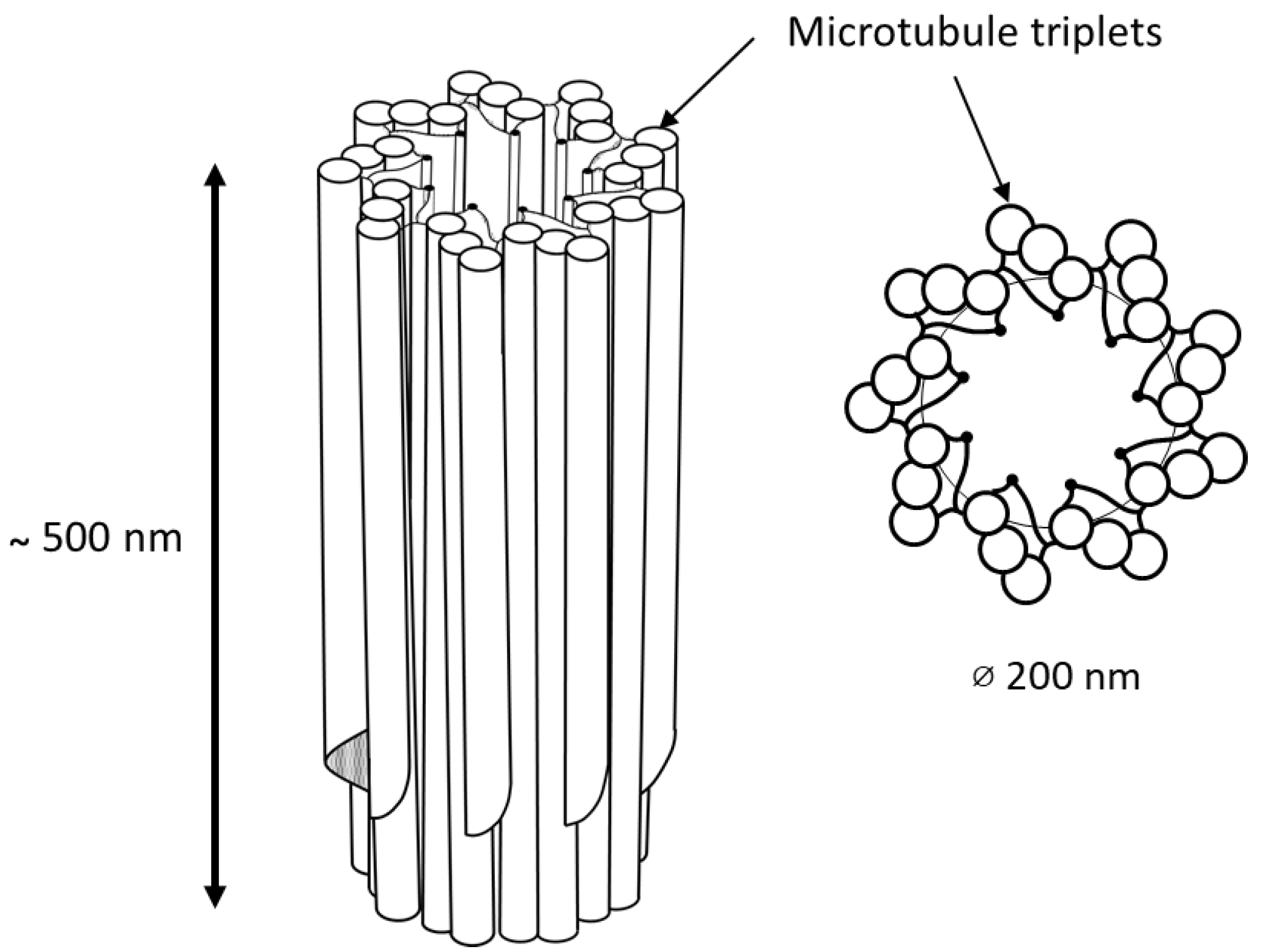{kind=link}
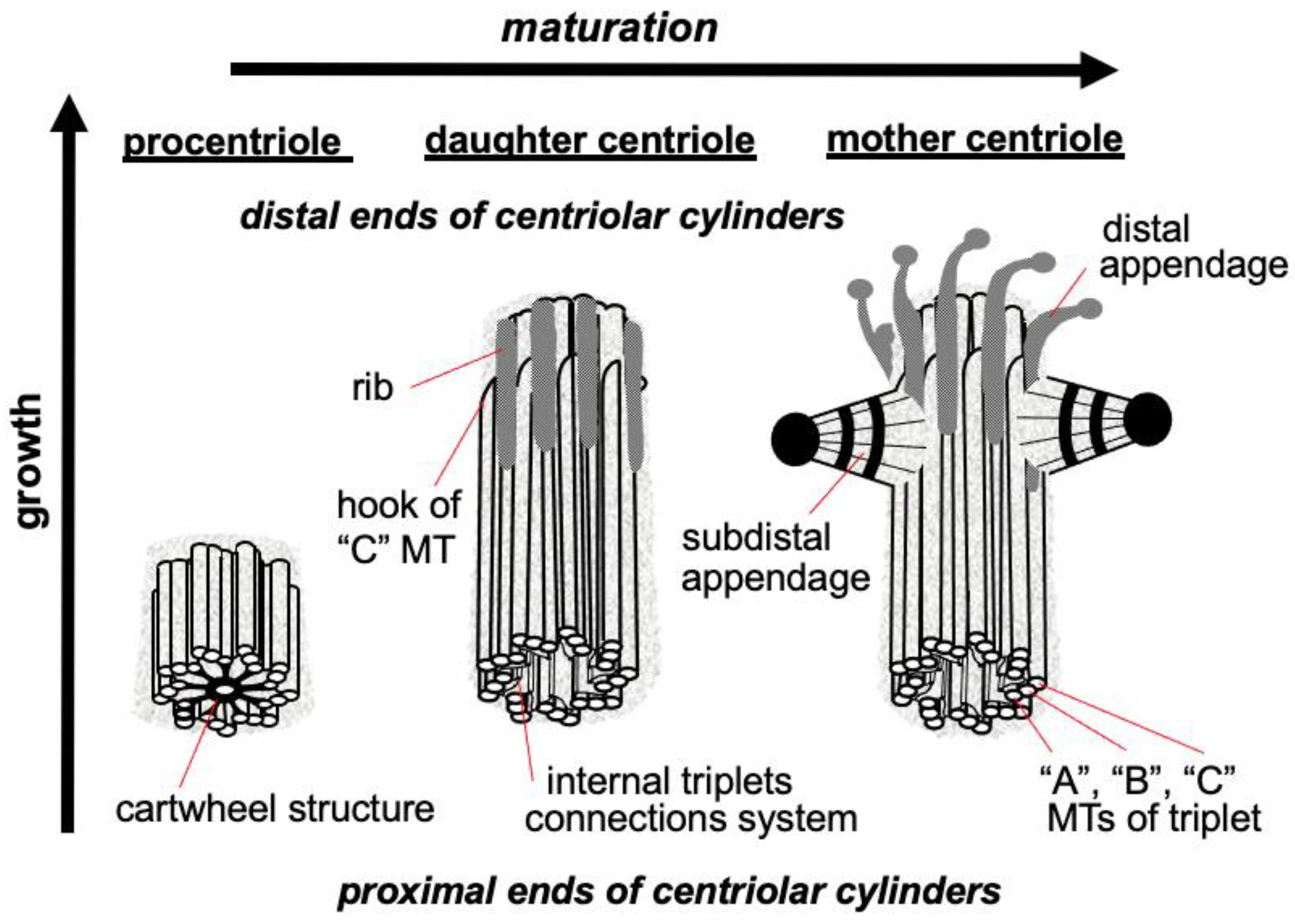{kind=link}
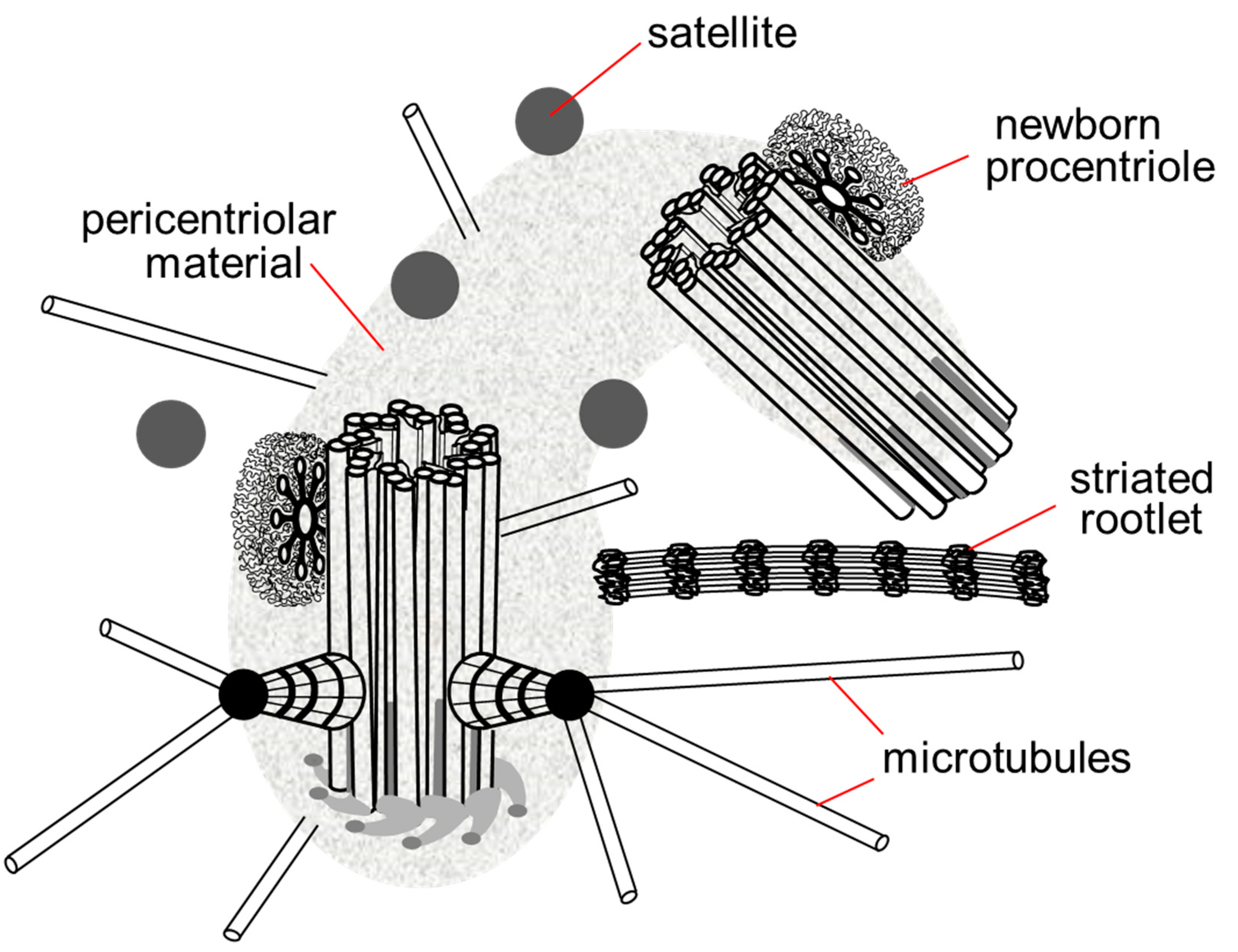{kind=link}
