
The Gluopsins: Opsins without the Retinal Binding Lysine


{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Material and Methods
2.1. Protein Sequence Collection
2.2. Sequence Pruning
2.3. Rogue and Long Branch Removal
2.4. Phylogenetic Reconstruction
2.5. Tree Visualization
2.6. Finding Position 2967.43 in Other Opsins and GPCRs
2.7. Annotate the Sequences with Higher Taxa
2.8. Sequence Logo
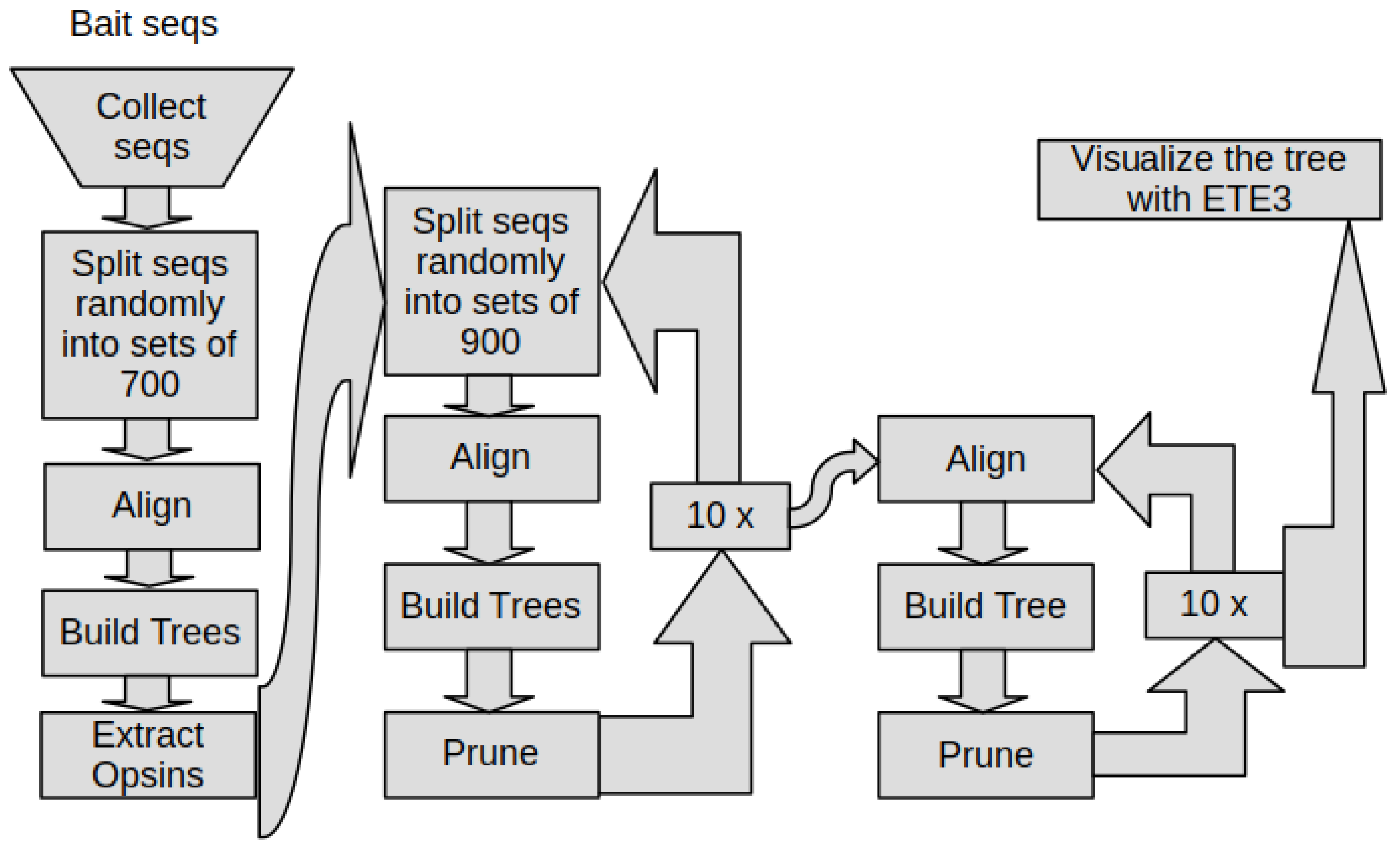
2.9. The Phylogeny Pipeline
2.10. Opsin Nomenclature
3. Results
3.1. The Phylogeny Pipeline
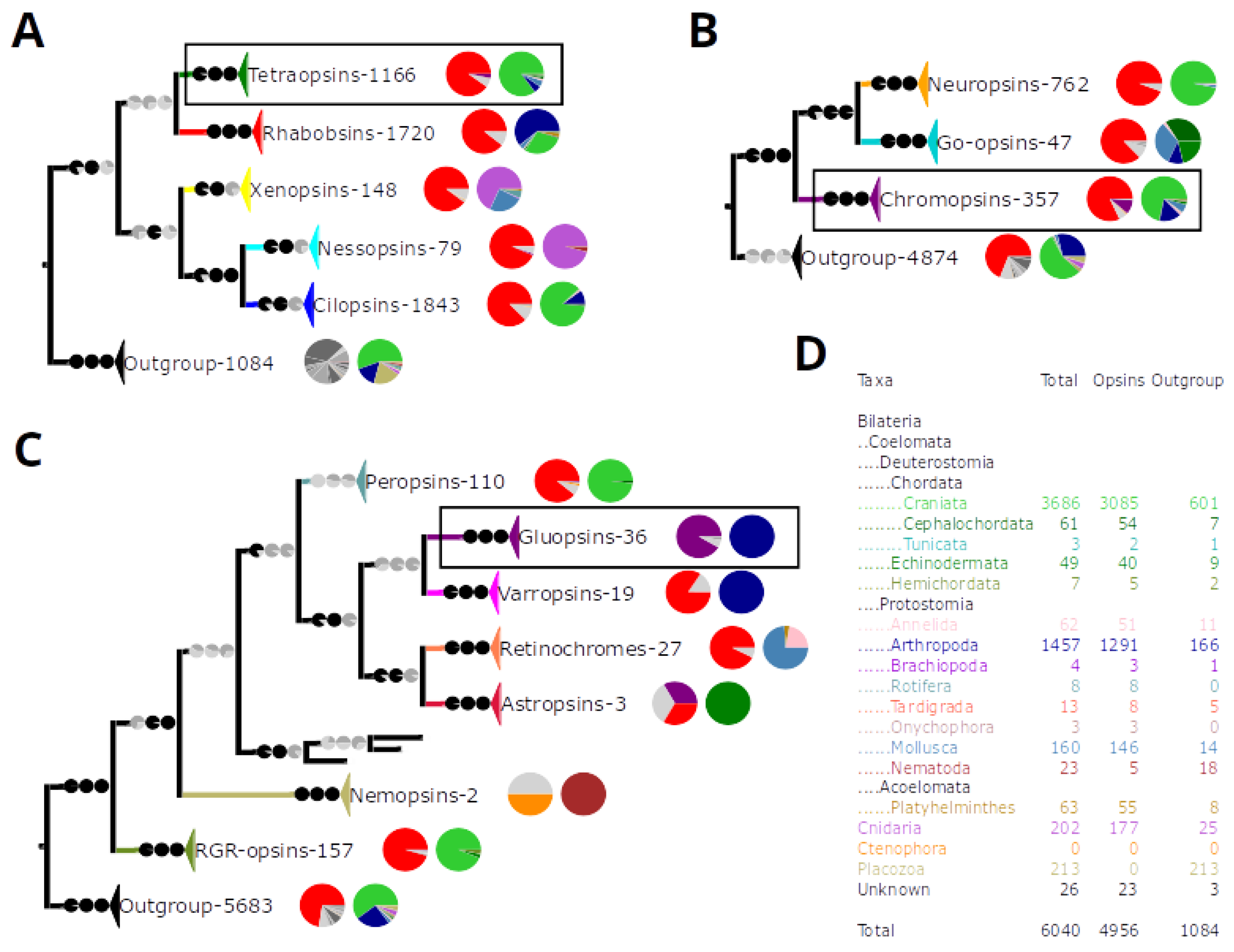
3.2. Five Basal Types of Opsins
3.3. The Chromopsins
3.4. The Nemopsins Have Arginine at Position 296
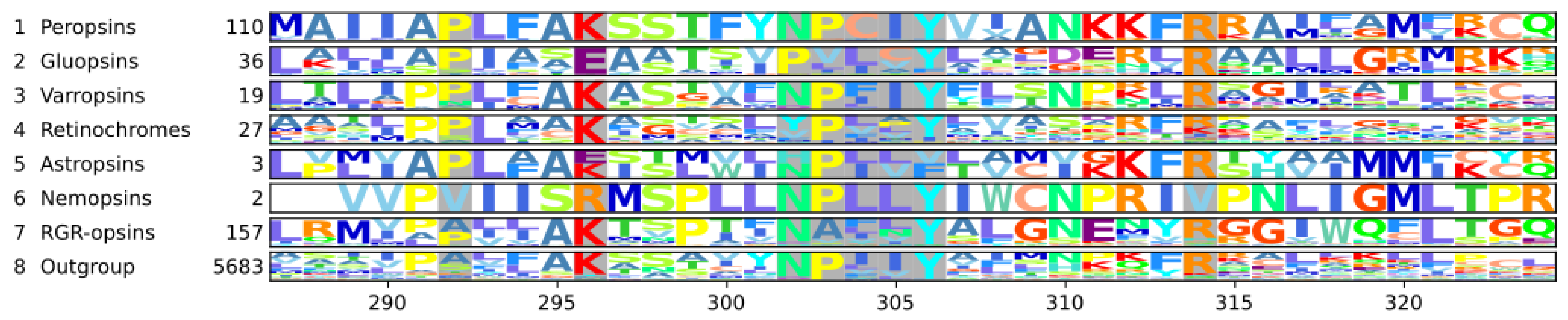
3.5. The Gluopsins Have Glutamic Acid at Position 296
3.6. The NPxxY7.53 Motif Is Derived in Some Chromopsins
4. Discussion
4.1. The Number of Chromopsins in the Urbilaterian
4.2. The Function of the Chromopsins
4.3. Photoisomerases
4.4. The Varropsins
4.5. The Peropsins
4.6. The RGR-Opsins
4.7. The Retinochromes
4.8. The Gluopsins: Opsins without Lysine 2967.43
5. Summary
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Casey, P.J.; Gilman, A.G. G protein involvement in receptor-effector coupling. J. Biol. Chem. 1988, 263, 2577–2580. [Google Scholar] [CrossRef]
- Attwood, T.K.; Findlay, J.B.C. Fingerprinting G-protein-coupled receptors. Protein Eng. Des. Sel. 1994, 7, 195–203. [Google Scholar] [CrossRef] [PubMed]
- Dixon, R.A.F.; Kobilka, B.K.; Strader, D.J.; Benovic, J.L.; Dohlman, H.G.; Frielle, T.; Bolanowski, M.A.; Bennett, C.D.; Rands, E.; Diehl, R.E.; et al. Cloning of the gene and cDNA for mammalian β-adrenergic receptor and homology with rhodopsin. Nature 1986, 321, 75–79. [Google Scholar] [CrossRef] [PubMed]
- Dixon, R.A.F.; Sigal, I.S.; Rands, E.; Register, R.B.; Candelore, M.R.; Blake, A.D.; Strader, C.D. Ligand binding to the β-adrenergic receptor involves its rhodopsin-like core. Nature 1987, 326, 73–77. [Google Scholar] [CrossRef]
- Wald, G. Carotenoids and the Vitamin A Cycle in Vision. Nature 1934, 134, 65. [Google Scholar] [CrossRef]
- Wald, G.; Brown, P.K.; Hubbard, R.; Oroshnik, W.; Brown, P.K.; Habburd, R.; Wald, G.; Hubbard, R.; Wald, G.; Brown, P.K.; et al. Hindered Cis Isomers of Vitamin A and Retinene: The Structure of the Neo-B Isomer. Proc. Natl. Acad. Sci. USA 1955, 41, 438–451. [Google Scholar] [CrossRef] [Green Version]
- Brown, P.; Wald, G. The neo-b isomer of vitamin A and retinene. J. Biol. Chem. 1956, 222, 865–877. [Google Scholar] [CrossRef]
- Oroshnik, W. The synthesis and configuration of neo-b vitamin A and neoretinene b. J. Am. Chem. Soc. 1956, 78, 2651–2652. [Google Scholar] [CrossRef]
- Oroshnik, W.; Brown, P.K.; Hubbard, R.; Wald, G. Hindered cis isomers of vitamin A and retinene: The structure of the neo-b isomer. Proc. Natl. Acad. Sci. USA 1956, 42, 578–580. [Google Scholar] [CrossRef] [Green Version]
