
Forcing the Antitumor Effects of HSPs Using a Modulated Electric Field


Abstract
:1. Heat Shock Proteins
HSPs: Friend or Foe in Oncology
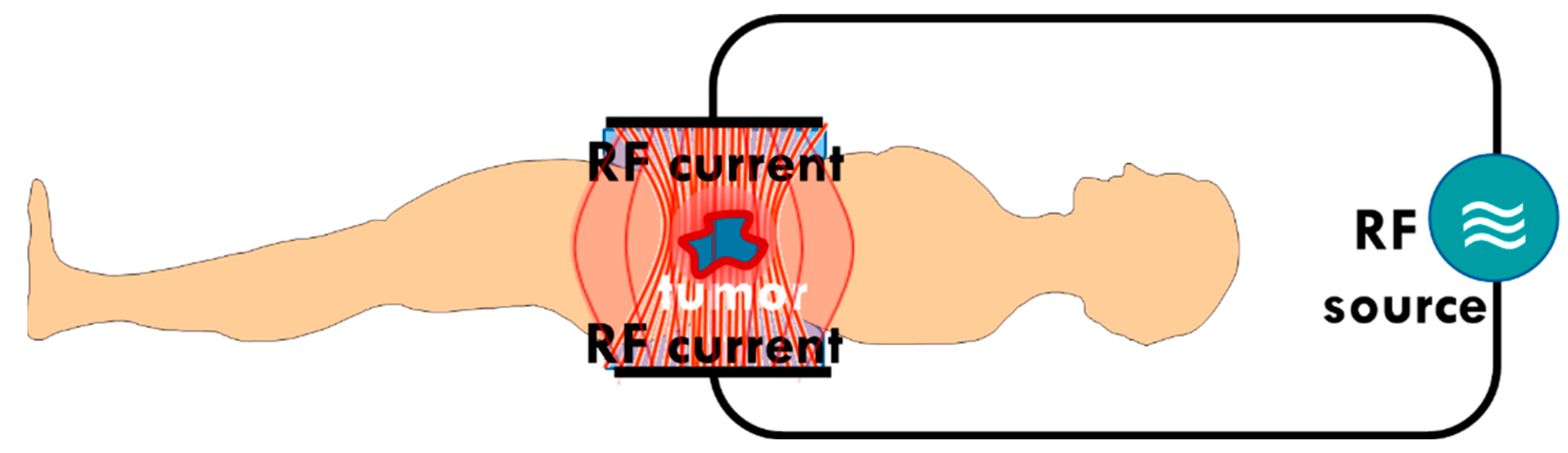
2. Electromagnetic Method of Hyperthermia in Oncology
- One way is to heat the whole tumor volume isothermally. The applied focusing techniques intend to maximize the temperature of the tumor volume and minimize it in healthy surroundings [70]. The original heating goal is necrosis. The applied dose compares the actual cellular distortion to the necrosis achieved in vitro at 43 °C [71];
- Another way heats small selected targets, either artificially or naturally available centers in the tumor volume:
- –
- –
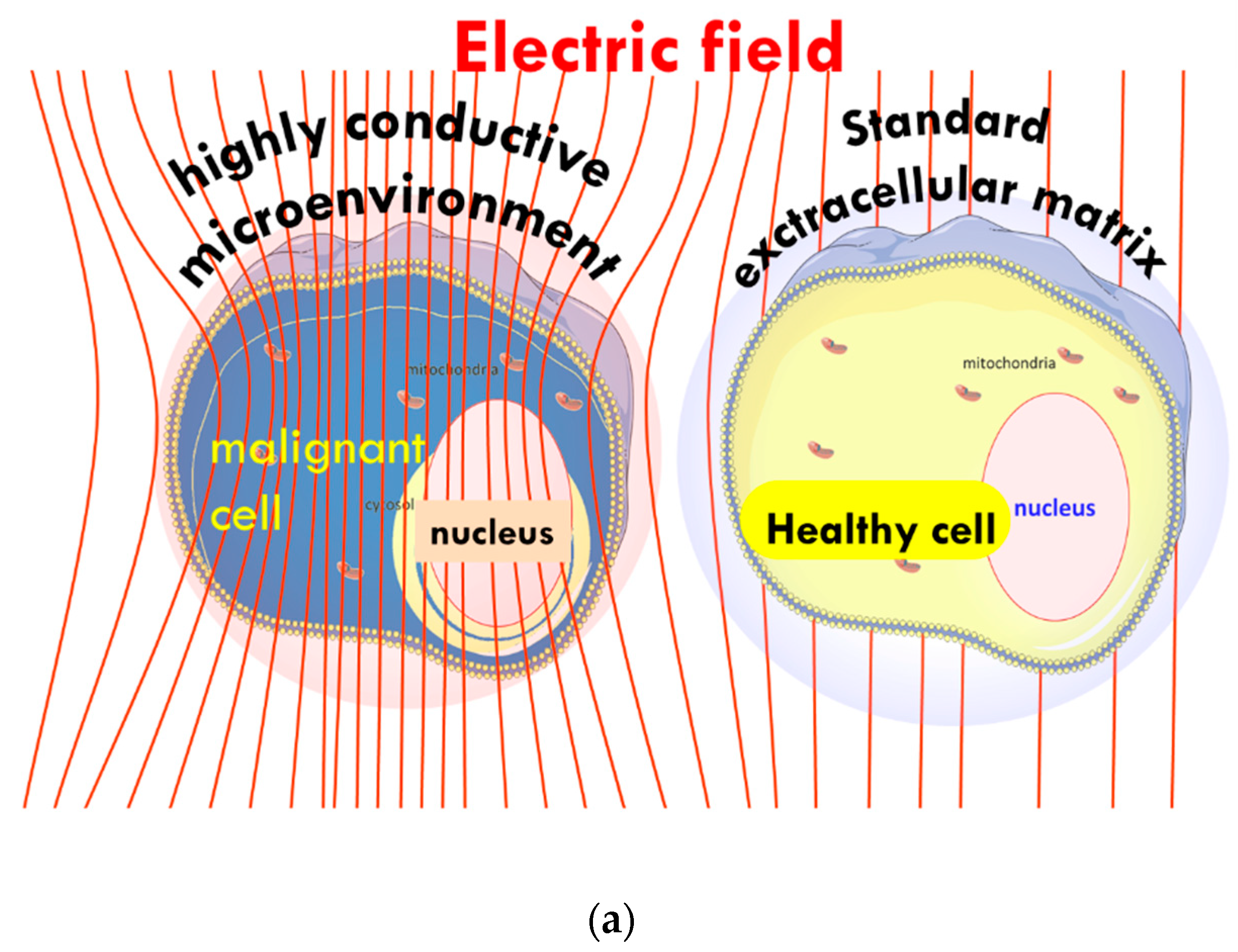
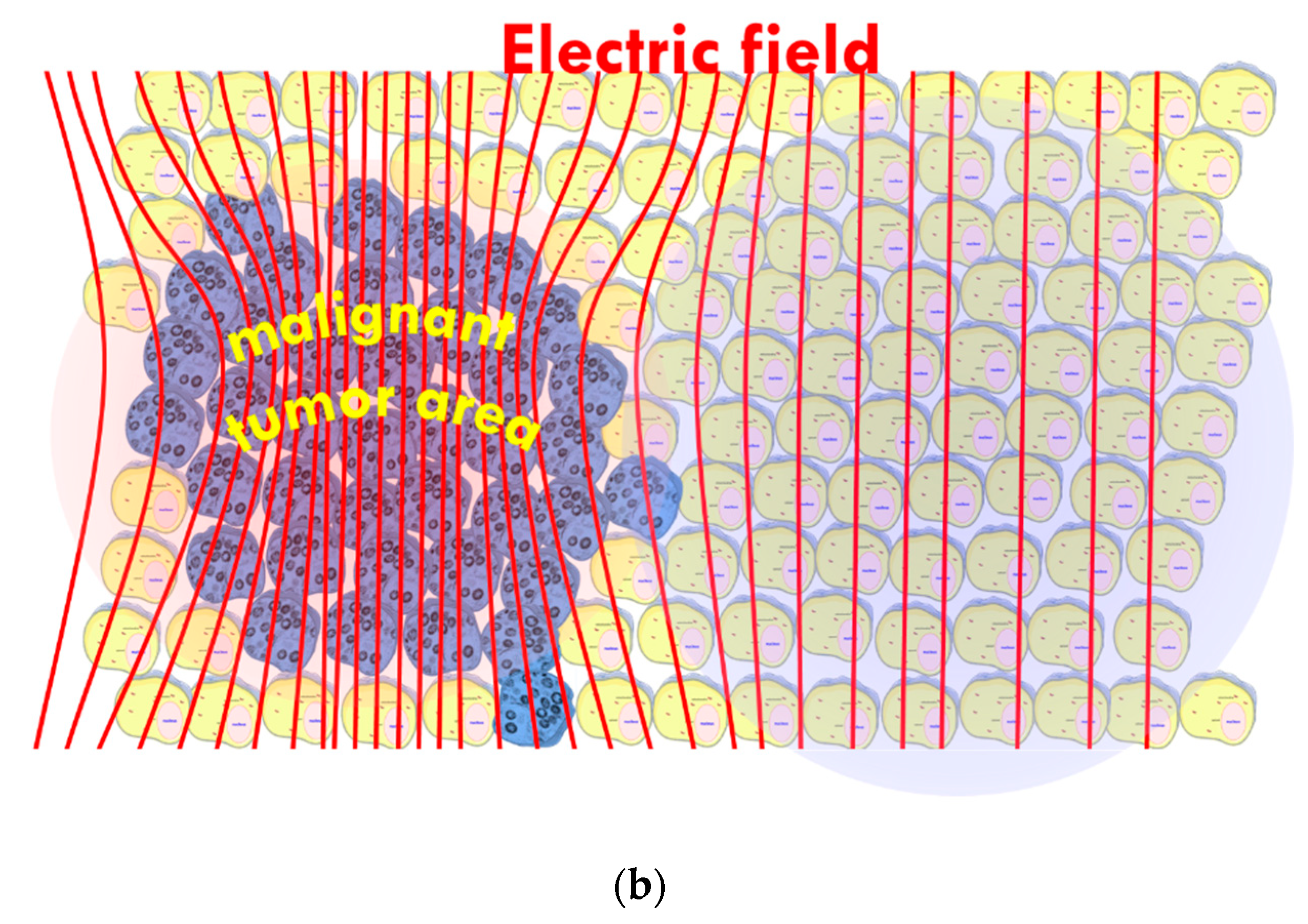
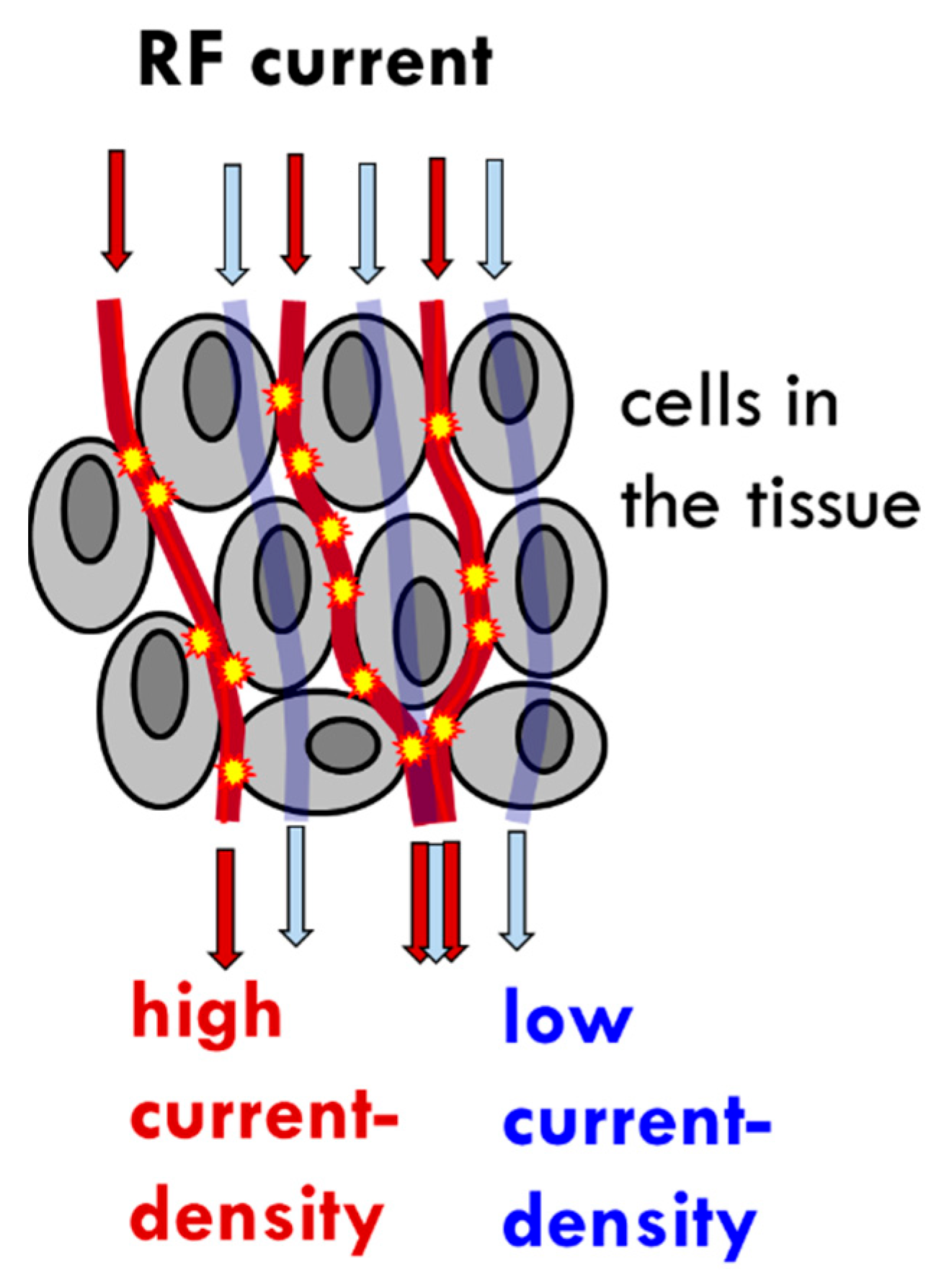
- The tumor’s heterogenic structure offers natural targets. The optimally chosen electromagnetic properties such as the frequency, the intensity, the phase, and the delivered time–information (modulation) allow for tuning to select the chosen particle (molecular cluster). For example, the membrane rafts (lipid micro-domains found in the plasma membranes of cells [74]) offer the perfect opportunity. The high electromagnetic contrast allows selection between the lipid-supported transmembrane proteins (membrane rafts) and their pure double lipid holding membrane material.
