
Mesenchymal Stem Cells, Bioactive Factors, and Scaffolds in Bone Repair: From Research Perspectives to Clinical Practice


Abstract
:1. Introduction
2. The Role of MSCs in Bone Formation and Healing
2.1. Bone Ossification
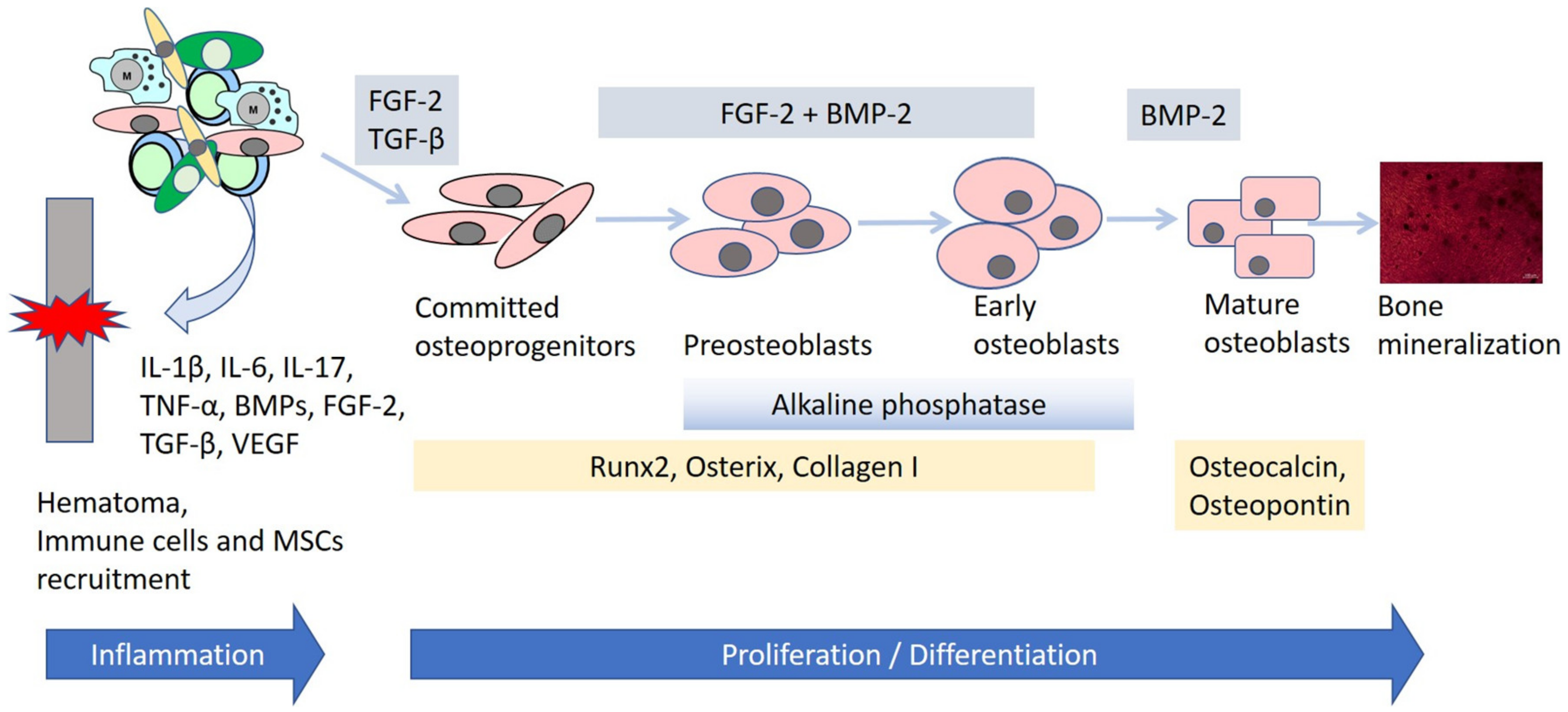
2.2. Bone Repair
3. MSC-Based Tissue Engineering Therapies in Bone Repair from a Research Perspective
3.1. Sources and Biological Properties of MSCs for Bone Regeneration
Safety and Limitation of MSC Therapy
3.2. Cytokines, Growth Factors and Signaling Pathways Enhancing Osteogenesis
3.3. Bone Scaffolds in Tissue Engineering
3.3.1. Scaffold Properties
3.3.2. Scaffold Types
- 1.
- Bioceramics
- 2.
- Biodegradable polymers
- 3.