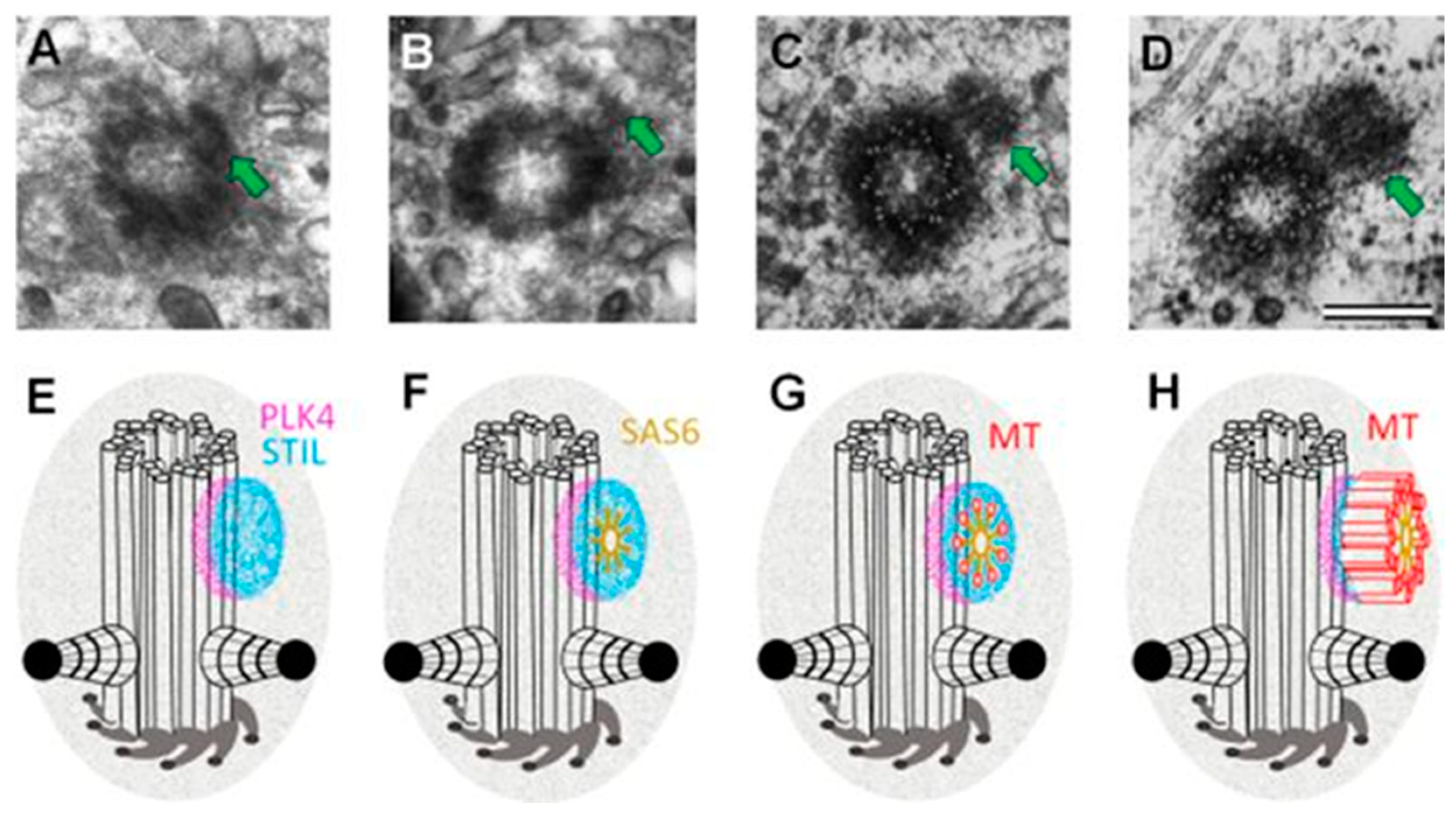{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. What Is the Centrosome?
