
RELA∙8-Oxoguanine DNA Glycosylase1 Is an Epigenetic Regulatory Complex Coordinating the Hexosamine Biosynthetic Pathway in RSV Infection


 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Cell Cultures and Treatment
2.2. Quantification of Cellular UDP-GlcNAc
2.3. Proteomics Analysis of N-glycosylation
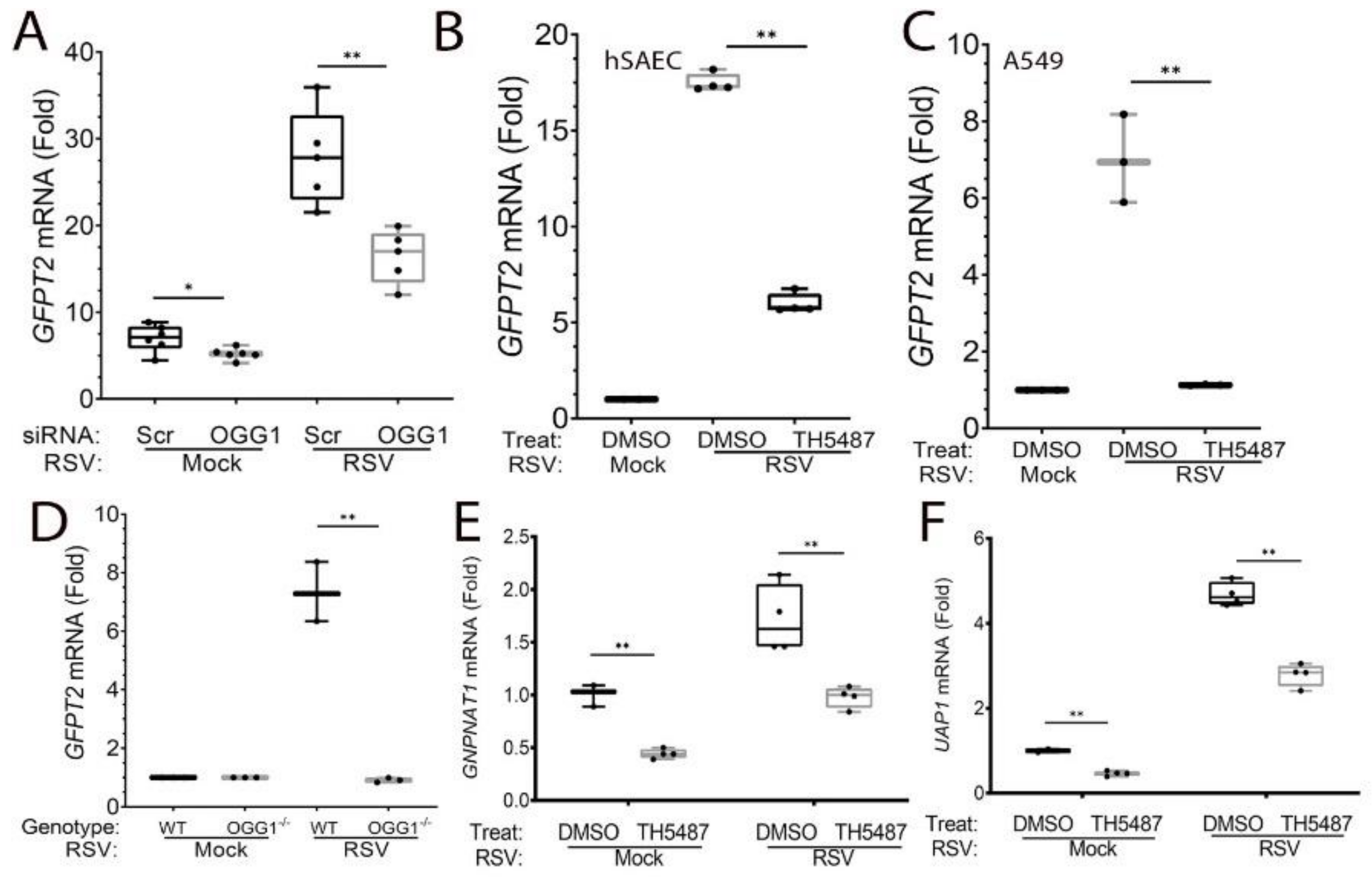
2.4. RNA Isolation and qRT-PCR
2.5. Western Blot
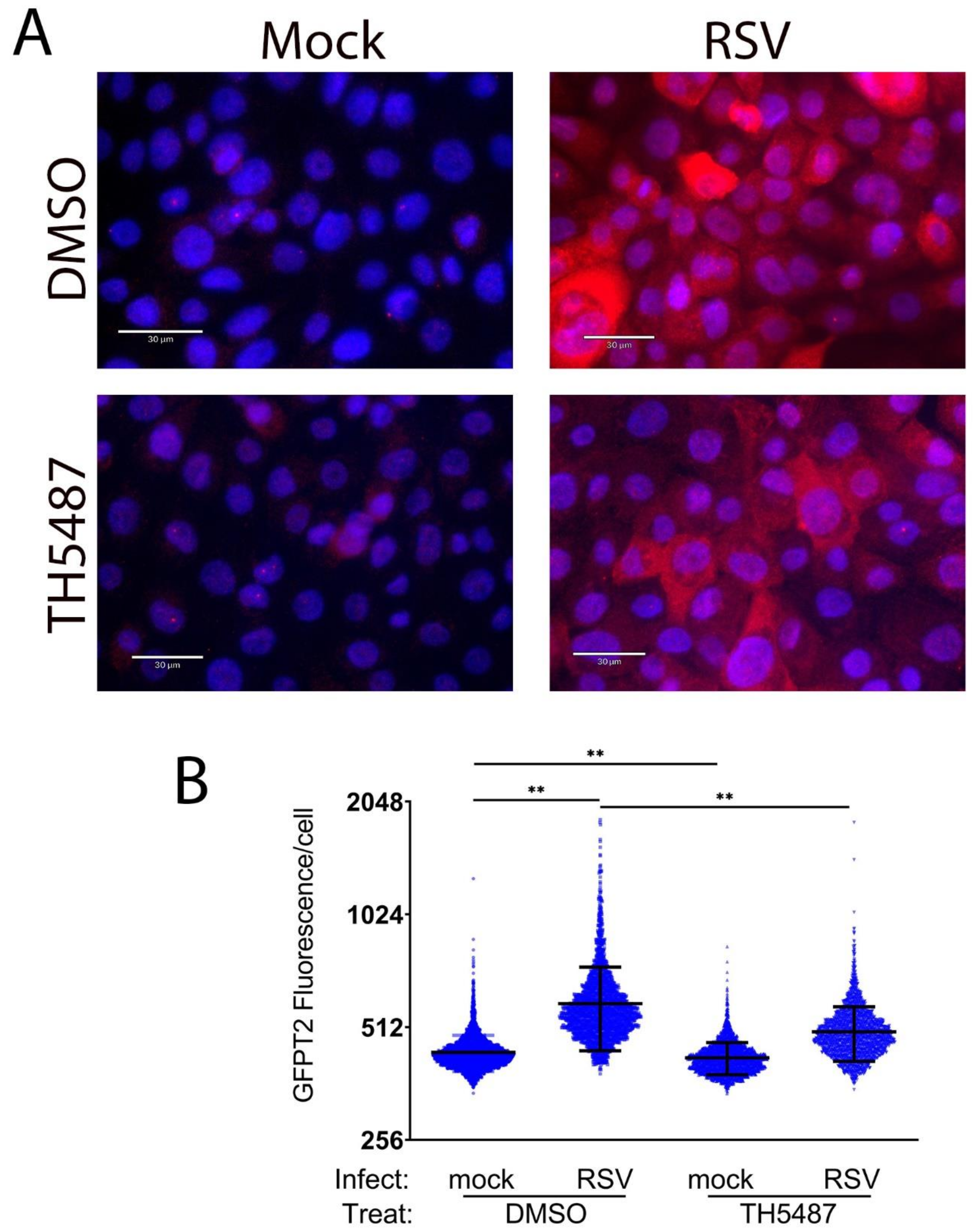
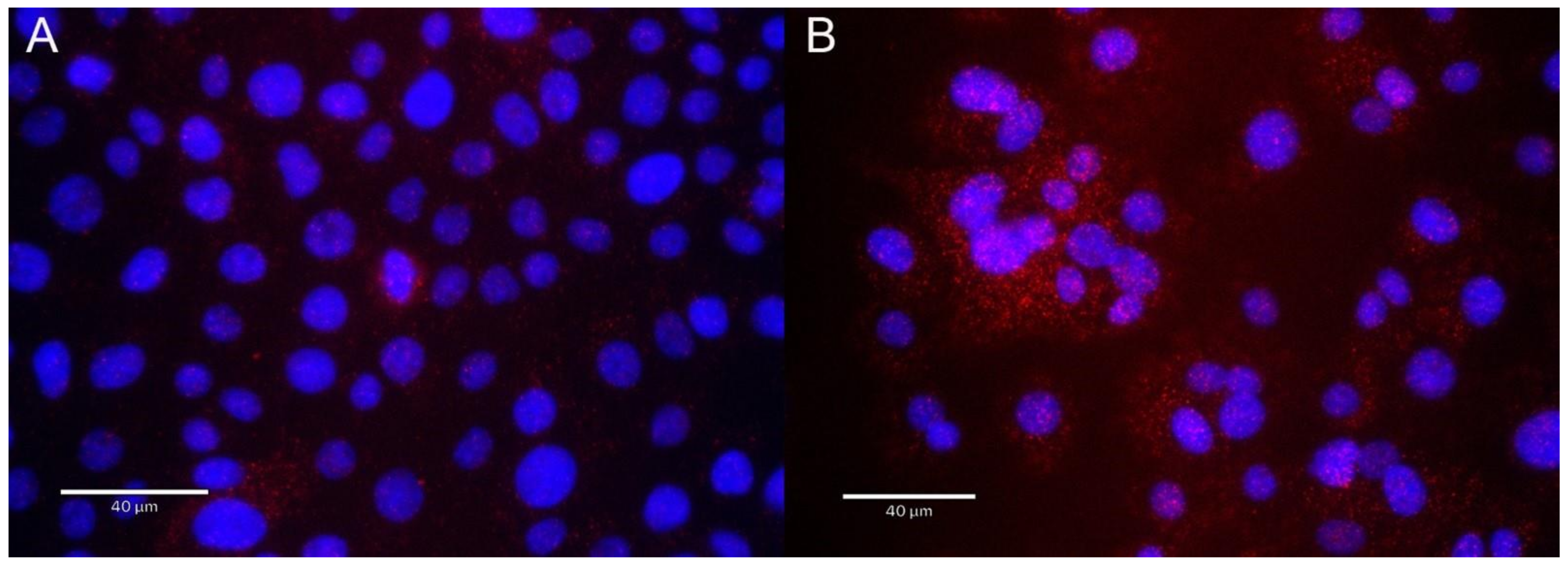
2.6. Immunofluorescence Microscopy and Flow Cytometry
2.7. Proximity Ligation Assay (PLA)
2.8. Co-Immunoprecipitation Assay
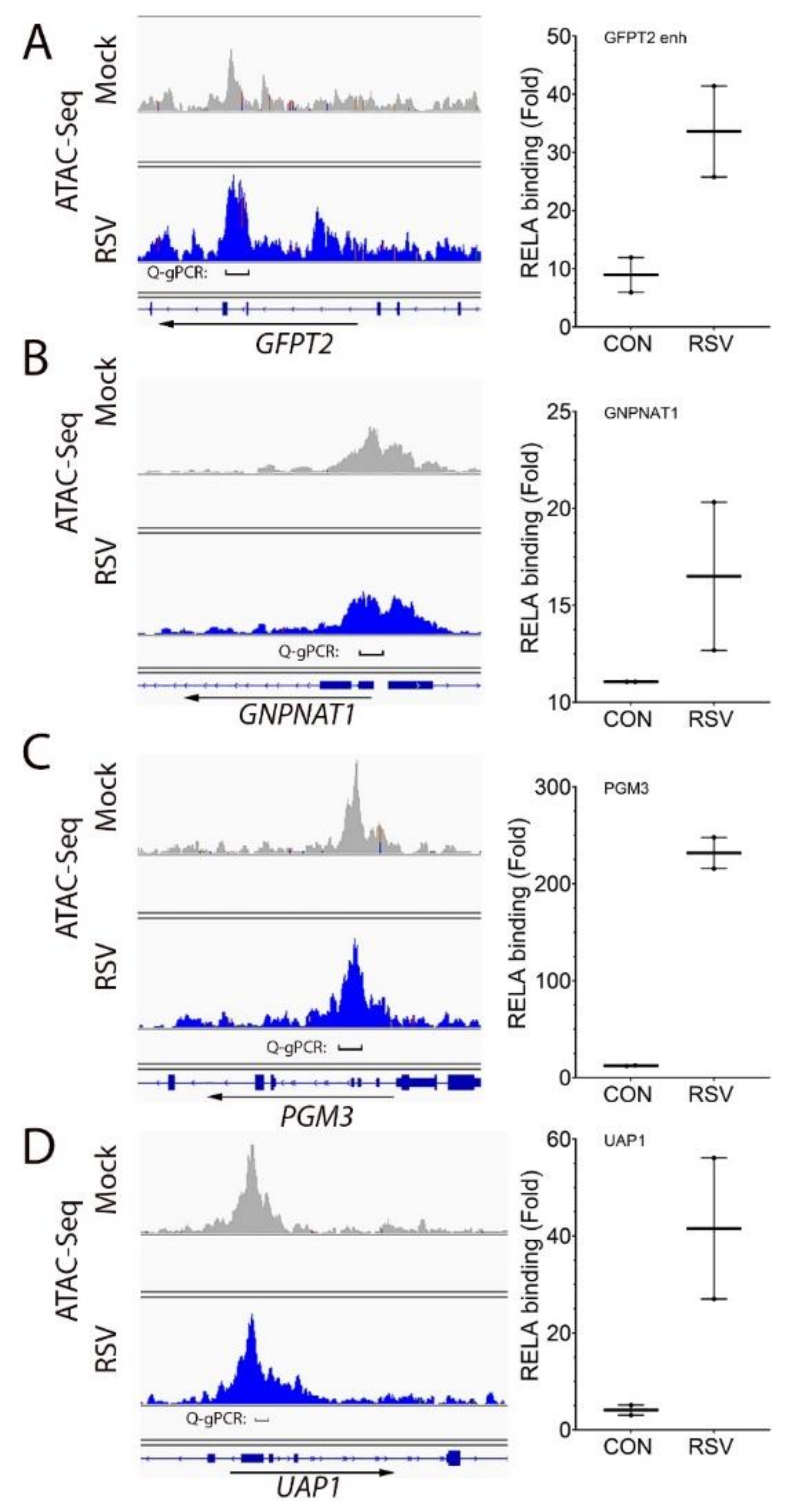
2.9. Two-Step Chromatin IP (XChIP)-Quantitative Genomic PCR (Q-gPCR)
2.10. Statistical Analyses
3. Results
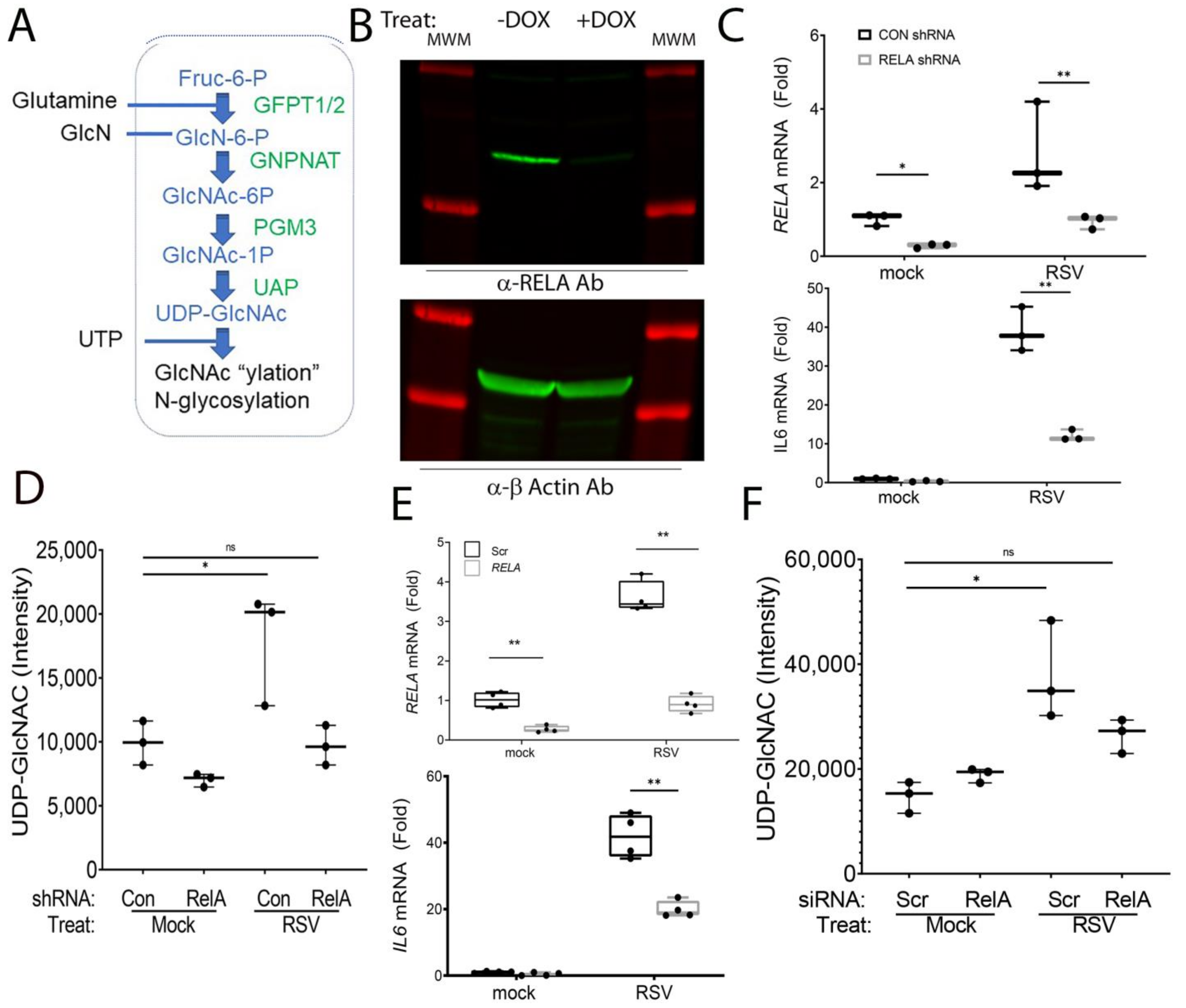
3.1. RSV Induces the HBP in an NFκB/RELA-Dependent Manner
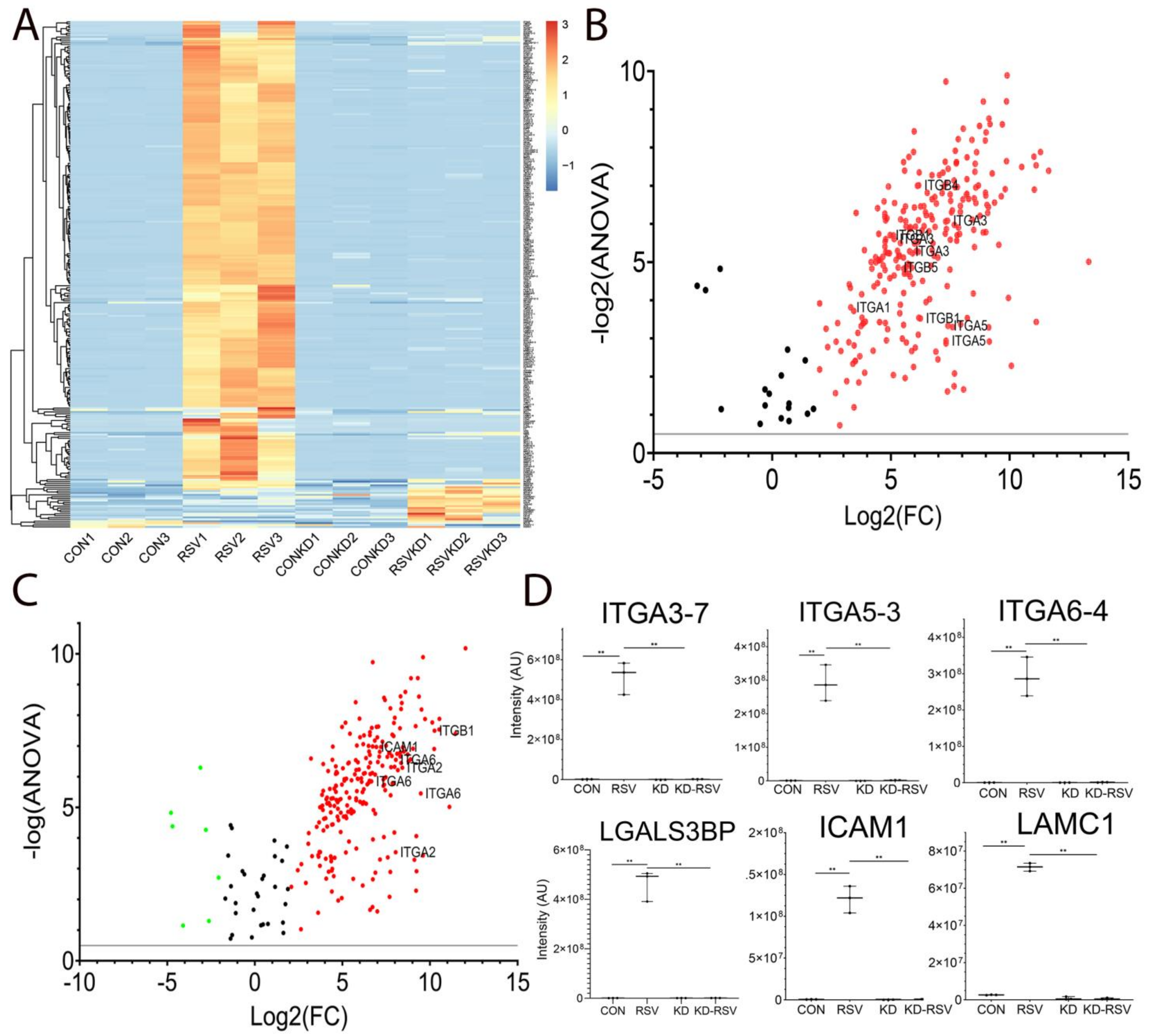
3.2. RELA Is Required for RSV-Induced N Glycoproteome Production
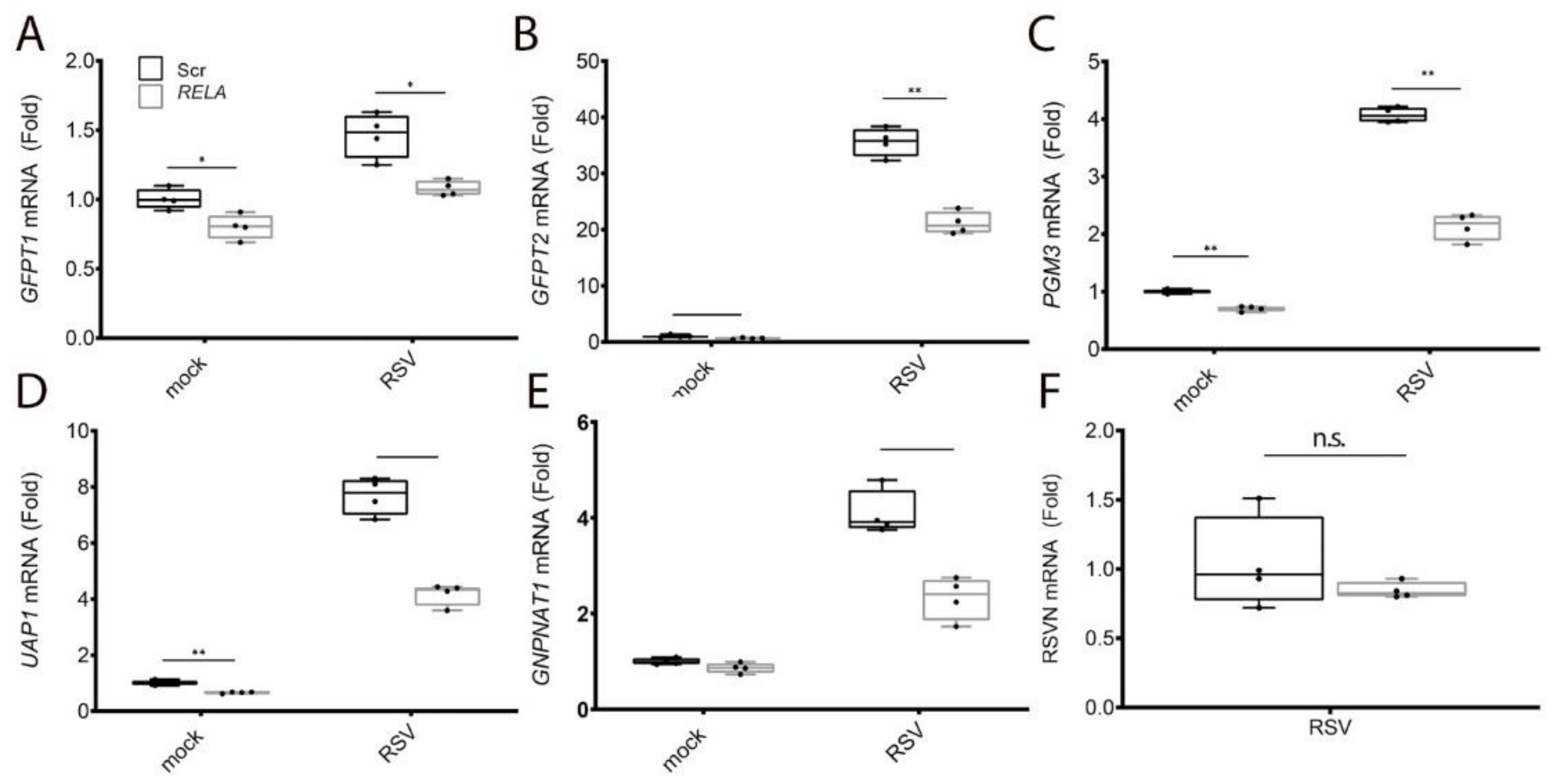
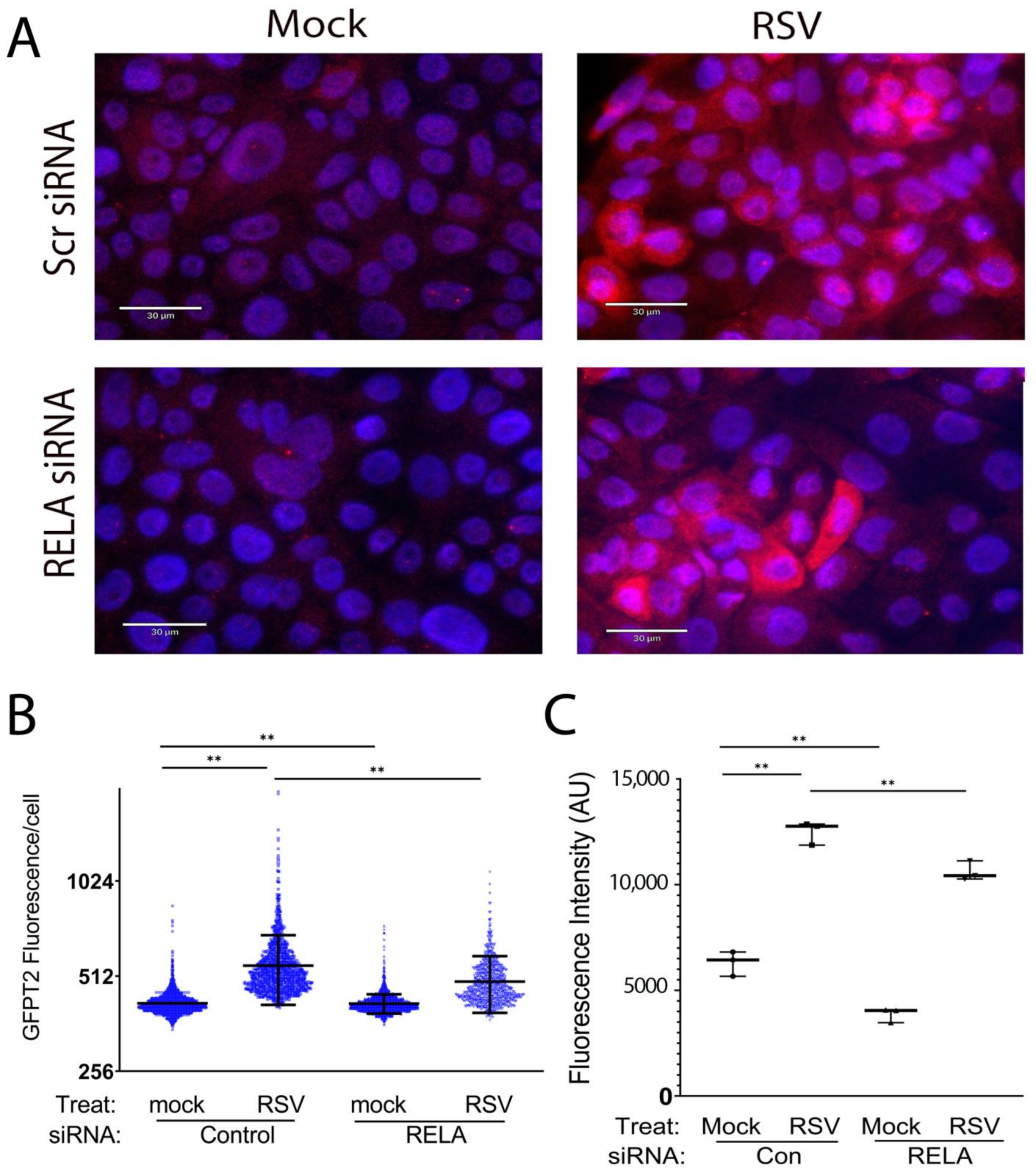
3.3. RELA Coordinately Regulates Core Enzymes of the HBP
3.4. RSV Induces RELA Binding to Open Chromatin in HBP Genes
3.5. HBP Pathway Genes Are OGG1-Dependent
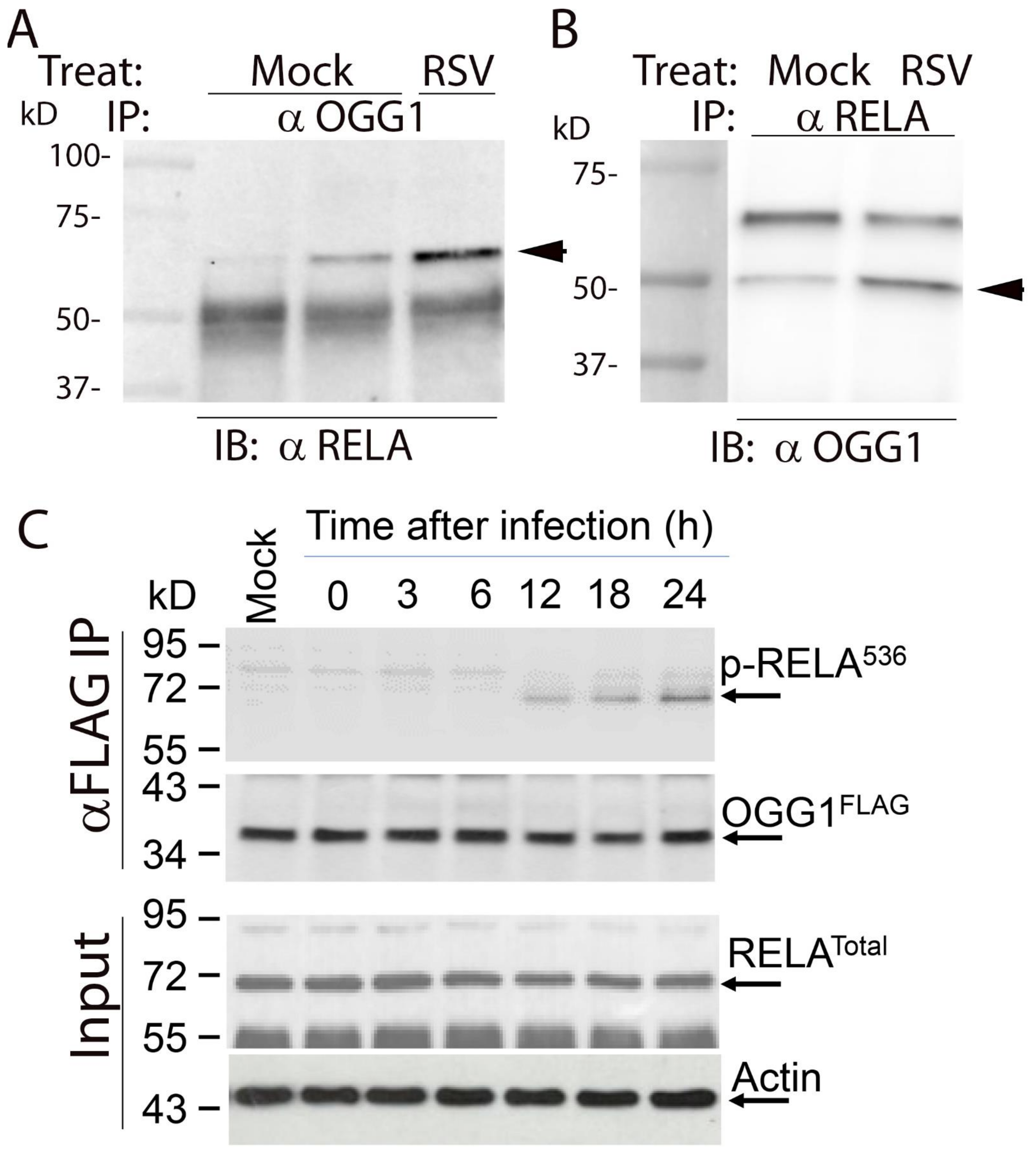
3.6. RSV Induces RELA∙OGG1 Complex Formation
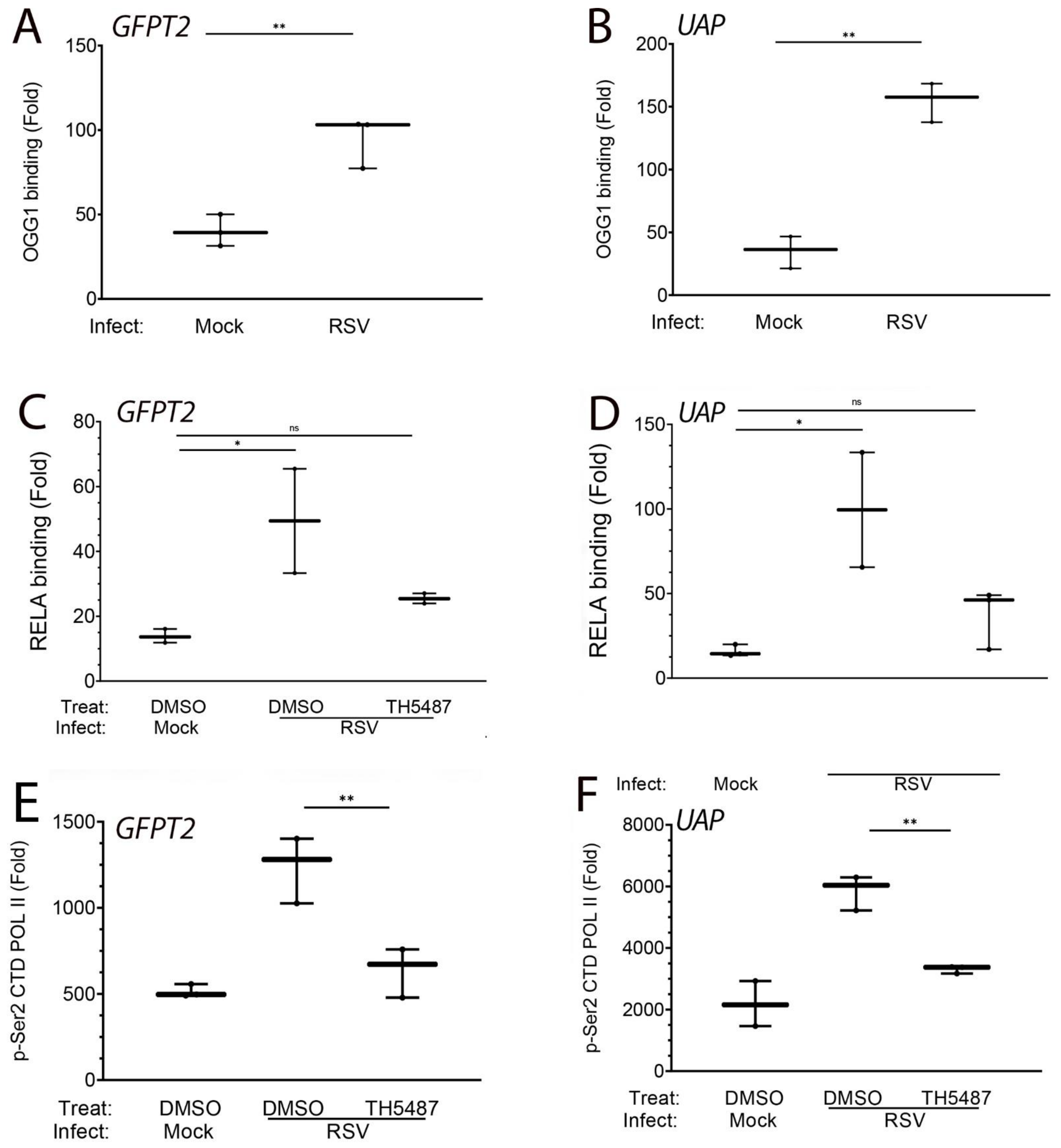
3.7. OGG1 Binds to the GFPT2 Intragenic Enhancer
3.8. RSV-Induced Recruitment of RELA Is OGG1-Dependent
3.9. OGG1 Is Required for Recruitment of Transcriptional Elongation-Competent Pol II
4. Discussion
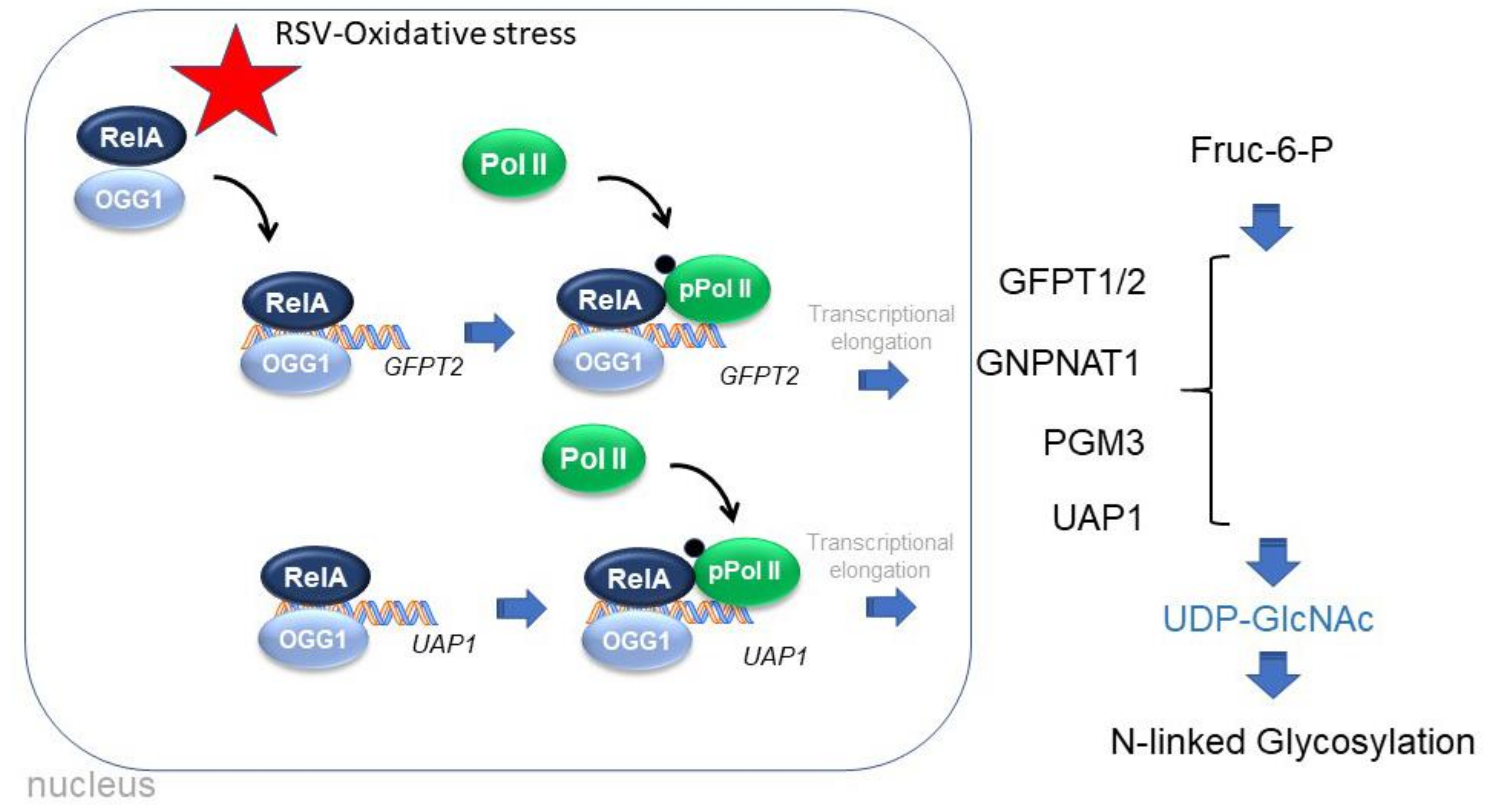
4.1. The Hexosamine Biosynthetic Pathway (HBP)
4.2. RELA-Dependent N-glycosylation of Integrin Signaling Pathway
4.3. RELA Is a Master Transcription Factor of Epithelial Plasticity Linking Innate Inflammation to Expression of EMT Regulators
4.4. OGG1-In Base Excision Repair and as an Epigenetic Regulator of HBP
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Borchers, A.T.; Chang, C.; Gershwin, M.E.; Gershwin, L.J. Respiratory syncytial virus--a comprehensive review. Clin. Rev. Allergy Immunol. 2013, 45, 331–379. [Google Scholar] [CrossRef] [PubMed]
- Stockman, L.J.; Curns, A.T.; Anderson, L.J.; Fischer-Langley, G. Respiratory syncytial virus-associated hospitalizations among infants and young children in the United States, 1997–2006. Pediatric Infect. Dis. J. 2012, 31, 5–9. [Google Scholar] [CrossRef] [PubMed]
