
Potential Variables for Improved Reproducibility of Neuronal Cell Grafts at Stroke Sites

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
3. Results
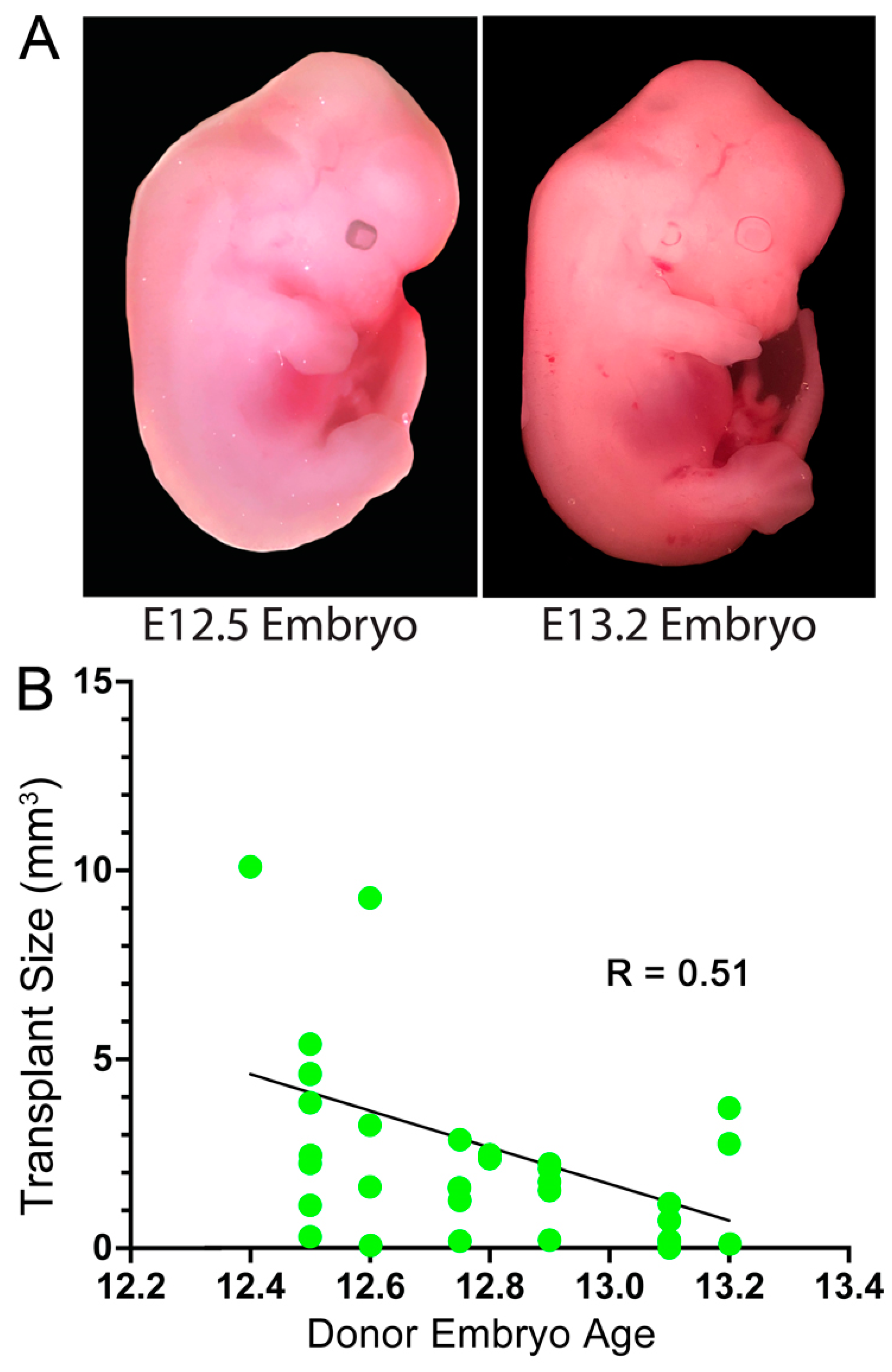
3.1. Embryonic Stage of the Donor Cells
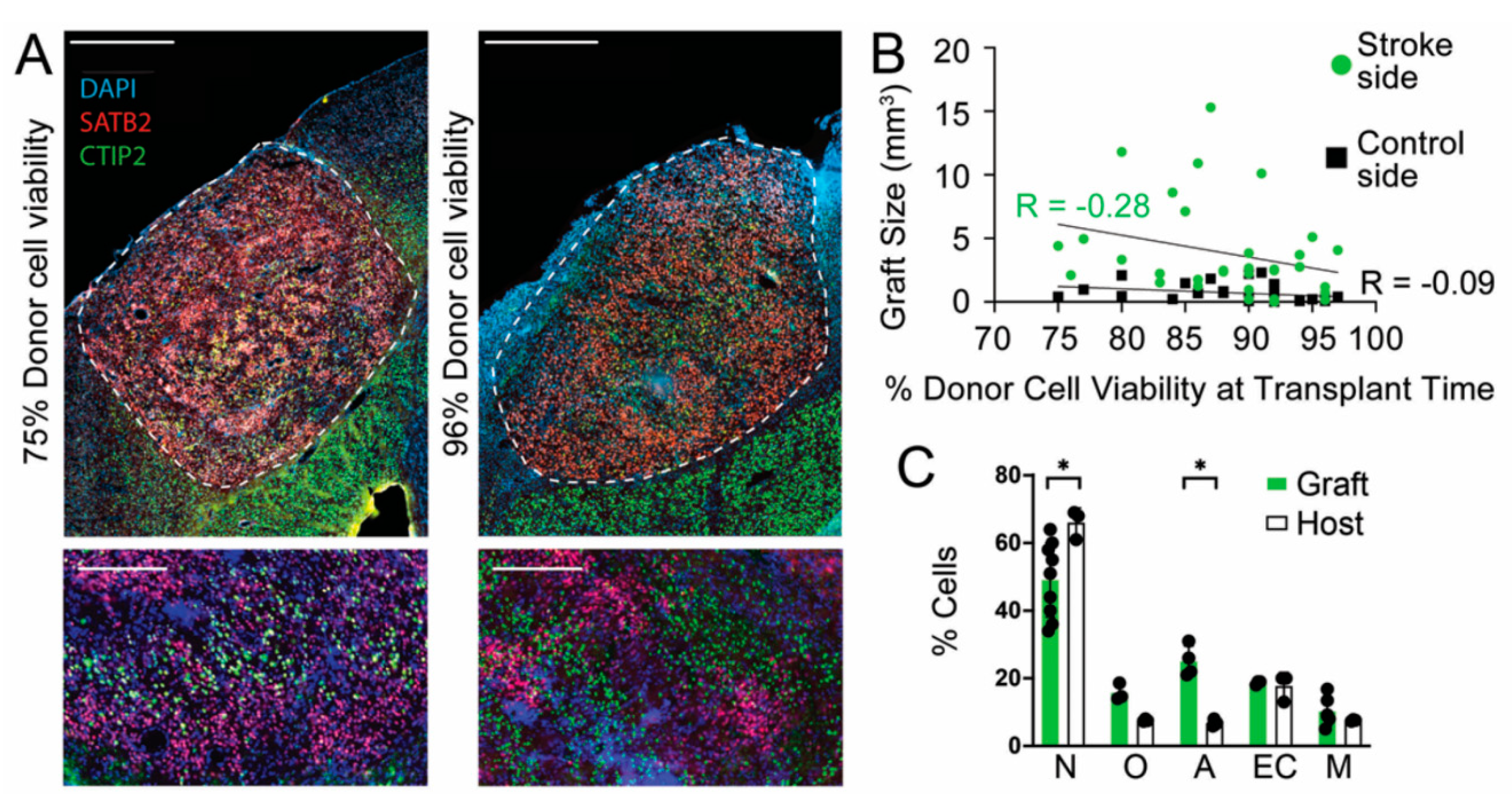
3.2. Donor Cell Viability at Time of Transplantation
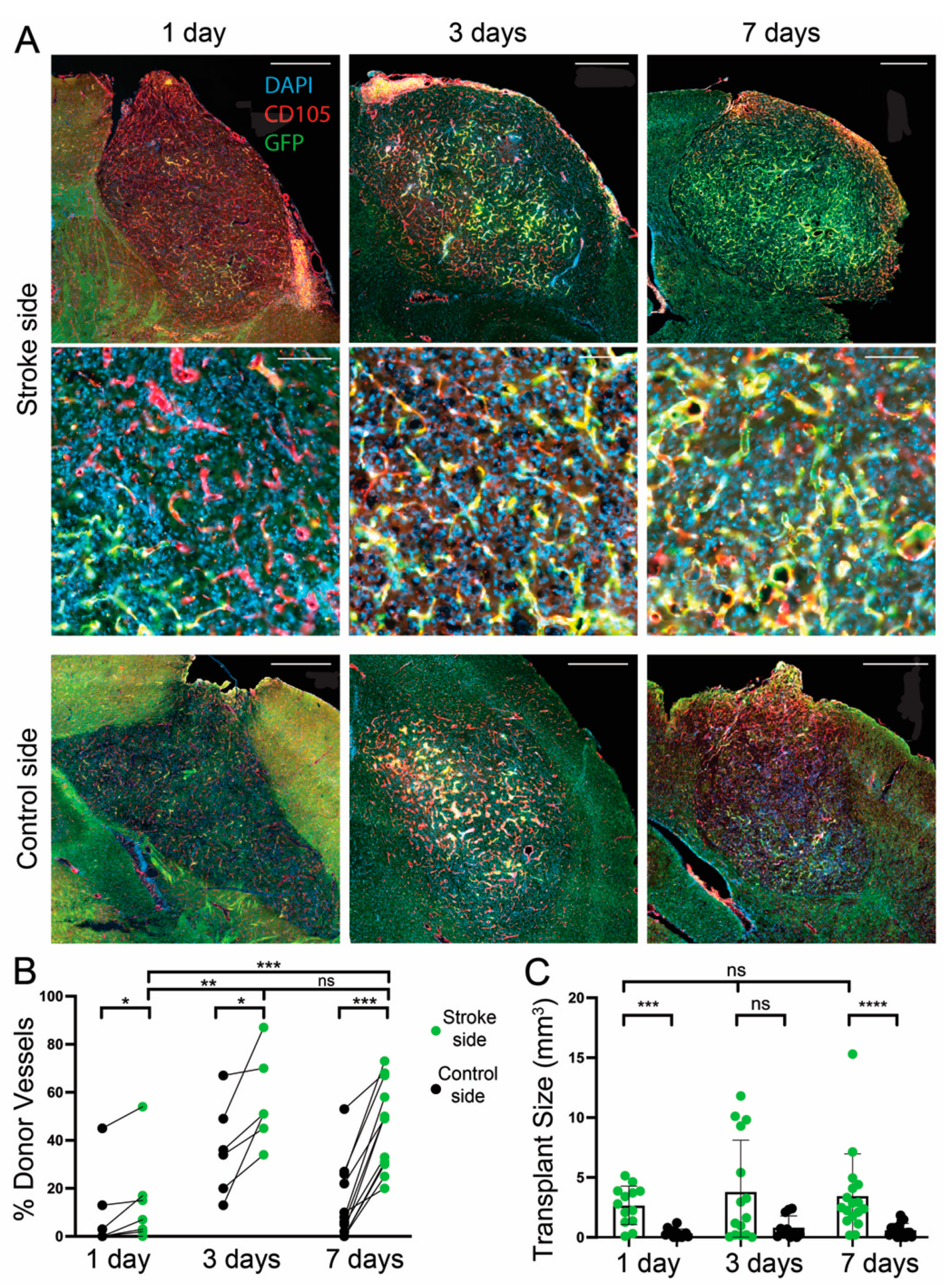
3.3. Effect of a Transplantation Delay of 1, 3, and 7 Days after dMCAO
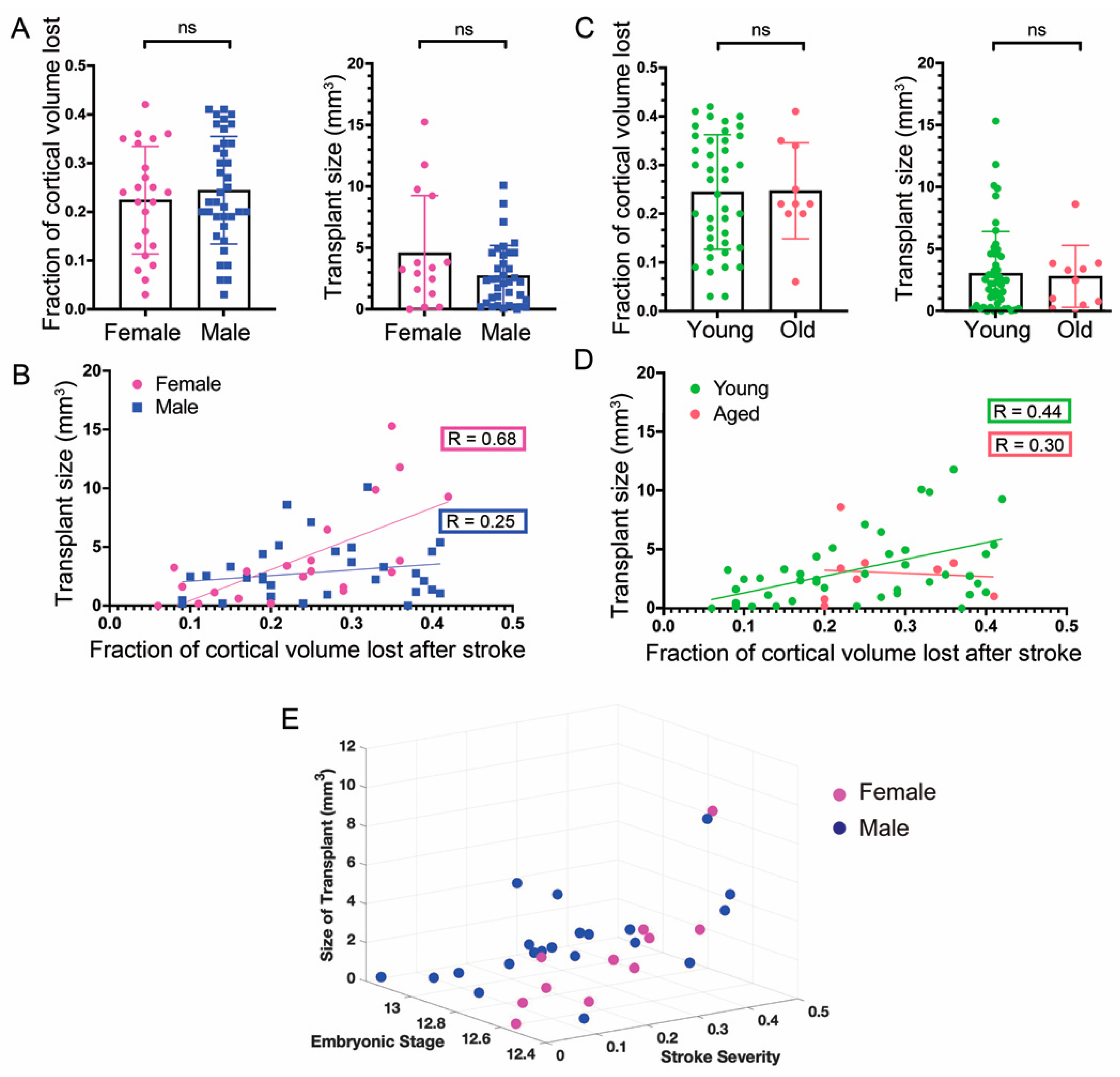
3.4. Sex of the Host Mice
3.5. Age of Hosts
3.6. General Linear Model of Stroke Severity and Embryonic Stage of Donor Cells
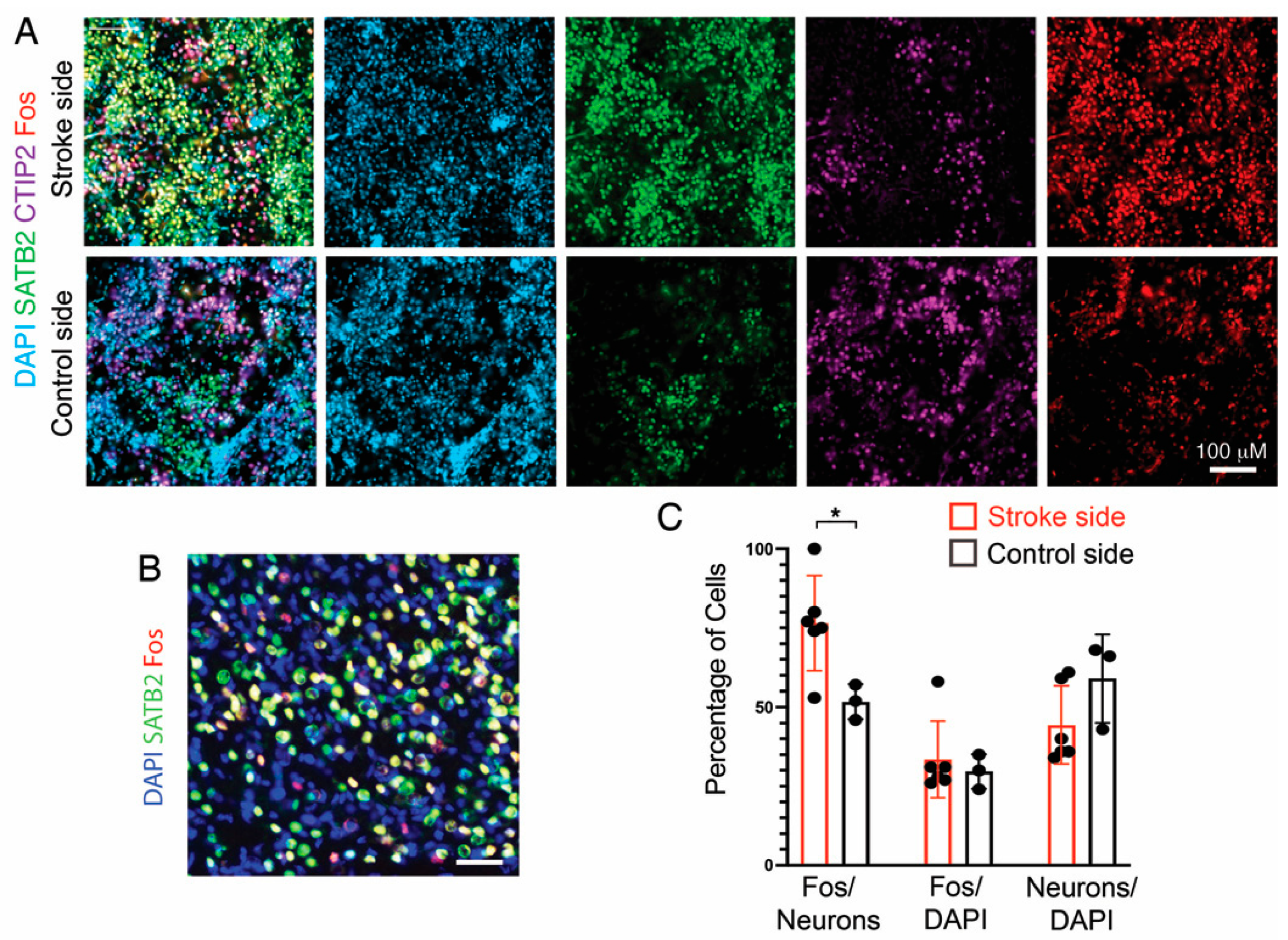
3.7. Fos Expression as a Proxy for Neuronal Activity
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Feigin, V.L.; GBD 2019 Stroke Collaborators. Global, regional, and national burden of stroke and its risk factors, 1990-2019: A systematic analysis for the global burden of disease study 2019. Lancet Neurol. 2021, 20, 795–820. [Google Scholar] [CrossRef]
- Pu, H.; Shi, Y.; Zhang, L.; Lu, Z.; Ye, Q.; Leak, R.K.; Xu, F.; Ma, S.; Mu, H.; Wei, Z.; et al. Protease-independent action of tissue plasminogen activator in brain plasticity and neurological recovery after ischemic stroke. Proc. Natl. Acad. Sci. USA 2019, 116, 9115–9124. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bang, O.Y. Clinical Trials of Adult Stem Cell Therapy in Patients with Ischemic Stroke. J. Clin. Neurol. 2016, 12, 14–20. [Google Scholar] [CrossRef] [PubMed]