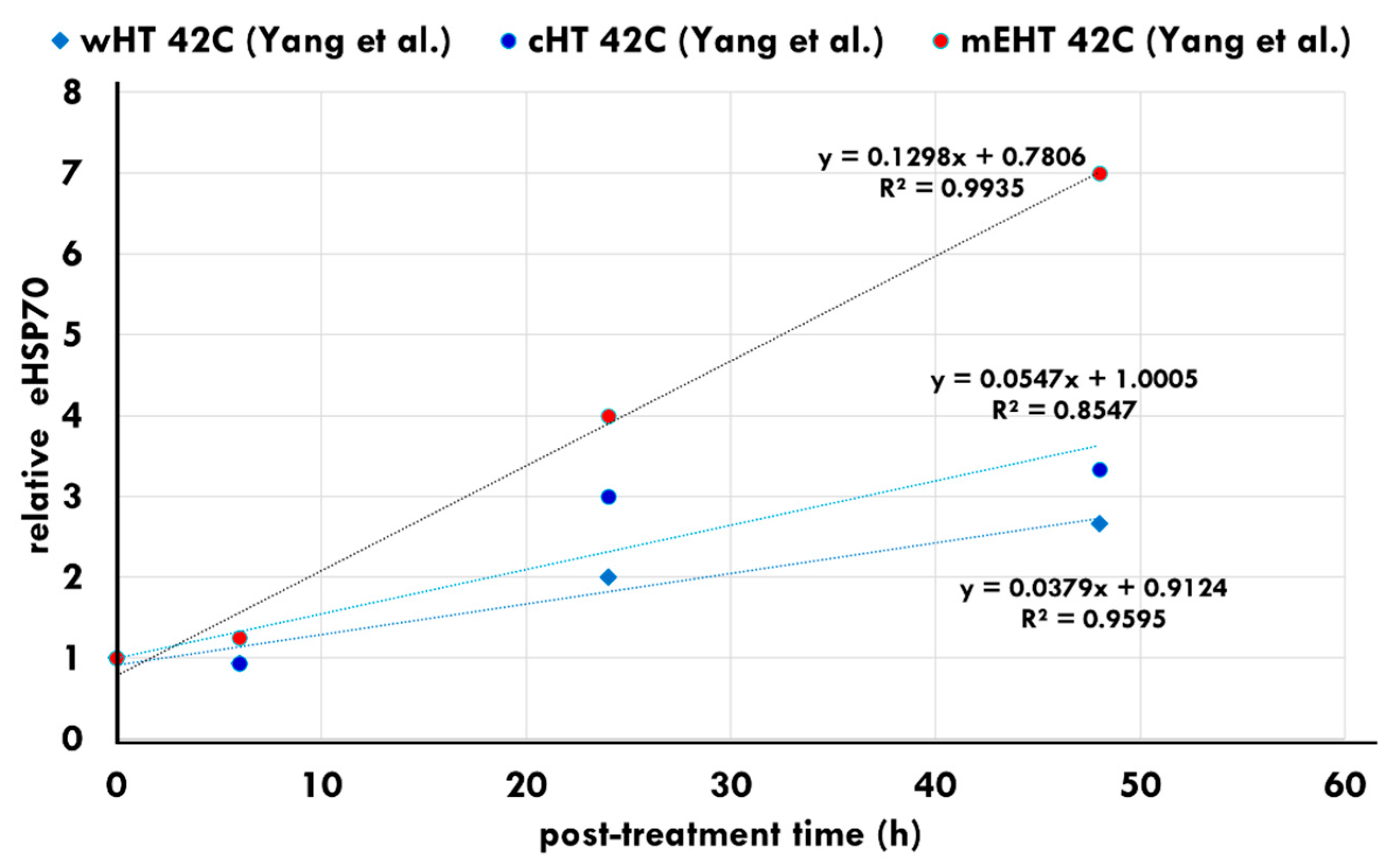
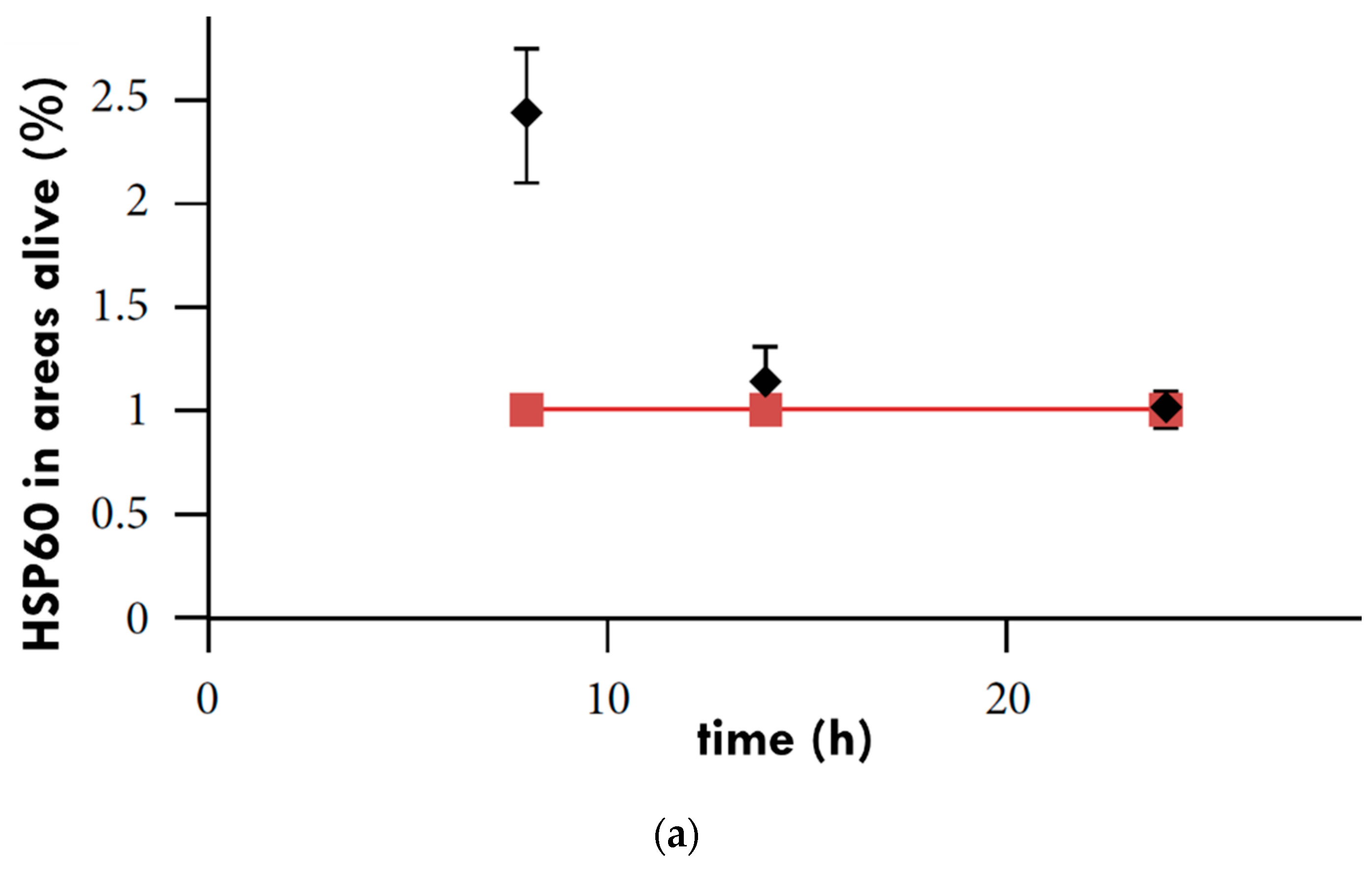
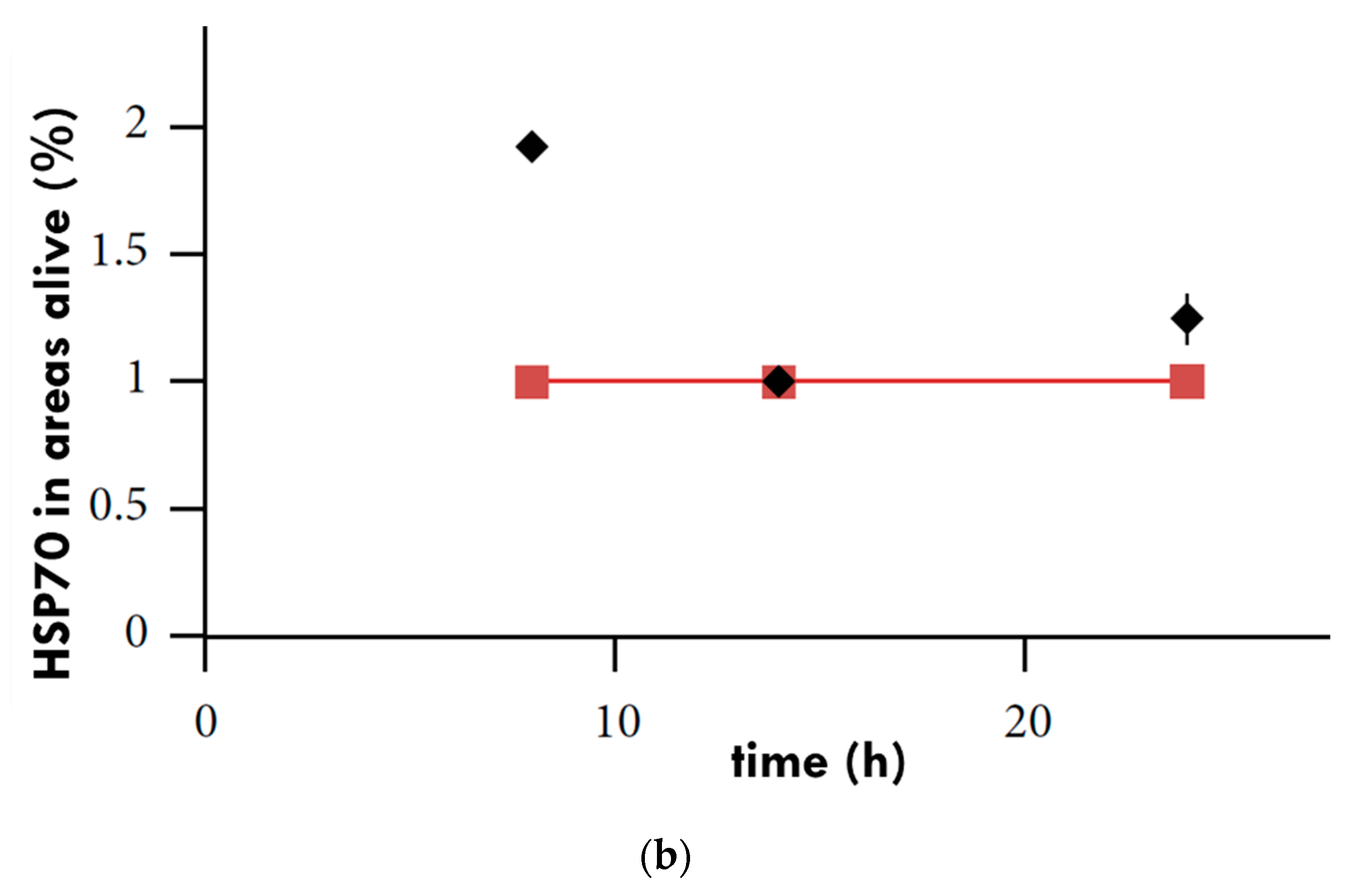
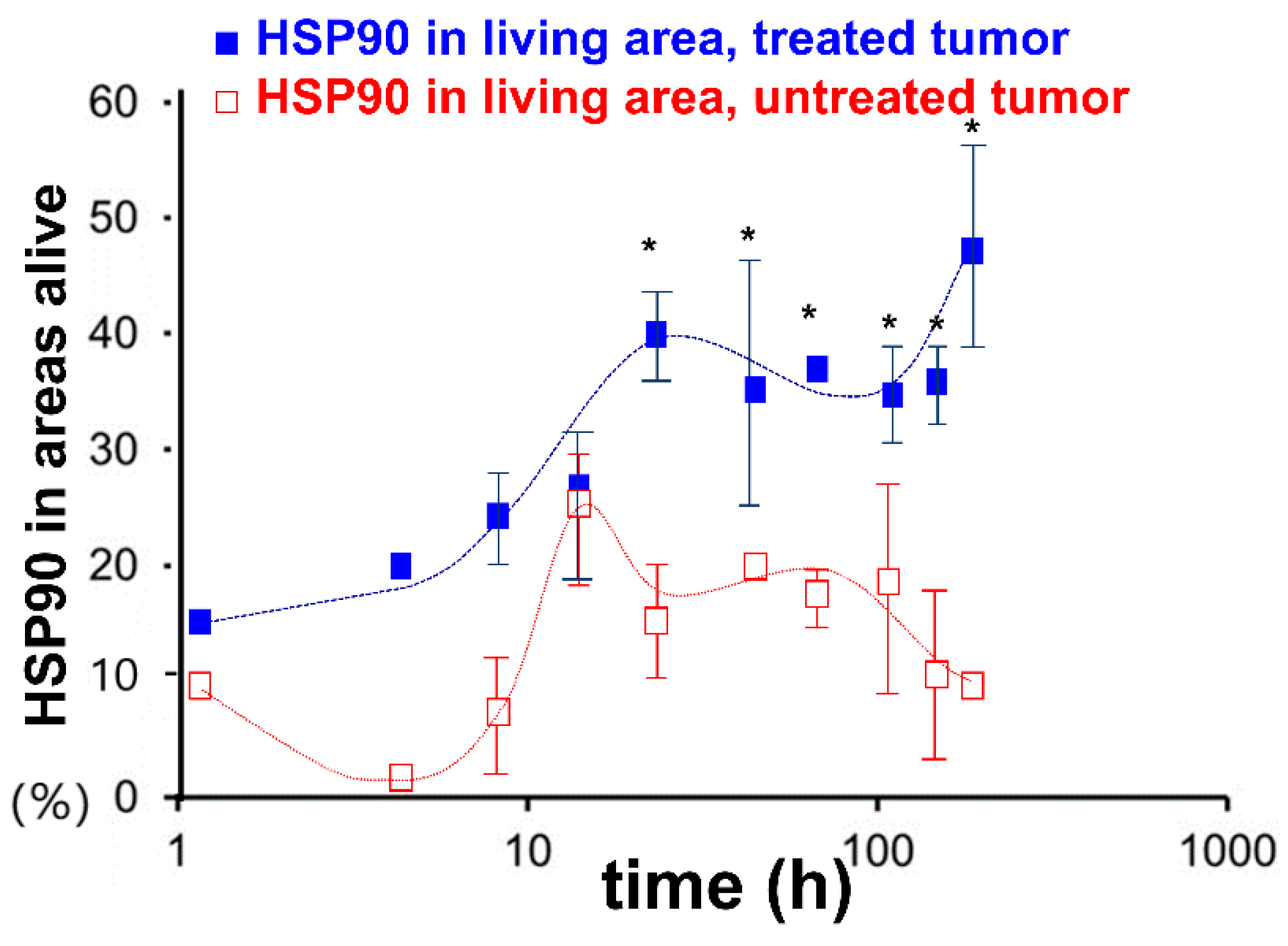
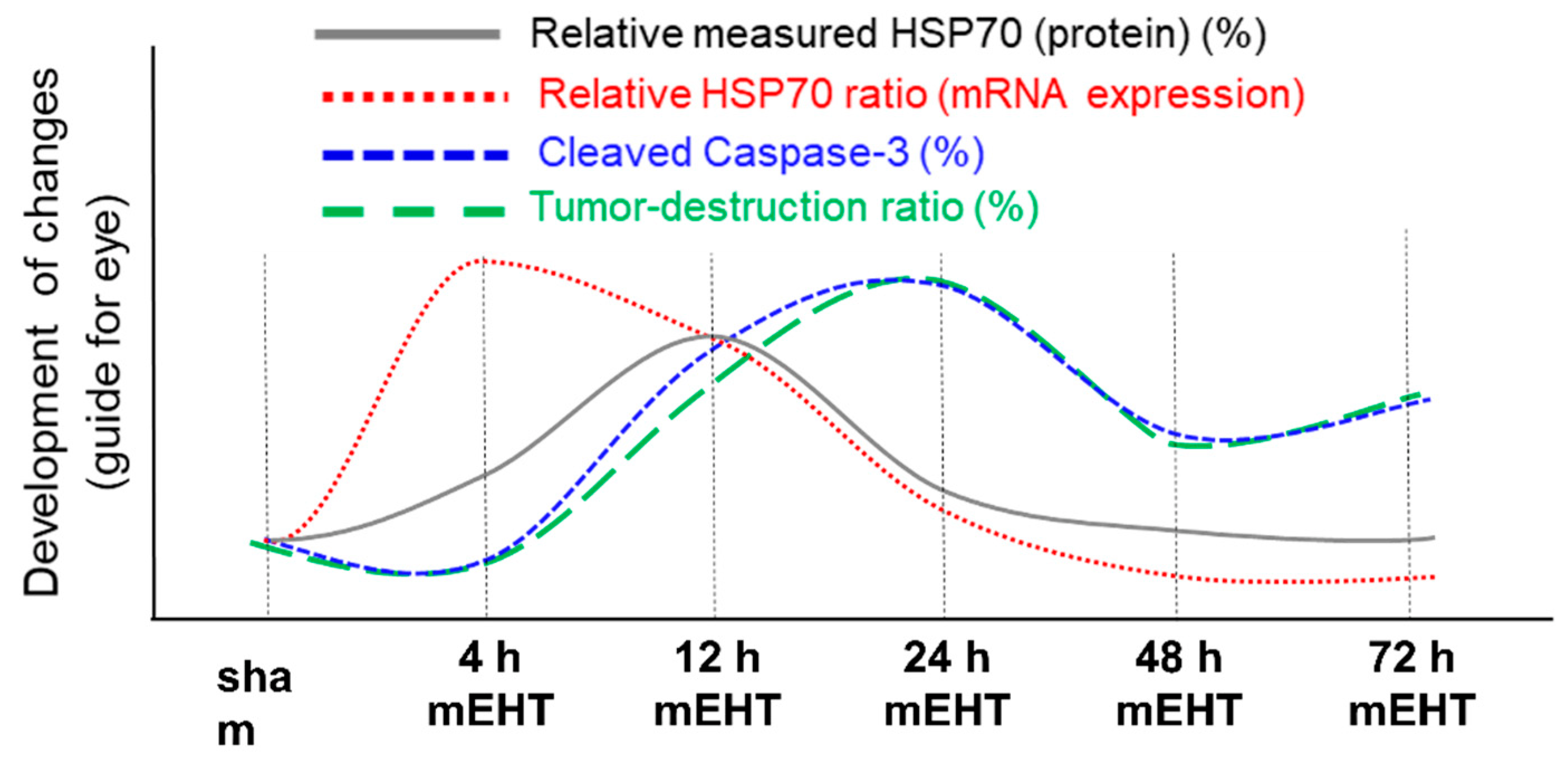
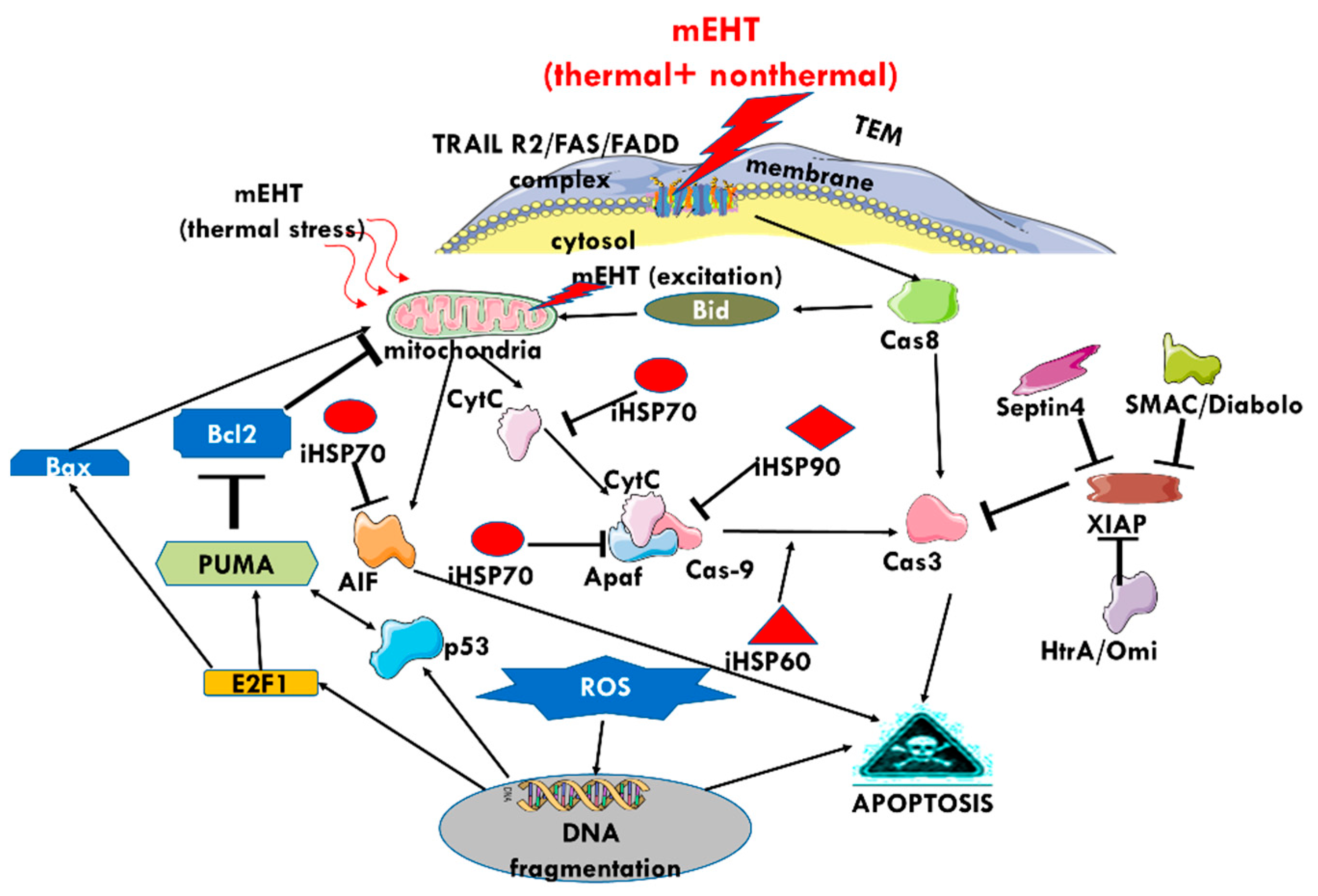
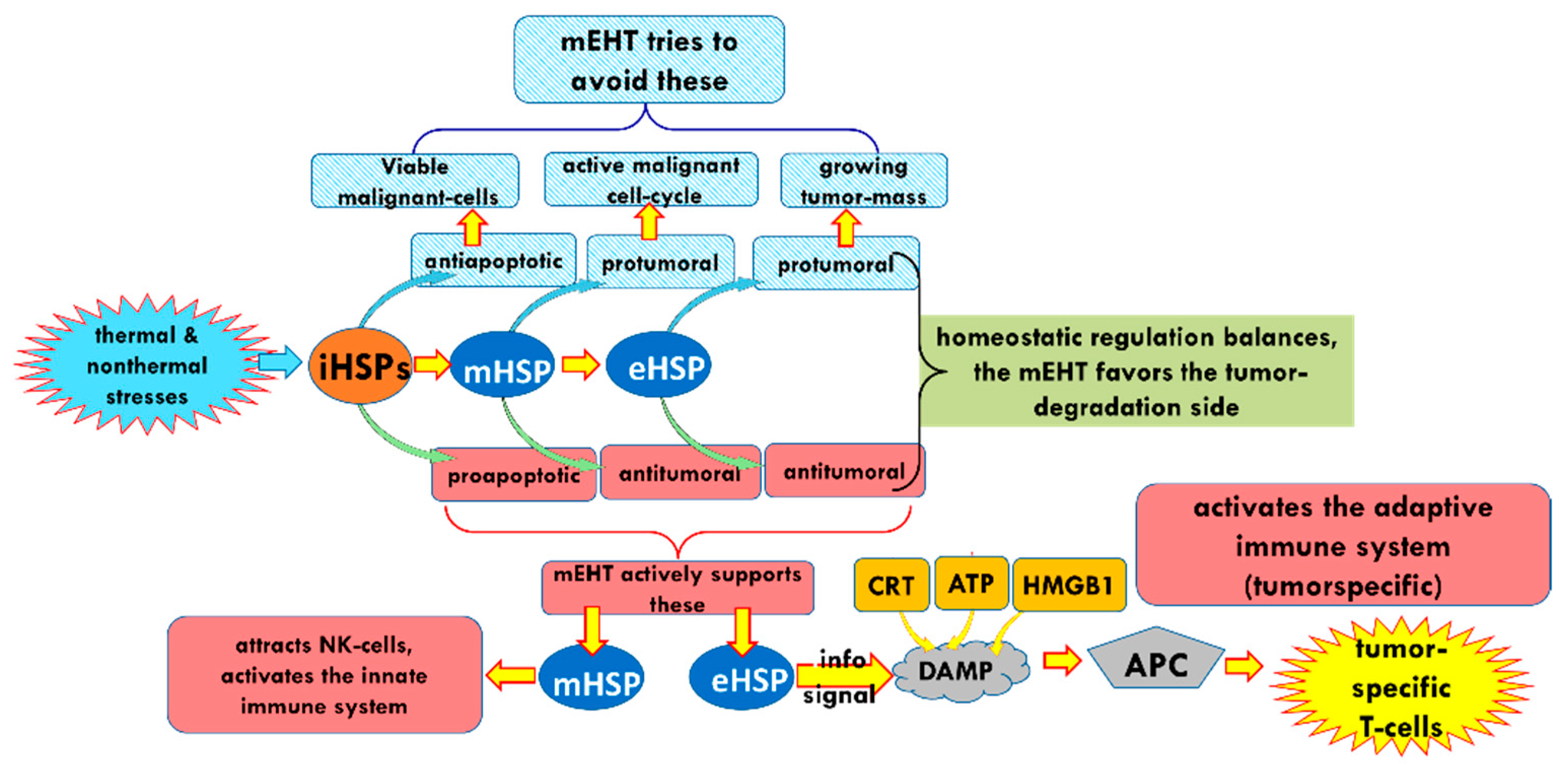
3. The Effect of mEHT on HSPs
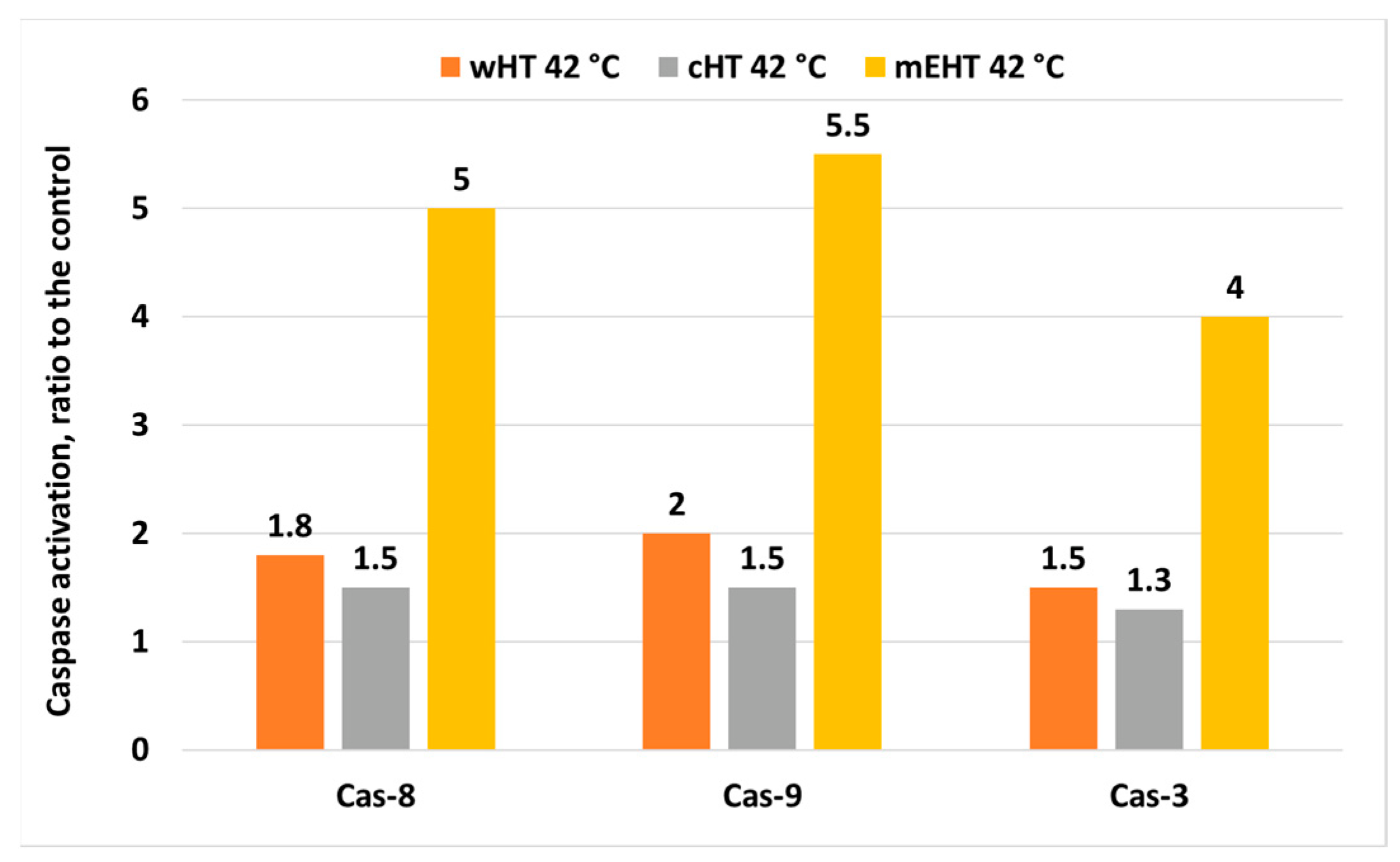
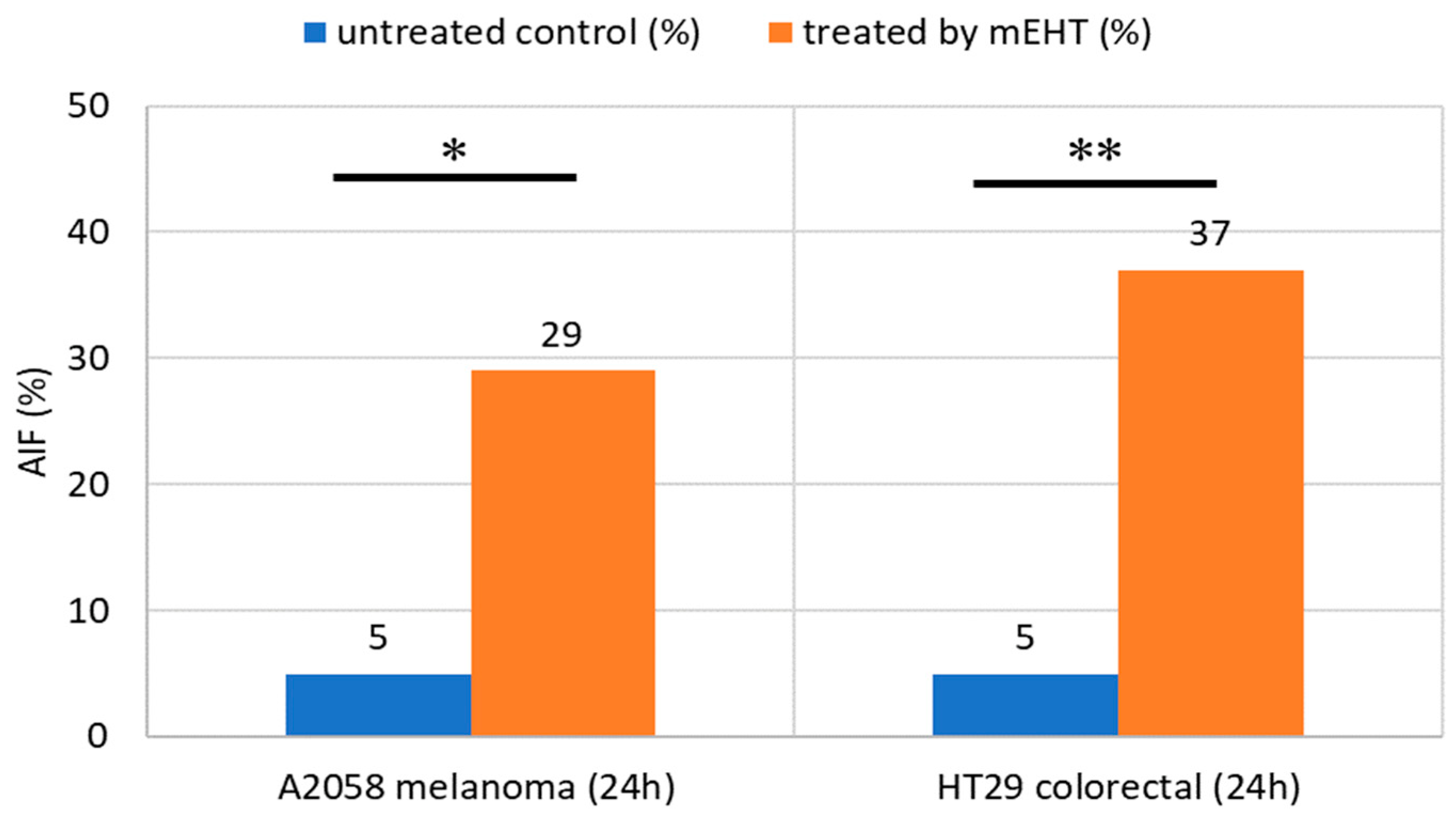
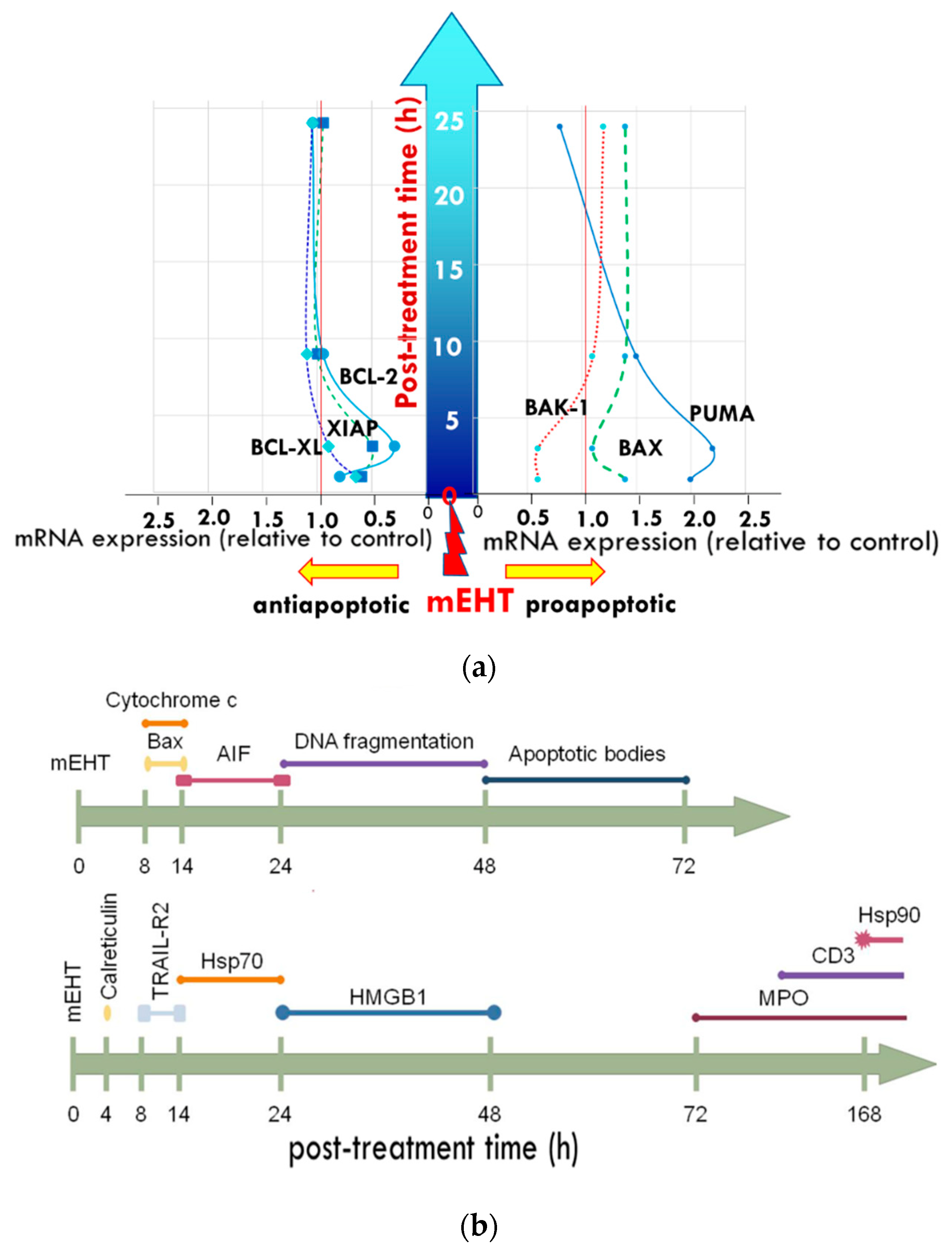
4. The Effect of mEHT on Apoptosis
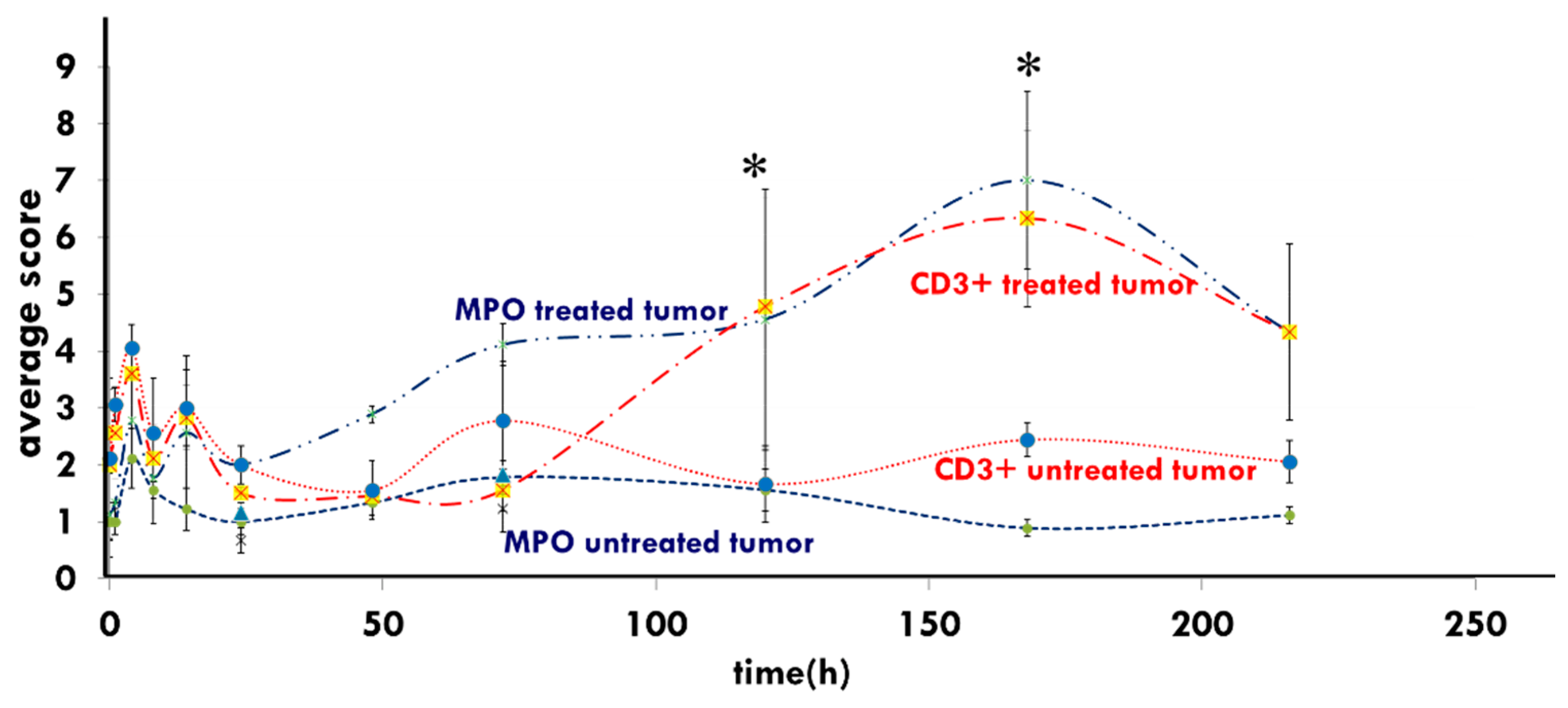
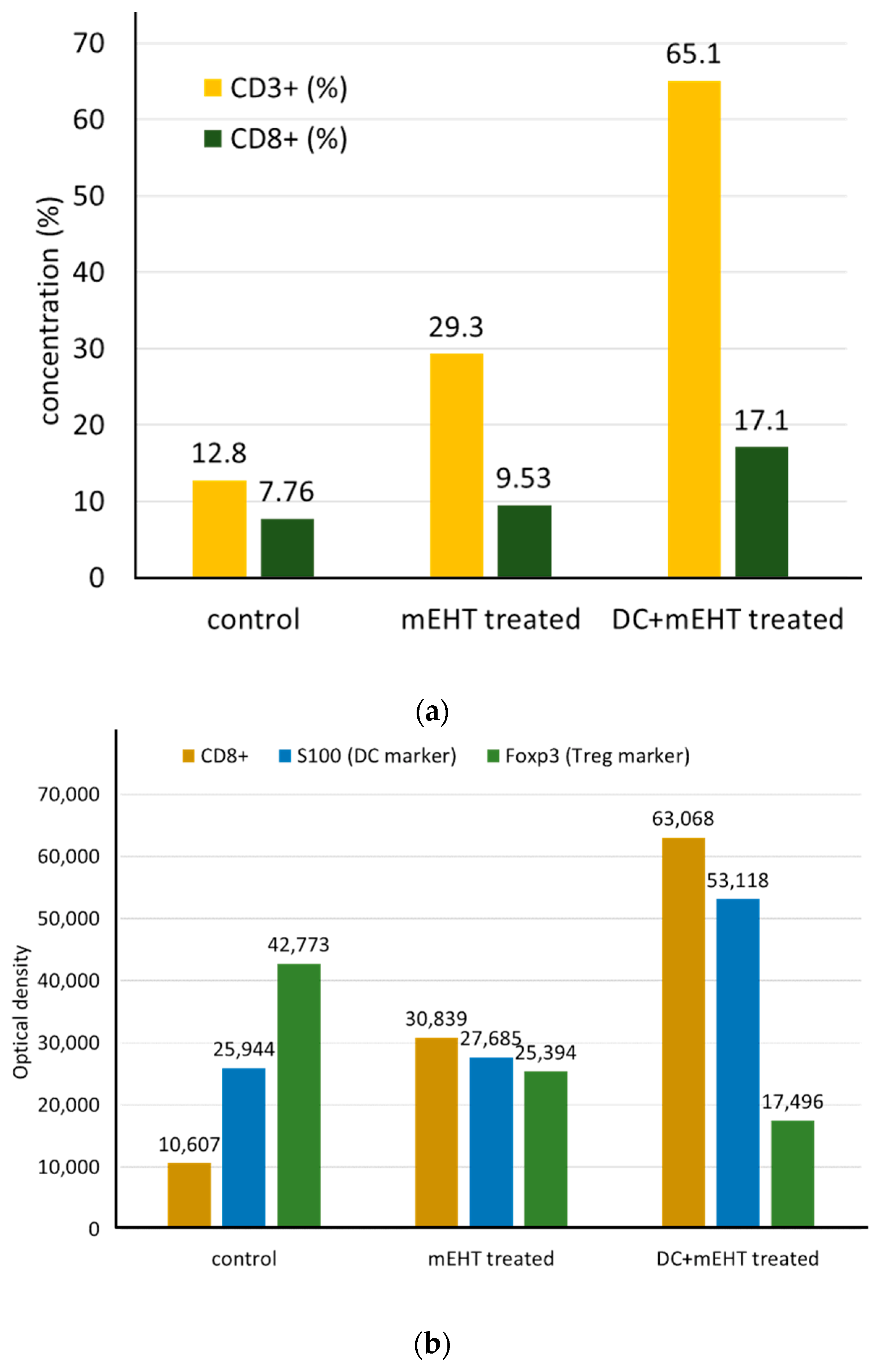
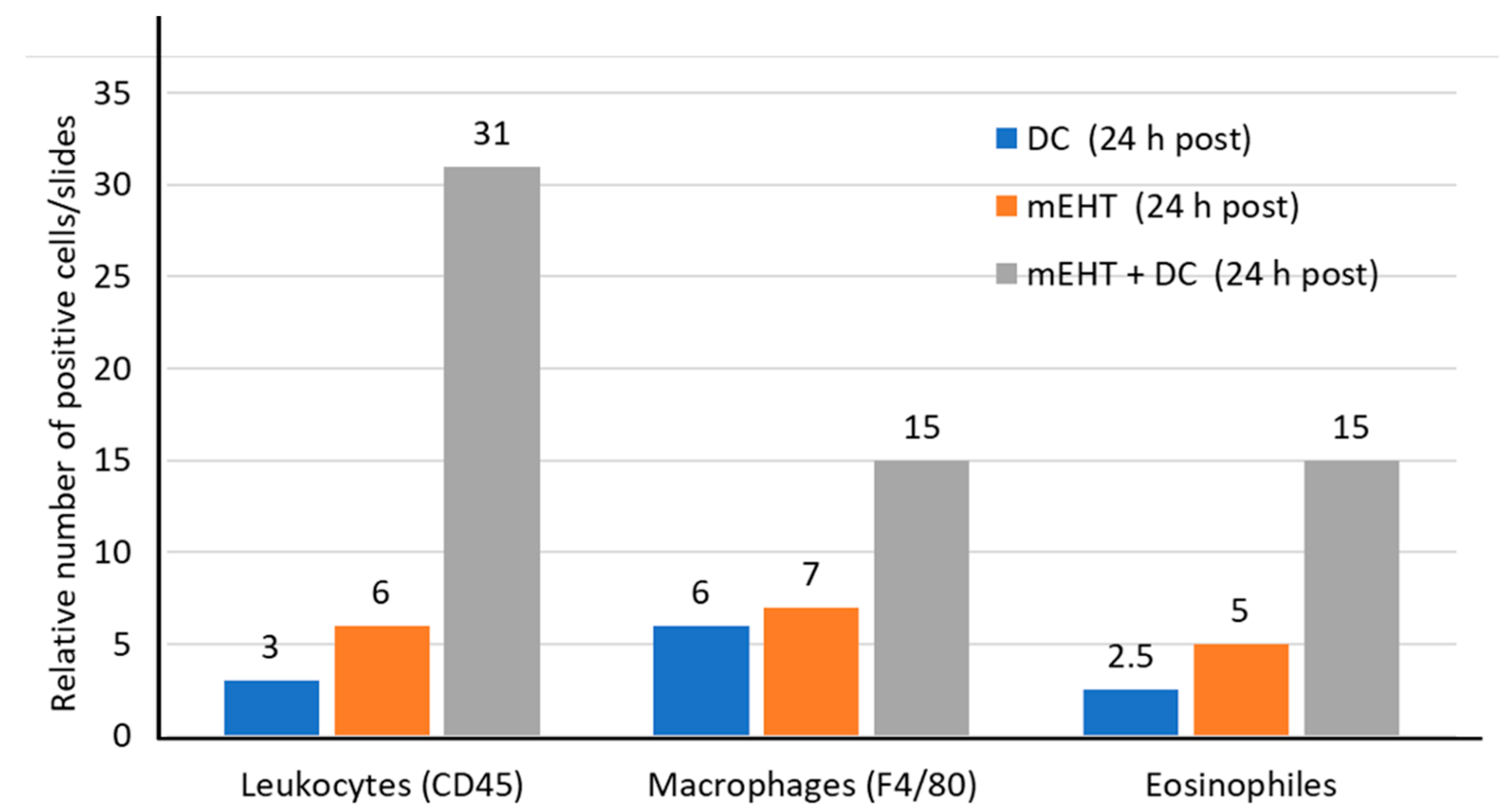
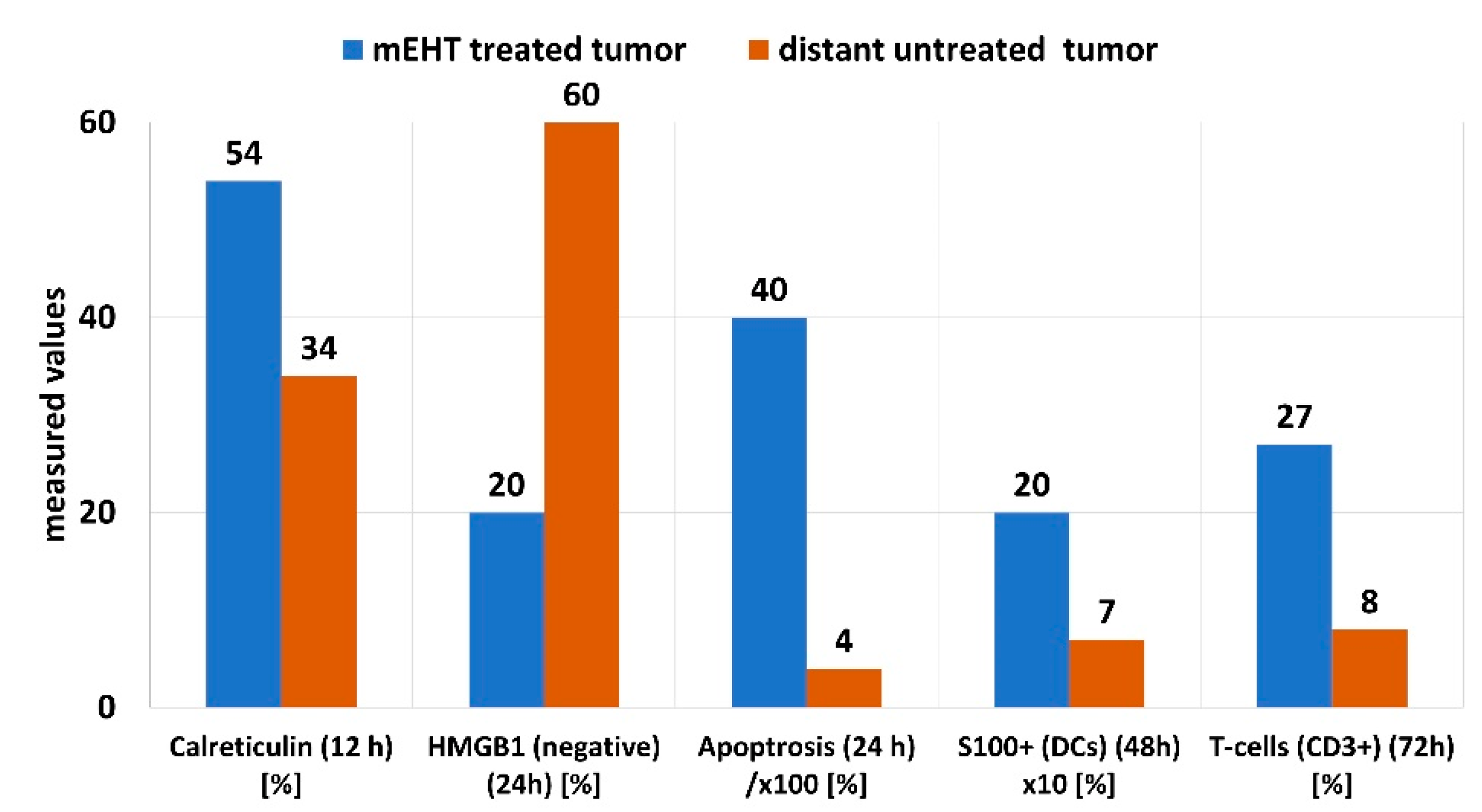
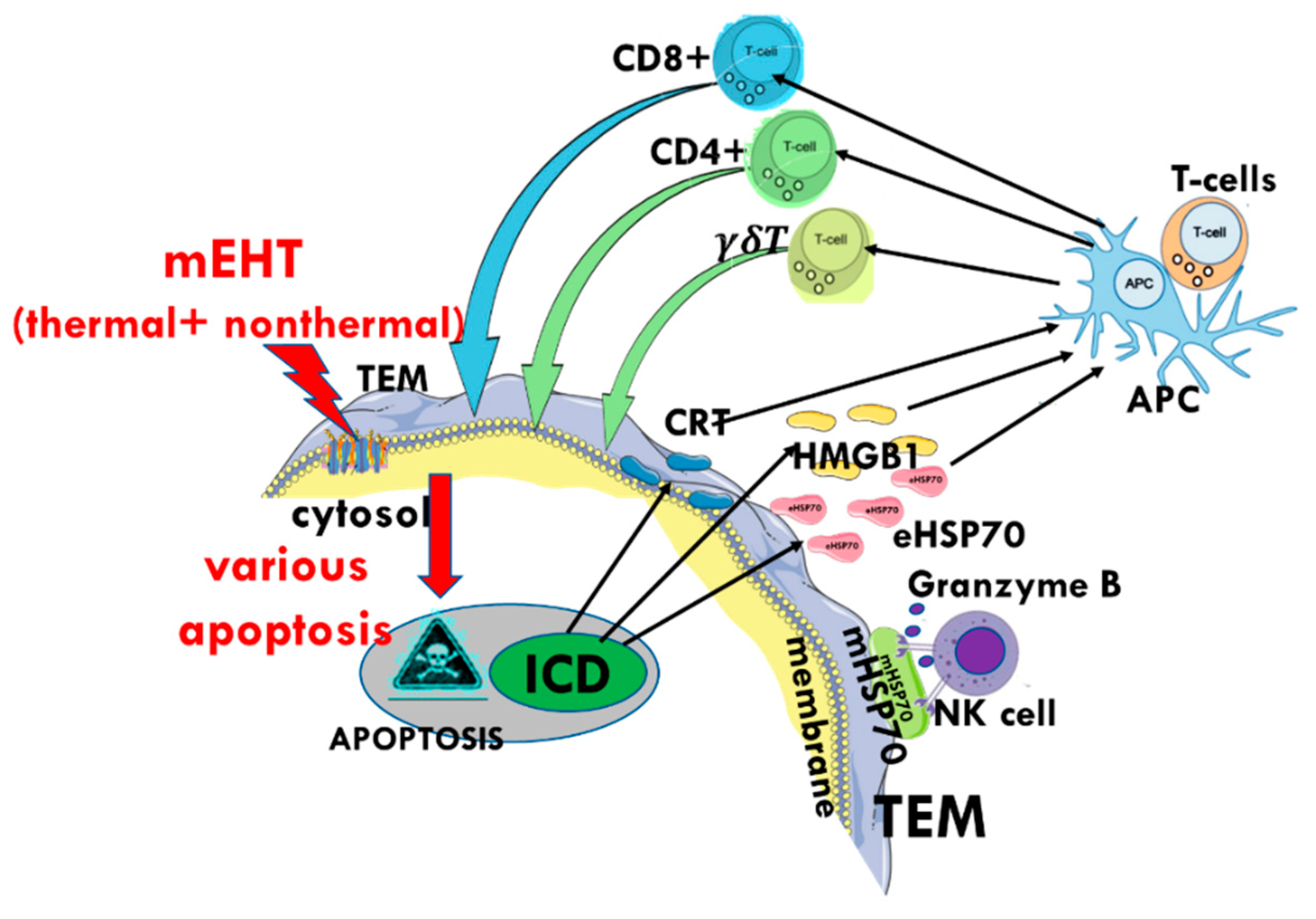
5. Clinical Evidence of a Systemic Immune Effect Induced by mEHT
6. Discussion
7. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Latchman, D.S. Stress Proteins; Springer: Berlin/Heidelberg, Germany, 1999; ISBN 978-3-642-58259-2. [Google Scholar] [CrossRef]
- Zügel, U.; Kaufmann, S.H. Role of heat shock proteins in protection from and pathogenesis of infectious diseases. Clin. Microbiol. Rev. 1999, 12, 19–39. [Google Scholar] [CrossRef] [Green Version]
- Miller, D.J.; Fort, P.E. Heat Shock Proteins Regulatory Role in Neurodevelopment. Front. Neurosci. 2018, 12, 821. [Google Scholar] [CrossRef] [Green Version]
- Horvath, I.; Multhoff, G.; Sonnleitner, A.; Vigh, L. Membrane-associated stress proteins: More than simply chaperones. Biochim. Biophys. Acta 2008, 1778, 1653–1664. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, B.; Feder, M.E.; Kang, L. Evolution of heat-shock protein expression underlying adaptive responses to environmental stress. Mol. Ecol. 2018, 27, 3040–3054. [Google Scholar] [CrossRef] [Green Version]
- Chowdhury, R.; Sahu, G.K.; Das, J. Stress response in pathogenic bacteria. J. Biosci. 1996, 21, 149–160. [Google Scholar] [CrossRef]
- Kaul, G.; Thippeswami, H. Role of heat shock proteins in diseases and their therapeutic potential. Indian J. Microbiol. 2011, 51, 124–131. [Google Scholar] [CrossRef] [Green Version]
- Sriram, K.; Rodriguez-Fernandez, M.; Doyle, F.J. A detailed modular análisis of heat-shock protein dynamics under acute and chronic stress and its implication in anxiety disorders. PLoS ONE 2021, 7, e42958. [Google Scholar] [CrossRef]
- Sorger, P.K. Heat shock factor and the heat shock response. Cell 2012, 65, 363–366. [Google Scholar] [CrossRef]
- Asea, A.A.A.; Calderwood, S.K. Cellular Trafficking of Cell Stress Proteins in Health and Disease; Springer: Berlin/Heidelberg, Germany, 2012; ISBN -13 978-9400747395. [Google Scholar]
- Anckar, J.; Sistonen, L. Regulation of HSF1 function in the heat stress response: Implications in aging and disease. Annu. Rev. Biochem. 2011, 80, 1089–1115. [Google Scholar] [CrossRef]
- Taha, E.A.; Ono, K.; Eguchi, T. Roles of extracellular HSPs as biomarkers in immune surveillance and immune evasion. Int. J. Mol. Sci. 2019, 20, 4588. [Google Scholar] [CrossRef] [Green Version]
- Piacentivi, L.; Fanti, L.; Berloco, M.; Perrii, B.; Piminelli, S. Heterochromatin protein 1 (HP1) is associated with induced gene expressio in Drosophila euchromatin. J. Cell Biol. 2003, 16, 707–714. [Google Scholar] [CrossRef]
- Guan, Y.; Zhu, X.; Liang, J.; Wei, M.; Huang, S.; Pan, X. Upregulation of HSPA1A/HSPA1B/HSPA7 and Downregulation of HSPA9 Were Related to Poor Survival in Colon Cancer. Front. Oncol. 2021, 11, 749673. [Google Scholar] [CrossRef]
- Watanabe, M.; Suzuki, K.; Kodama, S.; Sugahara, T. Normal human cells at confluence get heat resistance by efficient accumulation of HSP72 in nucleus. Carcinogenesis 1995, 16, 2373–2380. [Google Scholar] [CrossRef]
- Macario, A.J.L.; Conway de Macario, E. Heat Shock Response, Overview. In The Encyclopedia of Stress; Academic Press: San Diego, CA, USA, 2000. [Google Scholar]
- Matarredona, L.; Camacho, M.; Zafrilla, B.; Bonete, M.-J.; Esclapez, J. The role of stress proteins in haloarchaea and their adaptive response to environmental shifts. Biomolecules 2020, 10, 1390. [Google Scholar] [CrossRef] [PubMed]
- Calderwood, S.K.; Theriault, J.R.; Gong, J. Message in a bottle: Role of th 70-kDa heat shock protein family I anti-tumor immuity. Eur. J. Immunol. 2005, 35, 2518–2527. [Google Scholar] [CrossRef]
- Albakova, Z.; Mangasarova, Y. The HSP Immune Network in Cancer. Front. Immunol. 2021, 12, 796493. [Google Scholar] [CrossRef]
- Mambula, S.S.; Stevenson, M.A.; Ogawa, K.; Calderwood, S.K. Mechanisms for Hsp70 secretion: Crossing membranes without a leader. Methods 2007, 43, 168–175. [Google Scholar] [CrossRef] [Green Version]