- Shi, T.; McAllister, D.A.; O’Brien, K.L.; Simoes, E.A.F.; Madhi, S.A.; Gessner, B.D.; Polack, F.P.; Balsells, E.; Acacio, S.; Aguayo, C.; et al. Global, regional, and national disease burden estimates of acute lower respiratory infections due to respiratory syncytial virus in young children in 2015: A systematic review and modelling study. Lancet 2017, 390, 946–958. [Google Scholar] [CrossRef] [Green Version]
- Johnson, J.E.; Gonzales, R.A.; Olson, S.J.; Wright, P.F.; Graham, B.S. The histopathology of fatal untreated human respiratory syncytial virus infection. Mod. Pathol. 2007, 20, 108–119. [Google Scholar] [CrossRef]
- Martinez, F.D. The origins of asthma and chronic obstructive pulmonary disease in early life. Proc. Am. Thorac. Soc. 2009, 6, 272–277. [Google Scholar] [CrossRef] [Green Version]
- Fauroux, B.; Simoes, E.A.F.; Checchia, P.A.; Paes, B.; Figueras-Aloy, J.; Manzoni, P.; Bont, L.; Carbonell-Estrany, X. The Burden and Long-term Respiratory Morbidity Associated with Respiratory Syncytial Virus Infection in Early Childhood. Infect. Dis. Ther. 2017, 6, 173–197. [Google Scholar] [CrossRef] [Green Version]
- Falsey, A.R.; Walsh, E.E. Respiratory syncytial virus infection in adults. Clin. Microbiol. Rev. 2000, 13, 371–384. [Google Scholar] [CrossRef]
- Johnston, N.W.; Sears, M.R. Asthma exacerbations 1: Epidemiology. Thorax 2006, 61, 722–728. [Google Scholar] [CrossRef] [Green Version]
- Miller, M.K.; Lee, J.H.; Miller, D.P.; Wenzel, S.E.; Group, T.S. Recent asthma exacerbations: A key predictor of future exacerbations. Respir. Med. 2007, 101, 481–489. [Google Scholar] [CrossRef] [Green Version]
- Calhoun, W.J.; Haselkorn, T.; Miller, D.P.; Omachi, T.A. Asthma exacerbations and lung function in patients with severe or difficult-to-treat asthma. J. Allergy Clin. Immunol. 2015, 136, 1125–1127.e1124. [Google Scholar] [CrossRef]
- Bai, T.R.; Vonk, J.M.; Postma, D.S.; Boezen, H.M. Severe exacerbations predict excess lung function decline in asthma. Eur. Respir. J. 2007, 30, 452–456. [Google Scholar] [CrossRef] [Green Version]
- Jozwik, A.; Habibi, M.S.; Paras, A.; Zhu, J.; Guvenel, A.; Dhariwal, J.; Almond, M.; Wong, E.H.C.; Sykes, A.; Maybeno, M.; et al. RSV-specific airway resident memory CD8+ T cells and differential disease severity after experimental human infection. Nat. Commun. 2015, 6, 10224. [Google Scholar] [CrossRef]
- Tian, B.; Yang, J.; Zhao, Y.; Ivanciuc, T.; Sun, H.; Wakamiya, M.; Garofalo, R.P.; Brasier, A.R. Central Role of the NF-kappaB Pathway in the Scgb1a1-Expressing Epithelium in Mediating Respiratory Syncytial Virus-Induced Airway Inflammation. J. Virol. 2018, 92, e00441-18. [Google Scholar] [CrossRef] [Green Version]