2. Common Features between the DNA and the Centrosome Cycle
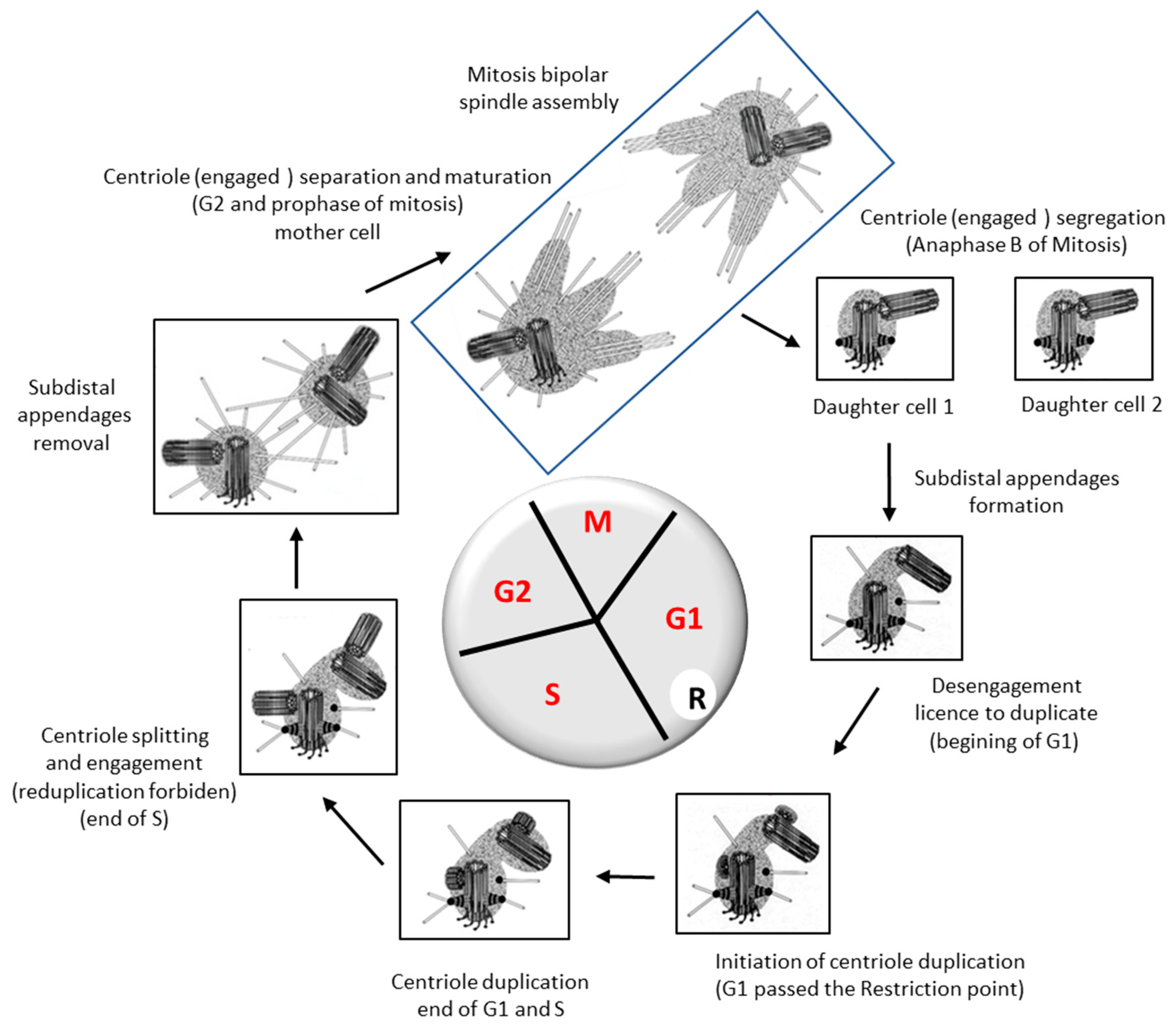
3. Behavior of the Centrosome during a Cell Cycle
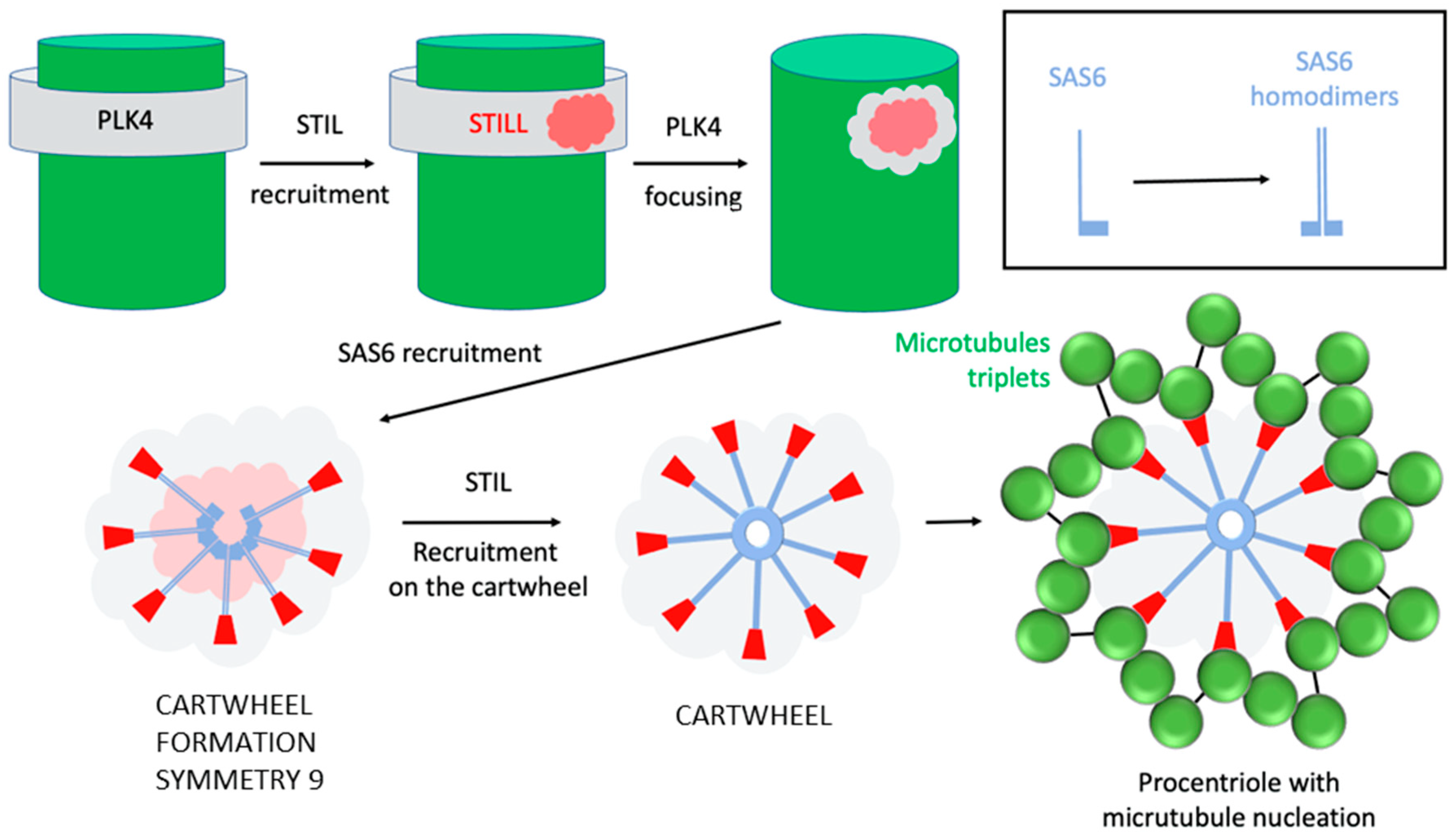
4. One and Only One Procentriole per Centriole per Cell Cycle
What Are the Proteins Involved in This Engagement?
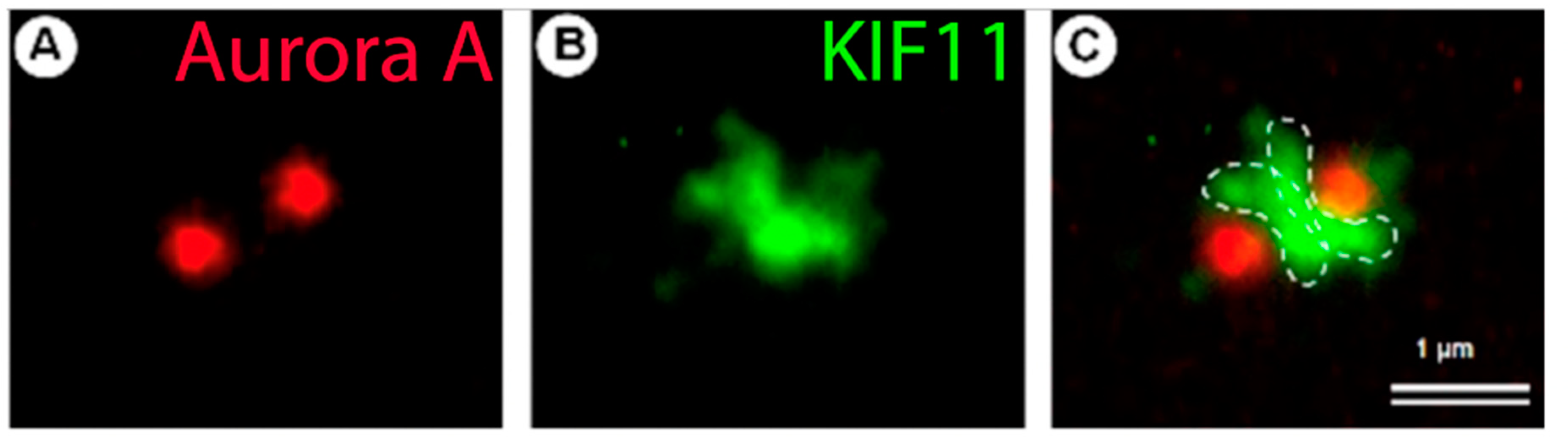
5. Centrosome Splitting, Separation and Segregation
6. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| BICD2 (dynein adaptor) | Bicaudal-D2 (BICD2) is a dynein activating adaptor protein that plays a critical role in microtubule-based minus-end-directed transport |
| CHO cells | Chinese hamster ovary cells |
| G1 phase (from Gap 1) | phase after mitosis before DNA replication |
| G2 phase (from Gap 2) | phase of cell cycle after S pahse and before mitosis |
| CEP68 | Centrosomal Protein 68 kDa |
| CEP120 | Centrosomal Protein 120 kDa |
| CEP135 | Centrosomal Protein 135 kDa |
| Cep152 | Centrosomal Protein 152 kDa |
| Cep192 | Centrosomal Protein 192 kDa |
| CEP215 | Centrosomal Protein 215 kDa (or CDK5RAP2 for Cdk5 regulatory subunit associated protein 2) |
| CNAP1 (Centrosomal Protein 250 kDa–Cep250) | centrosomal Nek2-associated protein 1 |
| CPAP | centriolar protein (centrosomal P4.1-associated protein) |
| EM | electron microscopy |
| LRRC45 | Leucine Rich Repeat Containing 45 kDa |
| Kinesin-like protein KIF11 (or Kinesin 5 or XlEg5) | plus end directed microtubule associated motor. |
| M phase | mitosis |
| MT | microtubule |
| Nek2 | for NIMA-related kinase 2 |
| Nup133 protein | nuclear pore protein 133kDa |
| p150 (Glued) protein | Dynactin subunit, potent anti-catastrophe factor for microtubules |
| PCM | pericentriolar material |
| PLK4 | Polo-like kinase 4 (Serine/threonine-protein kinase) |
| S phase (from synthesis) | phase of cell cycle during which DNA replicated |
| SAS6 | Centriolar Assembly Protein (Spindle Assembly 6 Homolog from Caenorabditis Elegans) |
| STIL protein | Centriolar Assembly Protein, the SCL/TAL interrupting locus is a human oncogene that was originally identified from human T-cell lymphoblastic leukemia, where it functions as a hematopoietic transcription factor. |
References
- Bornens, M. The centrosome in cells and organisms. Science 2012, 335, 422–426. [Google Scholar] [CrossRef] [PubMed]
- Uzbekov, R.; Prigent, C. Clockwise or anticlockwise, turning the centriole triplets in the right direction! FEBS Lett. 2007, 581, 1251–1254. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Uzbekov, R.; Alieva, I. Who are you, subdistal appendages of centriole? Open Biol. 2018, 8, 180062. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tanos, B.E.; Yang, H.J.; Soni, R.; Wang, W.J.; Macaluso, F.P.; Asara, J.M.; Tsou, M.F.B. Centriole distal appendages promote membrane docking, leading to cilia initiation. Genes Dev. 2013, 27, 163–168. [Google Scholar] [CrossRef] [Green Version]
- Uzbekov, R.E.; Avidor-Reiss, T. Principal postulates of centrosomal biology. Cells 2020, 9, 2156. [Google Scholar] [CrossRef]
- Delgehyr, N.; Sillibourne, J.; Bornens, M. Microtubule nucleation and anchoring at the centrosome are independent processes linked by ninein function. J. Cell Sci. 2005, 118 Pt 8, 1565–1575. [Google Scholar] [CrossRef] [Green Version]
- Vorobjev, I.A.; Chentsov, Y.S. Centrioles in the Cell Cycle. I Epithelial Cells. J. Cell Biol. 1982, 98, 938–949. [Google Scholar] [CrossRef]
- Kitagawa, D.; Vakonakis, I.; Olieric, N.; Hilbert, M.; Keller, D.; Olieric, V.; Bortfeld, M.; Erat, M.C.; Fluckiger, I.; Gonczy, P.; et al. Structural basis of the 9-fold symmetry of centrioles. Cell 2011, 144, 364–375. [Google Scholar] [CrossRef] [Green Version]
- van Breugel, M.; Hirono, M.; Andreeva, A.; Yanagisawa, H.A.; Yamaguchi, S.; Nakazawa, Y.; Morgner, N.; Petrovich, M.; Ebong, I.O.; Robinson, C.V.; et al. Structures of sas-6 suggest its organization in centrioles. Science 2011, 331, 1196–1199. [Google Scholar] [CrossRef] [Green Version]
