
Degenerated, Undifferentiated, Rearranged, Lost: High Variability of Sex Chromosomes in Geometridae (Lepidoptera) Identified by Sex Chromatin

 , , , and
, , , and
Abstract
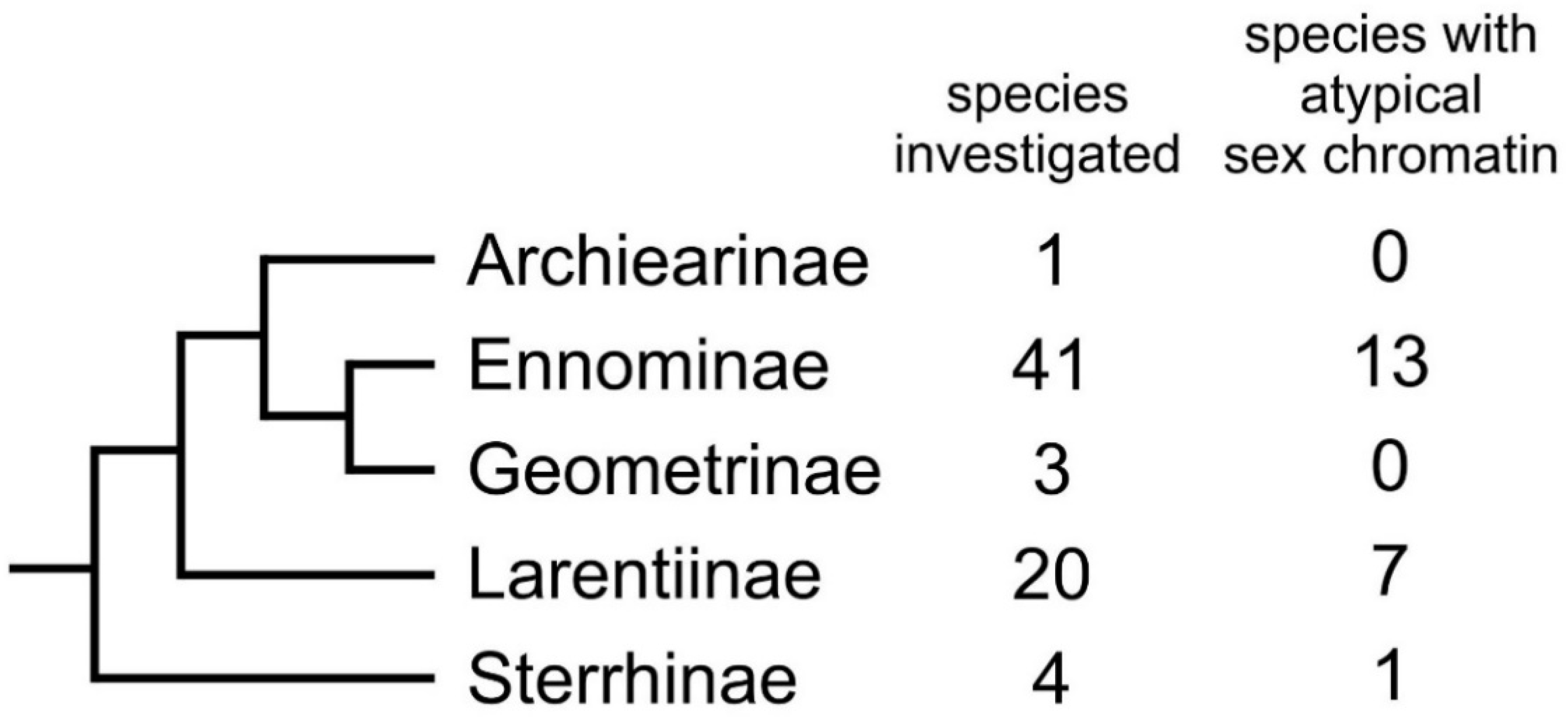
:1. Introduction
2. Materials and Methods
2.1. Specimens
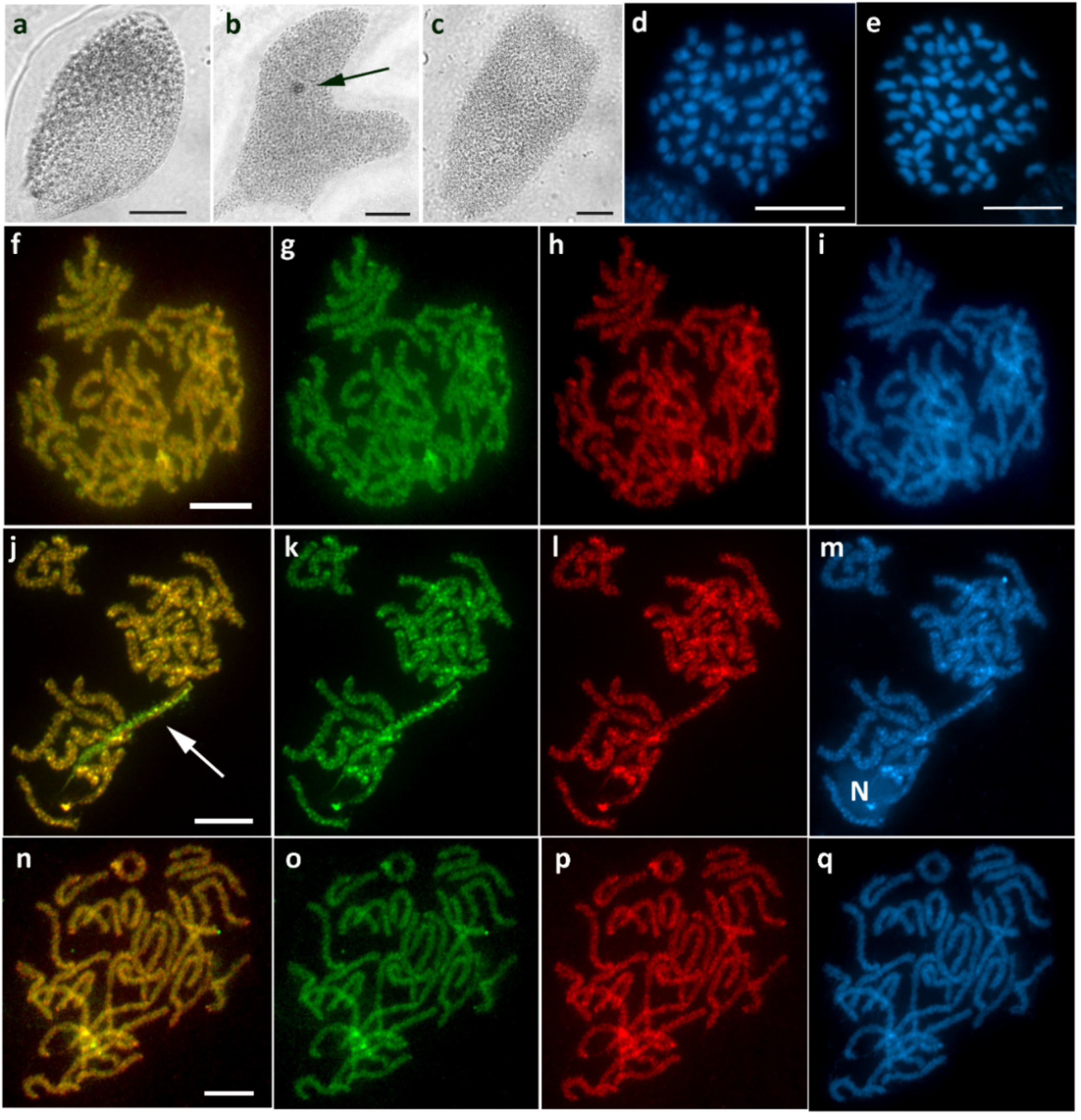
2.2. Chromosomal Preparations and Sex Chromatin Assay
2.3. DNA Isolation
2.4. Comparative Genomic Hybridization (CGH)
2.5. Genomic In Situ Hybridization (GISH) with 18S rDNA Probe
2.6. Reprobing and Fluorescence In Situ Hybridization (FISH) with Telomeric Probe
2.7. FISH with W-Painting Probe
2.8. Microscopy and Image Processing
3. Results
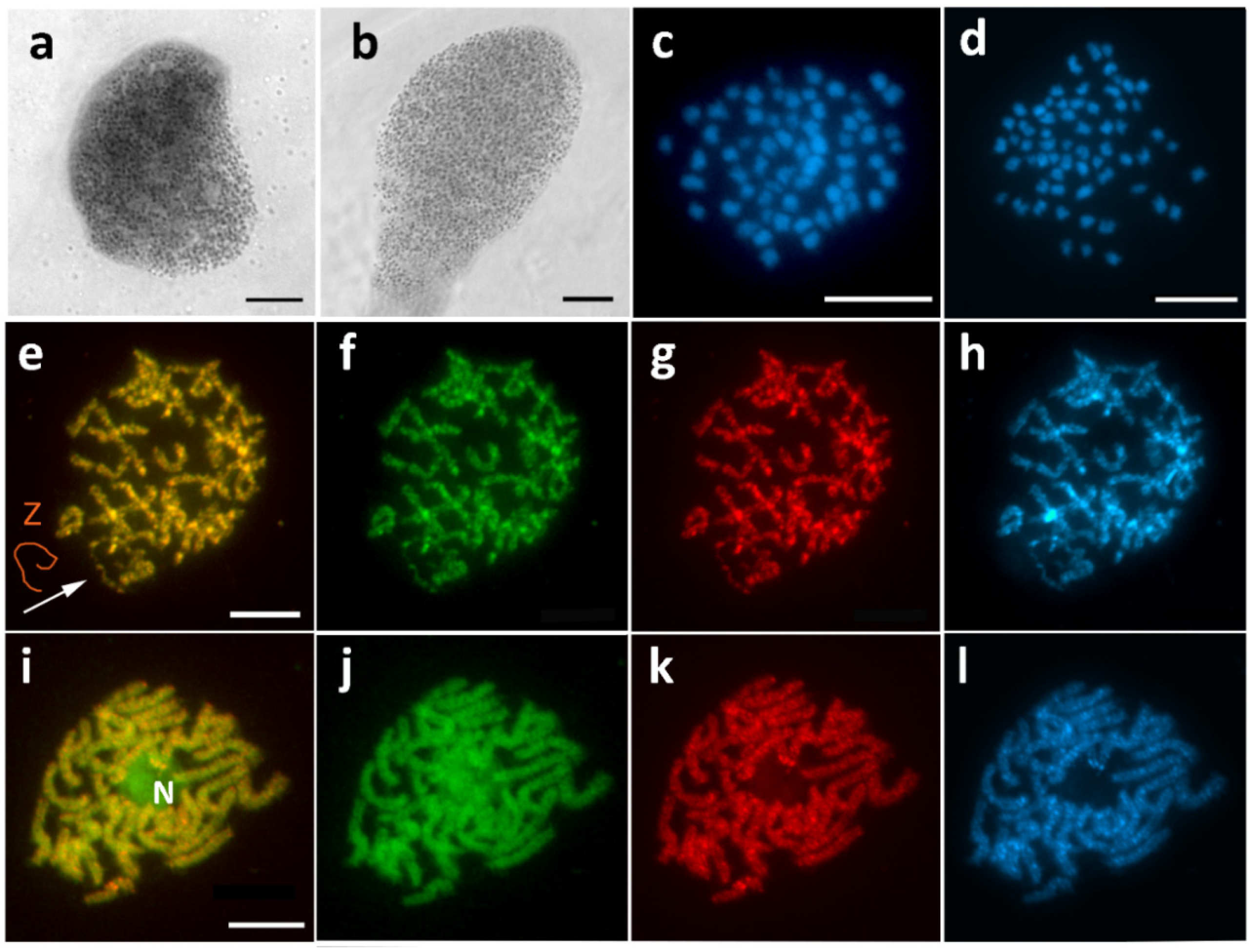
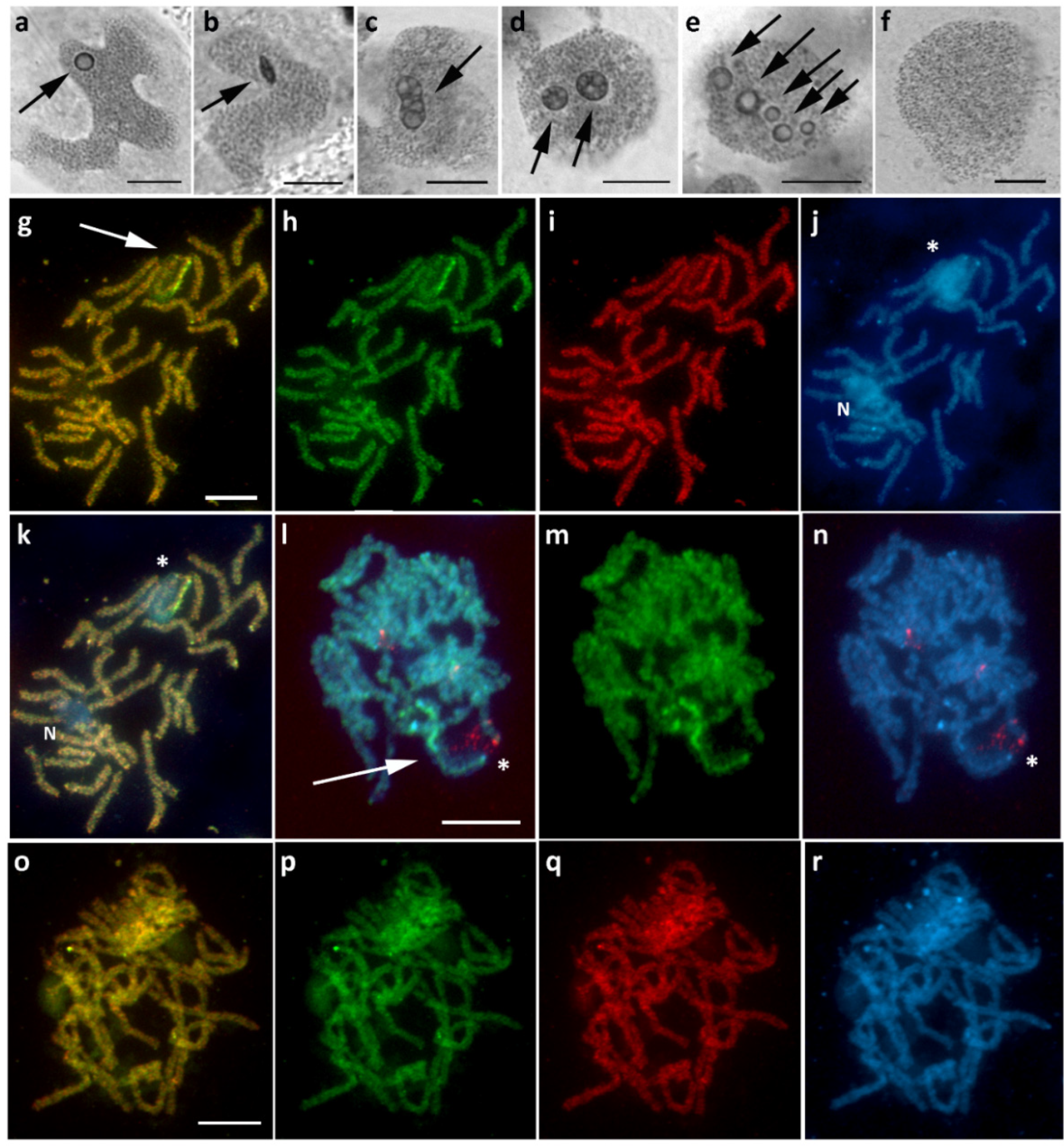
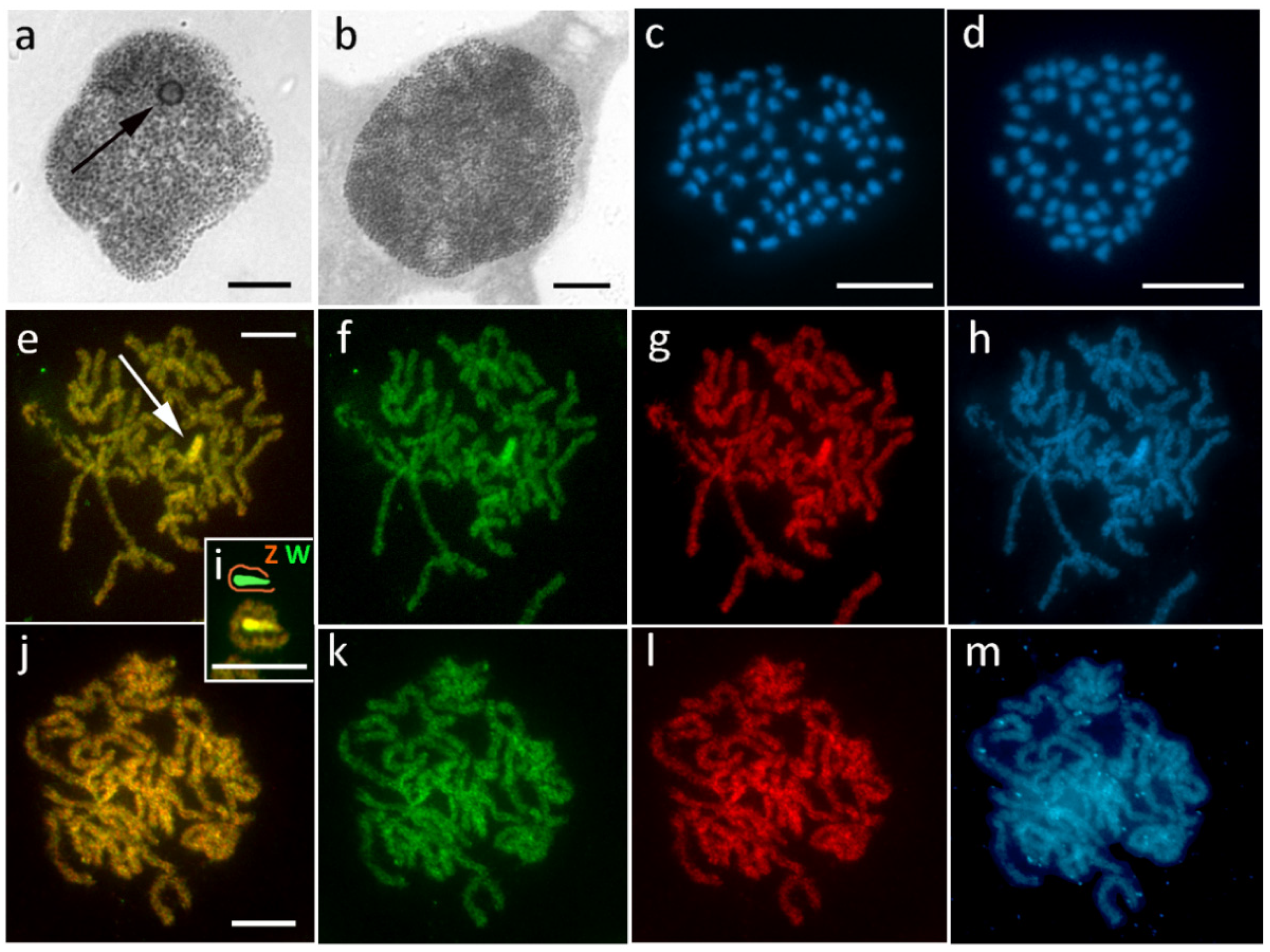
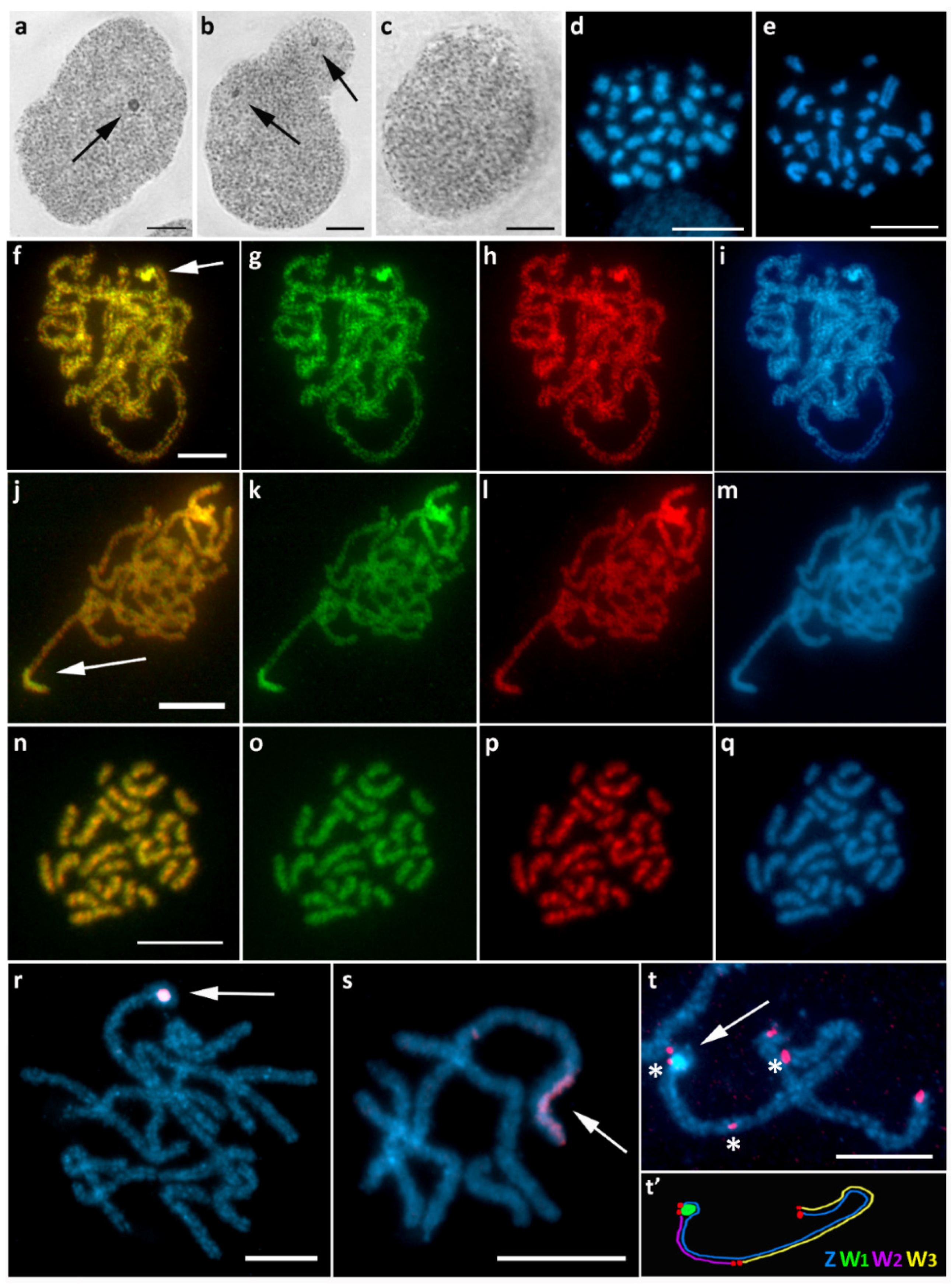
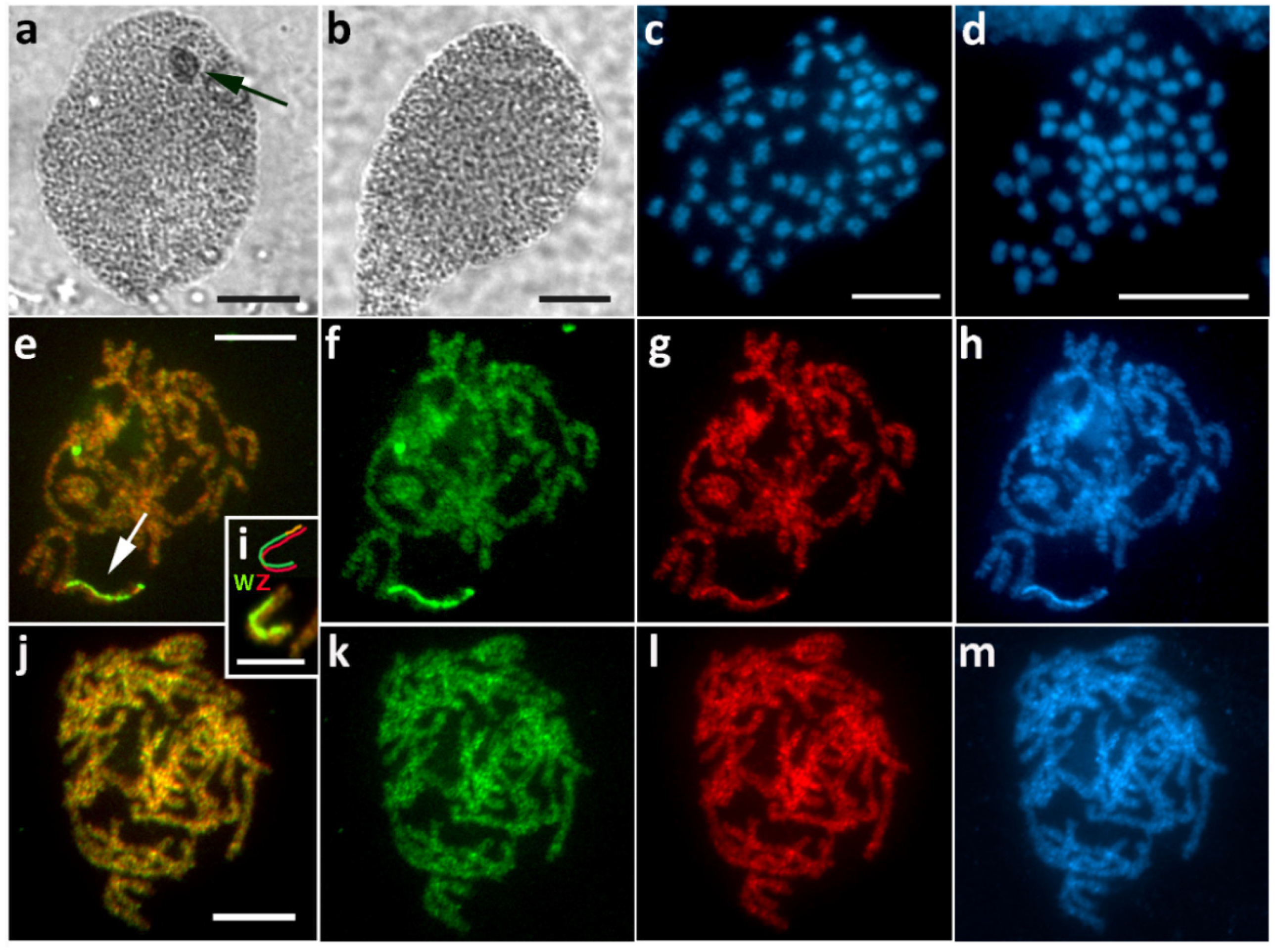
3.1. Aethalura Punctulata (Ennominae)
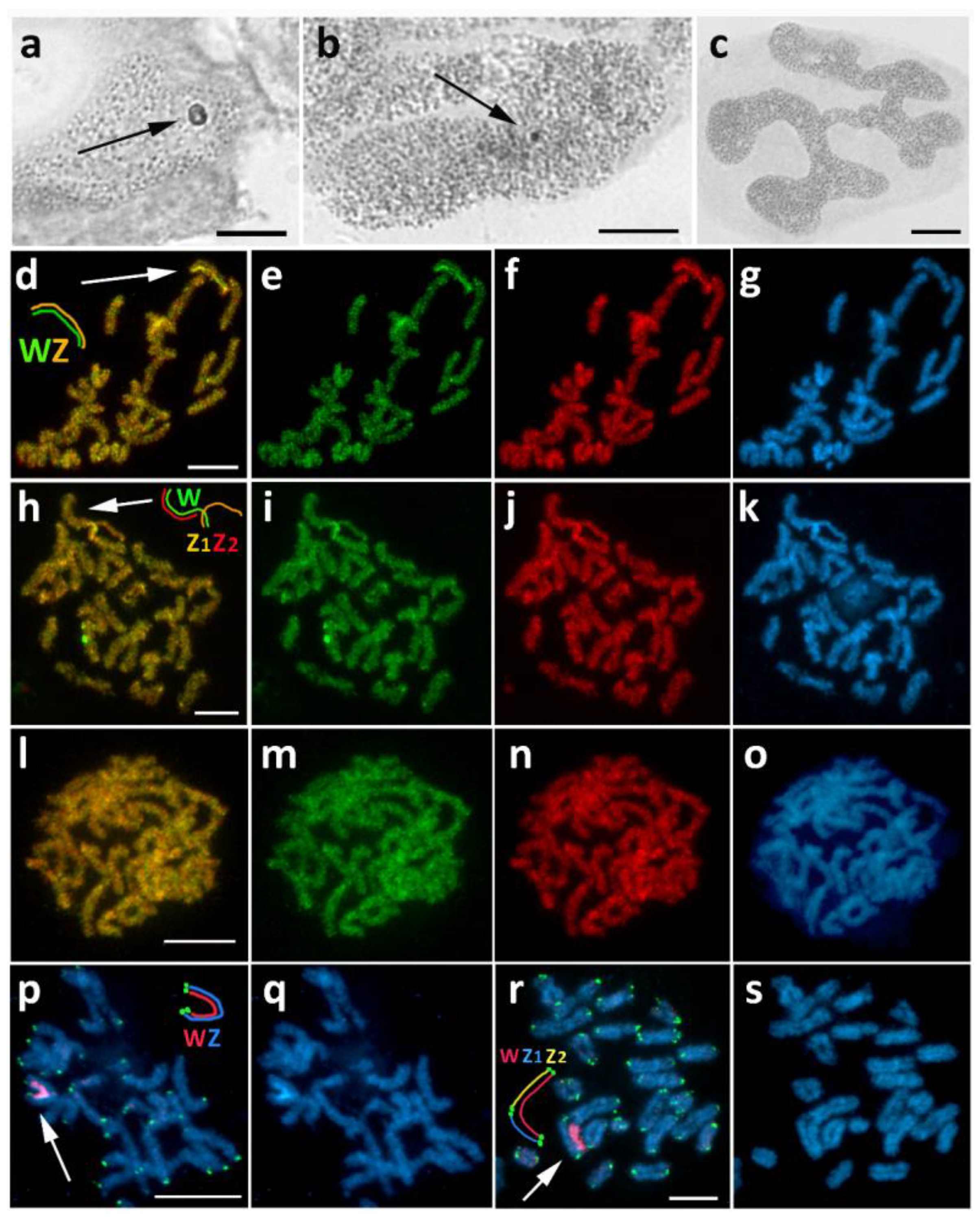
3.2. Chiasmia Clathrata (Ennominae)
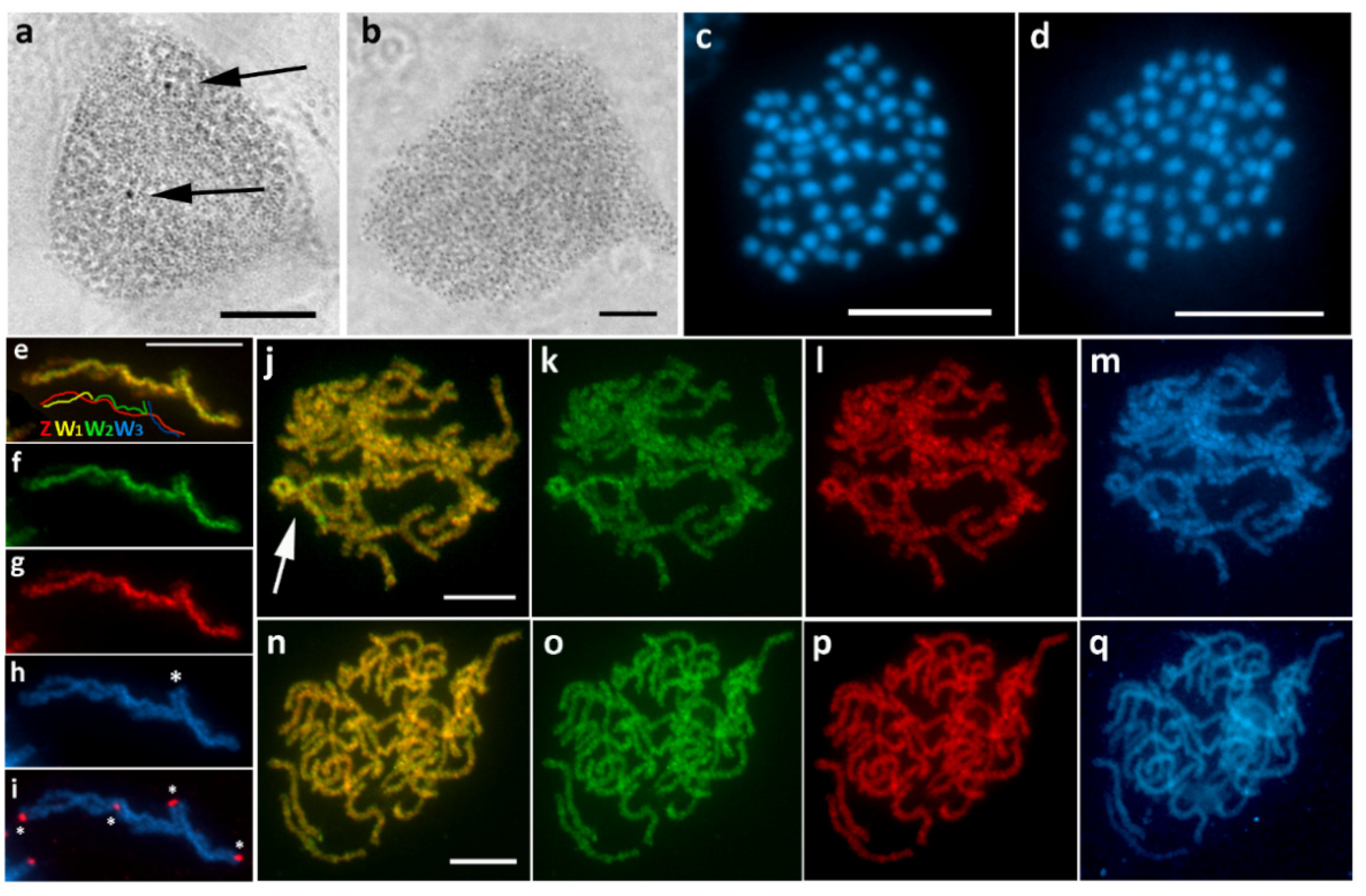
3.3. Epirrhoe Alternata (Larentiinae)
3.4. Hylaea Fasciaria (Ennominae)
3.5. Hypomecis Atomaria (Ennominae)
3.6. Operophtera Brumata (Larentiinae)
3.7. Peribatodes Rhomboidaria (Ennominae)
3.8. Pseudopanthera Macularia (Ennominae)

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | 2n ♀/♂ | SC ♀ | W Composition + Features | Sex Chromatin |
|---|---|---|---|---|
| A. punctulata | 61/62 | Z0 | absent (Z univalent recorded) | absent |
| C. clathrata | n.d. | WZ | female enriched/common repetitive (slightly DAPI+) | normal |
| n.d. | WZ1Z2 | neo-W with 2 parts: female enriched/common repetitive (slightly DAPI+) and undifferentiated | scattered | |
| E. alternata | 64/62 | W1W2W3Z | common repeats/female enriched, DAPI+ blocks on W2, W3 | scattered |
| H. fasciaria | n.d. | WZ | 2 parts: female enriched (DAPI+) and undifferentiated (nucleolus-associated) | normal/multiple |
| H. atomaria | 62/62 | WZ | common repeats, DAPI+, small size | normal |
| O. brumata | 30/28 | W1W2W3Z | W1 female enriched/common repeats, DAPI+, W2 and W3 undifferentiated; neo-Z | normal/double |
| P. rhomboidaria | 62/62 | WZ | female enriched, DAPI+ | normal |
