
5′-Nucleotidase Plays a Key Role in Uric Acid Metabolism of Bombyx mori

 ,
, 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Silkworm Strain
2.2. Phylogenetic Analysis
2.3. Qualitative and Quantitative Real-Time PCR
2.4. Plasmid Construction
2.5. Germline Transformation
2.6. CRISPR/Cas9 Genome Editing
2.7. Mutagenesis Analysis
2.8. Quantification of UA Content
2.9. Feeding Experiments
2.10. LC-MS Sample Preparation and Experiments
2.11. Metabolomics Data Analysis
2.12. RNA Sequencing and Analysis
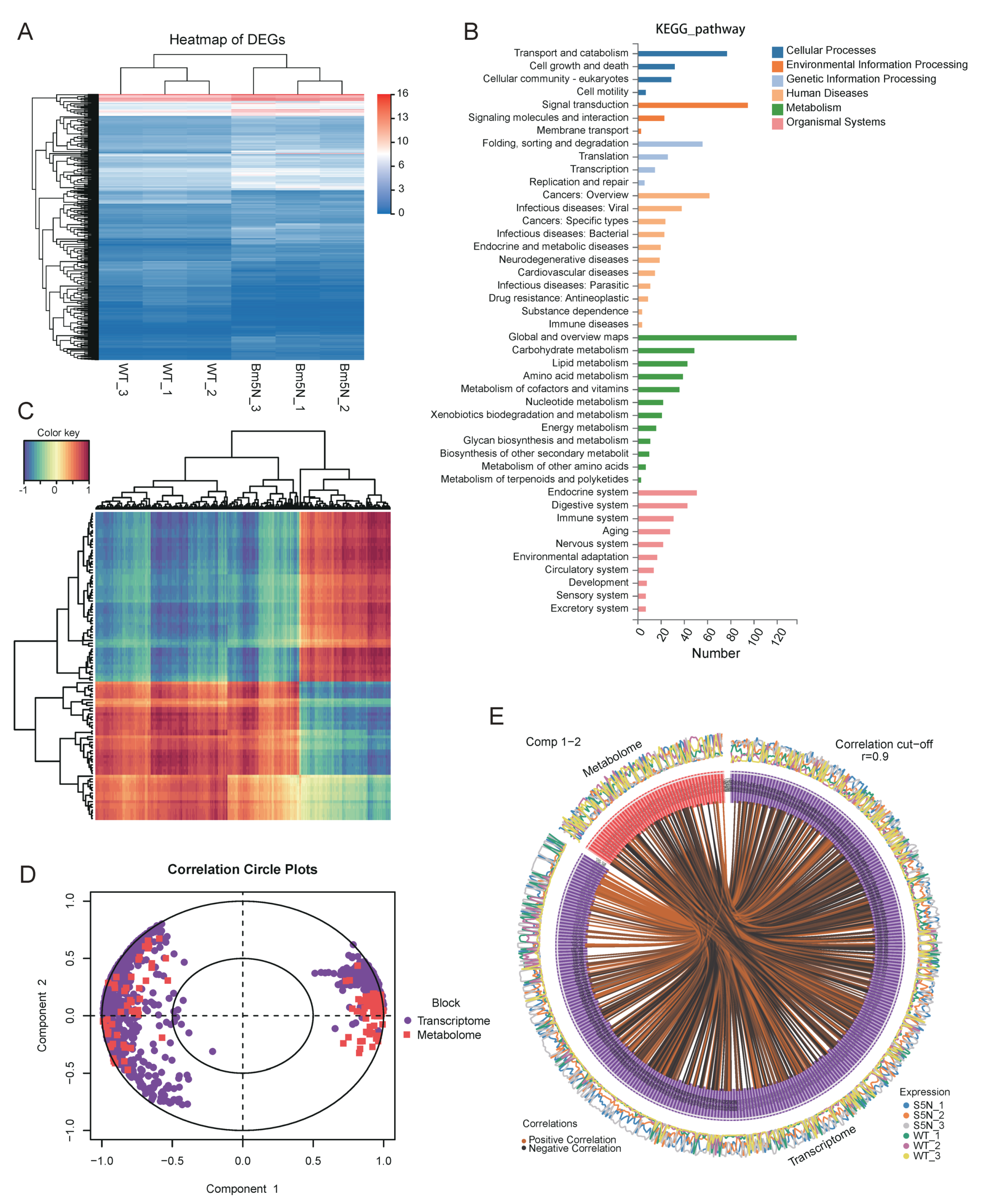
2.13. The Metabolites and Genes Correlation Analysis
2.14. Statistical Analysis
3. Results
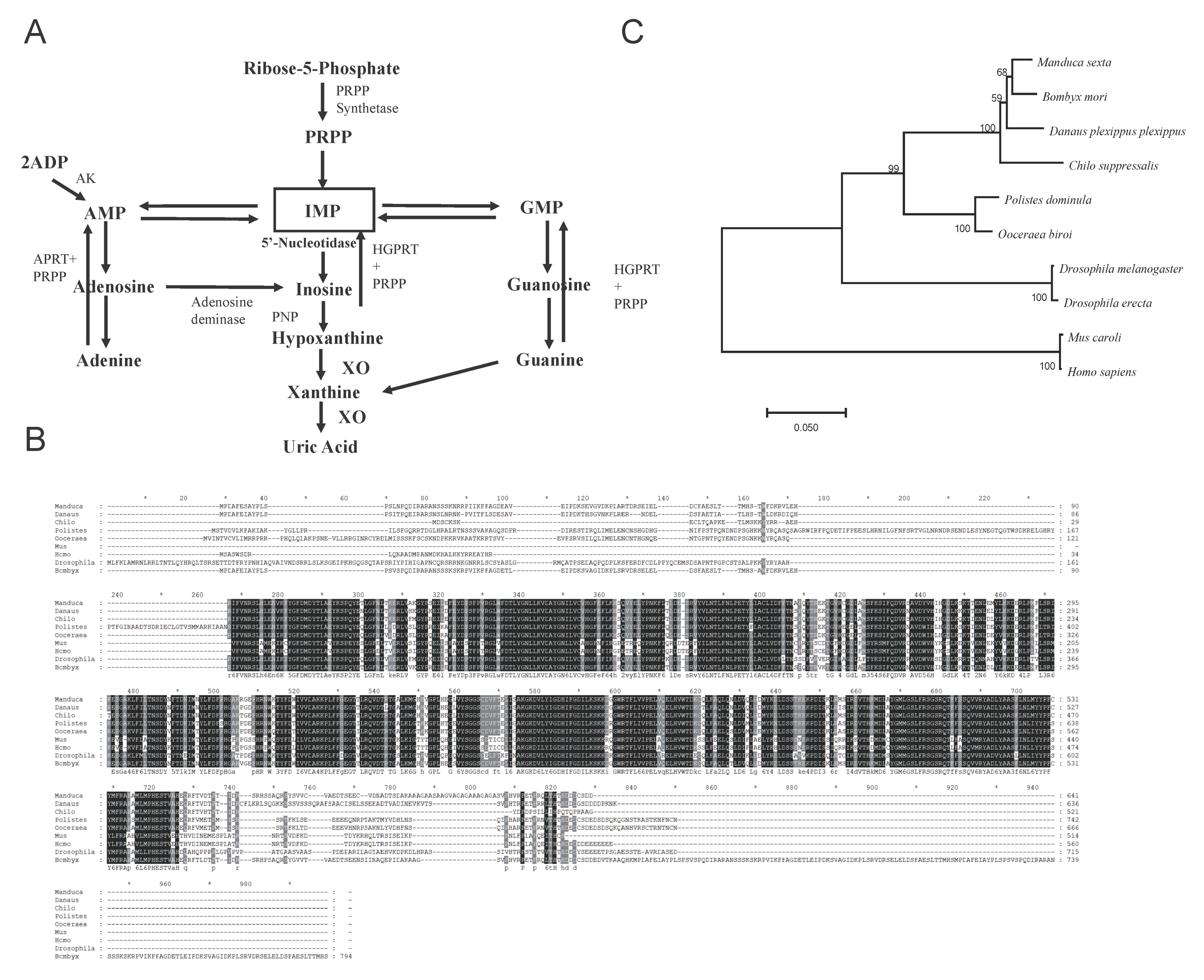
3.1. Phylogenetic Identification of Bm5′N
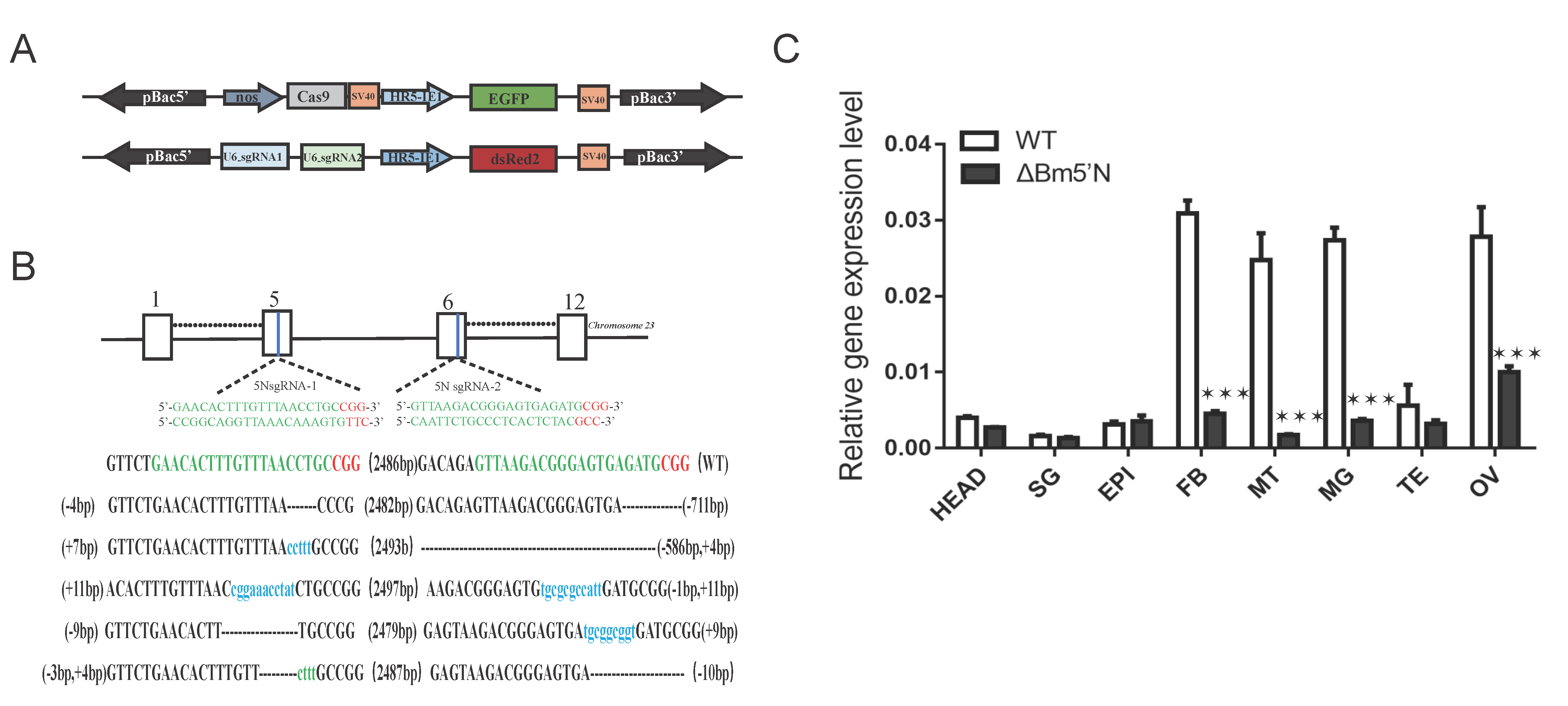
3.2. Targeted Mutagenesis Using the CRISPR/Cas9 System
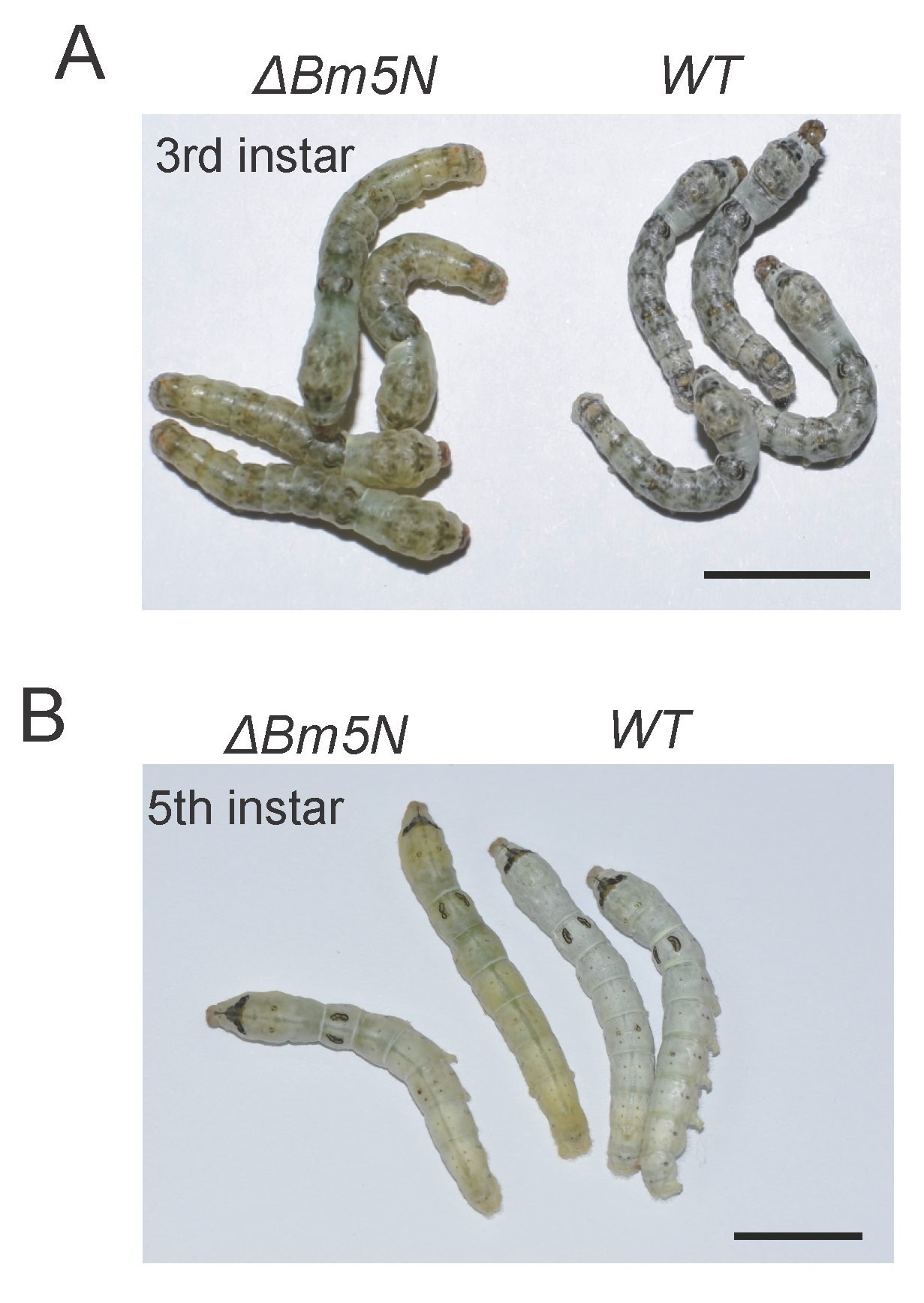
3.3. Loss Function of Bm5′N Results in Translucency of the Larval Integument
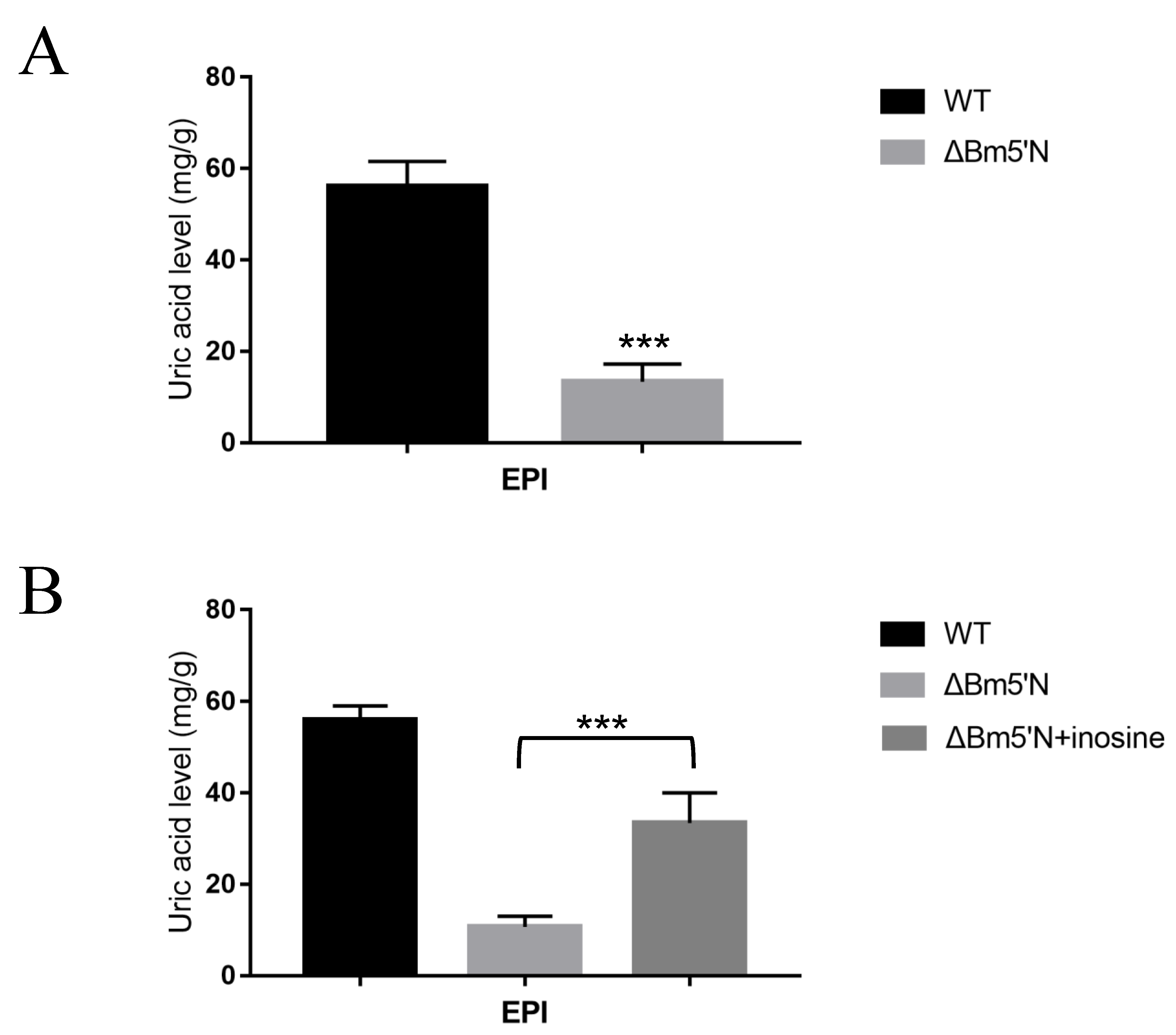
3.4. Feeding Experiments Indicate That Bm5′N Is Necessary for UA Biosynthesis
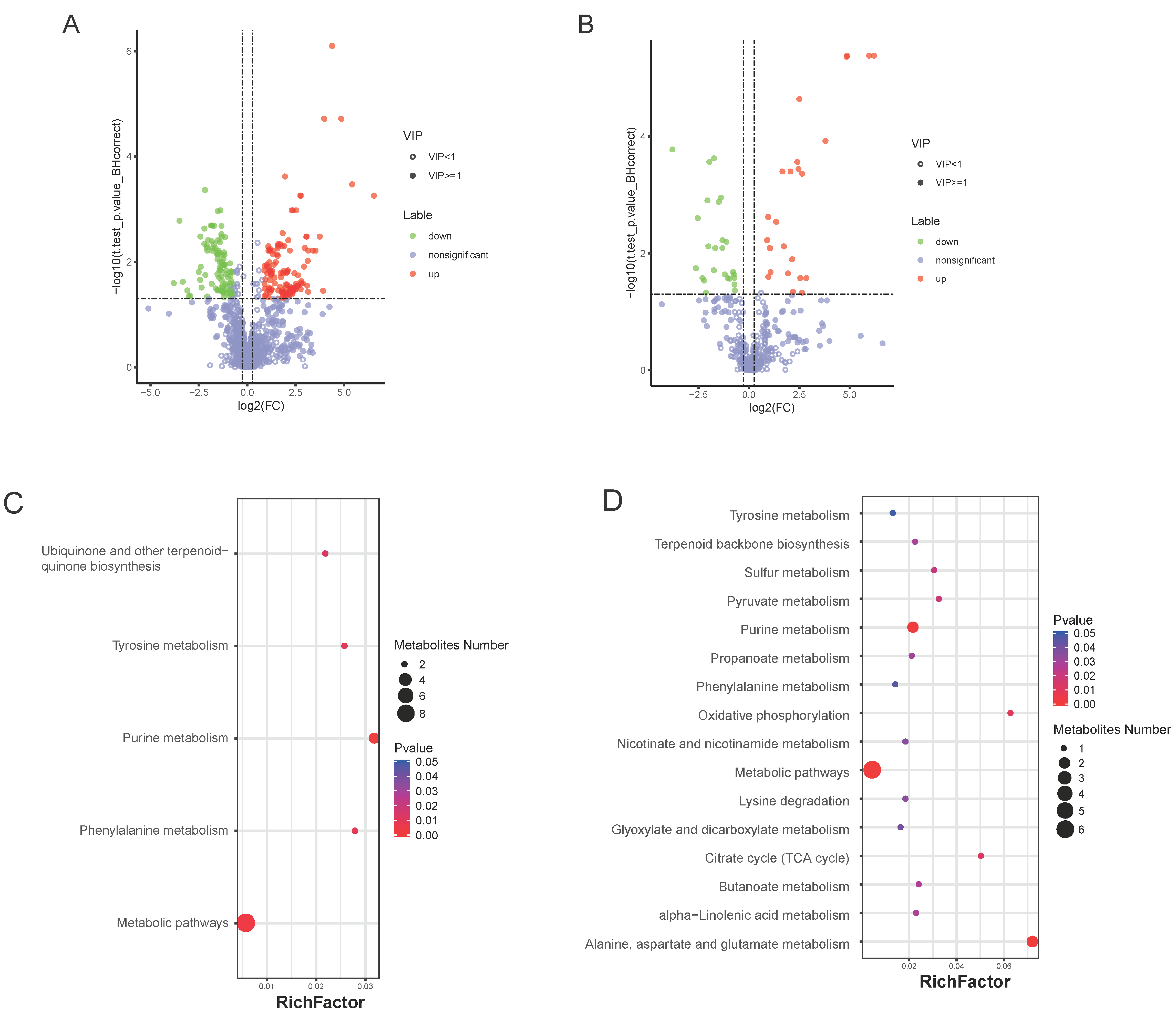
3.5. Bm5′N Mutant and WT Metabolite Profiles Differ in Purine Metabolism
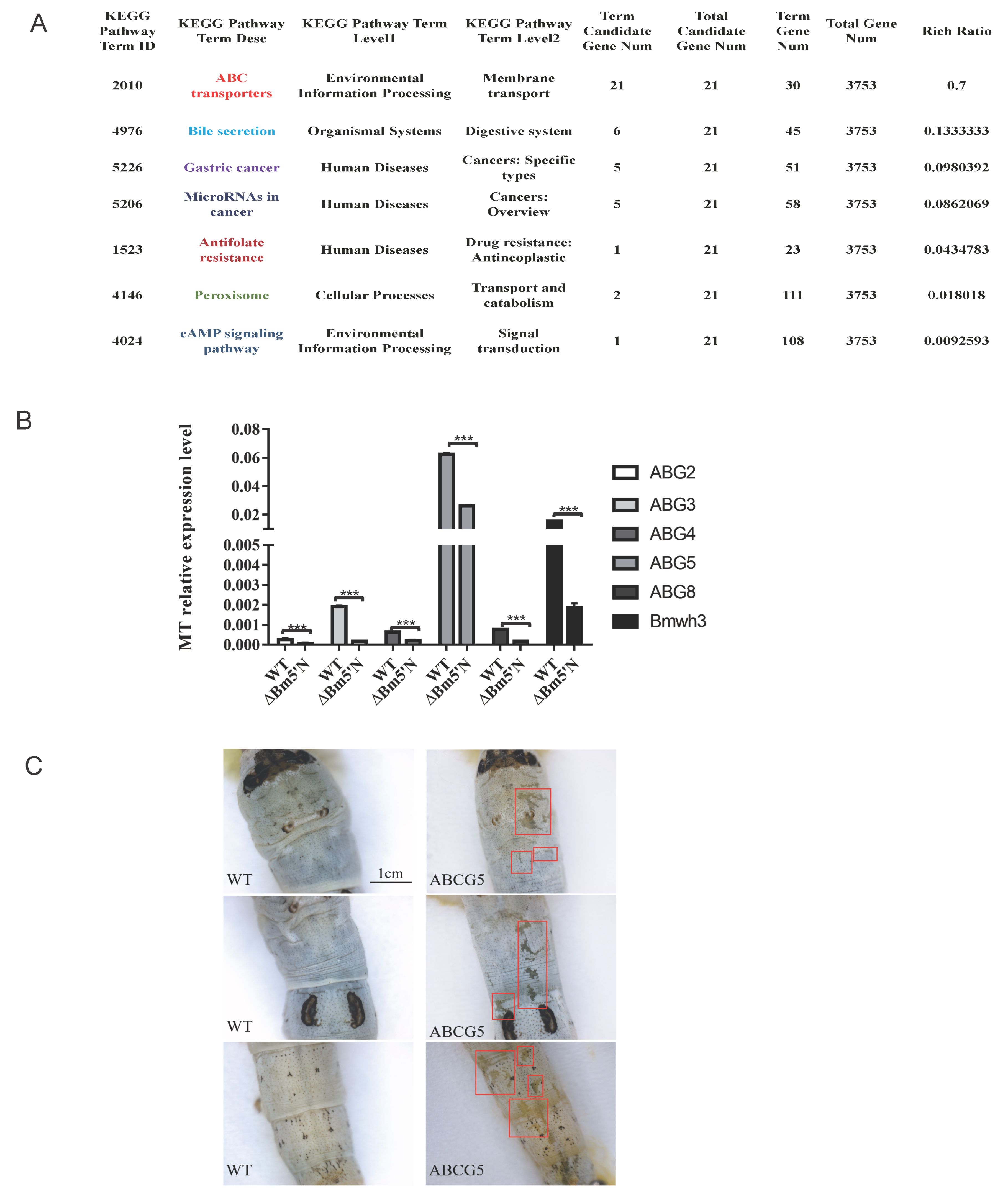
3.6. Level of ABCG Transporter Genes Were Down Regulated in Bm5′N Mutant
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Grassi, D.; Ferri, L.; Desideri, G.; Di Giosia, P.; Cheli, P.; Del Pinto, R.; Properzi, G.; Ferri, C. Chronic hyperuricemia, uric acid deposit and cardiovascular risk. Curr. Pharm. Des. 2013, 19, 2432–2438. [Google Scholar] [CrossRef] [Green Version]
- Tan, P.K.; Farrar, J.E.; Gaucher, E.A.; Miner, J.N. Coevolution of URAT1 and uricase during primate evolution: Implications for serum urate homeostasis and gout. Mol. Biol. Evol. 2016, 33, 2193–2200. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Watanabe, S.; Kang, D.H.; Feng, L.; Nakagawa, T.; Kanellis, J.; Lan, H.; Mazzali, M.; Johnson, R.J. Uric acid, hominoid evolution, and the pathogenesis of salt-sensitivity. Hypertension 2002, 40, 355–360. [Google Scholar] [CrossRef]