- Bownds, D. Site of Attachment of Retinal in Rhodopsin. Nature 1967, 216, 1178–1181. [Google Scholar] [CrossRef]
- Hargrave, P.A.; McDowell, J.H.; Curtis, D.R.; Wang, J.K.; Juszczak, E.; Fong, S.-L.; Rao, J.K.M.; Argos, P. The structure of bovine rhodopsin. Biophys. Struct. Mech. 1983, 9, 235–244. [Google Scholar] [CrossRef]
- Palczewski, K.; Kumasaka, T.; Hori, T.; Behnke, C.A.; Motoshima, H.; Fox, B.A.; Le Trong, I.; Teller, D.C.; Okada, T.; Stenkamp, R.E.; et al. Crystal Structure of Rhodopsin: A G Protein-Coupled Receptor. Science 2000, 289, 739–745. [Google Scholar] [CrossRef] [Green Version]
- Murakami, M.; Kouyama, T. Crystal structure of squid rhodopsin. Nature 2008, 453, 363–367. [Google Scholar] [CrossRef]
- Hubbard, R.; Kropf, A. The action of light on rhodopsin. Proc. Natl. Acad. Sci. USA 1958, 44, 130–139. [Google Scholar] [CrossRef] [Green Version]
- Kropf, A.; Hubbard, R. The mechanism of bleaching rhodopsin. Ann. N. Y. Acad. Sci. 1958, 74, 266–280. [Google Scholar] [CrossRef]
- Choe, H.-W.; Kim, Y.J.; Park, J.H.; Morizumi, T.; Pai, E.F.; Krauß, N.; Hofmann, K.P.; Scheerer, P.; Ernst, O.P. Crystal structure of metarhodopsin II. Nature 2011, 471, 651–655. [Google Scholar] [CrossRef]
- Wald, G. The Molecular Basis of Visual Excitation. Nature 1968, 219, 800–807. [Google Scholar] [CrossRef]
- Ballesteros, J.A.; Weinstein, H. Integrated methods for the construction of three-dimensional models and computational probing of structure-function relations in G protein-coupled receptors. Methods Neurosci. 1995, 25, 366–428. [Google Scholar]
- Fritze, O.; Filipek, S.; Kuksa, V.; Palczewski, K.; Hofmann, K.P.; Ernst, O.P. Role of the conserved NPxxY(x)5,6F motif in the rhodopsin ground state and during activation. Proc. Natl. Acad. Sci. USA 2003, 100, 2290–2295. [Google Scholar] [CrossRef] [Green Version]
- Wess, J.; Nanavati, S.; Vogel, Z.; Maggio, R. Functional role of proline and tryptophan residues highly conserved among G protein-coupled receptors studied by mutational analysis of the m3 muscarinic receptor. EMBO J. 1993, 12, 331–338. [Google Scholar] [CrossRef]
- Galés, C.; Kowalski-Chauvel, A.; Dufour, M.N.; Seva, C.; Moroder, L.; Pradayrol, L.; Vaysse, N.; Fourmy, D.; Silvente-Poirot, S. Mutation of Asn-391 within the Conserved NPXXY Motif of the Cholecystokinin B Receptor Abolishes Gq Protein Activation without Affecting Its Association with the Receptor. J. Biol. Chem. 2000, 275, 17321–17327. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mazna, P.; Grycova, L.; Balik, A.; Zemkova, H.; Friedlova, E.; Obsilova, V.; Obsil, T.; Teisinger, J. The role of proline residues in the structure and function of human MT2 melatonin receptor. J. Pineal Res. 2008, 45, 361–372. [Google Scholar] [CrossRef] [PubMed]