- Kregel, K.C. Heat shock proteins: Modifying factors in physiological stress responses and acquired thermotolerance. J. Appl. Physiol. 2002, 92, 2177–2186. [Google Scholar] [CrossRef] [Green Version]
- Calderwood, S.K.; Mambula, S.S.; Gay, P.J., Jr.; Theriault, J.R. Extracellular heat shock proteins in cell signaling. FEBS Lett. 2007, 581, 3689–3694. [Google Scholar] [CrossRef] [Green Version]
- Shevtsov, M.; Balogi, Z.s.; Khachatryan, W.; Gao, H.; Vigh, L.; Multhoff, G. Membrane-associated heat shock proteins in oncology: From basic research to new theranostic targets. Cells 2020, 9, 1263. [Google Scholar] [CrossRef]
- Tang, D.; Kang, R.; Berghe, T.V.; Vandnabele, P.; Kroemer, G. The molecular machinery of regulated cell death. Cell Res. 2019, 29, 347–364. [Google Scholar] [CrossRef] [Green Version]
- Giri, B.; Sethi, V.; Modi, S.; Garg, B.; Banerjee, S.; Saluja, A.; Dudeja, V. Heat shock protein 70 in pancreatic diseases: Friend or foe. J. Surg. Oncol. 2017, 116, 114–122. [Google Scholar] [CrossRef] [PubMed]
- Pockley, A.G.; Multhoff, G. Cell stress proteins in extracellular fluids: Friend or foe? Novartis Found. Symp. 2008, 291, 86–95. [Google Scholar] [CrossRef]
- Wu, T.; Tanguay, R.M. Antibodies against heat shock proteins in environmental stresses and diseases: Friend or foe? Cell Stress Chaperones 2006, 11, 1. [Google Scholar] [CrossRef] [Green Version]
- Hance, M.W.; Nolan, K.D.; Isaacs, J.S. The double-edged sword: Conserved functions of extracellular Hsp90 in wound healing and cancer. Cancers 2014, 6, 1065–1097. [Google Scholar] [CrossRef] [PubMed]
- Tittelmeier, J.; Nachman, E.; Nussbaum-Krammer, C. Molecular chaperones: A double-sword in neurodegenerative diseases. Front. Aging Neurosci. 2020, 12, 581374. [Google Scholar] [CrossRef]
- Trigos, A.S.; Pearson, R.B.; Papenfuss, A.T.; Goode, D.L. How the evolution of multicellularity set the stage for cancer. Br. J. Cancer 2018, 118, 145–152. [Google Scholar] [CrossRef]
- Popkin, G. Physics sheds light on cancer and bacteria evolution. APC News 2011, 20, 5. [Google Scholar]
- Aktipis, C.A.; Bobby, A.M.; Jansen, G.; Hibner, U.; Hochberg, M.; Maley, C.C.; Wilkinso, G.S. Cancer across the tree of life: Cooperation and cheating in multicellularity. Philos. Trans. R. Soc. B Biol. Sci. 2015, 370, 20140219. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Davidson, C.D.; Wang, W.Y.; Zaimi, I.; Jayco, D.K.P.; Baker, B.M. Cell force-mediated matrix reorganization underlies multicellular network assembly. Sci. Rep. 2019, 9, 12. [Google Scholar] [CrossRef] [Green Version]
- Dvorak, H.F. Tumors: Wounds that do not heal, Similarities between tumor stroma generation and wound healing. N. Engl. J. Med. 1986, 315, 1650–1659. [Google Scholar] [PubMed]
- Punyiczki, M.; Fesus, L. Heat Shock and Apoptosis: The Two Defense Systems of the Organisms May Have Overlapping Molecular Elements. Ann. N. Y. Acad. Sci. 1998, 951, 67–74. [Google Scholar] [CrossRef] [PubMed]
- Young, R.A. Stress Proteins and Immunology. Ann. Rev. Immunol. 1990, 8, 401–420. [Google Scholar] [CrossRef]
- Das, J.K.; Xiong, X.; Ren, X.; Yan, J.M.; Song, J. Heat shock proteins in cancer immunotherapy. J. Oncol. 2019, 2019, 3267207. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moreira-de-Sousa, C.; de Souza, R.B.; Fontanetti, C.S. HSP70 as a biomarker: An excellent tool in environmental contamination analysis—A review. Water Air Soil Pollut. 2018, 229, 264. [Google Scholar] [CrossRef] [Green Version]
- Siebert, C.; Ciato, D.; Murakami, M.; Frei-Stuber, L.; Perez-Rivas, L.G.; Moteseri-Garcia, J.L.; Nölting, S.; Maurer, J.; Perez-Rivas, L.G.; Walch, A.K.; et al. Heat shock protein 90 as a prognostic marker and therapeutic target for adrenocortical carcinoma. Front. Endocrinol. 2019, 10, 487. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Carpenter, R.L.; Gokmen-Polar, Y. HSF1 as a cancer biomarker and therapeutic target. Curr. Cancer Drug Targets 2019, 19, 515–524. [Google Scholar] [CrossRef]
- Soti, C.; Csermely, P. Molecular Chaperones in the Etiology and Therapy of Cancer. Pathol. Oncol. Res. 1998, 4, 316–321. [Google Scholar] [CrossRef] [PubMed]
- Soti, C.; Nagy, E.; Giricz, Z.; Vigh, L.; Csermely, P.; Ferdinady, P. Heat shock proteins as emerging therapeutic targets. Br. J. Pharmacol. 2005, 146, 769–780. [Google Scholar] [CrossRef] [Green Version]
- Xu, M.; Wright, W.D.; Higashikubo, R.; Roti, J.L.R. Chronic Thermotolerance with Continued Cell Proliferation. Int. J. Hyperth. 1996, 12, 645–660. [Google Scholar] [CrossRef]
- Pirity, M.; Hever-Szabo, A.; Venetianer, A. Overexpression of P-glycoprotein in Heta and/or Drug Resistant Hepatoma Variants. Cytotechnology 1996, 19, 207–214. [Google Scholar] [CrossRef] [PubMed]