| P. macularia | 62/62 | WZ | female enriched | normal |
| undifferentiated | absent |
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Acknowledgments
Conflicts of Interest
References
- Charlesworth, D.; Charlesworth, B.; Marais, G. Steps in the evolution of heteromorphic sex chromosomes. J. Hered. 2005, 95, 118–128. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abbott, J.K.; Nordén, A.K.; Hansson, B. Sex chromosome evolution: Historical insights and future perspectives. Proc. Biol. Sci. 2017, 284, 20162806. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Charlesworth, B. The evolution of chromosomal sex determination and dosage compensation. Curr. Biol. 1996, 6, 149–162. [Google Scholar] [CrossRef] [Green Version]
- Furman, B.L.S.; Metzger, D.C.H.; Darolti, I.; Wright, A.E.; Sandkam, B.A.; Almeida, P.; Shu, J.J.; Mank, J.E. Sex chromosome evolution: So many exceptions to the rules. Genome Biol. Evol. 2020, 12, 750–763. [Google Scholar] [CrossRef]
- Nokkala, S.; Grozeva, S.; Kuznetsova, V.; Maryanska-Nadachowska, A. The origin of the achiasmatic XY sex chromosome system in Cacopsylla peregrina (Frst.) (Psylloidea, Homoptera). Genetica 2003, 119, 327–332. [Google Scholar] [CrossRef] [PubMed]
- Clark, F.E.; Kocher, T.D. Changing sex for selfish gain: B chromosomes of Lake Malawi cichlid fish. Sci. Rep. 2019, 9, 20213. [Google Scholar] [CrossRef]
- Traut, W.; Marec, F. Sex chromosome differentiation in some species of Lepidoptera (Insecta). Chromosome Res. 1997, 5, 283–291. [Google Scholar] [CrossRef]
- Yoshido, A.; Marec, F.; Sahara, K. Resolution of sex chromosome constitution by genomic in situ hybridization and fluorescence in situ hybridization with (TTAGG)n telomeric probe in some species of Lepidoptera. Chromosoma 2005, 114, 193–202. [Google Scholar] [CrossRef] [Green Version]
- Vlašánek, P.; Bartoňová, A.; Marec, F.; Konvička, M. Elusive Parnassius mnemosyne (Linnaeus, 1758) larvae: Habitat selection, sex determination and sex ratio (Lepidoptera: Papilionidae). SHILAP Rev. Lepidopt. 2017, 45, 561–569. [Google Scholar]