- Borroto-Escuela, D.O.; Romero-Fernandez, W.; García-Negredo, G.; Correia, P.A.; Garriga, P.; Fuxe, K.; Ciruela, F. Dissecting the Conserved NPxxY Motif of the M3 Muscarinic Acetylcholine Receptor: Critical Role of Asp-7.49 for Receptor Signaling and Multiprotein Complex Formation. Cell. Physiol. Biochem. 2011, 28, 1009–1022. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhou, Q.; Yang, D.; Wu, M.; Guo, Y.; Guo, W.; Zhong, L.; Cai, X.; Dai, A.; Jang, W.; Shakhnovich, E.I.; et al. Common activation mechanism of class A GPCRs. eLife 2019, 8, e50279. [Google Scholar] [CrossRef]
- Ovchinnikov, Y. Rhodopsin and bacteriorhodopsin: Structure-function relationships. FEBS Lett. 1982, 148, 179–191. [Google Scholar] [CrossRef] [Green Version]
- Leung, N.Y.; Thakur, D.P.; Gurav, A.S.; Kim, S.H.; Di Pizio, A.; Niv, M.Y.; Montell, C. Functions of Opsins in Drosophila Taste. Curr. Biol. 2020, 30, 1367–1379.e6. [Google Scholar] [CrossRef]
- Ramirez, M.D.; Oakley, T.H. Eye-independent, light-activated chromatophore expansion (LACE) and expression of phototransduction genes in the skin of Octopus bimaculoides. J. Exp. Biol. 2015, 218, 1513–1520. [Google Scholar] [CrossRef] [Green Version]
- Porter, M.L.; Blasic, J.R.; Bok, M.J.; Cameron, E.G.; Pringle, T.; Cronin, T.W.; Robinson, P.R. Shedding new light on opsin evolution. Proc. R. Soc. B Boil. Sci. 2011, 279, 3–14. [Google Scholar] [CrossRef] [Green Version]
- Feuda, R.; Hamilton, S.C.; McInerney, J.O.; Pisani, D. Metazoan opsin evolution reveals a simple route to animal vision. Proc. Natl. Acad. Sci. USA 2012, 109, 18868–18872. [Google Scholar] [CrossRef] [Green Version]
- Shen, W.L.; Kwon, Y.; Adegbola, A.A.; Luo, J.; Chess, A.; Montell, C. Function of Rhodopsin in Temperature Discrimination in Drosophila. Science 2011, 331, 1333–1336. [Google Scholar] [CrossRef]
- Senthilan, P.R.; Piepenbrock, D.; Ovezmyradov, G.; Nadrowski, B.; Bechstedt, S.; Pauls, S.; Winkler, M.; Möbius, W.; Howard, J.; Göpfert, M.C. Drosophila Auditory Organ Genes and Genetic Hearing Defects. Cell 2012, 150, 1042–1054. [Google Scholar] [CrossRef] [Green Version]
- Feuda, R.; Menon, A.K.; Göpfert, M.C. Rethinking Opsins. Mol. Biol. Evol. 2022, 39, msac033. [Google Scholar] [CrossRef]
- Leung, N.Y.; Montell, C. Unconventional Roles of Opsins. Annu. Rev. Cell Dev. Biol. 2017, 33, 241–264. [Google Scholar] [CrossRef]
- Altschul, S.F.; Gish, W.; Miller, W.; Myers, E.W.; Lipman, D.J. Basic local alignment search tool. J. Mol. Biol. 1990, 215, 403–410. [Google Scholar] [CrossRef]
- Ramirez, M.D.; Pairett, A.N.; Pankey, M.S.; Serb, J.; Speiser, D.I.; Swafford, A.J.; Oakley, T.H. The last common ancestor of most bilaterian animals possessed at least 9 opsins. Genome Biol. Evol. 2016, 8, 3640–3652. [Google Scholar] [CrossRef] [Green Version]
- Lowe, E.K.; Garm, A.L.; Ullrich-Lüter, E.; Cuomo, C.; Arnone, M.I. The crowns have eyes: Multiple opsins found in the eyes of the crown-of-thorns starfish Acanthaster planci. BMC Evol. Biol. 2018, 18, 168. [Google Scholar] [CrossRef]