- Rust, R.; Gantner, C.; Schwab, M.E. Pro- and antiangiogenic therapies: Current status and clinical implications. FASEB J. 2019, 33, 34–48. [Google Scholar] [CrossRef] [PubMed]
- Kokaia, Z.; Tornero, D.; Lindvall, O. Transplantation of reprogrammed neurons for improved recovery after stroke. Prog. Brain Res. 2017, 231, 245–263. [Google Scholar] [CrossRef]
- Rikhtegar, R.; Yousefi, M.; Dolati, S.; Kasmaei, H.D.; Charsouei, S.; Nouri, M.; Shakouri, S.K. Stem cell-based cell therapy for neuroprotection in stroke: A review. J. Cell. Biochem. 2019, 120, 8849–8862. [Google Scholar] [CrossRef]
- Stonesifer, C.; Corey, S.; Ghaneka, S.; Diamandis, Z.; Acost, A.S.; Brolongan, C.V. Stem cell therapy for abrogating stroke-induced neuroinflammation and relevant secondary cell death mechanisms. Prog. Neurobiol. 2017, 158, 94–131. [Google Scholar] [CrossRef]
- Espuny-Camacho, I.; Michelsen, K.A.; Gall, D.; Linaro, D.; Hasche, A.; Bonnefont, J.; Bali, C.; Orduz, D.; Bilheu, A.; Herpoel, A.; et al. Pyramidal Neurons Derived from Human Pluripotent Stem Cells Integrate Efficiently into Mouse Brain Circuits In Vivo. Neuron 2013, 77, 440–456. [Google Scholar] [CrossRef] [Green Version]