- Xu, X.; Qiao, D.; Dong, C.; Mann, M.; Garofalo, R.P.; Keles, S.; Brasier, A.R. The SWI/SNF-Related, Matrix Associated, Actin-Dependent Regulator of Chromatin A4 Core Complex Represses Respiratory Syncytial Virus-Induced Syncytia Formation and Subepithelial Myofibroblast Transition. Front. Immunol. 2021, 12, 633654. [Google Scholar] [CrossRef]
- Xu, X.; Qiao, D.; Mann, M.; Garofalo, R.P.; Brasier, A.R. Respiratory Syncytial Virus Infection Induces Chromatin Remodeling to Activate Growth Factor and Extracellular Matrix Secretion Pathways. Viruses 2020, 12, 804. [Google Scholar] [CrossRef]
- Qiao, D.; Skibba, M.; Xu, X.; Garofalo, R.P.; Zhao, Y.; Brasier, A.R. Paramyxovirus replication induces the hexosamine biosynthetic pathway and mesenchymal transition via the IRE1alpha-XBP1s arm of the unfolded protein response. Am. J. Physiol. Lung Cell Mol. Physiol. 2021, 321, L576–L594. [Google Scholar] [CrossRef]
- Tian, B.; Yang, J.; Zhao, Y.; Ivanciuc, T.; Sun, H.; Garofalo, R.P.; Brasier, A.R. BRD4 Couples NF-kappaB/RelA with Airway Inflammation and the IRF-RIG-I Amplification Loop in Respiratory Syncytial Virus Infection. J. Virol. 2017, 91, e00007-17. [Google Scholar] [CrossRef] [Green Version]
- Jamaluddin, M.; Wiktorowicz, J.E.; Soman, K.V.; Boldogh, I.; Forbus, J.D.; Spratt, H.; Garofalo, R.P.; Brasier, A.R. Role of peroxiredoxin 1 and peroxiredoxin 4 in protection of respiratory syncytial virus-induced cysteinyl oxidation of nuclear cytoskeletal proteins. J. Virol. 2010, 84, 9533–9545. [Google Scholar] [CrossRef] [Green Version]
- Garofalo, R.P.; Kolli, D.; Casola, A. Respiratory syncytial virus infection: Mechanisms of redox control and novel therapeutic opportunities. Antioxid. Redox Signal 2013, 18, 186–217. [Google Scholar] [CrossRef] [Green Version]
- Svilar, D.; Goellner, E.M.; Almeida, K.H.; Sobol, R.W. Base excision repair and lesion-dependent subpathways for repair of oxidative DNA damage. Antioxid. Redox Signal 2011, 14, 2491–2507. [Google Scholar] [CrossRef] [Green Version]
- Hao, W.; Qi, T.; Pan, L.; Wang, R.; Zhu, B.; Aguilera-Aguirre, L.; Radak, Z.; Hazra, T.K.; Vlahopoulos, S.A.; Bacsi, A.; et al. Effects of the stimuli-dependent enrichment of 8-oxoguanine DNA glycosylase1 on chromatinized DNA. Redox Biol. 2018, 18, 43–53. [Google Scholar] [CrossRef]
- Choudhary, S.; Boldogh, I.; Brasier, A.R. Inside-Out Signaling Pathways from Nuclear Reactive Oxygen Species Control Pulmonary Innate Immunity. J. Innate Immun. 2016, 8, 143–155. [Google Scholar] [CrossRef]