- D’Aniello, S.; Delroisse, J.; Valero-Gracia, A.; Lowe, E.; Byrne, M.; Cannon, J.; Halanych, K.; Elphick, M.; Mallefet, J.; Kaul-Strehlow, S.; et al. Opsin evolution in the Ambulacraria. Mar. Genom. 2015, 24, 177–183. [Google Scholar] [CrossRef]
- Conzelmann, M.; Williams, E.A.; Krug, K.; Franz-Wachtel, M.; Macek, B.; Jékely, G. The neuropeptide complement of the marine annelid Platynereis dumerilii. BMC Genom. 2013, 14, 906. [Google Scholar] [CrossRef]
- Speiser, D.I.; Pankey, M.S.; Zaharoff, A.K.; Battelle, B.A.; Bracken-Grissom, H.D.; Breinholt, J.W.; Bybee, S.M.; Cronin, T.W.; Garm, A.; Lindgren, A.R.; et al. Using phylogenetically-informed annotation (PIA) to search for light-interacting genes in transcriptomes from non-model organisms. BMC Bioinform. 2014, 15, 350. [Google Scholar] [CrossRef] [Green Version]
- Pérez-Moreno, J.L.; DeLeo, D.M.; Palero, F.; Bracken-Grissom, H.D. Phylogenetic annotation and genomic architecture of opsin genes in Crustacea. Hydrobiologia 2018, 825, 159–175. [Google Scholar] [CrossRef]
- Shen, W.; Le, S.; Li, Y.; Hu, F. SeqKit: A Cross-Platform and Ultrafast Toolkit for FASTA/Q File Manipulation. PLoS ONE 2016, 11, e0163962. [Google Scholar] [CrossRef]
- Li, W.; Godzik, A. Cd-hit: A fast program for clustering and comparing large sets of protein or nucleotide sequences. Bioinformatics 2006, 22, 1658–1659. [Google Scholar] [CrossRef] [Green Version]
- Fu, L.; Niu, B.; Zhu, Z.; Wu, S.; Li, W. CD-HIT: Accelerated for clustering the next-generation sequencing data. Bioinformatics 2012, 28, 3150–3152. [Google Scholar] [CrossRef]
- Mirarab, S.; Nguyen, N.-P.; Guo, S.; Wang, L.-S.; Kim, J.; Warnow, T. PASTA: Ultra-Large Multiple Sequence Alignment for Nucleotide and Amino-Acid Sequences. J. Comput. Biol. 2015, 22, 377–386. [Google Scholar] [CrossRef]
- Minh, B.Q.; Schmidt, H.A.; Chernomor, O.; Schrempf, D.; Woodhams, M.D.; von Haeseler, A.; Lanfear, R.; Teeling, E. IQ-TREE 2: New Models and Efficient Methods for Phylogenetic Inference in the Genomic Era. Mol. Biol. Evol. 2020, 37, 1530–1534. [Google Scholar] [CrossRef] [Green Version]
- Junier, T.; Zdobnov, E.M. The Newick utilities: High-throughput phylogenetic tree processing in the UNIX shell. Bioinformatics 2010, 26, 1669–1670. [Google Scholar] [CrossRef] [Green Version]
- Aberer, A.J.; Krompass, D.; Stamatakis, A. Pruning Rogue Taxa Improves Phylogenetic Accuracy: An Efficient Algorithm and Webservice. Syst. Biol. 2013, 62, 162–166. [Google Scholar] [CrossRef] [Green Version]
- Saunders, A.M.; Ashlock, D.; Graether, S.P. Testing the rogue taxa hypothesis for clustering instability. J. Theor. Biol. 2019, 472, 36–45. [Google Scholar] [CrossRef]
- Aberer, A.J.; Krompaß, D.; Stamatakis, A. RogueNaRok: An Efficient and Exact Algorithm for Rogue Taxon Identification. Heidelberg Institute for Theoretical Studies: Exelixis-RRDR-2011–10. 2011. Available online: https://cme.h-its.org/exelixis/pubs/Exelixis-RRDR-2011-10.pdf (accessed on 1 June 2022).