- Zhu, Y.; Pandya, B.J.; Choi, H.K. Comorbidities of gout and hyperuricemia in the US general population: NHANES 2007-2008. Am. J. Med. 2012, 125, 679–687. [Google Scholar] [CrossRef] [PubMed]
- Fisher, M.C.; Rai, S.K.; Lu, N.; Zhang, Y.; Choi, H.K. The unclosing premature mortality gap in gout: A general population-based study. Ann. Rheum. Dis. 2017, 76, 1289–1294. [Google Scholar] [CrossRef]
- Nakatochi, M.; Kanai, M.; Nakayama, A.; Hishida, A.; Kawamura, Y.; Ichihara, S.; Akiyama, M.; Ikezaki, H.; Furusyo, N.; Shimizu, S.; et al. Genome-wide meta-analysis identifies multiple novel loci associated with serum uric acid levels in Japanese individuals. Commun. Biol. 2019, 2, 115. [Google Scholar] [CrossRef]
- Lang, S.; Hilsabeck, T.A.; Wilson, K.A.; Sharma, A.; Bose, N.; Brackman, D.J.; Beck, J.N.; Chen, L.; Watson, M.A.; Killilea, D.W.; et al. A conserved role of the insulin-like signaling pathway in diet-dependent uric acid pathologies in Drosophila melanogaster. PLoS Genet. 2019, 15, e1008318. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lu, J.; Hou, X.; Yuan, X.; Cui, L.; Liu, Z.; Li, X.; Ma, L.; Cheng, X.; Xin, Y.; Wang, C.; et al. Knockout of the urate oxidase gene provides a stable mouse model of hyperuricemia associated with metabolic disorders. Kidney Int. 2018, 93, 69–80. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, X.; Wakamiya, M.; Vaishnav, S.; Geske, R.; Montgomery, C., Jr.; Jones, P.; Bradley, A.; Caskey, C.T. Hyperuricemia and urate nephropathy in urate oxidase-deficient mice. Proc. Natl. Acad. Sci. USA 1994, 91, 742–746. [Google Scholar] [CrossRef] [Green Version]
- Buckner, J.S.; Newman, S.M. Uric-acid storage in the epidermal-cells of Manduca-Sexta-Localization and movement during the larval pupal transformation. J. Insect Physiol. 1990, 36, 219–229. [Google Scholar] [CrossRef]
- Cochran, D.G. Comparative analysis of excreta and fat-body from various cockroach species. Comp. Biochem. Phys. A 1979, 64, 1–4. [Google Scholar] [CrossRef]