- Falkner, S.; Grade, S.; Dimou, L.; Conzelmann, K.-K.; Bonhoeffer, T.; Götz, M.; Hübener, M. Transplanted embryonic neurons integrate into adult neocortical circuits. Nature 2016, 539, 248–253. [Google Scholar] [CrossRef]
- Michelsen, K.A.; Acosta-Verdugo, S.; Benoit-Marand, M.; Camacho, I.E.; Gaspard, N.; Saha, B.; Gaillard, A.; Vanderhaeghen, P. Area-Specific Reestablishment of Damaged Circuits in the Adult Cerebral Cortex by Cortical Neurons Derived from Mouse Embryonic Stem Cells. Neuron 2015, 85, 982–997. [Google Scholar] [CrossRef] [Green Version]
- Tang, Y.-H.; Ma, Y.-Y.; Zhang, Z.-J.; Wang, Y.-T.; Yang, G.-Y. Opportunities and Challenges: Stem Cell-Based Therapy for the Treatment of Ischemic Stroke. CNS Neurosci. Ther. 2015, 21, 337–347. [Google Scholar] [CrossRef] [PubMed]
- Krzyspiak, J.; Yan, J.; Ghosh, H.S.; Galinski, B.; Lituma, P.; Alvina, K.; Quezada, A.; Kee, S.; Grońska-Pęski, M.; Tai, Y.D.; et al. Donor-derived vasculature supports neocortical cell grafts after stroke. Stem Cell Res. 2022, 59, 102642. [Google Scholar] [CrossRef] [PubMed]
- Gaillard, A.; Prestoz, L.; Dumartin, B.; Cantereau, A.; Morel, F.; Roger, M.; Jaber, M. Reestablishment of damaged adult motor pathways by grafted embryonic cortical neurons. Nat. Neurosci. 2007, 10, 1294–1299. [Google Scholar] [CrossRef] [PubMed]