- Osborn, M.; Weber, K. Cytoplasmic microtubules in tissue culture cells appear to grow from an organizing structure towards the plasma membrane. Proc. Natl. Acad. Sci. USA 1976, 73, 867–871. [Google Scholar] [CrossRef] [Green Version]
- Diffley, J.F. Once and only once upon a time: Specifying and regulating origins of DNA replication in eukaryotic cells. Genes Dev. 1996, 10, 2819–2830. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zitouni, S.; Francia, M.E.; Leal, F.; Montenegro Gouveia, S.; Nabais, C.; Duarte, P.; Gilberto, S.; Brito, D.; Moyer, T.; Kandels-Lewis, S.; et al. CDK1 Prevents Unscheduled PLK4-STIL Complex Assembly in Centriole Biogenesis. Curr. Biol. 2016, 26, 1127–1137. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Watson, J.D.; Crick, F.H. Genetical implications of the structure of deoxyribonucleic acid. Nature 1953, 171, 964–967. [Google Scholar] [CrossRef] [PubMed]
- Meselson, M.; Stahl, F.W. The replication of DNA in Escherichia coli. Proc. Natl. Acad. Sci. USA 1958, 44, 671–682. [Google Scholar] [CrossRef] [Green Version]
- Gönczy, P.; Hatzopoulos, G.N. Centriole assembly at a glance. J. Cell Sci. 2019, 132, jcs228833. [Google Scholar] [CrossRef] [Green Version]
- Dirksen, E.R. The presence of centrioles in artificially activated sea urchin eggs. J. Biophys. Biochem. Cytol. 1961, 11, 244–247. [Google Scholar] [CrossRef] [Green Version]
- Szöllösi, D.; Calarco, P.; Donahue, R.P. Absence of centrioles in the first and second meiotic spindles of mouse oocytes. J. Cell Sci. 1972, 11, 521–541. [Google Scholar] [CrossRef]
- Uzbekov, R.E. Centriole duplication in PE (SPEV) cells starts before beginning of DNA replication. Biochem. (Mosc.) Suppl. Ser. A Membr. Cell Biol. 2007, 1, 206–211. [Google Scholar] [CrossRef]
- Rattner, J.B.; Phillips, S.G. Independence of centriole formation and DNA synthesis. J. Cell Biol. 1973, 57, 359–372. [Google Scholar] [CrossRef] [Green Version]
- Sluder, G. Centrosomes and the cell cycle. J. Cell Sci. Suppl. 1989, 12, 253–275. [Google Scholar] [CrossRef] [Green Version]
- Balczon, R.; Bao, L.; Zimmer, W.E.; Brown, K.; Zinkowski, R.P.; Brinkley, B.R. Dissociation of centrosome replication events from cycles of DNA synthesis and mitotic division in hydroxyurea-arrested Chinese hamster ovary cells. J. Cell Biol. 1995, 130, 105–115. [Google Scholar] [CrossRef] [PubMed]
- O’Connell, K.F.; Caron, C.; Kopish, K.R.; Hurd, D.D.; Kemphues, K.J.; Li, Y.; White, J.G. The C. elegans zyg-1 gene encodes a regulator of centrosome duplication with distinct maternal and paternal roles in the embryo. Cell 2001, 105, 547–558. [Google Scholar] [CrossRef] [Green Version]
- Habedanck, R.; Stierhof, Y.D.; Wilkinson, C.J.; Nigg, E.A. The Polo kinase Plk4 functions in centriole duplication. Nat. Cell Biol. 2005, 7, 1140–1146. [Google Scholar] [CrossRef] [PubMed]
- Bettencourt-Dias, M.; Rodrigues-Martins, A.; Carpenter, L.; Riparbelli, M.; Lehmann, L.; Gatt, M.K.; Carmo, N.; Balloux, F.; Callaini, G.; Glover, D.M. SAK/PLK4 is required for centriole duplication and flagella development. Curr. Biol. 2005, 15, 2199–2207. [Google Scholar] [CrossRef] [Green Version]
- Kim, T.S.; Park, J.E.; Shukla, A.; Choi, S.; Murugan, R.N.; Lee, J.H.; Ahn, M.; Rhee, K.; Bang, J.K.; Kim, B.Y.; et al. Hierarchical recruitment of Plk4 and regulation of centriole biogenesis by two centrosomal scaffolds, Cep192 and Cep152. Proc. Natl. Acad. Sci. USA 2013, 110, E4849–E4857. [Google Scholar] [CrossRef] [Green Version]
- Kemp, C.A.; Kopish, K.R.; Zipperlen, P.; Ahringer, J.; O'Connell, K.F. Centrosome maturation and duplication in C. elegans require the coiled-coil protein SPD-2. Dev. Cell 2004, 6, 511–523. [Google Scholar] [CrossRef] [Green Version]
- Pelletier, L.; Ozlü, N.; Hannak, E.; Cowan, C.; Habermann, B.; Ruer, M.; Müller-Reichert, T.; Hyman, A.A. The Caenorhabditis elegans centrosomal protein SPD-2 is required for both pericentriolar material recruitment and centriole duplication. Curr. Biol. 2004, 14, 863–873. [Google Scholar] [CrossRef] [Green Version]