- Grützner, F.; Deakin, J.; Rens, W.; El-Mogharbel, N.; Marshall Graves, J. The monotreme genome: A patchwork of reptile, mammal and unique features? Comp. Biochem. Physiol. A Mol. Integr. Physiol. 2003, 136, 867–881. [Google Scholar] [CrossRef]
- Yoshido, A.; Šíchová, J.; Pospíšilová, K.; Nguyen, P.; Šafář, J.; Provazník, J.; Voleníková, A.; Vila, R.; Marec, F. Evolution of multiple sex-chromosomes associated with dynamic genome reshuffling in Leptidea wood-white butterflies. Heredity 2020, 125, 138–154. [Google Scholar] [CrossRef] [PubMed]
- Šíchová, J.; Ohno, M.; Dincă, V.; Watanabe, M.; Sahara, K.; Marec, F. Fissions, fusions, and translocations shaped the karyotype and multiple sex chromosome constitution of the northeast-Asian wood white butterfly, Leptidea amurensis. Biol. J. Linn. Soc. 2016, 118, 457–471. [Google Scholar] [CrossRef] [Green Version]
- Zrzavá, M.; Hladová, I.; Dalíková, M.; Šíchová, J.; Õunap, E.; Kubíčková, S.; Marec, F. Sex chromosomes of the iconic moth Abraxas grossulariata (Lepidoptera, Geometridae) and its congener A. sylvata. Genes 2018, 9, 279. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chirino, M.G.; Fourastie, M.F.; Cemteno, N.D.; Bressa, M.J. Unusual chromosome polymorphism and heterochromatin variation in the Argentinean population of the necrophagous fly Lucilia sericata (Diptera: Calliphoridae), comparison with other populations and evolutionary aspects. Eur. J. Entomol. 2020, 117, 295–301. [Google Scholar] [CrossRef]
- Sahara, K.; Yoshido, A.; Traut, W. Sex chromosome evolution in moths and butterflies. Chromosome Res. 2012, 20, 83–94. [Google Scholar] [CrossRef] [Green Version]
- Wright, A.E.; Dean, R.; Zimmer, F.; Mank, J.E. How to make a sex chromosome. Nat. Commun. 2016, 7, 12087. [Google Scholar] [CrossRef]
- Dalíková, M.; Zrzavá, M.; Hladová, I.; Nguyen, P.; Šonský, I.; Flegrová, M.; Kubíčková, S.; Voleníková, A.; Kawahara, A.Y.; Peters, R.S.; et al. New insights into the evolution of the W chromosome in Lepidoptera. J. Hered. 2017, 108, 709–719. [Google Scholar] [CrossRef]
- Marec, F.; Traut, W. Sex chromosome pairing and sex chromatin bodies in W-Z translocation strains of Ephestia kuehniella (Lepidoptera). Genome 1994, 37, 426–435. [Google Scholar] [CrossRef]