- Zheng, X.; Wang, K.; Pan, L.; Hao, W.; Xue, Y.; Bacsi, A.; Vlahopoulos, S.A.; Radak, Z.; Hazra, T.K.; Brasier, A.R.; et al. Innate Immune Responses to RSV Infection Facilitated by OGG1, an Enzyme Repairing Oxidatively Modified DNA Base Lesions. J. Innate Immun. 2022, 10, 1–22. [Google Scholar] [CrossRef]
- Visnes, T.; Cázares-Körner, A.; Hao, W.; Wallner, O.; Masuyer, G.; Loseva, O.; Mortusewicz, O.; Wiita, E.; Sarno, A.; Manoilov, A.; et al. Small-molecule inhibitor of OGG1 suppresses proinflammatory gene expression and inflammation. Science 2018, 362, 834–839. [Google Scholar] [CrossRef] [Green Version]
- Morris, D.R.; Qu, Y.; Agrawal, A.; Garofalo, R.P.; Casola, A. HIF-1α Modulates Core Metabolism and Virus Replication in Primary Airway Epithelial Cells Infected with Respiratory Syncytial Virus. Viruses 2020, 12, 1088. [Google Scholar] [CrossRef]
- Connelly, A.R.; Jeong, B.M.; Coden, M.E.; Cao, J.Y.; Chirkova, T.; Rosas-Salazar, C.; Cephus, J.Y.; Anderson, L.J.; Newcomb, D.C.; Hartert, T.V.; et al. Metabolic Reprogramming of Nasal Airway Epithelial Cells Following Infant Respiratory Syncytial Virus Infection. Viruses 2021, 13, 2055. [Google Scholar] [CrossRef]
- Bond, M.R.; Hanover, J.A. A little sugar goes a long way: The cell biology of O-GlcNAc. J. Cell Biol. 2015, 208, 869–880. [Google Scholar] [CrossRef] [Green Version]
- Tian, B.; Widen, S.G.; Yang, J.; Wood, T.G.; Kudlicki, A.; Zhao, Y.; Brasier, A.R. The NFkappaB subunit RELA is a master transcriptional regulator of the committed epithelial-mesenchymal transition in airway epithelial cells. J. Biol. Chem. 2018, 293, 16528–16545. [Google Scholar] [CrossRef] [Green Version]
- Zhao, Y.; Jamaluddin, M.; Zhang, Y.; Sun, H.; Ivanciuc, T.; Garofalo, R.P.; Brasier, A.R. Systematic Analysis of Cell-Type Differences in the Epithelial Secretome Reveals Insights into the Pathogenesis of Respiratory Syncytial Virus-Induced Lower Respiratory Tract Infections. J. Immunol. 2017, 198, 3345–3364. [Google Scholar] [CrossRef] [Green Version]
- Garofalo, R.; Sabry, M.; Jamaluddin, M.; Yu, R.K.; Casola, A.; Ogra, P.L.; Brasier, A.R. Transcriptional activation of the interleukin-8 gene by respiratory syncytial virus infection in alveolar epithelial cells: Nuclear translocation of the RelA transcription factor as a mechanism producing airway mucosal inflammation. J. Virol. 1996, 70, 8773–8781. [Google Scholar] [CrossRef] [Green Version]