- Delattre, M.; Leidel, S.; Wani, K.; Baumer, K.; Bamat, J.; Schnabel, H.; Feichtinger, R.; Schnabel, R.; Gönczy, P. Centriolar SAS-5 is required for centrosome duplication in C. elegans. Nat. Cell Biol. 2004, 6, 656–664. [Google Scholar] [CrossRef]
- Lettman, M.M.; Wong, Y.L.; Viscardi, V.; Niessen, S.; Chen, S.H.; Shiau, A.K.; Zhou, H.; Desai, A.; Oegema, K. Direct binding of SAS-6 to ZYG-1 recruits SAS-6 to the mother centriole for cartwheel assembly. Dev. Cell 2013, 25, 284–298. [Google Scholar] [CrossRef] [Green Version]
- Ohta, M.; Ashikawa, T.; Nozaki, Y.; Kozuka-Hata, H.; Goto, H.; Inagaki, M.; Oyama, M.; Kitagawa, D. Direct interaction of Plk4 with STIL ensures formation of a single procentriole per parental centriole. Nat. Commun. 2014, 5, 5267. [Google Scholar] [CrossRef] [Green Version]
- Dzhindzhev, N.S.; Tzolovsky, G.; Lipinszki, Z.; Schneider, S.; Lattao, R.; Fu, J.; Debski, J.; Dadlez, M.; Glover, D.M. Plk4 phosphorylates Ana2 to trigger Sas6 recruitment and procentriole formation. Curr. Biol. 2014, 24, 2526–2532. [Google Scholar] [CrossRef] [Green Version]
- Moyer, T.C.; Clutario, K.M.; Lambrus, B.G.; Daggubati, V.; Holland, A.J. Binding of STIL to Plk4 activates kinase activity to promote centriole assembly. J. Cell Biol. 2015, 209, 863–878. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kratz, A.-S.; Bärenz, F.; Richter, K.T.; Hoffmann, I. Plk4-dependent phosphorylation of STIL is required for centriole duplication. Biol. Open 2015, 4, 370–377. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Uzbekov, R.E.; Alieva, I.B. Centriole duplication or DNA replication—what starts earlier. In Cytoskeleton: Cell Movement, Cytokinesis and Organelles Organization (Editors S. Lansing and and T. Rousseau); Chapter 5; Nosva Science Publishers, Inc.: New York, NY, USA, 2010. [Google Scholar]
- Uzbekov, R.E.; Alieva, I.B. Centrosome–history of investigations and new discoveries. In From Cytoplasmic Granule to the Centre of Intracellular Regulation; Nadezhdina, E.S., Ed.; Moscow University Press: Moscow, Russia, 2013; p. 320. [Google Scholar]
- Kim, M.; O’Rourke, B.P.; Soni, R.K.; Jallepalli, P.V.; Hendrickson, R.C.; Tsou, M.-F.B. Promotion and Suppression of Centriole Duplication Are Catalytically Coupled through PLK4 to Ensure Centriole Homeostasis. Cell Rep. 2016, 16, 1195–1203. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sakharov, V.N.; Voronkova, L.N. The kinetics of the transition to DNA synthesis in the cell cycle of sister cells of the ESK line in culture. Tsitologiia 1993, 35, 79–85. [Google Scholar]
- Gibbons, I.R.; Grimstone, A.V. On flagellar structure in certain flagellates. J. Biophys. Biochem. Cytol. 1960, 7, 697–716. [Google Scholar] [CrossRef] [Green Version]
- Leidel, S.; Delattre, M.; Cerutti, L.; Baumer, K.; Gönczy, P. SAS-6 defines a protein family required for centrosome duplication in C. elegans and in human cells. Nat. Cell Biol. 2005, 7, 115–125. [Google Scholar] [CrossRef]
- Mahjoub, M.R.; Xie, Z.; Stearns, T. Cep120 is asymmetrically localized to the daughter centriole and is essential for centriole assembly. J. Cell Biol. 2010, 191, 331–346. [Google Scholar] [CrossRef] [Green Version]
- Lin, Y.N.; Wu, C.T.; Lin, Y.C.; Hsu, W.B.; Tang, C.J.; Chang, C.W.; Tang, T.K. CEP120 interacts with CPAP and positively regulates centriole elongation. J. Cell Biol. 2013, 202, 211–219. [Google Scholar] [CrossRef] [Green Version]
- Comartin, D.; Gupta, G.D.; Fussner, E.; Coyaud, É.; Hasegan, M.; Archinti, M.; Cheung, S.W.; Pinchev, D.; Lawo, S.; Raught, B.; et al. CEP120 and SPICE1 cooperate with CPAP in centriole elongation. Curr. Biol. 2013, 23, 1360–1366. [Google Scholar] [CrossRef] [Green Version]
- Leidel, S.; Gönczy, P. SAS-4 is essential for centrosome duplication in C elegans and is recruited to daughter centrioles once per cell cycle. Dev. Cell 2003, 4, 431–439. [Google Scholar] [CrossRef] [Green Version]