- Beldade, P.; Saenko, S.V.; Pul, N.; Long, A.D. A gene-based linkage map for Bicyclus anynana butterflies allows for a comprehensive analysis of synteny with the lepidopteran reference genome. PLoS Genet. 2009, 5, e1000366. [Google Scholar] [CrossRef] [Green Version]
- Van’t Hof, A.E.; Nguyen, P.; Dalíková, M.; Edmonds, N.; Marec, F.; Saccheri, I.J. Linkage map of the peppered moth, Biston betularia (Lepidoptera, Geometridae): A model of industrial melanism. Heredity 2013, 110, 283–295. [Google Scholar] [CrossRef] [Green Version]
- Fraïsse, C.; Picard, M.A.L.; Vicoso, B. The deep conservation of the Lepidoptera Z chromosome suggests a non-canonical origin of the W. Nat. Commun. 2017, 8, 1486. [Google Scholar] [CrossRef] [Green Version]
- Traut, W.; Sahara, K.; Marec, F. Sex chromosomes and sex determination in Lepidoptera. Sex Dev. 2007, 1, 332–346. [Google Scholar] [CrossRef] [PubMed]
- Traut, W.; Vogel, H.; Glöckner, G.; Hartmann, E.; Heckel, D.G. High-throughput sequencing of a single chromosome: A moth W chromosome. Chromosome Res. 2013, 21, 491–505. [Google Scholar] [CrossRef] [PubMed]
- Traut, W.; Marec, F. Sex chromatin in Lepidoptera. Q. Rev. Biol. 1996, 71, 239–256. [Google Scholar] [CrossRef] [PubMed]
- Lukhtanov, V.A. Sex chromatin and sex chromosome systems in nonditrysian Lepidoptera (Insecta). J. Zool. Syst. Evol. Res. 2000, 38, 73–79. [Google Scholar] [CrossRef] [Green Version]
- Šíchová, J.; Nguyen, P.; Dalíková, M.; Marec, F. Chromosomal evolution in tortricid moths: Conserved karyotypes with diverged features. PLoS ONE 2013, 8, e64520. [Google Scholar] [CrossRef] [Green Version]
- Hejníčková, M.; Koutecký, P.; Potocký, P.; Provazníková, I.; Voleníková, A.; Dalíková, M.; Visser, S.; Marec, F.; Zrzavá, M. Absence of W chromosome in Psychidae moths and implications for the theory of sex chromosome evolution in Lepidoptera. Genes 2019, 10, 1016. [Google Scholar] [CrossRef] [Green Version]
- Rathjens, B. Zur Funktion des W-Chromatins bei Ephestia kuehniella (Lepidoptera). Isolierung und Charakterisierung von W-Chromatin-Mutanten. Chromosoma 1974, 47, 21–44. [Google Scholar] [CrossRef]