- Zhang, J.; Jamaluddin, M.; Zhang, Y.; Widen, S.G.; Sun, H.; Brasier, A.R.; Zhao, Y. Type II Epithelial-Mesenchymal Transition Upregulates Protein N-Glycosylation To Maintain Proteostasis and Extracellular Matrix Production. J. Proteome Res. 2019, 18, 3447–3460. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Y.; Zhang, J.; Sun, H.; Brasier, A.R. Crosstalk of the IkappaB Kinase with Spliced X-Box Binding Protein 1 Couples Inflammation with Glucose Metabolic Reprogramming in Epithelial-Mesenchymal Transition. J. Proteome Res. 2021, 20, 3475–3488. [Google Scholar] [CrossRef] [PubMed]
- Wisniewski, J.R.; Zielinska, D.F.; Mann, M. Comparison of ultrafiltration units for proteomic and N-glycoproteomic analysis by the filter-aided sample preparation method. Anal. Biochem. 2011, 410, 307–309. [Google Scholar] [CrossRef] [PubMed]
- Cox, J.; Mann, M. MaxQuant enables high peptide identification rates, individualized p.p.b.-range mass accuracies and proteome-wide protein quantification. Nat. Biotechnol. 2008, 26, 1367–1372. [Google Scholar] [CrossRef]
- Cox, J.; Hein, M.Y.; Luber, C.A.; Paron, I.; Nagaraj, N.; Mann, M. Accurate proteome-wide label-free quantification by delayed normalization and maximal peptide ratio extraction, termed MaxLFQ. Mol. Cell Proteom. 2014, 13, 2513–2526. [Google Scholar] [CrossRef] [Green Version]
- Tyanova, S.; Temu, T.; Sinitcyn, P.; Carlson, A.; Hein, M.Y.; Geiger, T.; Mann, M.; Cox, J. The Perseus computational platform for comprehensive analysis of (prote)omics data. Nat. Methods 2016, 13, 731–740. [Google Scholar] [CrossRef]
- Pan, L.; Hao, W.; Zheng, X.; Zeng, X.; Ahmed Abbasi, A.; Boldogh, I.; Ba, X. OGG1-DNA interactions facilitate NF-κB binding to DNA targets. Sci. Rep. 2017, 7, 43297. [Google Scholar] [CrossRef] [Green Version]
- Nowak, D.E.; Tian, B.; Brasier, A.R. Two-step cross-linking method for identification of NF-kappaB gene network by chromatin immunoprecipitation. BioTechniques 2005, 39, 715–725. [Google Scholar] [CrossRef]
- Tian, B.; Yang, J.; Brasier, A.R. Two-step cross-linking for analysis of protein-chromatin interactions. Methods Mol. Biol. 2012, 809, 105–120. [Google Scholar] [CrossRef] [Green Version]
- Hoffmann, A.; Leung, T.H.; Baltimore, D. Genetic analysis of NF-kappaB/Rel transcription factors defines functional specificities. Embo j. 2003, 22, 5530–5539. [Google Scholar] [CrossRef]
- Wiśniewski, J.R.; Zougman, A.; Nagaraj, N.; Mann, M. Universal sample preparation method for proteome analysis. Nat. Methods 2009, 6, 359–362. [Google Scholar] [CrossRef]