- He, Z.; Zhao, M.; Wang, C.Y.; Sun, L.; Jiang, Y.Y.; Feng, Y. Purine and uric acid contents of common edible insects in Southwest China. J. Insects Food Feed. 2019, 5, 293–299. [Google Scholar] [CrossRef]
- Lee, J.; Kiuchi, T.; Kawamoto, M.; Shimada, T.; Katsuma, S. Accumulation of uric acid in the epidermis forms the white integument of Samia ricini larvae. PLoS ONE 2018, 13, e0205758. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Kiuchi, T.; Hirayama, C.; Banno, Y.; Katsuma, S.; Shimada, T. A reexamination on the deficiency of riboflavin accumulation in Malpighian tubules in larval translucent mutants of the silkworm, Bombyx mori. Genetica 2018, 146, 425–431. [Google Scholar] [CrossRef]
- Fujii, T.; Banno, Y. Identification of a novel function of the silkworm integument in nitrogen metabolism: Uric acid is synthesized within the epidermal cells in B. mori. Insect Biochem. Mol. Biol. 2019, 105, 43–50. [Google Scholar] [CrossRef]
- Fujii, T.; Daimon, T.; Uchino, K.; Banno, Y.; Katsuma, S.; Sezutsu, H.; Tamura, T.; Shimada, T. Transgenic analysis of the BmBLOS2 gene that governs the translucency of the larval integument of the silkworm, Bombyx mori. Insect Biochem. Mol. Biol. 2010, 19, 659–667. [Google Scholar] [CrossRef] [PubMed]
- Fujii, T.; Yamamoto, K.; Banno, Y. Translucent larval integument and flaccid paralysis caused by genome editing in a gene governing molybdenum cofactor biosynthesis in Bombyx mori. Insect Biochem. Mol. Biol. 2018, 99, 11–16. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.Y.; Yin, Y.R.; Wang, K.; Cao, J.; Cheng, T.C.; Liu, C.; Zhang, Y.; Zhu, Y. Bombyx mori monocarboxylate transporter 9 (BmMCT9) is involved in the transport of uric acid in silkworm integument. Genes Cells 2020, 25, 33–40. [Google Scholar] [CrossRef]
- Sala-Newby, G.B.; Freeman, N.V.; Skladanowski, A.C.; Newby, A.C. Distinct roles for recombinant cytosolic 5’-nucleotidase-I and -II in AMP and IMP catabolism in COS-7 and H9c2 rat myoblast cell lines. J. Biol. Chem. 2000, 275, 11666–11671. [Google Scholar] [CrossRef] [Green Version]