- Mai, U.; Mirarab, S. TreeShrink: Fast and accurate detection of outlier long branches in collections of phylogenetic trees. BMC Genom. 2018, 19, 272. [Google Scholar] [CrossRef] [Green Version]
- Capella-Gutiérrez, S.; Silla-Martínez, J.M.; Gabaldón, T. trimAl: A tool for automated alignment trimming in large-scale phylogenetic analyses. Bioinformatics 2009, 25, 1972–1973. [Google Scholar] [CrossRef]
- Guindon, S.; Dufayard, J.-F.; Lefort, V.; Anisimova, M.; Hordijk, W.; Gascuel, O. New Algorithms and Methods to Estimate Maximum-Likelihood Phylogenies: Assessing the Performance of PhyML 3.0. Syst. Biol. 2010, 59, 307–321. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Anisimova, M.; Gil, M.; Dufayard, J.-F.; Dessimoz, C.; Gascuel, O. Survey of Branch Support Methods Demonstrates Accuracy, Power, and Robustness of Fast Likelihood-based Approximation Schemes. Syst. Biol. 2011, 60, 685–699. [Google Scholar] [CrossRef] [PubMed]
- Minh, B.Q.; Nguyen, M.A.T.; Von Haeseler, A. Ultrafast Approximation for Phylogenetic Bootstrap. Mol. Biol. Evol. 2013, 30, 1188–1195. [Google Scholar] [CrossRef] [PubMed]
- Hoang, D.T.; Chernomor, O.; Von Haeseler, A.; Minh, B.Q.; Vinh, L.S. UFBoot2: Improving the Ultrafast Bootstrap Approximation. Mol. Biol. Evol. 2018, 35, 518–522. [Google Scholar] [CrossRef]
- Simmons, M.P.; Norton, A.P. Divergent maximum-likelihood-branch-support values for polytomies. Mol. Phylogenet. Evol. 2014, 73, 87–96. [Google Scholar] [CrossRef]
- Liu, K.; Raghavan, S.; Nelesen, S.; Linder, C.R.; Warnow, T. Rapid and Accurate Large-Scale Coestimation of Sequence Alignments and Phylogenetic Trees. Science 2009, 324, 1561–1564. [Google Scholar] [CrossRef] [Green Version]
- Ali, R.H.; Bogusz, M.; Whelan, S. Identifying Clusters of High Confidence Homologies in Multiple Sequence Alignments. Mol. Biol. Evol. 2019, 36, 2340–2351. [Google Scholar] [CrossRef]
- Chang, J.-M.; Di Tommaso, P.; Notredame, C. TCS: A New Multiple Sequence Alignment Reliability Measure to Estimate Alignment Accuracy and Improve Phylogenetic Tree Reconstruction. Mol. Biol. Evol. 2014, 31, 1625–1637. [Google Scholar] [CrossRef]
- Tan, G.; Muffato, M.; Ledergerber, C.; Herrero, J.; Goldman, N.; Gil, M.; Dessimoz, C. Current Methods for Automated Filtering of Multiple Sequence Alignments Frequently Worsen Single-Gene Phylogenetic Inference. Syst. Biol. 2015, 64, 778–791. [Google Scholar] [CrossRef] [Green Version]
- Huerta-Cepas, J.; Serra, F.; Bork, P. ETE 3: Reconstruction, Analysis, and Visualization of Phylogenomic Data. Mol. Biol. Evol. 2016, 33, 1635–1638. [Google Scholar] [CrossRef] [Green Version]
- Tareen, A.; Kinney, J.B. Logomaker: Beautiful sequence logos in Python. Bioinformatics 2020, 36, 2272–2274. [Google Scholar] [CrossRef]
- Hunter, J.D. Matplotlib: A 2D graphics environment. Comput. Sci. Eng. 2007, 9, 90–95. [Google Scholar] [CrossRef]
- Huson, D.H.; Scornavacca, C. Dendroscope 3: An Interactive Tool for Rooted Phylogenetic Trees and Networks. Syst. Biol. 2012, 61, 1061–1067. [Google Scholar] [CrossRef] [Green Version]
- Feuda, R.; Rota-Stabelli, O.; Oakley, T.; Pisani, D. The Comb Jelly Opsins and the Origins of Animal Phototransduction. Genome Biol. Evol. 2014, 6, 1964–1971. [Google Scholar] [CrossRef] [Green Version]
- Artigas, G.Q.; Lapébie, P.; Leclère, L.; Takeda, N.; Deguchi, R.; Jékely, G.; Momose, T.; Houliston, E. A gonad-expressed opsin mediates light-induced spawning in the jellyfish Clytia. eLife 2018, 7, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Rawlinson, K.A.; Lapraz, F.; Ballister, E.R.; Terasaki, M.; Rodgers, J.; McDowell, R.J.; Girstmair, J.; Criswell, K.E.; Boldogkoi, M.; Simpson, F.; et al. Extraocular, rod-like photoreceptors in a flatworm express xenopsin photopigment. eLife 2019, 8, e45465. [Google Scholar] [CrossRef]
- Hara, T.; Hara, R. New Photosensitive Pigment Found in the Retina of the Squid Ommastrephes. Nature 1965, 206, 1331–1334. [Google Scholar] [CrossRef]
- Hara, T.; Hara, R.; Takeuchi, J. Vision in Octopus and Squid: Rhodopsin and Retinochrome in the Octopus Retina. Nature 1967, 214, 572–573. [Google Scholar] [CrossRef]
- Jiang, M.; Pandey, S.; Fong, H.K. An opsin homologue in the retina and pigment epithelium. Investig. Ophthalmol. Vis. Sci. 1993, 34, 3669–3678. [Google Scholar]
- Shen, D.; Jiang, M.; Hao, W.; Tao, L.; Salazar, M.; Fong, H.K.W. A Human Opsin-Related Gene That Encodes a Retinaldehyde-Binding Protein. Biochemistry 1994, 33, 13117–13125. [Google Scholar] [CrossRef]
- Sun, H.; Gilbert, D.J.; Copeland, N.G.; Jenkins, N.A.; Nathans, J. Peropsin, a novel visual pigment-like protein located in the apical microvilli of the retinal pigment epithelium. Proc. Natl. Acad. Sci. USA 1997, 94, 9893–9898. [Google Scholar] [CrossRef] [Green Version]