- Santin, A.D.; Hermonat, P.L.; Ravaggi, A.; Chiriva-Iternati, M.; Hiserodt, J.C.; Batchu, R.B.; Pecorelli, S.; Parham, G.P. The Effects of Irradiation on the Expression of a Tumor Rejection Antigen (Heat Shock Protein GP96) in Human Cervical Cancer. Int. Radiat. Biol. 1998, 76, 699–704. [Google Scholar]
- Morgan, J.; Whitaker, J.E.; Oseroff, A.R. GRP78 Induction by Calcium Ionophore Potentiates Photodynamic Therapy Using the Mitochondrial Targeting Dye Victoria Blue BO. Photocem. Photobiol. 1998, 67, 155–164. [Google Scholar] [CrossRef]
- Calderwood, S.K.; Murshid, A. Molecular chaperone accumulation in cancer and decrease in Alzheimer’s disease: The potential roles of HSF1. Front. Neurosci. 2017, 11, 192. [Google Scholar] [CrossRef] [Green Version]
- Ciocca, D.R.; Calderwood, S.K. Heat shock proteins in cancer: Diagnostic, prognostic, predictive, and treatment implications. Cell Stress Chaperones 2005, 10, 86–103. [Google Scholar] [CrossRef]
- Rothammer, A.; Sage, E.K.; Werner, C.; Combs, S.E.; Multhoff, G. Increased heat shock protein 70 (Hsp70) serum levels and low NK cell counts after radiotherapy-potential markers for predicting breast cancer recurrence? Radiat. Oncol. 2019, 14, 78. [Google Scholar] [CrossRef] [PubMed]
- Mosser, D.D.; Caron, A.W.; Bourget, L.; Denis-Larose, C.; Massie, B. Role of the human heat shock protein hsp70 in protection against stress-induced apoptosis. Mol. Cell. Biol. 1997, 17, 5317–5327. [Google Scholar] [CrossRef] [Green Version]
- Vega, V.L.; Rodriguez-Silva, M.; Frey, T.; Gehrmann, M.; Diaz, J.C.; Stinem, C.; Multhoff, G.; Arispe, N.; De Maio, A. Hsp70 translocates into the plasma membrane after stress and is released into the extracellular environment in a membrane-associated form that activates macrophages. Immunology 2008, 180, 4299–4307. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gehrmann, M.; Marinhage, J.; Eichholtz-Wirtz, H.; Fritz, E.; Ellwart, J.; Jaattla, M.; Zilch, T.; Multhoff, D. Dual function of membrane-bound heat shock protein (Hsp70), Bag-4, and Hsp40, protection against radiation-induced effects and target structure for natural killer cells. Cell Death Differ. 2005, 12, 38–51. [Google Scholar] [CrossRef] [Green Version]
- Snigireva, A.V.; Vrublevskaya, V.V.; Skarga, Y.Y.; Morenkov, O.S. The role of membrane-bound heat shock Hsp90 proteins in the migration of tumor cells in vitro and the involvement of cell surface heparan sulfate proteoglycans in protein binding to the plasma membrane. Biophysics 2016, 61, 277–283. [Google Scholar] [CrossRef]
- Radons, J.; Multhoff, G. Immunostimulatory functions of membrane-bound and exported heat shock protein 70. Exerc. Immunol. Rev. 2005, 11, 17–33. [Google Scholar]
- Multhoff, G.; Botzler, C.; Jennen, L.; Schmidt, L.; Ellwart, J.; Issels, R. Heat shock protein 72 on tumor cells: A recognition structure for natural killer cells. J. Immunol. 1997, 158, 4341–4350. [Google Scholar] [PubMed]
- Ablakova, Z.; Siam, M.K.S.; Sacitharan, P.K.; Sacitharan, P.K.; Ziganshin, R.H.; Ryazantsev, D.Y.; Sapozhnikov, A.M. Extracellular heat shock proteins and cancer-New perspectives. Transl. Oncol. 2021, 14, 100995. [Google Scholar] [CrossRef]
- Gas, P. Essential facts on the history of hyperthermia and their connections with electromedicine. arXiv 2017, arXiv:1710.00652. [Google Scholar]
- Paulides, M.M.; Trefna, H.D.; Curto, S.; Rodrigues, D.B. Recent technological advancements in radiofrequency- and microwave-mediated hyperthermia for enhancing drug delivery. Adv. Drug Deliv. Rev. 2020, 163–164, 3–18. [Google Scholar] [CrossRef]
- Calderwood, S.K.; Gong, J. Heat shock proteins promote cancer: It’s a protection racket. Trends Biochem. Sci. 2016, 41, 311–323. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schilling, D.; Kuhnel, A.; Konrad, S.; Tetzlaff, F.; Bayer, C.; Yaglom, J.; Multhoff, G. Sensitizing tumor cells to radiation by targeting the heat shock response. Cancer Lett. 2015, 360, 294–301. [Google Scholar] [CrossRef] [Green Version]
- Desai, S.; Liu, Z.; Yao, J.; Pat, N.; Chen, J.; Wu, Y.; Ah, Y.; Fodstad, O.; Tan, M. Heat Shock Factor 1 (HSF1) Controls Chemoresistance and Autophagy through Transcriptioal Regulation of Autophagy-relatd Protein 7 (ATG7). J. Biol. Chem. 2013, 288, 9165–9176. [Google Scholar] [CrossRef] [Green Version]
- Chatterjee, S.; Burns, T.F. Targeting Heat Shock Proteins in Cancer-A Promising Therapeutic Approach. Int. J. Mol. Sci. 2017, 18, 1978. [Google Scholar] [CrossRef] [Green Version]
- Yoneda, A.; Miomi, K.; Tamura, Y. Heat shock protein 47 confers chemoresistance on pancreatic cancer cells by interacting with calreticulin and IRE1α. Cancer Sci. 2021, 112, 2803–2820. [Google Scholar] [CrossRef]