- Péron, S.; Droguerre, M.; Debarbieux, F.; Ballout, N.; Benoit-Marand, M.; Francheteau, M.; Brot, S.; Rougon, G.; Jaber, M.; Gaillard, A. A Delay between Motor Cortex Lesions and Neuronal Transplantation Enhances Graft Integration and Improves Repair and Recovery. J. Neurosci. 2017, 37, 1820–1834. [Google Scholar] [CrossRef] [Green Version]
- Saga, Y.; Miyagawa-Tomita, S.; Takagi, A.; Kitajima, S.; Miyazaki, J.; Inoue, T. MesP1 is expressed in the heart precursor cells and required for the formation of a single heart tube. Development 1999, 126, 3437–3447. [Google Scholar] [CrossRef]
- Sousa, V.H.; Miyoshi, G.; Hjerling-Leffler, J.; Karayannis, T.; Fishell, G. Characterization of Nkx6-2-derived neocortical interneuron lineages. Cereb. Cortex. 2009, 19 (Suppl. 1), i1–i10. [Google Scholar] [CrossRef] [Green Version]
- Loo, L.; Simon, J.M.; Xing, L.; McCoy, E.S.; Niehaus, J.K.; Guo, J.; Anton, E.S.; Zylka, M.J. Single-cell transcriptomic analysis of mouse neocortical development. Nat. Commun. 2019, 10, 134. [Google Scholar] [CrossRef] [Green Version]
- Zudaire, E.; Gambardella, L.; Kurcz, C.; Vermeren, S. A Computational Tool for Quantitative Analysis of Vascular Networks. PLoS ONE 2011, 6, e27385. [Google Scholar] [CrossRef] [Green Version]