- Dammermann, A.; Müller-Reichert, T.; Pelletier, L.; Habermann, B.; Desai, A.; Oegema, K. Centriole assembly requires both centriolar and pericentriolar material proteins. Dev. Cell 2004, 7, 815–829. [Google Scholar] [CrossRef] [Green Version]
- Kirkham, M.; Müller-Reichert, T.; Oegema, K.; Grill, S.; Hyman, A.A. SAS-4 is a C. elegans centriolar protein that controls centrosome size. Cell 2003, 112, 575–587. [Google Scholar] [CrossRef] [Green Version]
- Lin, Y.C.; Chang, C.W.; Hsu, W.B.; Tang, C.J.; Lin, Y.N.; Chou, E.J.; Wu, C.T.; Tang, T.K. Human microcephaly protein CEP135 binds to hSAS-6 and CPAP, and is required for centriole assembly. EMBO J. 2013, 32, 1141–1154. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tang, C.J.; Lin, S.Y.; Hsu, W.B.; Lin, Y.N.; Wu, C.T.; Lin, Y.C.; Chang, C.W.; Wu, K.S.; Tang, T.K. The human microcephaly protein STIL interacts with CPAP and is required for procentriole formation. EMBO J. 2011, 30, 4790–4804. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nigg, E.A.; Holland, A.J. Once and only once: Mechanisms of centriole duplication and their deregulation in disease. Nat. Rev. Mol. Cell Biol. 2018, 19, 297–312. [Google Scholar] [CrossRef]
- Leda, M.; Holland, A.J.; Goryachev, A.B. Autoamplification and Competition Drive Symmetry Breaking: Initiation of Centriole Duplication by the PLK4-STIL Network. iScience 2018, 8, 222–235. [Google Scholar] [CrossRef] [Green Version]
- Yamamoto, S.; Kitagawa, D. Self-organization of Plk4 regulates symmetry breaking in centriole duplication. Nat. Commun. 2019, 10, 1810. [Google Scholar] [CrossRef] [Green Version]
- Cunningham, N.H.J.; Bouhlel, I.B.; Conduit, P.T. Daughter centrioles assemble preferentially towards the nuclear envelope in Drosophila syncytial embryos. Open Biol. 2022, 12, 210343. [Google Scholar] [CrossRef]
- Nadezhdina, E.S.; Fais, D.; Chentsov, Y.S. Partial purification of centrioles from spleen cells. Cell Biol. Int. Rep. 1978, 2, 601–606. [Google Scholar] [CrossRef]
- Barrera, J.A.; Kao, L.R.; Hammer, R.E.; Seemann, J.; Fuchs, J.L.; Megraw, T.L. CDK5RAP2 regulates centriole engagement and cohesion in mice. Dev. Cell 2010, 18, 913–926. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vidwans, S.J.; Wong, M.L.; O'Farrell, P.H. Mitotic regulators govern progress through steps in the centrosome duplication cycle. J. Cell Biol. 1999, 147, 1371–1378. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Uhlmann, F.; Lottspeich, F.; Nasmyth, K. Sister-chromatid separation at anaphase onset is promoted by cleavage of the cohesin subunit Scc1. Nature 1999, 400, 37–42. [Google Scholar] [CrossRef] [PubMed]
- Uhlmann, F.; Wernic, D.; Poupart, M.A.; Koonin, E.V.; Nasmyth, K. Cleavage of cohesin by the CD clan protease separin triggers anaphase in yeast. Cell 2000, 103, 375–386. [Google Scholar] [CrossRef] [Green Version]
- Tsou, M.F.; Stearns, T. Mechanism limiting centrosome duplication to once per cell cycle. Nature 2006, 442, 947–951. [Google Scholar] [CrossRef] [PubMed]
- Ciosk, R.; Zachariae, W.; Michaelis, C.; Shevchenko, A.; Mann, M.; Nasmyth, K. An ESP1/PDS1 complex regulates loss of sister chromatid cohesion at the metaphase to anaphase transition in yeast. Cell 1998, 93, 1067–1076. [Google Scholar] [CrossRef] [Green Version]
- Matsuo, K.; Ohsumi, K.; Iwabuchi, M.; Kawamata, T.; Ono, Y.; Takahashi, M. Kendrin is a novel substrate for separase involved in the licensing of centriole duplication. Curr. Biol. 2012, 22, 915–921. [Google Scholar] [CrossRef] [Green Version]
- Pagan, J.K.; Marzio, A.; Jones, M.J.; Saraf, A.; Jallepalli, P.V.; Florens, L.; Washburn, M.P.; Pagano, M. Degradation of Cep68 and PCNT cleavage mediate Cep215 removal from the PCM to allow centriole separation, disengagement and licensing. Nat. Cell Biol. 2015, 17, 31–43. [Google Scholar] [CrossRef] [Green Version]