- Schwab, M.; Thunborg, K.; Azmzadeh, O.; Von Torne, C.; Werner, C.; Shevtsov, M.; De Genio, T.; Zdralevic, M.; Pouyssegur, J.; Renner, K.; et al. Targeting Cancer Metabolism Breaks Radioresistance by Impairing the Stress Response. Cancers 2021, 13, 3762. [Google Scholar] [CrossRef]
- Cheng, Y.; Weng, S.; Yu, L.; Zhu, N.; Yang, M.; Yuan, Y. The Role of Hyperthermia in the Multidisciplinary Treatment of Malignant Tumors. Integr. Cancer Ther. 2019, 18, 1534735419876345. [Google Scholar] [CrossRef]
- Datta, N.R.; Jain, B.M.; Mathi, Z.; Datta, S.; Johari, S.; Singh, A.R.; Kalbande, P.; Kale, P.; Shivkumar, V.; Bodis, S. Hyperthermia: A potential game-changer in the management of cancers in low-middl-income group countries. Cancers 2022, 14, 315. [Google Scholar] [CrossRef] [PubMed]
- Fiorentini, G.; Sarti, D.; Gadaleta, C.D.; Ballerini, M.; Fiorentini, C.; Garfango, T.; Ranieri, G.; Guadagni, S. A Narrative Review of Regional Hyperthermia-Updates from 2010 to 2019. Integr. Cancer Ther. 2020, 19, 1534735420932648. [Google Scholar] [CrossRef] [PubMed]
- Datta, N.R.; Kok, H.P.; Crezee, H.; Gaipi, U.S.; Bodis, S. Integrating loco-regional hyperthermia into the current oncology practice: SWOT and TOWS analyses. Front. Oncol. 2020, 10, 819. [Google Scholar] [CrossRef]
- Hurwitz, M.D. Hyperthermia and immunotherapy: Clinical opportunities. Int. J. Hyperth. 2019, 36, 4–9. [Google Scholar] [CrossRef]
- Roussakow, S. The History Of Hyperthermia Rise And Decline. Conf. Pap. Med. 2012, 2013, 428027. [Google Scholar] [CrossRef]
- Lee, S.Y.; Szigeti, G.P.; Szasz, A.M. Oncological hyperthermia: The correct dosing in clinical applications. Int. J. Oncol. 2019, 54, 627–643. [Google Scholar] [CrossRef]
- Cherukuri, P.; Glazer, E.S.; Curley, S.A. Targeted hyperthermia using metal nanoparticles. Adv. Drug Deliv. Rev. 2010, 62, 339–345. [Google Scholar] [CrossRef] [Green Version]
- Sohail, A.; Ahmed, Z.; Beg, O.A.; Arshad, S.; Sherin, L. A Review on hypertermia via nanoparticle-mediated therapy. Bull. Cancer 2017, 104, 452–461. [Google Scholar] [CrossRef]
- Nicolau, D.V.; Burrage, K.; Parton, R.G.; Hancock, J.F. Identifying Optimal Lipid Raft Characteristics Required to Promote Nanoscale Protein-Protein Interactions on the Plasma Membrane. J. Mol. Cell Biol. 2006, 26, 313–323. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, K.L.; Huang, C.C.; Chi, M.S.; Chiang, H.C.; Wang, Y.S.; Andocs, G.; Wang, H.-E.; Chi, K.-H. In vitro comparison of conventional hyperthermia and modulated electro-hyperthermia. Oncotarget 2016, 7, 84082–84092. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wust, P.; Kortum, B.; Strauss, U.; Nadobny, J.; Zschaeck, S.; Beck, M.; Stein, U.; Ghadjar, P. Non-thermal effects of radiofrequency electromagnetic fields. Sci. Rep. 2020, 10, 13488. [Google Scholar] [CrossRef] [PubMed]
- Wust, P.; Stein, U.; Ghadjar, P. Non-thermal membrane effects of electromagnetic fields and therapeutic applications in oncology. Int. J. Hyperth. 2021, 38, 715–731. [Google Scholar] [CrossRef] [PubMed]
- Frolich, H. What are non-thermal electric biological effects? Bioelectromagnetics 1982, 3, 45–46. [Google Scholar] [CrossRef]
- Vincze, G.; Szigeti, G.; Andocs, G.; Szasz, A. Nanoheating without Artificial Nanoparticles. Biol. Med. 2015, 7, 249. [Google Scholar]
- Sezgin, E.; Levental, I.; Mayor, S.; Eggelig, C. The mystery of membrane organization: Composition, regulation and roles of lipid rafts. Nat. Rev. Mol. Cell Biol. 2017, 18, 361–374. [Google Scholar] [CrossRef] [Green Version]
- Nicolson, G.L. The Fluid—Mosaic Model of Membrane Structure: Still relevant to understanding the structure, function and dynamics of biological membranes after more than 40 years. Biochim. Biophys. Acta 2014, 1838, 1451–1466. [Google Scholar] [CrossRef] [Green Version]
- Staunton, J.R.; Agus, D.B.; Alexander, J.F.; Arap, W.; Ashili, S.; Aslan, J.E.; Austin, R.H.; Backman, V.; Bethel, K.J.; Bonneau, R.; et al. The Physical Sciences-Oncology Centers Network; A physical sciences network characterization of non-tumorigenic and metastatic cells. Sci. Rep. 2008, 3, 1449. [Google Scholar]
- Cha, J.; Jeon, T.W.; Lee, C.G.; Oh, S.T.; Yang, H.-B.; Choi, K.-J.; Seo, D.; Yun, I.; Bain, I.H.; Park, K.R.; et al. Electro-hyperthermia inhibits glioma tumorigenicity through the induction of E2F1-mediated apoptosis. Int. J. Hyperth. 2015, 31, 784–792. [Google Scholar] [CrossRef]
- Blank, M. Evidence for Stress Response (Stress Proteins), Health Risk of Electromagnetic Fields: Research on the Stress Response. 2007. Available online: https://citeseerx.ist.psu.edu/viewdoc/download?doi=10.1.1.534.8023&rep=rep1&type=pdf (accessed on 21 October 2021).