- Wong, M.D.; van Eede, M.C.; Spring, S.; Jevtic, S.; Boughner, J.C.; Lerch, J.P.; Henkelman, R.M. 4D atlas of the mouse embryo for precise morphological staging. Development 2015, 142, 3583–3591. [Google Scholar] [CrossRef] [Green Version]
- Hanashima, C.; Shen, L.; Li, S.C.; Lai, E. Brain factor-1 controls the proliferation and differentiation of neocortical progenitor cells through independent mechanisms. J. Neurosci. 2002, 22, 6526–6536. [Google Scholar] [CrossRef] [Green Version]
- Jiang, M.Q.; Zhao, Y.-Y.; Cao, W.; Wei, Z.Z.; Gu, X.; Wei, L.; Yu, S.P. Long-term survival and regeneration of neuronal and vasculature cells inside the core region after ischemic stroke in adult mice. Brain Pathol. 2017, 27, 480–498. [Google Scholar] [CrossRef] [PubMed]
- Hayashi, T.; Noshita, N.; Sugawara, T.; Chan, P.H. Temporal Profile of Angiogenesis and Expression of Related Genes in the Brain After Ischemia. J. Cereb. Blood Flow Metab. 2003, 23, 166–180. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ballout, N.; Rochelle, T.; Brot, S.; Bonnet, M.-L.; Francheteau, M.; Prestoz, L.; Zibara, K.; Gaillard, A. Characterization of Inflammation in Delayed Cortical Transplantation. Front. Mol. Neurosci. 2019, 12, 160. [Google Scholar] [CrossRef] [Green Version]