- Kopff, M.; Zakrzewska, I.; FuchsKalinowska, J.; Klem, J.; Strzelczyk, M.; Puczkowski, S. 5’-Nucleotidase and adenosine deaminase activities in blood of patients with unstable angina pectoris. Haematologia 1997, 28, 223–231. [Google Scholar]
- Yoneyama, Y.; Suzuki, S.; Sawa, R.; Otsubo, Y.; Miura, A.; Kuwabara, Y.; Ishino, H.; Kiyokawa, Y.; Doi, D.; Kobayashi, H.; et al. Plasma 5’-nucleotidase activities and uric acid levels in women with pre-eclampsia. Gynecol. Obstet. Investig. 2002, 54, 168–171. [Google Scholar] [CrossRef] [PubMed]
- Tan, Y.; Liu, X.; Zhou, K.; He, X.; Lu, C.; He, B.; Niu, X.; Xiao, C.; Xu, G.; Bian, Z.; et al. The Potential Biomarkers to Identify the Development of Steatosis in Hyperuricemia. PLoS ONE 2016, 11, e0149043. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Y.; Zhang, H.; Chang, D.; Guo, F.; Pan, H.; Yang, Y. Metabolomics approach by (1)H NMR spectroscopy of serum reveals progression axes for asymptomatic hyperuricemia and gout. Arthritis Res. Ther. 2018, 20, 111. [Google Scholar] [CrossRef] [Green Version]
- Hoque, K.M.; Dixon, E.E.; Lewis, R.M.; Allan, J.; Gamble, G.D.; Phipps-Green, A.J.; Halperin Kuhns, V.L.; Horne, A.M.; Stamp, L.K.; Merriman, T.R.; et al. The ABCG2 Q141K hyperuricemia and gout associated variant illuminates the physiology of human urate excretion. Nat. Commun. 2020, 11, 2767. [Google Scholar] [CrossRef] [PubMed]
- Nigam, S.K.; Bhatnagar, V. The systems biology of uric acid transporters: The role of remote sensing and signaling. Curr. Opin. Nephrol. Hypertens. 2018, 27, 305–313. [Google Scholar] [CrossRef]
- Tan, A.; Fu, G.; Jin, L.; Guo, Q.; Li, Z.; Niu, B.; Meng, Z.; Morrison, N.I.; Alphey, L.; Huang, Y. Transgene-based, female-specific lethality system for genetic sexing of the silkworm, Bombyx mori. Proc. Natl. Acad. Sci. USA 2013, 110, 6766–6770. [Google Scholar] [CrossRef] [Green Version]
- Felsenstein, J. Confidence limits on phylogenies: An approach using the bootstrap. Evolution 1985, 39, 783–791. [Google Scholar] [CrossRef]
- Kumar, S.; Stecher, G.; Tamura, K. MEGA7: Molecular evolutionary genetics analysis version 7.0 for bigger datasets. Mol. Biol. Evol. 2016, 33, 1870–1874. [Google Scholar] [CrossRef] [Green Version]