- Blank, M.; Goodman, R. Electromagnetic fields stress living cells. Pathophysiology 2009, 16T, 71–78. [Google Scholar] [CrossRef] [Green Version]
- Schwan, H.P. Nonthermal Cellular Effects of Electromagnetic Fields: AC-Field Induced Ponderomotoric Forces. Br. J. Cancer 1982, 45, 220–224. [Google Scholar]
- Szasz, A. Therapeutic basis of electromagnetic resonances and signal-modulation. Open J. Biophys. 2021, 11, 314–350. [Google Scholar] [CrossRef]
- Szasz, A.; Iluri, N.; Szasz, O. Local Hyperthermia in Oncology–To Choose or Not to Choose? In Hyperthermia; Huilgol, N., Ed.; InTech: London, UK, 2013; Chapter 1; pp. 1–82. ISBN 980-953-307-019-8. [Google Scholar]
- Blank, M. Coupling of AC Electric Fields to Cellular Processes. In Proceedings of the First International Symposium on Nonthermal Medical/Biological Treatments Using Electromagnetic Fields and Ionized Gases, ElectroMed’99, Symposium Record Abstracts, Norfolk, VA, USA, 12–14 April 1999; p. 23. [Google Scholar]
- Zeni, O.; Simko, M.; Scarfi, M.R.; Mattsson, M.O. Cellular response to ELF-MF and heat: Evidence for a common involvement of heat shock proteins? Front. Public Health 2017, 5, 280. [Google Scholar] [CrossRef] [Green Version]
- Minnaar, C. Challenges Associated with Hyperthermia. In Challenges and Solutions of Oncological Hyperthermia; Szasz, A., Ed.; Cambridge Scholars: Newcastle upon Thyne, UK, 2020; Chapter 1; pp. 1–31. [Google Scholar]
- Szasz, O.; Szasz, A. Approaching complexity: Hyperthermia dose and its possible measurement in oncology. Open J. Biophys. 2021, 11, 68–132. [Google Scholar] [CrossRef]
- Lee, S.Y.; Lee, N.R.; Cho, D.H.; Kim, J.S. Treatment outcome analysis of chemotherapy combined with modulated electro-hyperthermia compared with chemotherapy alone for recurrent cervical cancer, following irradiation. Oncol. Lett. 2017, 14, 73–78. [Google Scholar] [CrossRef] [Green Version]
- Minnaar, C.; Kotzen, J.A.; Baeyens, A. Modulated Electro-Hyperthermia Improves Three Year Survival in Cervical Cancer Patients, Presentation number: PH-0551. In Proceedings of the ESTRO Conference, Madrid, Spain, 27–31 October 2021. [Google Scholar]
- Minnaar, C.A.; Kotzen, J.A.; Ayeni, O.A.; Naidoo, T.; Tunmer, M.; Sharma, V.; Vangu, M.-D.-V.; Bayes, A. The effect of modulated electro-hyperthermia on local disease control in HIV-positive and -negative cervical cancer women in South Africa: Early results from a phase III randomized controlled trial. PLoS ONE 2019, 14, e0217894. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, S.-Y.; Kim, J.-H.; Han, Y.-H.; Cho, D.-H. The effect of modulated electro-hyperthermia on temperature and blood flow in human cervical carcinoma. Int. J. Hyperth. 2018, 34, 953–960. [Google Scholar] [CrossRef]
- Minnaar, C.A.; Kotzen, J.A.; Naidoo, T.; Naidoo, T.; Tunmer, M.; Sharma, V.; Vangu, M.-D.-V.; Bayes, A. Analysis of the effects of mEHT on the treatment- related toxicity and quality of life of HIV-positive cervical cancer patients. Int. J. Hyperth. 2020, 37, 263–272. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Papp, E.; Vancsik, T.; Kiss, E.; Szasz, O. Energy absorption by the membrane rafts in the modulated electro-hyperthermia (mEHT). Open J. Biophys. 2017, 7, 216–229. [Google Scholar] [CrossRef] [Green Version]
- Szasz, A. The capacitive coupling modalities for oncological hyperthermia. Open J. Biophys. 2021, 11, 252–313. [Google Scholar] [CrossRef]
- Szasz, O.; Szasz, A.; Iluri, N. RF Hyperthermia Device for Personalized Treatment and Diagnosis. U.S. Patent 9,937,357 B2, 10 April 2018. [Google Scholar]
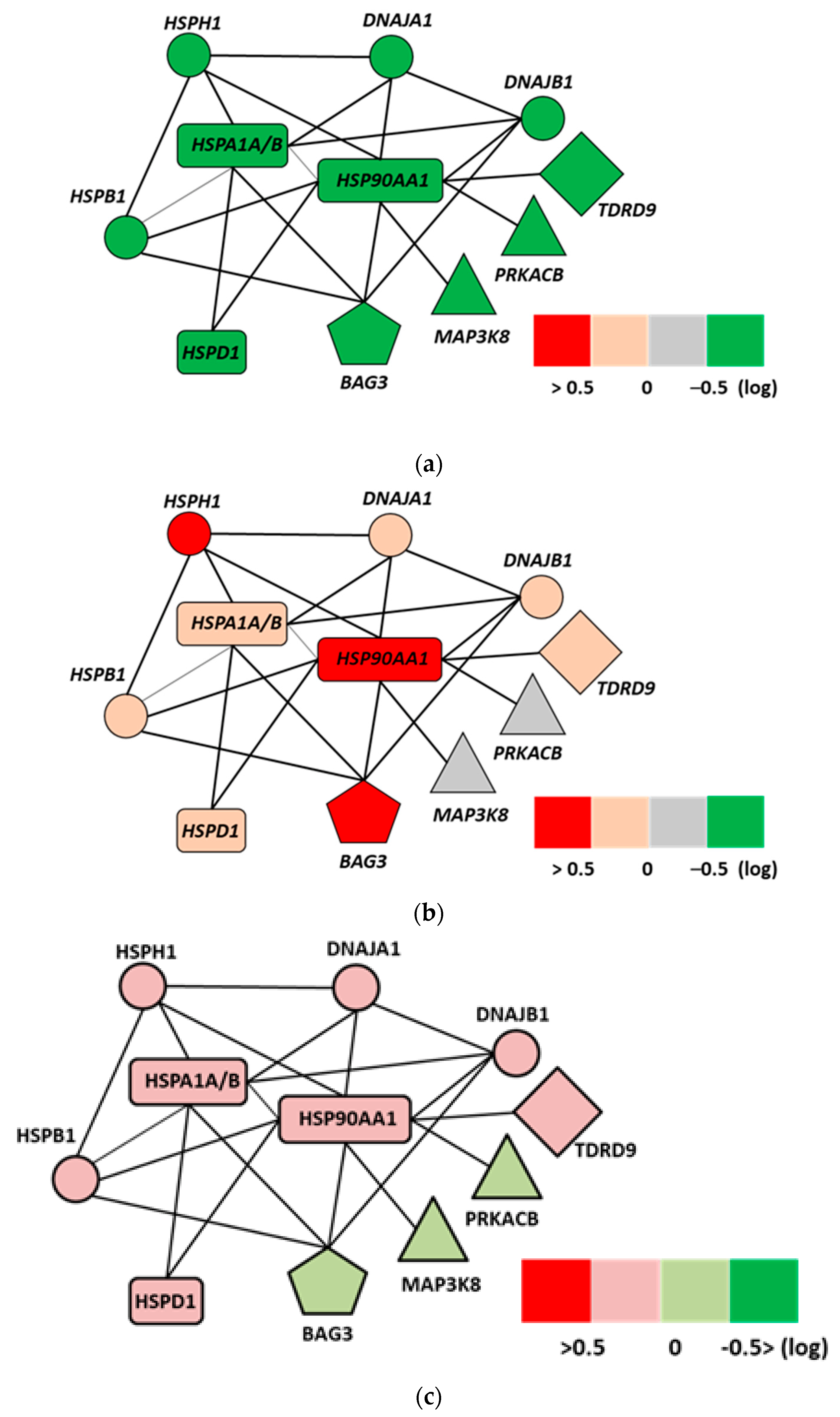
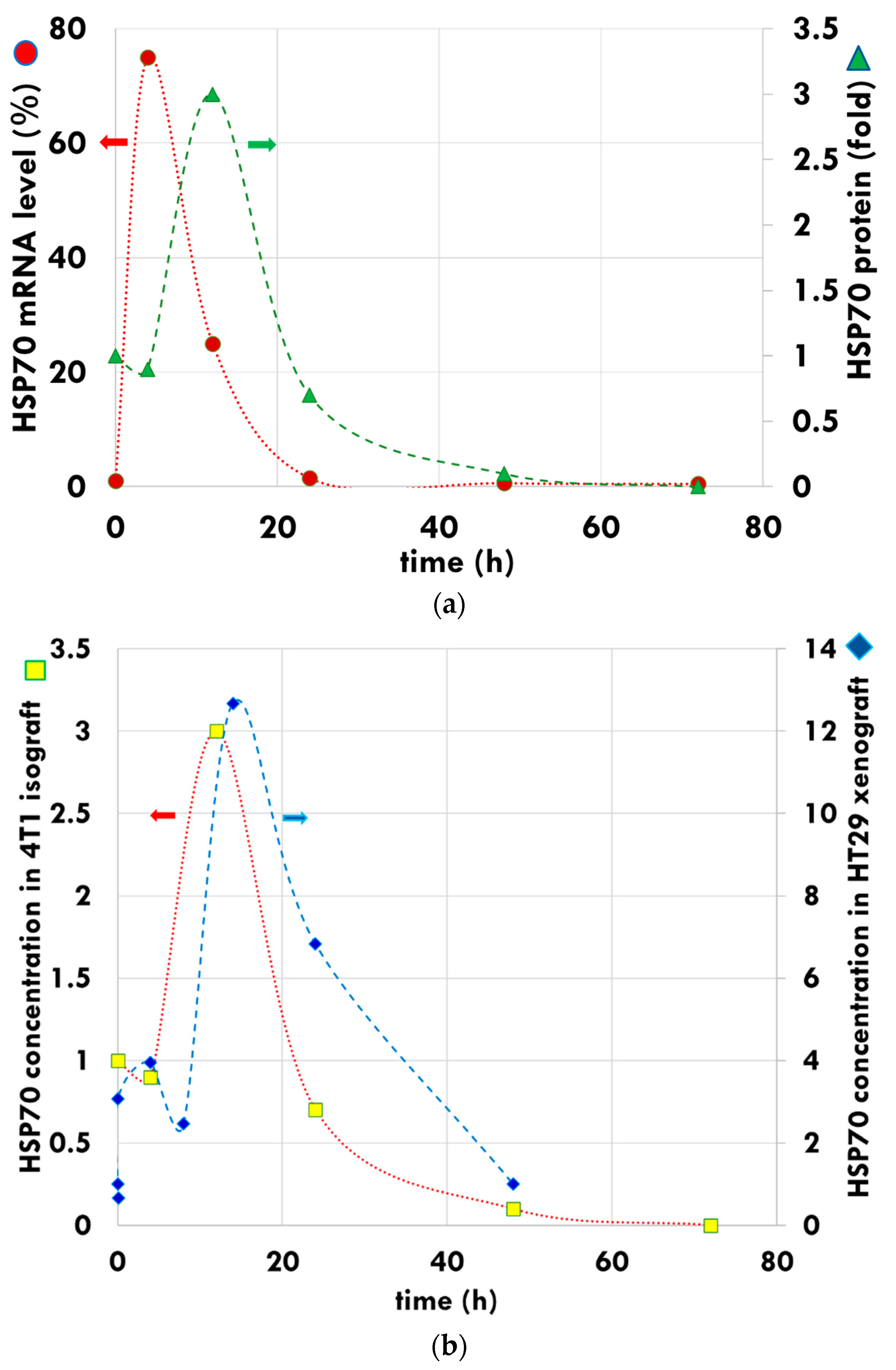
- Andocs, G.; Meggyeshazi, N.; Balogh, L.; Spisak, S.; Maros, M.E.; Balla, P.; Kiszner, G.; Teleki, I.; Kovago, C.; Krenacs, T. Upregulation of heat shock proteins and the promotion of damage-associated molecular pattern signals in a colorectal cancer model by modulated electrohyperthermia. Cell Stress Chaperones 2014, 20, 37–46. [Google Scholar] [CrossRef] [Green Version]
- Meggyeshazi, N.; Andocs, G.; Spisak, S.; Krenacs, T. Early Changes in mRNA and Protein Expression Related to Cancer Treatment by Modulated Electro-Hyperthermia. In Proceedings of the International Clinical Hyperthermia Society, Budapest, Hungary, 12–14 October 2012; Hindawi Publishing Corporation Conference Papers in Medicine: London, UK, 2013. [Google Scholar]
- Meggyeshazi, N. Studies on Modulated Electrohyperthermia Induced Tumor Cell Death in a Colorectal Carcinoma Model. Ph.D. Thesis, Semmelweis University, Budapest, Hungary, 2015. [Google Scholar]
- Schvarcz, C.A.; Danics, L.; Krenacs, T.; Viana, P.; Beres, R.; Vancsik, T.; Nagy, A.; Gyenesei, A.; Kun, J.; Fonovic, M.; et al. Modulated electro-hyperthermia induces a prominent local stress response and growth inhibition in mouse breast cancer isografts. Cancers 2021, 13, 1744. [Google Scholar] [CrossRef]
- Andocs, G.; Rehman, M.U.; Zhao, Q.L.; Tabuchi, Y.; Kanamori, M.; Kondo, T. Comparison of biological effects of modulated electro-hyperthermia and conventional heat treatment in human lymphoma U937 cell. Cell Death Discov. 2016, 2, 16039. [Google Scholar] [CrossRef] [Green Version]
- Danics, L.; Schvarcz, C.A.; Viana, P.; Vancsik, T.; Krenacs, T.; Benyo, Z.; Kaucsar, T.; Hamar, P. Exhaustion of protective heat shock response induces significant tumor damage by apoptosis after modulated electro-hyperthermia treatment of triple negative breast cancer isografts in mice. Cancers 2020, 12, 2581. [Google Scholar] [CrossRef]
- Besztercei, B.; Vancsik, T.; Benedek, A.; Marjor, E.; Thomas, M.J.; Schvarcz, C.A.; Krenacs, T.; Benyo, Z.; Balogh, A. Stress-induced, p53-mediated tumor growth inhibition of melanoma by modulated electrohyperthermia in mouse models without major immunogenic effects. Int. J. Mol. Sci. 2019, 20, 4019. [Google Scholar] [CrossRef] [Green Version]
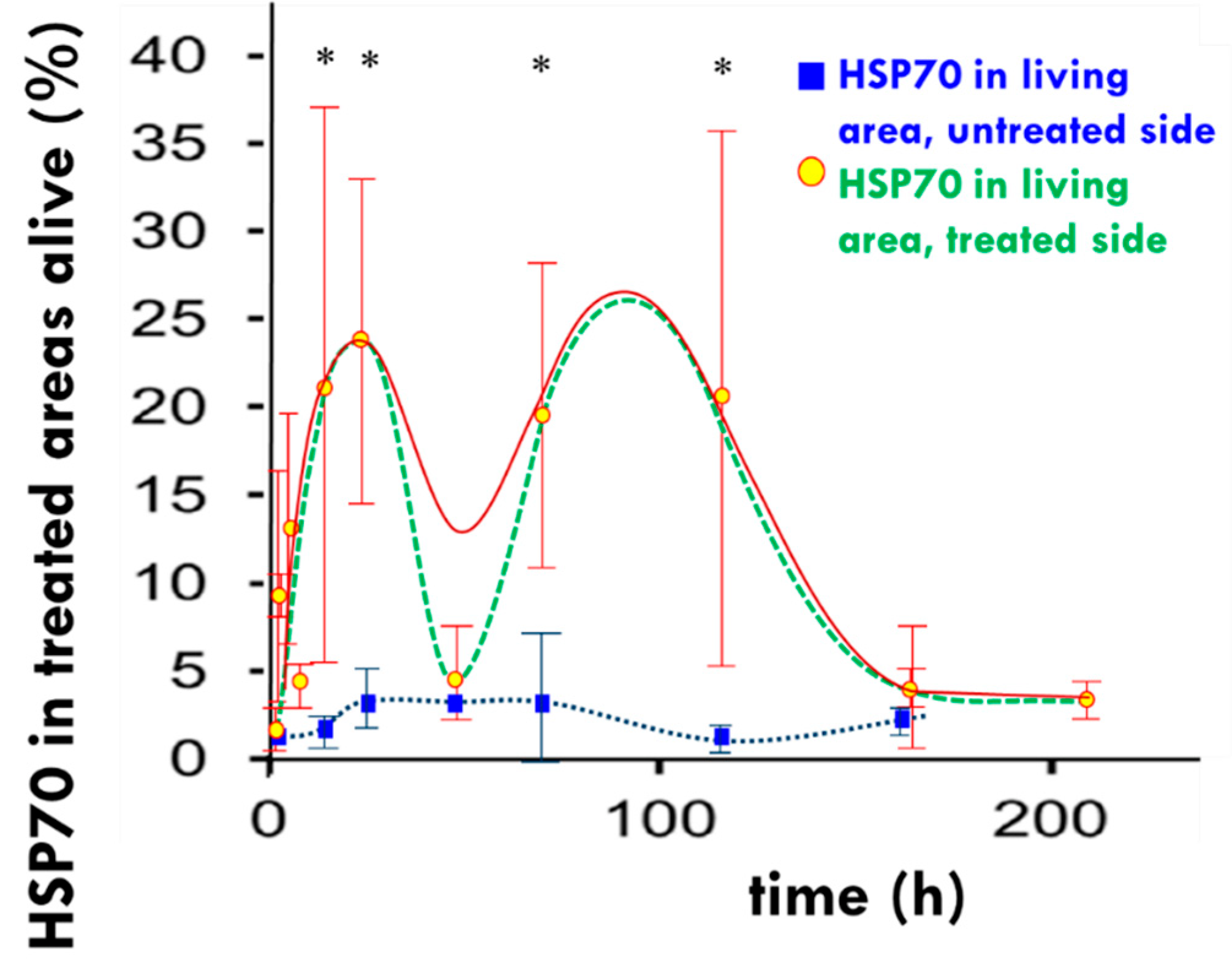
- Vancsik, T.; Kovago, C.; Kiss, E.; Papp, E.; Forika, G.; Benyo, Z.; Meggyeshazi, N.; Krenacs, T. Modulated electro-hyperthermia induced loco-regional and systemic tumor destruction in colorectal cancer allografts. J. Cancer 2018, 9, 41–53. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Qin, W.; Akutsu, Y.; Andocs, G.; Suganami, A.; Hu, X.; Yusup, G.; Komatsu-Akimoto, A.; Hoshino, I.; Hanari, N.; Mori, M.; et al. Modulated electro-hyperthermia enhances dendritic cell therapy through an abscopal effect in mice. Oncol. Rep. 2014, 32, 2373–2379. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Meggyeshazi, N.; Andocs, G.; Krenacs, T. Programmed Cell Death Induced by Modulated Electro-Hyperthermia. In Proceedings of the International Clinical Hyperthermia Society 2012, Budapest, Hungary, 12–14 October 2012; Hindawi Publishing Corporation Conference Papers in Medicine: London, UK, 2013; Volume 2013, p. 187835. [Google Scholar]
- Meggyesházi, N.; Andocs, G.; Balogh, L.; Balla, P.; Kiszner, G.; Teleki, I.; Jeney, A.; Krenács, T. DNA fragmentation and caspase-independent programmed cell death by modulated electrohyperthermia. Strahlenther. Onkol. 2014, 190, 815–822. [Google Scholar] [CrossRef] [PubMed]
- Sevrioukova, I.F. Apoptosis-inducing factor: Structure, function, and redox regulation. Antioxid. Redox Signal. 2011, 14, 2545–2579. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tsang, Y.-W.; Huang, C.-C.; Yang, K.-L.; Chi, M.-S.; Chiang, H.-C.; Wang, Y.-S.; Andocs, G.; Szasz, A.; Li, W.-T.; Chi, K.-H. Improving immunological tumor microenvironment using electro-hyperthermia followed by dendritic cell immunotherapy. BMC Cancer 2015, 15, 708. [Google Scholar] [CrossRef] [Green Version]
- Wust, P.; Ghadjar, P.; Nadobny, J. Physical analysis of temperature-dependent effects of amplitude-modulated electromagnetic hyperthermia. Int. J. Hyperth. 2019, 36, 1246–1254. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Warfel, N.A.; El-Deiry, W.S. p21WAF1 and tumourigenesis. Curr. Opin. Oncol. 2013, 25, 52–58. [Google Scholar] [CrossRef] [PubMed]
- Vancsik, T.; Máthé, D.; Horváth, I.; Várallyaly, A.A.; Benedek, A.; Bergmann, R.; Krenács, T.; Benyó, Z.; Balogh, A. Modulated electro-hyperthermia facilitates NK-cell infiltration and growth arrest of human A2058 melanoma in a xenograft model. Front. Oncol. 2021, 11, 164. [Google Scholar] [CrossRef]
- Multhoff, G. Activation of natural killer cells by heat shock protein 70. Int. J. Hyperth. 2002, 18, 576–585. [Google Scholar] [CrossRef]
- Yilmaz, M.T.; Elmali, A.; Yazici, G. Abscopal Effect, From Myth to Reality: From Radiation Oncologists’ Perspective. Cureus 2019, 11, e3860. [Google Scholar] [CrossRef] [Green Version]
- Wahl, R.L.; Jacene, H.; Kasamon, Y.; Lodge, M.A. From RECIST to PERCIST: Evolving Considerations for PET Response Criteria in Solid Tumors. J. Nucl. Med. 2009, 50, 122–151. [Google Scholar] [CrossRef] [Green Version]
- Brix, N.; Tiefenthaller, A.; Anders, H.; Belka, C.; Lauber, K. Abscopal, immunological effects of radiotherapy: Narrowing the gap between clinical and preclinical experiences. Immunol. Rev. 2017, 280, 249–279. [Google Scholar] [CrossRef] [PubMed]
- Tubin, S.; Casamassima, F.; Menichelli, C. A Case Report on Metastatic Thyroid Carcinoma: Radiation-induced Bystander or Abscopal Effect ? J. Cancer Sci. Ther. 2012, 4, 408–411. [Google Scholar] [CrossRef] [Green Version]
- Hlavata, Z.; Solinas, C.; De Silva, P.; Porcu, M.; Saba, L.; Willard-Gallo, K.; Scartozzi, M. The Abscopal Effect in the Era of Cancer Immunotherapy: A Spontaneous Synergism Boosting Anti-tumor Immunity? Target. Oncol. 2018, 13, 113–123. [Google Scholar] [CrossRef] [PubMed]
- Reynders, K.; Illidge, T.; Siva, S.; Chang, J.Y.; De Ruysscher, D. The abscopal effect of local radiotherapy: Using immunotherapy to make a rare event clinically relevant. Cancer Treat. Rev. 2015, 41, 503–510. [Google Scholar] [CrossRef] [Green Version]