- Roy-O’Reilly, M.; McCullough, L.D. Age and Sex Are Critical Factors in Ischemic Stroke Pathology. Endocrinology 2018, 159, 3120–3131. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, R.-L.; Balami, J.S.; Esiri, M.M.; Chen, L.-K.; Buchan, A.M. Ischemic stroke in the elderly: An overview of evidence. Nat. Rev. Neurol. 2010, 6, 256–265. [Google Scholar] [CrossRef]
- Popa-Wagner, A.; Badan, I.; Walker, L.; Groppa, S.; Patrana, N.; Kessler, C. Accelerated infarct development, cytogenesis and apoptosis following transient cerebral ischemia in aged rats. Acta Neuropathol. 2007, 113, 277–293. [Google Scholar] [CrossRef]
- Hudson, A.E. Genetic Reporters of Neuronal Activity: C-Fos and G-CaMP6. Methods Enzymol. 2018, 603, 197–220. [Google Scholar] [CrossRef]
- Luhmann, H.J.; Sinning, A.; Yang, J.W.; Reyes-Puerta, V.; Stuttgen, M.C.; Kirischuk, S.; Kilb, W. Spontaneous neuronal activity in developing neocortical networks: From single cells to large-scale interactions. Front. Neural Circ. 2016, 10, 40. [Google Scholar] [CrossRef] [Green Version]
- Molnar, Z.; Luhmann, H.J.; Kanold, P.O. Transient cortical circuits match spontaneous and sensory-driven activity during development. Science 2020, 370, eabb2153. [Google Scholar] [CrossRef]
- Darsalia, V.; Allison, S.J.; Cusulin, C.; Monni, E.; Kuzdas, D.; Kallur, T.; Lindvall, O.; Kokaia, Z. Cell number and timing of transplantation determine survival of human neural stem cell grafts in stroke-damaged rat brain. J. Cereb. Blood Flow Metab. 2011, 31, 235–242. [Google Scholar] [CrossRef] [Green Version]