- Fry, A.M.; Mayor, T.; Meraldi, P.; Stierhof, Y.D.; Tanaka, K.; Nigg, E.A. C-Nap1, a novel centrosomal coiled-coil protein and candidate substrate of the cell cycle-regulated protein kinase Nek2. J. Cell Biol. 1998, 141, 1563–1574. [Google Scholar] [CrossRef] [Green Version]
- Mayor, T.; Hacker, U.; Stierhof, Y.D.; Nigg, E.A. The mechanism regulating the dissociation of the centrosomal protein C-Nap1 from mitotic spindle poles. J. Cell Sci. 2002, 115 Pt 16, 3275–3284. [Google Scholar] [CrossRef]
- Bahe, S.; Stierhof, Y.D.; Wilkinson, C.J.; Leiss, F.; Nigg, E.A. Rootletin forms centriole-associated filaments and functions in centrosome cohesion. J. Cell Biol. 2005, 171, 27–33. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, J.; Adamian, M.; Li, T. Rootletin interacts with C-Nap1 and may function as a physical linker between the pair of centrioles/basal bodies in cells. Mol. Biol. Cell 2006, 17, 1033–1040. [Google Scholar] [CrossRef] [PubMed]
- Graser, S.; Stierhof, Y.D.; Nigg, E.A. Cep68 and Cep215 (Cdk5rap2) are required for centrosome cohesion. J. Cell Sci. 2007, 120 Pt 24, 4321–4331. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hadjihannas, M.V.; Brückner, M.; Behrens, J. Conductin/axin2 and Wnt signalling regulates centrosome cohesion. EMBO Rep. 2010, 11, 317–324. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- He, R.; Huang, N.; Bao, Y.; Zhou, H.; Teng, J.; Chen, J. LRRC45 is a centrosome linker component required for centrosome cohesion. Cell Rep. 2013, 4, 1100–1107. [Google Scholar] [CrossRef] [Green Version]
- Fang, G.; Zhang, D.; Yin, H.; Zheng, L.; Bi, X.; Yuan, L. Centlein mediates an interaction between C-Nap1 and Cep68 to maintain centrosome cohesion. J. Cell Sci. 2014, 127 Pt 8, 1631–1639. [Google Scholar] [CrossRef] [Green Version]
- Uzbekov, R.; Kireyev, I.; Prigent, C. Centrosome separation: Respective role of microtubules and actin filaments. Biol. Cell 2002, 94, 275–288. [Google Scholar] [CrossRef]
- Blangy, A.; Lane, H.A.; d’Hérin, P.; Harper, M.; Kress, M.; Nigg, E.A. Phosphorylation by p34cdc2 regulates spindle association of human Eg5, a kinesin-related motor essential for bipolar spindle formation in vivo. Cell 1995, 83, 1159–1169. [Google Scholar] [CrossRef] [Green Version]
- Mayer, T.U.; Kapoor, T.M.; Haggarty, S.J.; King, R.W.; Schreiber, S.L.; Mitchison, T.J. Small molecule inhibitor of mitotic spindle bipolarity identified in a phenotype-based screen. Science 1999, 286, 971–974. [Google Scholar] [CrossRef] [Green Version]
- Kashina, A.S.; Baskin, R.J.; Cole, D.G.; Wedaman, K.P.; Saxton, W.M.; Scholey, J.M. A bipolar kinesin. Nature 1996, 379, 270–272. [Google Scholar] [CrossRef]
- Kapitein, L.C.; Peterman, E.J.; Kwok, B.H.; Kim, J.H.; Kapoor, T.M.; Schmidt, C.F. The bipolar mitotic kinesin Eg5 moves on both microtubules that it crosslinks. Nature 2005, 435, 114–118. [Google Scholar] [CrossRef] [PubMed]
- Guo, Y.; Zheng, Y. Lamins position the nuclear pores and centrosomes by modulating dynein. Mol. Biol. Cell 2015, 26, 3379–3389. [Google Scholar] [CrossRef] [PubMed]
- Bolhy, S.; Bouhlel, I.; Dultz, E.; Nayak, T.; Zuccolo, M.; Gatti, X.; Vallee, R.; Ellenberg, J.; Doye, V. A Nup133-dependent NPC-anchored network tethers centrosomes to the nuclear envelope in prophase. J. Cell Biol. 2011, 192, 855–871. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Giet, R.; Uzbekov, R.; Cubizolles, F.; Le Guellec, K.; Prigent, C. The Xenopus laevis aurora-related protein kinase pEg2 associates with and phosphorylates the kinesin-related protein XlEg5. J. Biol. Chem. 1999, 274, 15005–15013. [Google Scholar] [CrossRef] [Green Version]
- Paintrand, M.; Moudjou, M.; Delacroix, H.; Bornens, M. Centrosome organization and centriole architecture: Their sensitivity to divalent cations. J. Struct. Biol. 1992, 108, 107–128. [Google Scholar] [CrossRef]
- Ibrahim, R.; Messaoudi, C.; Chichon, F.J.; Celati, C.; Marco, S. Electron tomography study of isolated human centrioles. Microsc. Res. Tech. 2009, 72, 42–48. [Google Scholar] [CrossRef]