- Kao, P.H.-J.; Chen, C.-H.; Tsang, Y.-W.; Lin, C.-S.; Chiang, H.-C.; Huang, C.-C.; Chi, M.-S.; Yang, K.-L.; Li, W.-T.; Kao, S.-J.; et al. Relationship between energy dosage and apoptotic cell death by modulated electro-hyperthermia. Sci. Rep. 2020, 10, 8936. [Google Scholar] [CrossRef]
- Krenacs, T.; Benyo, Z. Tumor specific stress and immune response induced by modulated electrohyperthermia in relation to tumor metabolic profiles. Oncothermia J. 2017, 20, 264–272. [Google Scholar]
- Yoon, S.-M.; Jung, S.L. Case of Abscopal effect with Metastatic Non-Small-Cell Lung Cancer. Oncothermia J. 2012, 5, 53–57. [Google Scholar]
- Fiorentini, G.; Yoon, S.M.; Yan, O.; Andocs, G.; Baronzio, G.F.; Laurent, S.; Balogh, L.; Szasz, A. Abscopal effect: New perspectives in Oncothermia. Oncothermia J. 2013, 7, 279–281. [Google Scholar]
- Chi, M.-S.; Wu, J.-H.; Shaw, S.; Wu, C.-J.; Chen, L.-K.; Hsu, H.-C.; Chi, K.-H. Marked local and distant response of heavily treated breast cancer with cardiac metastases treated by combined low dose radiotherapy, low dose immunotherapy and hyperthermia: A case report. Ther. Radiol. Oncol. 2021, 5, 17. [Google Scholar] [CrossRef]
- Schirrmacher, V.; Stücker, W.; Lulei, M.; Bihari, A.-S.; Sprenger, T. Long-term survival of a breast cancer patient with extensive liver metastases upon immune and virotherapy: A case report. Immunotherapy 2015, 7, 855–860. [Google Scholar] [CrossRef] [Green Version]
- Chi, M.-S.; Mehta, M.P.; Yang, K.-L.; Lai, H.-C.; Lin, Y.-C.; Ko, H.-L.; Wang, Y.-S.; Liao, K.-W.; Chi, K.-H. Putative abscopal effect in three patients treated by combined radiotherapy and modulated electrohyperthermia. Front. Oncol. 2020, 10, 254. [Google Scholar] [CrossRef]
- Chi, K.-H. Tumour-Directed Immunotherapy: Clinical Results of Radiotherapy with Modulated Electro-Hyperthermia. In Challenges and Solutions of Oncological Hyperthermia; Szasz, A., Ed.; Cambridge Scholars: London, UK, 2020; Chapter 12; pp. 206–226. [Google Scholar]
- Chi, K.H. Tumor-directed immunotherapy: Combined radiotherapy and oncothermia. Oncothermia J. 2018, 24, 196–235. [Google Scholar]
- Pang, C.L.K. The Immune Regulating Effect of Hyperthermia in Combination with TCM on Cancer Patients. Oncothermia J. 2016, 18, 170–179. [Google Scholar]
- Van Gool, S.W.; Makalowski, J.; Bonner, E.R.; Feyen, O.; Domogalla, M.P.; Prix, L.; Schirrmacher, V.; Nazarian, J.; Stuecker, W. Addition of multimodal immunotherapy to combination treatment strategies for children with DIPG: A single institution experience. Medicines 2020, 7, 29. [Google Scholar] [CrossRef] [PubMed]
- Van Gool, S.W.; Makalowski, J.; Fiore, S.; Sprenger, T.; Prix, L.; Schirrmacher, V.; Stuecker, W. Randomized controlled immunotherapy clinical trials for GBM challenged. Cancers 2021, 13, 32. [Google Scholar] [CrossRef]
- Van Gool, S.W.; Makalowski, J.; Feyen, O.; Prix, L.; Schirrmacher, V.; Stuecker, W. The induction of immunogenic cell death (ICD) during maintenance chemotherapy and subsequent multimodal immunotherapy for glioblastoma (GBM). Austin Oncol. Case Rep. 2018, 3, 1–8. [Google Scholar]
- Van Gool, S.W.; Makalowski, J.; Stuecker, W. Modulated electrohyperthermia (mEHT) as part of multimodal immunotherapy for brain tumors. Oncothermia J. 2018, 248. [Google Scholar]
- Van Gool, S.; Makalowski, J.; Marko, M. Hyperthermia as part of multimodal immunotherapy for patients with GBM. Oncothermia J. 2019, 27, 122–137. [Google Scholar]
- Van Gool, S.; Makalowski, J.; Marko, M. Multimodal immunotherapy for patients with ovarian cancer. Oncothermia J. 2019, 27, 138–152. [Google Scholar]
- Minnaar, C.A.; Kotzen, J.A.; Ayeni, O.A.; Vangu, M.-D.-T.; Baeyens, A. Potentiation of the Abscopal Effect by Modulated Electro-Hyperthermia in Locally Advanced Cervical Cancer Patients. Front. Oncol. 2020, 10, 376. [Google Scholar] [CrossRef] [Green Version]
- Blad, B.; Wendel, P.; Jonsson, M.; Lindstrom, K. An electrical impedance index to distinguish between normal and cancerous tissues. J. Med. Eng. Technol. 1999, 23, 57–62. [Google Scholar] [CrossRef] [PubMed]
- Zhou, J.; Wang, G.; Chen, Y.; Wang, H.; Hua, Y.; Cai, Z. Immunogenic cell death in cancer therapy: Present and emerging inducers. J. Cell. Mol. Med. 2019, 23, 4854–4865. [Google Scholar] [CrossRef]
- Szasz, A. Electromagnetic Effects in Nanoscale Range. In Cellular Response to Physical Stress and Therapeutic Applications; Tadamichi, S., Takashi, K., Eds.; Nova Science Publishers, Inc.: Hauppauge, NY, USA, 2013; Chapter 4. [Google Scholar]
- Andocs, G.; Rehman, M.U.; Zhao, Q.L.; Papp, E.; Kondo, T.; Szasz, A. Nanoheating without Artificial Nanoparticles Part II. Experimental support of the nanoheating concept of the modulated electro-hyperthermia method, using U937 cell suspension model. Biol. Med. 2015, 7, 4. [Google Scholar] [CrossRef] [Green Version]
- Szasz, A. Thermal and nonthermal effects of radiofrequency on living state and applications as an adjuvant with radiation therapy. J. Radiat. Cancer Res. 2019, 10, 1–17. [Google Scholar] [CrossRef]
- Szasz, O.; Szasz AMMinnaar, C.; Szasz, A. Heating preciosity-trends in modern oncological hyperthermia. Open J. Biophys. 2017, 7, 116–144. [Google Scholar] [CrossRef] [Green Version]
- Ronchi, R.; Marano, L.; Braidotti, P.; Bianciardi, P.; Calamia, M.; Fiorentini, C.; Samaja, M. Effects of broad band magnetic fields on HSP70 expression and ischemia-reperfusion in rat hearts. Life Sci. 2004, 75, 1925–1936. [Google Scholar] [CrossRef]
- Goodman, R.; Blank, M. Insights into electromagnetic interaction mechanisms. J. Cell. Physiol. 2002, 192, 16–22. [Google Scholar] [CrossRef]
- Sapozhnikov, A.M.; Ponomarev, E.D.; Tarasenko, T.N.; Telford, W.G. Spontaneous apoptosis and expression of cell-surface heat-shock proteins in cultured EL-4 lymphoma cells. Cell Prolif. 1999, 32, 363–378. [Google Scholar] [CrossRef]
- Horvath, I.; Vigh, L. Cell biology: Stability in times of stress. Nature 2010, 463, 436–438. [Google Scholar] [CrossRef]
- Hildebrandt, B.; Wust, P.; Ahlers, O.; Dieing, A.; Sreenivasa, G.; Kerner, T.; Felix, R.; Riess, H. The cellular and molecular basis of hyperthermia. Crit. Rev. Oncol. Hematol. 2002, 43, 33–56. [Google Scholar] [CrossRef]
- Nishida, T.; Akagi, K.; Tanaka, Y. Correlation between cell killing effect and cell membrane potential after heat treatment: Analysis using fluorescent dye and flow cytometry. Int. J. Hyperth. 1997, 13, 227–234. [Google Scholar] [CrossRef]
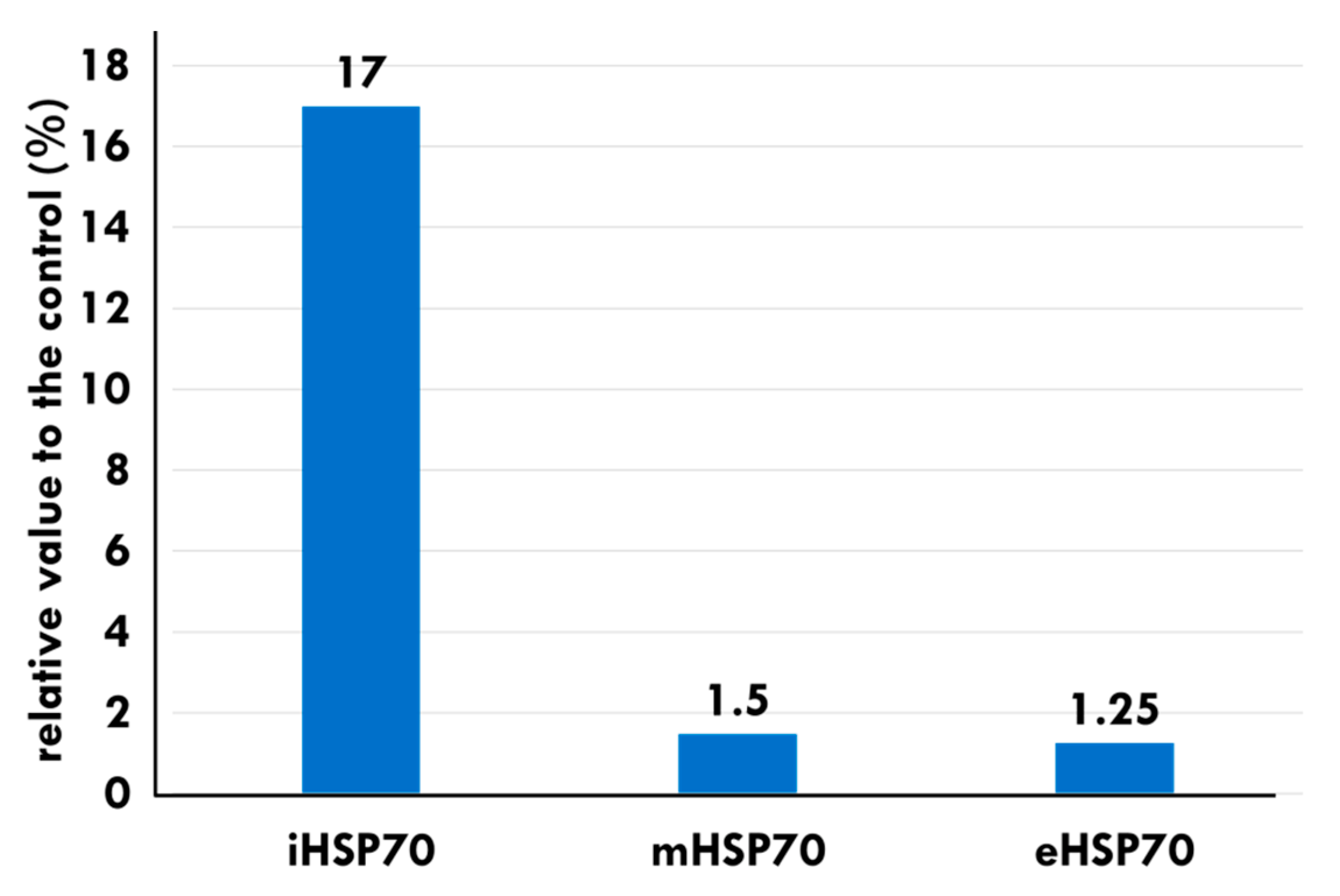
- Gehrmann, M.; Radons, J.; Molls, M.; Multhoff, G. The therapeutic implications of clinically applied modifiers of heat shock protein 70 (Hsp70) expression by tumor cells. Cell Stress Chaperones 2008, 13, 1–10. [Google Scholar] [CrossRef]
- Pfister, K.; Radons, J.; Busch, R.; Tidball, J.G.; Pfeifer, M.; Freitag, L.; Feldmann, H.J.; Milani, V.; Issels, R.; Multhoff, G. Patient survival by Hsp70 membrane phenotype: Association with different routes of metastasis. Cancer 2007, 110, 926–935. [Google Scholar] [CrossRef]
- Multhoff, G. Heat shock protein 70 (Hsp70): Membrane location, export and immunological relevance. Methods 2007, 43, 229–237. [Google Scholar] [CrossRef] [PubMed]
- Andreev, V.P. Cytoplasmic electric fields and electroosmosis: Possible solution for the paradoxes of the intracellular transport of biomolecules. PLoS ONE 2013, 8, e61884. [Google Scholar] [CrossRef] [Green Version]
- Multhoff, G.; Hightower, L.E. Distinguishing integral and receptor-bound heat shock protein 70 (Hsp70) on the cell surface by Hsp70-specific antibodies. Cell Stress Chaperones 2011, 16, 251–255. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Calderwood, S.K. Molecular co-chaperones: Tumor growth and cancer treatment. Scientifica 2013, 2013, 217513. [Google Scholar] [CrossRef] [Green Version]
- Spisek, R.; Charalambous, A.; Mazumder, A.; Vesole, D.H.; Jagannath, S.; Dhodapkar, M.V. Bortezomib enhances dendritic cell (DC)-mediated induction of immunity to human myeloma via exposure of cell surface heat shock protein 90 on dying tumor cells: Therapeutic implications. Blood 2007, 109, 4839–4845. [Google Scholar] [CrossRef]
- Multhoff, G.; Botzler, C.; Wiesnet, M.; Müller, E.; Meier, T.; Wilmanns, W.; Issels, R.D. A stress-inducible 72-kDa heat-shock protein (HSP72) is expressed on the surface of human tumor cells, but not on normal cells. Int. J. Cancer 1995, 61, 272–279. [Google Scholar] [CrossRef]
- Gehrmann, M.; Liebisch, G.; Schmitz, G.; Anderson, R.; Stinem, C.; De Maio, A.; Pockley, G.; Multhoff, G. Tumor-Specific Hsp70 Plasma Membrane Localization Is Enabled by the Glycosphingolipid Gb3. PLoS ONE 2008, 2, e1925–e1933. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xanthoudakis, S.; Roy, S.; Rasper, D.; Heessey, T.; Aubin, Y.; Cassady, R.; Tawa, P.; Ruel, R.; Rosen, A.; Nicholson, D.W. Hsp60 accelerates the maturation of pro-caspase-3 by upstream activator proteases during apoptosis. EMBO J. 1999, 18, 2049–2056. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Szasz, A.; Vincze, G.; Szasz, O.; Szasz, N. An energy analysis of extracellular hyperthermia. Electromagn. Biol. Med. 2003, 22, 103–115. [Google Scholar] [CrossRef]
- Jeon, T.W.; Yang, H.; Lee, C.G.; Oh, S.T.; Seo, D.; Baik, I.H.; Lee, E.H.; Yun, I.; Park, K.R.; Lee, Y.H. Electro-hyperthermia up-regulates tumor suppressor Septin 4 to induce apoptotic cell death in hepatocellular carcinoma. Int. J. Hyperth. 2016, 32, 648–656. [Google Scholar] [CrossRef] [Green Version]
- Hou, W.; Zhang, Q.; Yan, Z.; Chen, R.; Zeh, H.J., III; Kang, R.; Lotze, M.T.; Tang, D. Strange attractors: DAMPs and autophagy link tumor cell death and immunity. Cell Death Dis. 2013, 4, e966. [Google Scholar] [CrossRef]
- Murao, A.; Aziz, M.; Wang, H.; Brenner, M.; Wang, P. Release mechanisms of major DAMPs. Apoptosis 2021, 26, 152–162. [Google Scholar] [CrossRef] [PubMed]