- Li, H.; Zhang, N.; Lin, H.Y.; Yu, Y.; Cai, Q.Y.; Ma, L.; Ding, S. Histological, cellular and behavioral assessments of stroke outcomes after photothrombosis-induced ischemia in adult mice. BMC Neurosci. 2014, 15, 58. [Google Scholar] [CrossRef] [Green Version]
- Buga, A.-M.; Sascau, M.; Pisoschi, C.; Herndon, J.G.; Kessler, C.; Popa-Wagner, A. The genomic response of the ipsilateral and contralateral cortex to stroke in aged rats. J. Cell. Mol. Med. 2008, 12, 2731–2753. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Villeda, S.A.; Plambeck, K.E.; Middeldorp, J.; Castellano, J.M.; Mosher, K.I.; Luo, J.; Smith, L.K.; Bieri, G.; Lin, K.; Berdnik, D.; et al. Young blood reverses age-related impairments in cognitive function and synaptic plasticity in mice. Nat. Med. 2014, 20, 659–663. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yousef, H.; Czupalla, C.J.; Lee, D.; Chen, M.B.; Burke, A.N.; Zera, K.A.; Zandstra, J.; Berber, E.; Lehallier, B.; Mathur, V.; et al. Aged blood impairs hippocampal neural precursor activity and activates microglia via brain endothelial cell VCAM1. Nat. Med. 2019, 25, 988–1000. [Google Scholar] [CrossRef]