- Xu, J.; Liu, W.; Yang, D.; Chen, S.; Chen, K.; Liu, Z.; Yang, X.; Meng, J.; Zhu, G.; Dong, S.; et al. Regulation of olfactory-based sex behaviors in the silkworm by genes in the sex-determination cascade. PLoS Genet. 2020, 16, e1008622. [Google Scholar] [CrossRef]
- Xu, J.; Chen, S.Q.; Zeng, B.S.; James, A.A.; Tan, A.J.; Huang, Y.P. Bombyx mori P-element somatic inhibitor (BmPSI) is a key auxiliary factor for silkworm male sex determination. PLoS Genet. 2017, 13, e1006576. [Google Scholar] [CrossRef]
- Zeng, B.S.; Huang, Y.P.; Xu, J.; Shiotsuki, T.; Bai, H.; Palli, S.R.; Huang, Y.P.; Tan, A.J. The FOXO transcription factor controls insect growth and development by regulating juvenile hormone degradation in the silkworm, Bombyx mori. J. Biol. Chem. 2017, 292, 11659–11669. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.; Li, Z.; Xu, J.; Zeng, B.; Ling, L.; You, L.; Chen, Y.; Huang, Y.; Tan, A. The CRISPR/Cas system mediates efficient genome engineering in Bombyx mori. Cell Res. 2013, 23. [Google Scholar] [CrossRef] [Green Version]
- Wang, X.; Gao, M.; Wang, Z.; Cui, W.; Zhang, J.; Zhang, W.; Xia, Y.; Wei, B.; Tang, Y.; Xu, X. Hepatoprotective effects of oridonin against bisphenol A induced liver injury in rats via inhibiting the activity of xanthione oxidase. Sci. Total Environ. 2021, 770, 145301. [Google Scholar] [CrossRef] [PubMed]
- Liu, R.X.; Hong, J.; Xu, X.Q.; Feng, Q.; Zhang, D.Y.; Gu, Y.Y.; Shi, J.; Zhao, S.Q.; Liu, W.; Wang, X.K.; et al. Gut microbiome and serum metabolome alterations in obesity and after weight-loss intervention. Nat. Med. 2017, 23, 859–868. [Google Scholar] [CrossRef]
- Zhong, H.Z.; Fang, C.; Fan, Y.Q.; Lu, Y.; Wen, B.; Ren, H.H.; Hou, G.X.; Yang, F.M.; Xie, H.L.; Jie, Z.Y.; et al. Lipidomic profiling reveals distinct differences in plasma lipid composition in healthy, prediabetic, and type 2 diabetic individuals. Gigascience 2017, 6, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Lee, K.R.; Lin, X.W.; Park, D.C.; Eslava, S. Megavariate data analysis of mass spectrometric proteomics data using latent variable projection method. Proteomics 2003, 3, 1680–1686. [Google Scholar] [CrossRef] [PubMed]