- Garg, A.D.; Nowis, D.; Golab, J.; Vandenabeele, P.; Krysko, D.V.; Agostinis, P. Immunogenic cell death, DAMPs and anticancer therapeutics: An emerging amalgamation. Biochim. Biophys. Acta 2010, 1805, 53–71. [Google Scholar] [CrossRef] [PubMed]
- Lin, F.C.; Hsu, C.H. Nano-therapeutic cancer immunotherapy using hyperthermia-induced heat shock proteins: Insights from mathematical modeling. Int. J. Nanomed. 2018, 13, 3529–3539. [Google Scholar] [CrossRef] [Green Version]
- Ito, A.; Honda, H.; Kobayashi, T. Cancer immunotherapy based on intracellular hyperthermia using magnetite nanoparticles: A novel concept of “heat-controlled necrosis” with heat shock protein expression. Cancer Immunol. Immunother. 2005, 55, 320–328. [Google Scholar] [CrossRef]
- Ablakova, Z.; Armeev, G.A.; Kanevskiy, L.M.; Kovalenko, E.I.; Sapozhikov, A.M. HSP70 multi-funtionality in cancer. Cells 2020, 9, 587. [Google Scholar] [CrossRef] [Green Version]
- Stocki, P.; Dickinson, A.M. The immunosuppressive activity of heat shock protein 70. Autoimmune Dis. 2012, 2012, 617213. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Beachy, S.H.; Repasky, E.A. Toward establishment of temperature thresholds for immunological impact of heat exposure in humans. Int. J. Hyperth. 2011, 27, 344–352. [Google Scholar] [CrossRef]
- Vakkila, J.; Lotze, M.T. Inflammation and necrosis promote tumour growth. Nat. Rev. Immunol. 2004, 4, 641–648. [Google Scholar] [CrossRef]
- Feyerabend, T.; Wiedemann, G.J.; Jager, B.; Vesely, H.; Mahlmann, B.; Richter, E. Local hyperthermia, radiation, and chemotherapy in recurrent breast cancer is feasible and effective except for inflammatory disease. Int. J. Radiat. Oncol. Biol. Phys. 2001, 49, 1317–1325. [Google Scholar] [CrossRef]
- Kumar, S.; Deepak, P.; Kumar, S.; Kishore, D.; Acharya, A. Autologous Hsp70 induces antigen specific Th1 immune responses in a murine T-cell lymphoma. Immunol. Investig. 2009, 38, 449–465. [Google Scholar] [CrossRef] [PubMed]
- Jolesch, A.; Elmer, K.; Bendz, H.; Issels, R.D.; Nossner, E. Hsp70, a messenger from hyperthermia for the immune system. Eur. J. Cell Biol. 2011, 91, 48–52. [Google Scholar] [CrossRef] [Green Version]
- Hicman-Miller, H.D.; Hildbrand, H. The immune response under stress: The role of HSP-derived peptides. Trends Immunol. 2004, 25, 427–433. [Google Scholar] [CrossRef] [PubMed]
- Binder, R.J. Functions of heat shock proteins in pathways of the innate and adaptive immune system. J. Immunol. 2014, 193, 5765–5771. [Google Scholar] [CrossRef] [Green Version]
- Keep, O.; Galluzzi, L.; Senovilla, L.; Panarkakis, T.; Tesiere, A.; Schlemmer, F.; Madeo, F.; Zitvogel, L.; Kroemer, G. Viral subversion of immunogenic cell death. Cell Cycle 2009, 8, 860–869. [Google Scholar] [CrossRef]
- Krysko, D.V.; Ravichandran, K.S.; VAndenabeele, P. Macrophages regulate the clearance of living cells by calreticulin. Nat. Commun. 2018, 9, 4644. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Obeid, M.; Tesniere, A.; Ghiringhelli, F.; Fimia, G.M.; Apetoh, L.; Perfettini, J.-L.; Castedo, M.; Mignot, G.; Panaretakis, T.; Casares, N.; et al. Calreticulin exposure dictates the immunogenicity of cancer cell death. Nat. Med. 2007, 13, 54–61. [Google Scholar] [CrossRef] [PubMed]
- Xu, Z.; Yang, Y.; Zhou, J.; Huang, Y.; Wang, Y.; Zhang, Y.; Lan, Y.; Liang, J.; Liu, X.; Zhong, N.; et al. Role of plasma calreticulin in the prediction of severity in septic patients. Dis. Markers 2019, 2019, 8792640. [Google Scholar] [CrossRef]
- Gold, L.I.; Eggleton, P.; Sweetwyne, M.T.; Van Duyn, L.B.; Greives, M.R.; Naylor, S.M.; Michalak, M.; Murphy-Ullrich, J.E. Calreticulin: Non-endoplamic reticulum functions in physiology and disease. FASEB J. 2009, 24, 665–683. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kwon, M.S.; Park, C.S.; Choi, K.r.; Park, C.-S.; Ahnn, J.; Kim, J., II; Eom, S.H.; Kaufman, S.J.; Song, W.K. Calreticulin couples calcium release and calcium influx in integrin-mediatd calcium signaling. Mol. Biol. Cell 2000, 11, 1433–1443. [Google Scholar] [CrossRef] [Green Version]
- Klune, J.R.; Dhuper, R.; Cardinal, J.; Billiar, T.R.; Tsung, A. HMGB1, Endogenous Danger Signaling. Mol. Med. 2008, 14, 476–484. [Google Scholar] [CrossRef]
- Kang, R.; Chen, R.; Zhag, Q.; Hou, W.; Wu, S.; Cao, L.; Huang, J.; Yu, Y.; Fan, Y.-G.; Yan, Z.; et al. HMGB1 in health and disease. Mol. Aspects Med. 2008, 40, 1–116. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kazama, H.; Ricci, J.-E.; Herndon, J.M.; Hoppe, G.; Gren, D.R.; Ferguson, T.A. Induction of immunological tolerance by apoptotic cells requires caspase-dependent oxidation of high-mobility group Box-1 protein. Immunity 2008, 29, 21–32. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, C.; Zhang, Y.; Cheng, X.; Yuan, H.; Zhu, S.; Liu, J.; Wen, Q.; Xie, Y.; Liu, J.; Kroemer, G.; et al. PINK1 and PARK2 suppress pancreatic tumorigenesis through control of mitochondrial iron-mediated immunometabolism. Dev. Cell. 2018, 46, 441–455.e448. [Google Scholar] [CrossRef] [Green Version]
- Yu, Y.; Tang, D.; Kang, R. Oxidative stress-mediated HMGB1 biology. Front. Physiol. 2015, 6, 93. [Google Scholar] [CrossRef] [Green Version]
- Galluzzi, L.; Buque, A.; Kepp, O.; Zitvogel, L.; Kroemer, G. Immunogenic cell death in cancer and infectious disease. Nat. Rev. Immunol. 2017, 17, 97–111. [Google Scholar] [CrossRef]
- Medina, C.B.; Ravichandran, K.S. Do not let death do us part-‘find-me’ signals in communication between dying cells and the phygocytes. Cell Death Differ. 2016, 23, 979–989. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Michaud, M.; Martins, I.; Sukkurwala, A.Q.; Adjemian, S.; Ma, Y.; Pellegatti, P.; Shen, S.; Kepp, O.; Scoazec, M.; Mignot, G.; et al. Autophagy-dependent anticancer immune responses induced by chemotherapeutic agents in mice. Science 2011, 334, 1573–1577. [Google Scholar] [CrossRef]
- Land, W.G. The role of damage-associated molecular patterns (DAMPs) in human diseases. Sultan Qaboos Univ. Med. J. 2015, 15, e157–e170. [Google Scholar]
- Hernandez, C.; Huebener, P.; Schwabe, R.F. Damage-associated molecular patterns in cancer: A double-edged sword. Oncogene 2016, 35, 5931–5941. [Google Scholar] [CrossRef] [PubMed]
- Sangiuliano, B.; Perez, N.M.; Moreira, D.F.; Belizario, J.E. Cell death-associated molecular-pattern molecules: Inflammatory signaling and control. Mediat. Inflamm. 2014, 2014, 821043. [Google Scholar] [CrossRef] [PubMed]
- Hegyi, G.; Vincze, G.; Szasz, A. On the Dynamic Equilibrium in Homeostasis. Open J. Biophys. 2012, 2, 64–71. [Google Scholar] [CrossRef] [Green Version]
- Szigeti, G.P.; Szasz, A.M.; Szasz, A. The growth of healthy and cancerous tissues. Open J. Biophys. 2020, 10, 113–128. [Google Scholar] [CrossRef]
- Levin, M. Large-scale biophysics: Ion flows and regeneration. Trends Cell Biol. 2007, 17, 261–269. [Google Scholar] [CrossRef] [PubMed]
- Szasz, O.; Szigeti, G.P.; Szasz, A.; Benyo, Z. Role of electrical forces in angiogenesis. Open J. Biophys. 2018, 8, 49–67. [Google Scholar] [CrossRef] [Green Version]
- Derer, A.; Deloch, L.; Rubner, Y.; Fietkau, R.; Frey, B.; Gaipl, U.S. Radio-immunotherapy-induced immunogenic cancer cells as basis for induction of systemic anti-tumor immune responses–pre-clinical evidence and ongoing clinical applications. Front. Immunol. 2015, 6, 505. [Google Scholar] [CrossRef] [Green Version]
- Stagg, A.J.; Knight, S.C. Antigen-presenting cells. Nature 2001, 1–8. Available online: http://labs.icb.ufmg.br/lbcd/pages2/bernardo/Bernardo/Artigos/Antigen-presenting%20Cells.pdf (accessed on 7 October 2020).
- Kepp, O.; Galluzzi, L.; Martins, I.; Schlemmer, F.; Adjemian, S.; Michaud, M.; Sukkurwala, A.Q.; Menger, L.; Zitvogel, L.; Kroemer, G. Molecular determinants of immunogenic cell death elicited by anticancer chemotherapy. Cancer Metastasis Rev. 2011, 30, 61–69. [Google Scholar] [CrossRef] [PubMed]
- Sachamitr, P.; Fairchild, P.J. Cross presentation of antigen by dendritic cells: Mechanisms and implications for immunotherapy. Expert. Rev. Clin. Immunol. 2012, 8, 547–555. [Google Scholar] [CrossRef]
- Chi, K.-W. Tumor-Directed Immunotherapy: Combined Radiotherapy and Oncothermia. In Proceedings of the 36th Conference of the International Clinical Hyperthermia Society, Budapest, Hungary, 28–29 September 2018. [Google Scholar]
- Holtmeier, W.; Kabelitz, D. γδ T-cells link innate and adaptive immune responses. Chem. Immunol. Allergy 2005, 86, 151–183. [Google Scholar]
- Andocs, G.; Szasz, A.; Szasz, I.; Szasz, N. Tumor. Vaccination Patent U.S. 2015/0217099, 6 October 2020. Available online: http://www.freepatentsonline.com/20150217099.pdf (accessed on 10 January 2021).
























{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Tumor (A2058 Melanoma (24 h)) | Dead Area (%) | FI Signal Intensity | cCas3 (%) | ||||||
|---|---|---|---|---|---|---|---|---|---|
| mEHT + Primary NK | mEHT + NK92MI | mEHT Alone | mEHT + Primary NK | mEHT + NK92MI | mEHT Alone | mEHT + Primary NK | mEHT + NK92MI | mEHT Alone | |
| Untreated control (%) | 5 | 9 | 7 | 0.9 | 0.8 | 0.9 | 5 | 6 | 8.5 |
| Treated by mEHT (%) | 63 | 90 | 60 | 1.35 | 1.39 | 1.26 | 26 | 33 | 19 |
| Significance | *** | ** | *** | ** | *** | ||||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Minnaar, C.A.; Szasz, A. Forcing the Antitumor Effects of HSPs Using a Modulated Electric Field. Cells 2022, 11, 1838. https://doi.org/10.3390/cells11111838
Minnaar CA, Szasz A. Forcing the Antitumor Effects of HSPs Using a Modulated Electric Field. Cells. 2022; 11(11):1838. https://doi.org/10.3390/cells11111838
Chicago/Turabian StyleMinnaar, Carrie Anne, and Andras Szasz. 2022. "Forcing the Antitumor Effects of HSPs Using a Modulated Electric Field" Cells 11, no. 11: 1838. https://doi.org/10.3390/cells11111838