- Henze, M.J.; Oakley, T.H. The Dynamic Evolutionary History of Pancrustacean Eyes and Opsins. Integr. Comp. Biol. 2015, 55, 830–842. [Google Scholar] [CrossRef] [Green Version]
- Nagata, T.; Koyanagi, M.; Tsukamoto, H.; Terakita, A. Identification and characterization of a protostome homologue of peropsin from a jumping spider. J. Comp. Physiol. A Sens. Neural Behav. Physiol. 2010, 196, 51–59. [Google Scholar] [CrossRef]
- Battelle, B.-A.; Kempler, K.E.; Saraf, S.R.; Marten, C.E.; Dugger, D.R.; Speiser, D.; Oakley, T.H. Opsins in Limulus eyes: Characterization of three visible light-sensitive opsins unique to and co-expressed in median eye photoreceptors and a peropsin/RGR that is expressed in all eyes. J. Exp. Biol. 2014, 218, 466–479. [Google Scholar] [CrossRef] [Green Version]
- Marlow, H.; Tosches, M.A.; Tomer, R.; Steinmetz, P.R.; Lauri, A.; Larsson, T.; Arendt, D. Larval body patterning and apical organs are conserved in animal evolution. BMC Biol. 2014, 12, 7. [Google Scholar] [CrossRef] [Green Version]
- Troemel, E.R.; Chou, J.H.; Dwyer, N.D.; Colbert, H.A.; Bargmann, C.I. Divergent seven transmembrane receptors are candidate chemosensory receptors in C. elegans. Cell 1995, 83, 207–218. [Google Scholar] [CrossRef] [Green Version]
- Futahashi, R.; Kawahara-Miki, R.; Kinoshita, M.; Yoshitake, K.; Yajima, S.; Arikawa, K.; Fukatsu, T. Extraordinary diversity of visual opsin genes in dragonflies. Proc. Natl. Acad. Sci. USA 2015, 112, E1247–E1256. [Google Scholar] [CrossRef] [Green Version]
- Böhm, A.; Meusemann, K.; Misof, B.; Pass, G. Hypothesis on monochromatic vision in scorpionflies questioned by new transcriptomic data. Sci. Rep. 2018, 8, 9872. [Google Scholar] [CrossRef] [Green Version]
- Tsukamoto, H.; Terakita, A. Diversity and functional properties of bistable pigments. Photochem. Photobiol. Sci. 2010, 9, 1435. [Google Scholar] [CrossRef]
- Terakita, A. The opsins. Genome Biol. 2005, 6, 213. [Google Scholar] [CrossRef] [Green Version]
- Chen, K.; Durand, D.; Farach-Colton, M. NOTUNG: A Program for Dating Gene Duplications and Optimizing Gene Family Trees. J. Comput. Biol. 2000, 7, 429–447. [Google Scholar] [CrossRef] [PubMed]
- Szöllősi, G.J.; Tannier, E.; Daubin, V.; Boussau, B. The Inference of Gene Trees with Species Trees. Syst. Biol. 2014, 64, e42–e62. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Musser, J.M.; Arendt, D. Loss and gain of cone types in vertebrate ciliary photoreceptor evolution. Dev. Biol. 2017, 431, 26–35. [Google Scholar] [CrossRef] [PubMed]
- Colbourne, J.K.; Pfrender, M.E.; Gilbert, D.; Thomas, W.K.; Tucker, A.; Oakley, T.H.; Tokishita, S.; Aerts, A.; Arnold, G.J.; Basu, M.K.; et al. The Ecoresponsive Genome of Daphnia pulex. Science 2011, 331, 555–561. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, X.; Yuan, J.; Sun, Y.; Li, S.; Gao, Y.; Yu, Y.; Liu, C.; Wang, Q.; Lv, X.; Zhang, X.; et al. Penaeid shrimp genome provides insights into benthic adaptation and frequent molting. Nat. Commun. 2019, 10, 356. [Google Scholar] [CrossRef] [Green Version]
- Suvorov, A.; Jensen, N.O.; Sharkey, C.R.; Fujimoto, M.S.; Bodily, P.; Wightman, H.M.C.; Ogden, T.H.; Clement, M.J.; Bybee, S.M. Opsins have evolved under the permanent heterozygote model: Insights from phylotranscriptomics of Odonata. Mol. Ecol. 2017, 26, 1306–1322. [Google Scholar] [CrossRef]
- Battelle, B.-A.; Ryan, J.F.; Kempler, K.E.; Saraf, S.R.; Marten, C.E.; Warren, W.C.; Minx, P.J.; Montague, M.J.; Green, P.J.; Schmidt, S.A.; et al. Opsin Repertoire and Expression Patterns in Horseshoe Crabs: Evidence from the Genome of Limulus polyphemus (Arthropoda: Chelicerata). Genome Biol. Evol. 2016, 8, 1571–1589. [Google Scholar] [CrossRef] [Green Version]
- Porter, M.L.; Awata, H.; Bok, M.J.; Cronin, T.W. Exceptional diversity of opsin expression patterns in Neogonodactylus oerstedii (Stomatopoda) retinas. Proc. Natl. Acad. Sci. USA 2020, 117, 8948–8957. [Google Scholar] [CrossRef]
- Feuda, R.; Goulty, M.; Zadra, N.; Gasparetti, T.; Rosato, E.; Pisani, D.; Rizzoli, A.; Segata, N.; Ometto, L.; Stabelli, O.R. Phylogenomics of Opsin Genes in Diptera Reveals Lineage-Specific Events and Contrasting Evolutionary Dynamics in Anopheles and Drosophila. Genome Biol. Evol. 2021, 13, evab170. [Google Scholar] [CrossRef]
- Koyanagi, M.; Terakita, A.; Kubokawa, K.; Shichida, Y. Amphioxus homologs of Go-coupled rhodopsin and peropsin having 11-cis- and all -trans -retinals as their chromophores. FEBS Lett. 2002, 531, 525–528. [Google Scholar] [CrossRef] [Green Version]
- Nagata, T.; Koyanagi, M.; Lucas, R.; Terakita, A. An all-trans-retinal-binding opsin peropsin as a potential dark-active and light-inactivated G protein-coupled receptor. Sci. Rep. 2018, 8, 3535. [Google Scholar] [CrossRef] [Green Version]