- Ba, X.; Boldogh, I. 8-Oxoguanine DNA glycosylase 1: Beyond repair of the oxidatively modified base lesions. Redox Biol. 2018, 14, 669–678. [Google Scholar] [CrossRef]
- Lia, D.; Reyes, A.; de Melo Campos, J.T.A.; Piolot, T.; Baijer, J.; Radicella, J.P.; Campalans, A. Mitochondrial maintenance under oxidative stress depends on mitochondrially localised α-OGG1. J. Cell Sci. 2018, 131, 100. [Google Scholar] [CrossRef] [Green Version]
- Aherne, W.; Bird, T.; Court, S.D.; Gardner, P.S.; McQuillin, J. Pathological changes in virus infections of the lower respiratory tract in children. J. Clin. Pathol. 1970, 23, 7–18. [Google Scholar] [CrossRef] [Green Version]
- Tian, B.; Patrikeev, I.; Ochoa, L.; Vargas, G.; Belanger, K.K.; Litvinov, J.; Boldogh, I.; Ameredes, B.T.; Motamedi, M.; Brasier, A.R. NF-kappaB Mediates Mesenchymal Transition, Remodeling, and Pulmonary Fibrosis in Response to Chronic Inflammation by Viral RNA Patterns. Am. J. Respir. Cell Mol. Biol. 2017, 56, 506–520. [Google Scholar] [CrossRef] [Green Version]
- Vander Heiden, M.G.; Cantley, L.C.; Thompson, C.B. Understanding the Warburg effect: The metabolic requirements of cell proliferation. Science 2009, 324, 1029–1033. [Google Scholar] [CrossRef] [Green Version]
- Moremen, K.W.; Tiemeyer, M.; Nairn, A.V. Vertebrate protein glycosylation: Diversity, synthesis and function. Nat. Rev. Mol. Cell Biol. 2012, 13, 448–462. [Google Scholar] [CrossRef] [Green Version]
- Kroef, V.; Ruegenberg, S.; Horn, M.; Allmeroth, K.; Ebert, L.; Bozkus, S.; Miethe, S.; Elling, U.; Schermer, B.; Baumann, U.; et al. GFPT2/GFAT2 and AMDHD2 act in tandem to control the hexosamine pathway. eLife 2022, 11, e69223. [Google Scholar] [CrossRef]
- Cai, X.; Thinn, A.M.M.; Wang, Z.; Shan, H.; Zhu, J. The importance of N-glycosylation on β3 integrin ligand binding and conformational regulation. Sci. Rep. 2017, 7, 4656. [Google Scholar] [CrossRef] [Green Version]
- Ohtsubo, K.; Marth, J.D. Glycosylation in Cellular Mechanisms of Health and Disease. Cell 2006, 126, 855–867. [Google Scholar] [CrossRef] [Green Version]
- Creyghton, M.P.; Cheng, A.W.; Welstead, G.G.; Kooistra, T.; Carey, B.W.; Steine, E.J.; Hanna, J.; Lodato, M.A.; Frampton, G.M.; Sharp, P.A.; et al. Histone H3K27ac separates active from poised enhancers and predicts developmental state. Proc. Natl. Acad. Sci. USA 2010, 107, 21931–21936. [Google Scholar] [CrossRef] [Green Version]