- Kanehisa, M.; Araki, M.; Goto, S.; Hattori, M.; Hirakawa, M.; Itoh, M.; Katayama, T.; Kawashima, S.; Okuda, S.; Tokimatsu, T.; et al. KEGG for linking genomes to life and the environment. Nucleic Acids Res. 2008, 36, D480–D484. [Google Scholar] [CrossRef]
- Wishart, D.S.; Feunang, Y.D.; Marcu, A.; Guo, A.C.; Liang, K.; Vazquez-Fresno, R.; Sajed, T.; Johnson, D.; Li, C.R.; Karu, N.; et al. HMDB 4.0: The human metabolome database for 2018. Nucleic Acids Res. 2018, 46, D608–D617. [Google Scholar] [CrossRef]
- Xin, B.; Tao, F.; Wang, Y.; Liu, H.Y.; Ma, C.Q.; Xu, P. Coordination of metabolic pathways: Enhanced carbon conservation in 1,3-propanediol production by coupling with optically pure lactate biosynthesis. Metab. Eng. 2017, 41, 102–114. [Google Scholar] [CrossRef]
- Conesa, A.; Gotz, S.; Garcia-Gomez, J.M.; Terol, J.; Talon, M.; Robles, M. Blast2GO: A universal tool for annotation, visualization and analysis in functional genomics research. Bioinformatics 2005, 21, 3674–3676. [Google Scholar] [CrossRef] [Green Version]
- Gonzalez, I.; Le Cao, K.A.; Davis, M.J.; Dejean, S. Visualising associations between paired ‘omics’ data sets. Biodata Min. 2012, 5. [Google Scholar] [CrossRef] [Green Version]
- Rohart, F.; Gautier, B.; Singh, A.; Le Cao, K.A. mixOmics: An R package for ‘omics feature selection and multiple data integration. PLoS Comput. Biol. 2017, 13, e1005752. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lima, W.G.; Martins-Santos, M.E.S.; Chaves, V.E. Uric acid as a modulator of glucose and lipid metabolism. Biochimie 2015, 116, 17–23. [Google Scholar] [CrossRef]
- Xu, J.; Chen, R.M.; Chen, S.Q.; Chen, K.; Tang, L.M.; Yang, D.H.; Yang, X.; Zhang, Y.; Song, H.S.; Huang, Y.P. Identification of a germline-expression promoter for genome editing in Bombyx mori. Insect Sci. 2019, 26, 991–999. [Google Scholar] [CrossRef]