- Hao, W.; Fong, H.K.W. The Endogenous Chromophore of Retinal G Protein-coupled Receptor Opsin from the Pigment Epithelium. J. Biol. Chem. 1999, 274, 6085–6090. [Google Scholar] [CrossRef] [Green Version]
- Gehring, W.J. The evolution of vision. Wiley Interdiscip. Rev. Dev. Biol. 2014, 3, 1–40. [Google Scholar] [CrossRef]
- Kato, M.; Sugiyama, T.; Sakai, K.; Yamashita, T.; Fujita, H.; Sato, K.; Tomonari, S.; Shichida, Y.; Ohuchi, H. Two Opsin 3-Related Proteins in the Chicken Retina and Brain: A TMT-Type Opsin 3 Is a Blue-Light Sensor in Retinal Horizontal Cells, Hypothalamus, and Cerebellum. PLoS ONE 2016, 11, e0163925. [Google Scholar] [CrossRef]
- Wald, G. The Photochemical Basis of Rod Vision. J. Opt. Soc. Am. 1951, 41, 949–955. [Google Scholar] [CrossRef]
- Hao, W.; Fong, H.K.W. Blue and Ultraviolet Light-Absorbing Opsin from the Retinal Pigment Epithelium. Biochemistry 1996, 35, 6251–6256. [Google Scholar] [CrossRef]
- Kaylor, J.J.; Xu, T.; Ingram, N.T.; Tsan, A.; Hakobyan, H.; Fain, G.; Travis, G.H. Blue light regenerates functional visual pigments in mammals through a retinyl-phospholipid intermediate. Nat. Commun. 2017, 8, 16. [Google Scholar] [CrossRef] [Green Version]
- Pepe, I.M.; Cugnoli, C. New trends in photobiology. J. Photochem. Photobiol. B Biol. 1992, 13, 5–17. [Google Scholar] [CrossRef]
- Goldsmith, T.H. Evolutionary tinkering with visual photoreception. Vis. Neurosci. 2012, 30, 21–37. [Google Scholar] [CrossRef]
- Koyanagi, M.; Terakita, A. Diversity of animal opsin-based pigments and their optogenetic potential. Biochim. Biophys. Acta 2013, 1837, 710–716. [Google Scholar] [CrossRef] [Green Version]
- Verasztó, C.; Gühmann, M.; Jia, H.; Rajan, V.B.V.; Bezares-Calderón, L.A.; Piñeiro-Lopez, C.; Randel, N.; Shahidi, R.; Michiels, N.K.; Yokoyama, S.; et al. Ciliary and rhabdomeric photoreceptor-cell circuits form a spectral depth gauge in marine zooplankton. eLife 2018, 7, e36440. [Google Scholar] [CrossRef]
- Arendt, D.; Tessmar-Raible, K.; Snyman, H.; Dorresteijn, A.W.; Wittbrodt, J. Ciliary Photoreceptors with a Vertebrate-Type Opsin in an Invertebrate Brain. Science 2004, 306, 869–871. [Google Scholar] [CrossRef] [Green Version]
- Eriksson, B.J.; Fredman, D.; Steiner, G.; Schmid, A. Characterisation and localisation of the opsin protein repertoire in the brain and retinas of a spider and an onychophoran. BMC Evol. Biol. 2013, 13, 186. [Google Scholar] [CrossRef] [Green Version]
- Cook, J.D.; Ng, S.Y.; Lloyd, M.; Eddington, S.; Sun, H.; Nathans, J.; Bok, D.; Radu, R.A.; Travis, G.H. Peropsin modulates transit of vitamin A from retina to retinal pigment epithelium. J. Biol. Chem. 2017, 292, 21407–21416. [Google Scholar] [CrossRef] [Green Version]
- Toh, P.P.C.; Yap, A.M.Y.; Sriram, G.; Bigliardi, P.; Bigliardi-Qi, M. Expression of peropsin in human skin is related to phototransduction of violet light in keratinocytes. Exp. Dermatol. 2016, 25, 1002–1005. [Google Scholar] [CrossRef] [Green Version]
- Bailey, M.J.; Cassone, V.M. Opsin Photoisomerases in the Chick Retina and Pineal Gland: Characterization, Localization, and Circadian Regulation. Investig. Opthalmol. Vis. Sci. 2004, 45, 769–775. [Google Scholar] [CrossRef]
- Ksantini, M.; Sénéchal, A.; Humbert, G.; Arnaud, B.; Hamel, C.P. RRH, Encoding the RPE-Expressed Opsin-Like Peropsin, Is Not Mutated in Retinitis Pigmentosa and Allied Diseases. Ophthalmic Genet. 2007, 28, 31–37. [Google Scholar] [CrossRef] [Green Version]
- Rivolta, C.; Berson, E.L.; Dryja, T.P. Mutation Screening of the Peropsin Gene, a Retinal Pigment Epithelium Specific Rhodopsin Homolog, in Patients with Retinitis Pigmentosa and Allied Diseases. Mol. Vis. 2006, 12, 1511–1515. [Google Scholar]
- Morimura, H.; Saindelle-Ribeaudeau, F.; Berson, E.L.; Dryja, T.P. Mutations in RGR, encoding a light-sensitive opsin homologue, in patients with retinitis pigmentosa. Nat. Genet. 1999, 23, 393–394. [Google Scholar] [CrossRef]
- Chen, P.; Hao, W.; Rife, L.; Wang, X.P.; Shen, D.; Chen, J.; Ogden, T.; Van Boemel, G.B.; Wu, L.; Yang, M.; et al. A photic visual cycle of rhodopsin regeneration is dependent on Rgr. Nat. Genet. 2001, 28, 256–260. [Google Scholar] [CrossRef]
- Pandey, S.; Blanks, J.C.; Spee, C.; Jiang, M.; Fong, H.K. Cytoplasmic Retinal Localization of an Evolutionary Homolog of the Visual Pigments. Exp. Eye Res. 1994, 58, 605–613. [Google Scholar] [CrossRef] [PubMed]
- Shichida, Y.; Matsuyama, T. Evolution of opsins and phototransduction. Philos. Trans. R. Soc. B: Biol. Sci. 2009, 364, 2881–2895. [Google Scholar] [CrossRef] [PubMed]
- Nagata, T.; Koyanagi, M.; Terakita, A. Evolution and Functional Diversity of Opsin-Based Photopigments. Available online: http://photobiology.info/Terakita.html (accessed on 3 August 2021).