- Dotson, A.L.; Wang, J.; Saugstad, J.; Murphy, S.J.; Offner, H. Splenectomy reduces infarct volume and neuroinflammation in male but not female mice in experimental stroke. J. Neuroimmunol. 2014, 278, 289–298. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Androvic, P.; Kirdajova, D.; Tureckova, J.; Zucha, D.; Rohlova, E.; Abaffy, P.; Kriska, J.; Valny, M.; Anderova, M.; Kubista, M.; et al. Decoding the Transcriptional Response to Ischemic Stroke in Young and Aged Mouse Brain. Cell Rep. 2020, 31, 107777. [Google Scholar] [CrossRef]
- Buga, A.M.; Margaritescu, C.; Scholz, C.J.; Radu, E.; Zelenak, C.; Popa-Wagner, A. Transcriptomics of post-stroke angiogenesis in the aged brain. Front. Aging Neurosci. 2014, 6, 44. [Google Scholar] [CrossRef] [PubMed]





Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Krzyspiak, J.; Khodakhah, K.; Hébert, J.M. Potential Variables for Improved Reproducibility of Neuronal Cell Grafts at Stroke Sites. Cells 2022, 11, 1656. https://doi.org/10.3390/cells11101656
Krzyspiak J, Khodakhah K, Hébert JM. Potential Variables for Improved Reproducibility of Neuronal Cell Grafts at Stroke Sites. Cells. 2022; 11(10):1656. https://doi.org/10.3390/cells11101656
Chicago/Turabian StyleKrzyspiak, Joanna, Kamran Khodakhah, and Jean M. Hébert. 2022. "Potential Variables for Improved Reproducibility of Neuronal Cell Grafts at Stroke Sites" Cells 11, no. 10: 1656. https://doi.org/10.3390/cells11101656