- Traut, W.; Weith, A.; Traut, G. Structural mutants of the W chromosome in Ephestia (Insecta, Lepidoptera). Genetica 1986, 70, 69–79. [Google Scholar] [CrossRef]
- Makee, H.; Tafesh, N. Sex chromatin body as a cytogenetic marker of W chromosome aberrations in Cydia pomonella females. In Area-Wide Control of Insect Pests; Springer: Dordrecht, The Netherlands, 2007; pp. 113–118. [Google Scholar] [CrossRef]
- Traut, W.; Clarke, C.A. Karyotype evolution by chromosome fusion in the moth genus Orgyia. Hereditas 1997, 126, 77–84. [Google Scholar] [CrossRef]
- Fuková, I.; Traut, W.; Vítková, M.; Nguyen, P.; Kubíčková, S.; Marec, F. Probing the W chromosome of the codling moth, Cydia pomonella, with sequences from microdissected sex chromatin. Chromosoma 2007, 116, 135–145. [Google Scholar] [CrossRef] [PubMed]
- Vítková, M.; Fuková, I.; Kubíčková, S.; Marec, F. Molecular divergence of the W chromosomes in pyralid moths (Lepidoptera). Chromosome Res. 2007, 15, 917–930. [Google Scholar] [CrossRef] [PubMed]
- Traut, W.; Sahara, K.; Otto, T.D.; Marec, F. Molecular differentiation of sex chromosomes probed by comparative genomic hybridization. Chromosoma 1999, 108, 173–180. [Google Scholar] [CrossRef]
- Sahara, K.; Marec, F.; Eickhoff, U.; Traut, W. Moth sex chromatin probed by comparative genomic hybridization (CGH). Genome 2003, 46, 339–342. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Green, J.E.; Dalíková, M.; Sahara, K.; Akam, M.; Marec, F. XX/XY system of sex determination in the geophilomorph centipede Strigamia maritima. PLoS ONE 2016, 11, e0150292. [Google Scholar] [CrossRef] [Green Version]
- Sihvonen, P.; Mutanen, M.; Kaila, L.; Brehm, G.; Hausmann, A.; Staude, H.S. Comprehensive molecular sampling yields a robust phylogeny for geometrid moths (Lepidoptera: Geometridae). PLoS ONE 2011, 6, e20356. [Google Scholar] [CrossRef] [Green Version]
- Traut, W.; Mosbacher, G.C. Geschlechtschromatin bei Lepidopteren. Chromosoma. 1968, 25, 343–356. [Google Scholar] [CrossRef]
- Ennis, T.J. Sex chromatin and chromosome numbers in Lepidoptera. Can. J. Genet. Cytol. 1976, 18, 119–130. [Google Scholar] [CrossRef]