- Wang, L.Y.; Kiuchi, T.; Fujii, T.; Daimon, T.; Li, M.W.; Banno, Y.; Kikuta, S.; Kikawada, T.; Katsuma, S.; Shimada, T. Mutation of a novel ABC transporter gene is responsible for the failure to incorporate uric acid in the epidermis of ok mutants of the silkworm, Bombyx mori. Insect Biochem. Mol. Biol. 2013, 43, 562–571. [Google Scholar] [CrossRef]
- Fujii, T.; Kakino, K.; Tanaka, M.; Lee, J.M.; Kusakabe, T.; Banno, Y. A defect in purine nucleotide metabolism in the silkworm, Bombyx mori, causes a translucent larval integument and male infertility. Insect Biochem. Mol. Biol. 2020, 126, 103458. [Google Scholar] [CrossRef]
- Mandal, A.K.; Mount, D.B. The molecular physiology of uric acid homeostasis. Annu. Rev. Physiol. 2015, 77, 323–345. [Google Scholar] [CrossRef]
- Odonnell, M.J.; Maddrell, S.H.P.; Gardiner, B.O.C. Transport of Uric-Acid by the Malpighian Tubules of Rhodnius-Prolixus and Other Insects. J. Exp. Biol. 1983, 103, 169–184. [Google Scholar] [CrossRef]
- Balasubramaniam, S.; Duley, J.A.; Christodoulou, J. Inborn errors of purine metabolism: Clinical update and therapies. J. Inherit. Metab. Dis. 2014, 37, 669–686. [Google Scholar] [CrossRef]
- Komoto, N.; Quan, G.X.; Sezutsu, H.; Tamura, T. A single-base deletion in an ABC transporter gene causes white eyes, white eggs, and translucent larval skin in the silkworm w-3(oe) mutant. Insect Biochem. Mol. Biol. 2009, 39, 152–156. [Google Scholar] [CrossRef] [PubMed]
- Maiuolo, J.; Oppedisano, F.; Gratteri, S.; Muscoli, C.; Mollace, V. Regulation of uric acid metabolism and excretion. Int. J. Cardiol. 2016, 213, 8–14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fujii, T.; Yamamoto, K.; Banno, Y. Molybdenum cofactor deficiency causes translucent integument, male-biased lethality, and flaccid paralysis in the silkworm Bombyx mori. Insect Biochem. Mol. Biol. 2016, 73, 20–26. [Google Scholar] [CrossRef] [PubMed]
- Tabunoki, H.; Ono, H.; Ode, H.; Ishikawa, K.; Kawana, N.; Banno, Y.; Shimada, T.; Nakamura, Y.; Yamamoto, K.; Satoh, J.; et al. Identification of key uric acid synthesis pathway in a unique mutant silkworm Bombyx mori model of Parkinson’s disease. PLoS ONE 2013, 8, e69130. [Google Scholar] [CrossRef] [PubMed]