- Chong, W.M.; Wang, W.J.; Lo, C.H.; Chiu, T.Y.; Chang, T.J.; Liu, Y.P.; Tanos, B.; Mazo, G.; Tsou, M.B.; Jane, W.N.; et al. Super-resolution microscopy reveals coupling between mammalian centriole subdistal appendages and distal appendages. Elife 2020, 9, e53580. [Google Scholar] [CrossRef] [Green Version]
- Chen, T.Y.; Syu, J.S.; Han, T.Y.; Cheng, H.L.; Lu, F.I.; Wang, C.Y. Cell Cycle-Dependent Localization of Dynactin Subunit p150 glued at Centrosome. J. Cell Biochem. 2015, 116, 2049–2060. [Google Scholar] [CrossRef]
- Kodani, A.; Salomé Sirerol-Piquer, M.; Seol, A.; Garcia-Verdugo, J.M.; Reiter, J.F. Kif3a interacts with Dynactin subunit p150 Glued to organize centriole subdistal appendages. EMBO J. 2013, 32, 597–607. [Google Scholar] [CrossRef] [Green Version]
- Vitrat, N.; Cohen-Solal, K.; Pique, C.; Le Couedic, J.P.; Norol, F.; Larsen, A.K.; Katz, A.; Vainchenker, W.; Debili, N. Endomitosis of human megakaryocytes are due to abortive mitosis. Blood 1998, 91, 3711–3723. [Google Scholar] [CrossRef]
- Anastasi, J. Some observations on the geometry of megakaryocyte mitotic figures: Buckyballs in the bone marrow. Blood 2011, 118, 6473–6474. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nigg, E.A. Centrosome aberrations: Cause or consequence of cancer progression? Nat. Rev. Cancer 2002, 2, 815–825. [Google Scholar] [CrossRef] [PubMed]
- Boveri, T. Concerning the origin of malignant tumours by Theodor Boveri. Translated and annotated by Henry Harris. J. Cell Sci. 2008, 121 (Suppl. S1), 1–84. [Google Scholar] [CrossRef] [PubMed]
- Hardy, P.A.; Zacharias, H. Reappraisal of the Hansemann-Boveri hypothesis on the origin of tumors. Cell Biol. Int. 2005, 29, 983–992. [Google Scholar] [CrossRef]
- Kalkan, B.M.; Ozcan, S.C.; Quintyne, N.J.; Reed, S.L.; Acilan, C. Keep Calm and Carry on with Extra Centrosomes. Cancers 2022, 14, 442. [Google Scholar] [CrossRef] [PubMed]
- Fan, G.; Sun, L.; Meng, L.; Hu, C.; Wang, X.; Shi, Z.; Hu, C.; Han, Y.; Yang, Q.; Cao, L.; et al. The ATM and ATR kinases regulate centrosome clustering and tumor recurrence by targeting KIFC1 phosphorylation. Nat. Commun. 2021, 12, 20. [Google Scholar] [CrossRef]
- Khodjakov, A.; Cole, R.W.; Oakley, B.R.; Rieder, C.L. Centrosome-independent mitotic spindle formation in vertebrates. Curr. Biol. 2000, 10, 59–67. [Google Scholar] [CrossRef] [Green Version]
- Basto, R.; Lau, J.; Vinogradova, T.; Gardiol, A.; Woods, C.G.; Khodjakov, A.; Raff, J.W. Flies without centrioles. Cell 2006, 125, 1375–1386. [Google Scholar] [CrossRef] [Green Version]
- Takumi, K.; Kitagawa, D. Experimental and Natural Induction of de novo Centriole Formation. Front. Cell Dev. Biol. 2022, 10, 861864. [Google Scholar] [CrossRef]
- Flemming, W. Studien in der Entwicklungsgeschichte der Najaden. Sitz. Akad. Wissensch Wien. 1875, 71, 81–147. (In German) [Google Scholar]
- Hertwig, O. Beitrage zur Kenntniss der Bildung, Befruchtung und Theilung des thierischen Eies. Morphol. Jb. 1875, 1, 347–434. (In German) [Google Scholar] [CrossRef]
- Van Beneden, E. Recherches sur les Dicyémides, survivants actuels d’un embranchement des mésozoaires. Bull. Acad. Roy. Méd. Belg. 1876, 41, 1160–1205. (In French) [Google Scholar]
- Bornens, M. Centrosome organization and functions. Curr. Opin. Struct. Biol. 2021, 66, 199–206. [Google Scholar] [CrossRef] [PubMed]







Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Prigent, C.; Uzbekov, R. Duplication and Segregation of Centrosomes during Cell Division. Cells 2022, 11, 2445. https://doi.org/10.3390/cells11152445
Prigent C, Uzbekov R. Duplication and Segregation of Centrosomes during Cell Division. Cells. 2022; 11(15):2445. https://doi.org/10.3390/cells11152445
Chicago/Turabian StylePrigent, Claude, and Rustem Uzbekov. 2022. "Duplication and Segregation of Centrosomes during Cell Division" Cells 11, no. 15: 2445. https://doi.org/10.3390/cells11152445