- Makino, S. An Atlas of the Chromosome Numbers in Animals, 2nd ed.; Iowa State College Press: Ames, IA, USA, 1951. [Google Scholar] [CrossRef] [Green Version]
- Robinson, R. Lepidoptera Genetics; Pergamon Press: Oxford, UK, 1970. [Google Scholar]
- Murillo-Ramos, L.; Brehm, G.; Sihvonen, P.; Hausmann, A.; Holm, S.; Reza Ghanavi, H.; Õunap, E.; Truuverk, A.; Staude, H.; Friedrich, E.; et al. A comprehensive molecular phylogeny of Geometridae (Lepidoptera) with a focus on enigmatic small subfamilies. Peer J. 2019, 27, e7386. [Google Scholar] [CrossRef] [Green Version]
- Mediouni, J.; Fuková, I.; Frydrychová, R.; Dhouibi, M.H.; Marec, F. Karyotype, sex chromatin and sex chromosome differentiation in the carob moth, Ectomyelois ceratoniae (Lepidoptera: Pyralidae). Caryologia 2004, 57, 184–194. [Google Scholar] [CrossRef]
- Winnepenninckx, B.; Backeljau, T.; de Wachter, R. Extraction of high molecular weight DNA from molluscs. Trends Genet. 1993, 12, 407. [Google Scholar] [CrossRef]
- Kato, A.; Albert, P.S.; Vega, J.M.; Birchler, J.A. Sensitive fluorescence in situ hybridization signal detection in maize using directly labeled probes produced by high concentration DNA polymerase nick translation. Biotech. Histochem. 2006, 81, 71–78. [Google Scholar] [CrossRef] [PubMed]
- Fuková, I.; Nguyen, P.; Marec, F. Codling moth cytogenetics: Karyotype, chromosomal location of rDNA, and molecular differentiation of sex chromosomes. Genome 2005, 48, 1083–1092. [Google Scholar] [CrossRef] [PubMed]
- Okazaki, S.; Tsuchida, K.; Maekawa, H.; Ishikawa, H.; Fujiwara, H. Identification of a pentanucleotide telomeric sequence, (TTAGG)n, in the silkworm Bombyx mori and in other insects. Mol. Cell. Biol. 1993, 13, 1424–1432. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ijdo, J.W.; Wells, R.A.; Baldini, A.; Reeders, S.T. Improved telomere detection using a telomere repeat probe (TTAGGG)n generated by PCR. Nucleic Acids Res. 1991, 19, 4780. [Google Scholar] [CrossRef] [Green Version]