- Horsley, A.; Helm, J.; Brennan, A.; Bright-Thomas, R.; Webb, K.; Jones, A. Gout and hyperuricaemia in adults with cystic fibrosis. J. R. Soc. Med. 2011, 104 (Suppl. 1), S36–S39. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vidanapathirana, D.M.; Jayasena, S.; Jasinge, E.; Stiburkova, B. A heterozygous variant in the SLC22A12 gene in a Sri Lanka family associated with mild renal hypouricemia. BMC Pediatr. 2018, 18, 210. [Google Scholar] [CrossRef] [PubMed]
- Xu, J.; Zhan, S.; Chen, S.; Zeng, B.; Li, Z.; James, A.A.; Tan, A.; Huang, Y. Sexually dimorphic traits in the silkworm, Bombyx mori, are regulated by doublesex. Insect Biochem. Mol. Biol. 2017, 80, 42–51. [Google Scholar] [CrossRef] [Green Version]
- Meng, X.; Zhu, F.F.; Chen, K.P. Silkworm: A promising model organism in life science. J. Insect. Sci. 2017, 17, 95. [Google Scholar]
- Tabunoki, H.; Bono, H.; Ito, K.; Yokoyama, T. Can the silkworm (Bombyx mori) be used as a human disease model? Drug Discov. Ther. 2016, 10, 3–8. [Google Scholar] [CrossRef] [Green Version]
- Zhang, X.; Xue, R.; Cao, G.; Pan, Z.; Zheng, X.; Gong, C. Silkworms can be used as an animal model to screen and evaluate gouty therapeutic drugs. Insect Sci. 2012, 12, 4. [Google Scholar] [CrossRef] [PubMed]







Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tang, L.; Yang, D.; Wang, Y.; Yang, X.; Chen, K.; Luo, X.; Xu, J.; Liu, Y.; Tang, Z.; Zhang, Q.; et al. 5′-Nucleotidase Plays a Key Role in Uric Acid Metabolism of Bombyx mori. Cells 2021, 10, 2243. https://doi.org/10.3390/cells10092243
Tang L, Yang D, Wang Y, Yang X, Chen K, Luo X, Xu J, Liu Y, Tang Z, Zhang Q, et al. 5′-Nucleotidase Plays a Key Role in Uric Acid Metabolism of Bombyx mori. Cells. 2021; 10(9):2243. https://doi.org/10.3390/cells10092243
Chicago/Turabian StyleTang, Linmeng, Dehong Yang, Yaohui Wang, Xu Yang, Kai Chen, Xingyu Luo, Jun Xu, Yujia Liu, Zheng Tang, Qianqian Zhang, and et al. 2021. "5′-Nucleotidase Plays a Key Role in Uric Acid Metabolism of Bombyx mori" Cells 10, no. 9: 2243. https://doi.org/10.3390/cells10092243