- Brasier, A.R. RSV Reprograms the CDK9*BRD4 Chromatin Remodeling Complex to Couple Innate Inflammation to Airway Remodeling. Viruses 2020, 12, 472. [Google Scholar] [CrossRef] [Green Version]
- Ijaz, T.; Jamaluddin, M.; Zhao, Y.; Zhang, Y.; Jay, J.; Finnerty, C.C.; Herndon, D.N.; Tilton, R.G.; Brasier, A.R. Coordinate activities of BRD4 and CDK9 in the transcriptional elongation complex are required for TGFbeta-induced Nox4 expression and myofibroblast transdifferentiation. Cell Death Dis. 2017, 8, e2606. [Google Scholar] [CrossRef] [Green Version]
- Tian, B.; Zhao, Y.; Sun, H.; Zhang, Y.; Yang, J.; Brasier, A.R. BRD4 mediates NF-kappaB-dependent epithelial-mesenchymal transition and pulmonary fibrosis via transcriptional elongation. Am. J. Physiol. Lung Cell Mol. Physiol. 2016, 311, L1183–L1201. [Google Scholar] [CrossRef]
- Kalita, M.; Tian, B.; Gao, B.; Choudhary, S.; Wood, T.G.; Carmical, J.R.; Boldogh, I.; Mitra, S.; Minna, J.D.; Brasier, A.R. Systems approaches to modeling chronic mucosal inflammation. BioMed Res. Int 2013, 2013, 505864. [Google Scholar] [CrossRef]
- Bertolusso, R.; Tian, B.; Zhao, Y.; Vergara, L.; Sabree, A.; Iwanaszko, M.; Lipniacki, T.; Brasier, A.R.; Kimmel, M. Dynamic cross talk model of the epithelial innate immune response to double-stranded RNA stimulation: Coordinated dynamics emerging from cell-level noise. PLoS One 2014, 9, e93396. [Google Scholar] [CrossRef] [Green Version]
- Mitra, S.; Izumi, T.; Boldogh, I.; Bhakat, K.K.; Hill, J.W.; Hazra, T.K. Choreography of oxidative damage repair in mammalian genomes. Free Radic. Biol. Med. 2002, 33, 15–28. [Google Scholar] [CrossRef]
- Castro, S.M.; Guerrero-Plata, A.; Suarez-Real, G.; Adegboyega, P.A.; Colasurdo, G.N.; Khan, A.M.; Garofalo, R.P.; Casola, A. Antioxidant treatment ameliorates respiratory syncytial virus-induced disease and lung inflammation. Am. J. Respir. Crit. Care Med. 2006, 174, 1361–1369. [Google Scholar] [CrossRef] [Green Version]
- Fleming, A.M.; Ding, Y.; Burrows, C.J. Oxidative DNA damage is epigenetic by regulating gene transcription via base excision repair. Proc. Natl. Acad. Sci. USA 2017, 114, 2604–2609. [Google Scholar] [CrossRef]
- Pan, L.; Zhu, B.; Hao, W.; Zeng, X.; Vlahopoulos, S.A.; Hazra, T.K.; Hegde, M.L.; Radak, Z.; Bacsi, A.; Brasier, A.R.; et al. Oxidized Guanine Base Lesions Function in 8-Oxoguanine DNA Glycosylase-1-mediated Epigenetic Regulation of Nuclear Factor kappaB-driven Gene Expression. J. Biol. Chem. 2016, 291, 25553–25566. [Google Scholar] [CrossRef] [Green Version]