- Wenzel, A.; Oberhauser, V.; Pugh, E.N.; Lamb, T.D.; Grimm, C.; Samardzija, M.; Fahl, E.; Seeliger, M.W.; Remé, C.E.; von Lintig, J. The Retinal G Protein-coupled Receptor (RGR) Enhances Isomerohydrolase Activity Independent of Light. J. Biol. Chem. 2005, 280, 29874–29884. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Radu, R.A.; Hu, J.; Peng, J.; Bok, D.; Mata, N.L.; Travis, G.H. Retinal Pigment Epithelium-Retinal G Protein Receptor-Opsin Mediates Light-dependent Translocation of All-trans-retinyl Esters for Synthesis of Visual Chromophore in Retinal Pigment Epithelial Cells. J. Biol. Chem. 2008, 283, 19730–19738. [Google Scholar] [CrossRef] [Green Version]
- Zhang, J.; Choi, E.H.; Tworak, A.; Salom, D.; Leinonen, H.; Sander, C.L.; Hoang, T.V.; Handa, J.T.; Blackshaw, S.; Palczewska, G.; et al. Photic generation of 11-cis-retinal in bovine retinal pigment epithelium. J. Biol. Chem. 2019, 294, 19137–19154. [Google Scholar] [CrossRef]
- Sato, K.; Yamashita, T.; Ohuchi, H.; Takeuchi, A.; Gotoh, H.; Ono, K.; Mizuno, M.; Mizutani, Y.; Tomonari, S.; Sakai, K.; et al. Opn5L1 is a retinal receptor that behaves as a reverse and self-regenerating photoreceptor. Nat. Commun. 2018, 9, 1255. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Z.; Fong, H.K. Coexpression of nonvisual opsin, retinal G protein-coupled receptor, and visual pigments in human and bovine cone photoreceptors. Mol. Vis. 2018, 24, 434–442. [Google Scholar]
- Morshedian, A.; Kaylor, J.J.; Ng, S.Y.; Tsan, A.; Frederiksen, R.; Xu, T.; Yuan, L.; Sampath, A.P.; Radu, R.A.; Fain, G.L.; et al. Light-Driven Regeneration of Cone Visual Pigments through a Mechanism Involving RGR Opsin in Müller Glial Cells. Neuron 2019, 102, 1172–1183.e5. [Google Scholar] [CrossRef]
- Hara, T.; Hara, R. Vision in Octopus and Squid: Rhodopsin and Retinochrome in the Squid Retina. Nature 1967, 214, 573–575. [Google Scholar] [CrossRef]
- Hara, T.; Hara, R. Regeneration of Squid Retinochrome. Nature 1968, 219, 450–454. [Google Scholar] [CrossRef]
- Ozaki, K.; Hara, R.; Hara, T. Histochemical localization of retinochrome and rhodopsin studied by fluorescence microscopy. Cell Tissue Res. 1983, 233, 335–345. [Google Scholar] [CrossRef]
- Hara, T.; Hara, R. Distribution of rhodopsin and retinochrome in the squid retina. J. Gen. Physiol. 1976, 67, 791–805. [Google Scholar] [CrossRef] [Green Version]
- Terakita, A.; Hara, R.; Hara, T. Retinal-binding protein as a shuttle for retinal in the rhodopsin-retinochrome system of the squid visual cells. Vis. Res. 1989, 29, 639–652. [Google Scholar] [CrossRef]
- Kingston, A.C.N.; Wardill, T.J.; Hanlon, R.T.; Cronin, T.W. An Unexpected Diversity of Photoreceptor Classes in the Longfin Squid, Doryteuthis pealeii. PLoS ONE 2015, 10, e0135381. [Google Scholar] [CrossRef] [Green Version]
- Hara, T.; Hara, R. Retinochrome and rhodopsin in the extraocular photoreceptor of the squid, Todarodes. J. Gen. Physiol. 1980, 75, 1–19. [Google Scholar] [CrossRef] [Green Version]
- Sea Urchin Genome Sequencing Consortium; Sodergren, E.; Weinstock, G.M.; Davidson, E.H.; Cameron, R.A.; Gibbs, R.A.; Angerer, R.C.; Angerer, L.M.; Arnone, M.I.; Burgess, D.R.; et al. The Genome of the Sea Urchin Strongylocentrotus purpuratus. Science 2006, 314, 941–952. [Google Scholar] [CrossRef] [Green Version]
- Davidson, P.L.; Guo, H.; Wang, L.; Berrio, A.; Zhang, H.; Chang, Y.; Soborowski, A.L.; McClay, D.R.; Fan, G.; Wray, G.A. Chromosomal-Level Genome Assembly of the Sea Urchin Lytechinus variegatus Substantially Improves Functional Genomic Analyses. Genome Biol. Evol. 2020, 12, 1080–1086. [Google Scholar] [CrossRef]
- Devine, E.L.; Oprian, D.D.; Theobald, D.L. Relocating the active-site lysine in rhodopsin and implications for evolution of retinylidene proteins. Proc. Natl. Acad. Sci. USA 2013, 110, 13351–13355. [Google Scholar] [CrossRef] [Green Version]



Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gühmann, M.; Porter, M.L.; Bok, M.J. The Gluopsins: Opsins without the Retinal Binding Lysine. Cells 2022, 11, 2441. https://doi.org/10.3390/cells11152441
Gühmann M, Porter ML, Bok MJ. The Gluopsins: Opsins without the Retinal Binding Lysine. Cells. 2022; 11(15):2441. https://doi.org/10.3390/cells11152441
Chicago/Turabian StyleGühmann, Martin, Megan L. Porter, and Michael J. Bok. 2022. "The Gluopsins: Opsins without the Retinal Binding Lysine" Cells 11, no. 15: 2441. https://doi.org/10.3390/cells11152441