- Sahara, K.; Marec, F.; Traut, W. TTAGG telomeric repeats in chromosomes of some insects and other arthropods. Chromosome Res. 1999, 7, 449–460. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shibata, F.; Sahara, K.; Naito, Y.; Yasukochi, Y. Reprobing multicolor FISH preparations in lepidopteran chromosome. Zool. Sci. 2009, 26, 187–193. [Google Scholar] [CrossRef]
- Murillo-Ramos, L.; Chazot, N.; Sihvonen, P.; Õunap, E.; Jiang, N.; Han, H.; Clarke, J.T.; Davis, R.B.; Tammaru, T.; Wahlberg, N. Molecular phylogeny, classification, biogeography and diversification patterns of a diverse group of moths (Geometridae: Boarmiini). Mol. Phylogenet. Evol. 2021, 162, 107198. [Google Scholar] [CrossRef] [PubMed]
- Van’t Hof, A.E.; Marec, F.; Saccheri, I.J.; Brakefield, P.M.; Zwaan, B.J. Cytogenetic characterization and AFLP-based genetic linkage mapping for the butterfly Bicyclus anynana, covering all 28 karyotyped chromosomes. PLoS ONE 2008, 3, e3882. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Smith, D.A.S.; Gordon, I.J.; Traut, W.; Herren, J.; Collins, S.; Martins, D.J.; Saitoti, K.; Ireri, P.; Ffrench-Constant, R. A neo-W chromosome in a tropical butterfly links colour pattern, male-killing, and speciation. Proc. R. Soc. B 2016, 283, 20160821. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Loda, A.; Brandsma, J.H.; Vassilev, I.; Servant, N.; Loos, F.; Amirnasr, A.; Splinter, E.; Barillot, E.; Poot, R.A.; Heard, E.; et al. Genetic and epigenetic features direct differential efficiency of Xist-mediated silencing at X-chromosomal and autosomal locations. Nat. Commun. 2017, 8, 690. [Google Scholar] [CrossRef] [Green Version]
- Kataoka, K.; Noto, T.; Mochizuki, K. Phosphorylation of an HP1-like protein regulates heterochromatin body assembly for DNA elimination. Dev. Cell. 2015, 35, 775–788. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mongue, A.J.; Nguyen, P.; Voleníková, A.; Walters, J.R. Neo-sex chromosomes in the Monarch butterfly, Danaus plexippus. G3 Genes Genomes Genet. 2017, 7, 3281–3294. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yoshido, A.; Marec, F.; Sahara, K. The fate of W chromosomes in hybrids between wild silkmoths, Samia cynthia ssp.: No role in sex determination and reproduction. Heredity 2016, 116, 424–433. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kiuchi, T.; Koga, H.; Kawamoto, M.; Shoji, K.; Sakai, H.; Arai, Y.; Ishihara, G.; Kawaoka, S.; Sugano, S.; Shimada, T.; et al. A single female-specific piRNA is the primary determiner of sex in the silkworm. Nature 2014, 509, 633–636. [Google Scholar] [CrossRef]
- Yoshido, A.; Sahara, K.; Marec, F.; Matsuda, Y. Step-by-step evolution of neo-sex chromosomes in geographical populations of wild silkmoths, Samia cynthia ssp. Heredity 2011, 106, 614–624. [Google Scholar] [CrossRef] [Green Version]
- Carabajal Paladino, L.Z.; Provazníková, I.; Berger, M.; Bass, C.; Aratchige, N.S.; López, S.N.; Marec, F.; Nguyen, P. Sex chromosome turnover in moths of the diverse superfamily Gelechioidea. Genome Biol. Evol. 2019, 11, 1307–1319. [Google Scholar] [CrossRef]
- Šíchová, J.; Voleníková, A.; Dincă, V.; Nguyen, P.; Vila, R.; Sahara, K.; Marec, F. Dynamic karyotype evolution and unique sex determination systems in Leptidea wood white butterflies. BMC Evol. Biol. 2015, 15, 89. [Google Scholar] [CrossRef] [Green Version]









Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hejníčková, M.; Dalíková, M.; Potocký, P.; Tammaru, T.; Trehubenko, M.; Kubíčková, S.; Marec, F.; Zrzavá, M. Degenerated, Undifferentiated, Rearranged, Lost: High Variability of Sex Chromosomes in Geometridae (Lepidoptera) Identified by Sex Chromatin. Cells 2021, 10, 2230. https://doi.org/10.3390/cells10092230
Hejníčková M, Dalíková M, Potocký P, Tammaru T, Trehubenko M, Kubíčková S, Marec F, Zrzavá M. Degenerated, Undifferentiated, Rearranged, Lost: High Variability of Sex Chromosomes in Geometridae (Lepidoptera) Identified by Sex Chromatin. Cells. 2021; 10(9):2230. https://doi.org/10.3390/cells10092230
Chicago/Turabian StyleHejníčková, Martina, Martina Dalíková, Pavel Potocký, Toomas Tammaru, Marharyta Trehubenko, Svatava Kubíčková, František Marec, and Magda Zrzavá. 2021. "Degenerated, Undifferentiated, Rearranged, Lost: High Variability of Sex Chromosomes in Geometridae (Lepidoptera) Identified by Sex Chromatin" Cells 10, no. 9: 2230. https://doi.org/10.3390/cells10092230