- Albensi, B.C. What Is Nuclear Factor Kappa B (NF-kappaB) Doing in and to the Mitochondrion? Front. Cell Dev. Biol. 2019, 7, 154. [Google Scholar] [CrossRef] [PubMed]
- Sasaki, C.Y.; Barberi, T.J.; Ghosh, P.; Longo, D.L. Phosphorylation of RelA/p65 on serine 536 defines an IkBa-independent NF-kB pathway. J. Biol. Chem. 2005, 280, 34538–34547. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cui, R.; Tieu, B.; Recinos, A.; Tilton, R.G.; Brasier, A.R. RhoA Mediates Angiotensin II Induced Phospho-Ser536 Nuclear Factor -kB/RelA Subunit Exchange on the Interleukin-6 Promoter in VSMCs. Circ. Res. 2006, 99, 723–730. [Google Scholar] [CrossRef] [PubMed] [Green Version]












{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Forward | Reverse |
|---|---|---|
| GFPT2 | 5′-GGGCATCCTGAGCGTGATTC-3′ | 5′-CCATGTAGCATCCCTGCTGT-3′ |
| GNPNAT1 | 5′-ATCCTGGAGAAGGCTTGGTT-3′ | 5′-CAGAGTTGCCGTAGCAACAA-3′ |
| PGM3 | 5′-TCATGTTTCGCATGGGATTA-3′ | 5′-AAACAGGTGGCATGTTCCTC-3′ |
| UAP1 | 5′-GAGGCATTTGGAGCATTCAT-3′ | 5′-TCCGTCTGAGCTTCGTTTTT-3′ |
| OGT | 5′-TGTGGCAGCTTATCTTCGTG-3′ | 5′-GAGAGCATTGGCTAGGTTGC-3′ |
| MGAT1 | 5′-GGTGGAGAAAGTGAGGACCA-3′ | 5′-CGGAACTGGAAGGTGACAAT-3′ |
| OGG1 | 5′-GCAGCAGCTACGAGAGTCCT-3′ | 5′-TTCCCAGTTCCTTGTTGGTC-3′ |
| Genic Region | Forward | Reverse |
|---|---|---|
| GFPT2 enhancer | 5′-GGAGTTGGGACGGAAAGTCA-3′ | 5′-GAAGCTCACCCTTGCCACTA-3′ |
| GNPNAT1 promoter | 5′-GGGGTAGGAGCCTAGGAAAA-3′ | 5′-GCGTGGGAAATGAGACAGTT-3′ |
| PGM3 promoter | 5′-GCCTAGGTCCACGTACCAGA-3′ | 5′-CTCGGAGTTGAGAAGGGAGA-3′ |
| UAP1 promoter | 5′-AGTGGGACAGGAGATCGTTG-3′ | 5′-AGAGAGGGGAAACCCAGAAA-3′ |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Xu, X.; Qiao, D.; Pan, L.; Boldogh, I.; Zhao, Y.; Brasier, A.R. RELA∙8-Oxoguanine DNA Glycosylase1 Is an Epigenetic Regulatory Complex Coordinating the Hexosamine Biosynthetic Pathway in RSV Infection. Cells 2022, 11, 2210. https://doi.org/10.3390/cells11142210
Xu X, Qiao D, Pan L, Boldogh I, Zhao Y, Brasier AR. RELA∙8-Oxoguanine DNA Glycosylase1 Is an Epigenetic Regulatory Complex Coordinating the Hexosamine Biosynthetic Pathway in RSV Infection. Cells. 2022; 11(14):2210. https://doi.org/10.3390/cells11142210
Chicago/Turabian StyleXu, Xiaofang, Dianhua Qiao, Lang Pan, Istvan Boldogh, Yingxin Zhao, and Allan R. Brasier. 2022. "RELA∙8-Oxoguanine DNA Glycosylase1 Is an Epigenetic Regulatory Complex Coordinating the Hexosamine Biosynthetic Pathway in RSV Infection" Cells 11, no. 14: 2210. https://doi.org/10.3390/cells11142210