- Composite biomaterials
3.3.3. 3D Printing of Bone Scaffolds
3.4. Animal Models for In Vivo Studies
3.5. Preclinical Studies
4. Clinical Trials
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Chen, X.; Fan, H.; Deng, X.; Wu, L.; Yi, T.; Gu, L.; Zhou, C.; Fan, Y.; Zhang, X. Scaffold Structural Microenvironmental Cues to Guide Tissue Regeneration in Bone Tissue Applications. Nanomaterial 2018, 8, 960. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Winkler, T.; Sass, F.A.; Duda, G.N.; Schmidt-Bleek, K. A review of biomaterials in bone defect healing, remaining shortcomings and future opportunities for bone tissue engineering: The unsolved challenge. Bone Jt. Res. 2018, 7, 232–243. [Google Scholar] [CrossRef] [PubMed]
- Perez, J.R.; Kouroupis, D.; Li, D.J.; Best, T.M.; Kaplan, L.; Correa, D. Tissue Engineering and Cell-Based Therapies for Fractures and Bone Defects. Front. Bioeng. Biotechnol. 2018, 6, 105. [Google Scholar] [CrossRef] [Green Version]
- Nielsen, J.J.; Low, S.A. Bone-Targeting Systems to Systemically Deliver Therapeutics to Bone Fractures for Accelerated Healing. Curr. Osteoporos. Rep. 2020, 18, 449–459. [Google Scholar] [CrossRef]
- Chen, Y.; Xu, J.; Huang, Z.; Yu, M.; Zhang, Y.; Chen, H.; Ma, Z.; Liao, H.; Hu, J. An Innovative Approach for Enhancing Bone Defect Healing Using PLGA Scaffolds Seeded with Extracorporeal-shock-wave-treated Bone Marrow Mesenchymal Stem Cells (BMSCs). Sci. Rep. 2017, 7, 44130. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Su, P.; Tian, Y.; Yang, C.; Ma, X.; Wang, X.; Pei, J.; Qian, A. Mesenchymal Stem Cell Migration during Bone Formation and Bone Diseases Therapy. Int. J. Mol. Sci. 2018, 19, 2343. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Breeland, G.; Sinkler, M.A.; Menezes, R.G. Embryology, Bone Ossification; StatPearls: Treasure Island, FL, USA, 2020. [Google Scholar]
- White, A.; Wallis, G. Endochondral ossification: A delicate balance between growth and mineralisation. Curr. Biol. 2001, 11, R589–R591. [Google Scholar] [CrossRef] [Green Version]
- Pountos, I.; Giannoudis, P.V. Fracture Healing: Back to Basics and Latest Advances. In Fracture Reduction and Fixation Techniques: Upper Extremities; Giannoudis, P.V., Ed.; Springer International Publishing: Cham, Switzerland, 2018; pp. 3–17. [Google Scholar]
- Iaquinta, M.R.; Mazzoni, E.; Bononi, I.; Rotondo, J.C.; Mazziotta, C.; Montesi, M.; Sprio, S.; Tampieri, A.; Tognon, M.; Martini, F. Adult Stem Cells for Bone Regeneration and Repair. Front. Cell Dev. Biol. 2019, 7, 268. [Google Scholar] [CrossRef] [Green Version]
- Schell, H.; Duda, G.N.; Peters, A.; Tsitsilonis, S.; Johnson, K.A.; Schmidt-Bleek, K. The haematoma and its role in bone healing. J. Exp. Orthop. 2017, 4, 5. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Walmsley, G.G.; Ransom, R.C.; Zielins, E.R.; Leavitt, T.; Flacco, J.S.; Hu, M.S.; Lee, A.S.; Longaker, M.T.; Wan, D.C. Stem Cells in Bone Regeneration. Stem Cell Rev. Rep. 2016, 12, 524–529. [Google Scholar] [CrossRef] [Green Version]
- Sheen, J.R.; Garla, V.V. Fracture Healing Overview; StatPearls: Treasure Island, FL, USA, 2020. [Google Scholar]
- Friedenstein, A.J.; Piatetzky, S., II; Petrakova, K.V. Osteogenesis in transplants of bone marrow cells. J. Embryol. Exp. Morphol. 1966, 16, 381–390. [Google Scholar] [PubMed]
- Caplan, A.I. Mesenchymal stem cells. J. Orthop. Res. 1991, 9, 641–650. [Google Scholar] [CrossRef] [PubMed]
- Pittenger, M.F.; Discher, D.E.; Péault, B.M.; Phinney, D.G.; Hare, J.M.; Caplan, A.I. Mesenchymal stem cell perspective: Cell biology to clinical progress. NPJ Regen. Med. 2019, 4, 22. [Google Scholar] [CrossRef] [Green Version]
- Knight, M.N.; Hankenson, K.D. Mesenchymal Stem Cells in Bone Regeneration. Adv. Wound Care 2013, 2, 306–316. [Google Scholar] [CrossRef] [Green Version]
- Logeart-Avramoglou, D.; Anagnostou, F.; Bizios, R.; Petite, H. Engineering bone: Challenges and obstacles. J. Cell. Mol. Med. 2005, 9, 72–84. [Google Scholar] [CrossRef] [PubMed]
- Viateau, V.; Logeart-Avramoglou, D.; Guillemin, G.; Petite, H. Animal Models for Bone Tissue Engineering Purposes. In Sourcebook of Models for Biomedical Research; Conn, P.M., Ed.; Humana Press: Totowa, NJ, USA, 2008; pp. 725–736. [Google Scholar]
- Safarova, Y.; Umbayev, B.; Hortelano, G.; Askarova, S. Mesenchymal stem cells modifications for enhanced bone targeting and bone regeneration. Regen. Med. 2020, 15, 1579–1594. [Google Scholar] [CrossRef] [Green Version]
- Klimczak, A.; Kozlowska, U. Mesenchymal Stromal Cells and Tissue-Specific Progenitor Cells: Their Role in Tissue Homeostasis. Stem Cells Int. 2016, 2016, 4285215. [Google Scholar] [CrossRef] [Green Version]
- Kozlowska, U.; Krawczenko, A.; Futoma, K.; Jurek, T.; Rorat, M.; Patrzalek, D.; Klimczak, A. Similarities and differences between mesenchymal stem/progenitor cells derived from various human tissues. World J. Stem Cells 2019, 11, 347–374. [Google Scholar] [CrossRef] [PubMed]
- Amati, E.; Perbellini, O.; Rotta, G.; Bernardi, M.; Chieregato, K.; Sella, S.; Rodeghiero, F.; Ruggeri, M.; Astori, G. High-throughput immunophenotypic characterization of bone marrow- and cord blood-derived mesenchymal stromal cells reveals common and differentially expressed markers: Identification of angiotensin-converting enzyme (CD143) as a marker differentially expressed between adult and perinatal tissue sources. Stem Cell Res. Ther. 2018, 9, 10. [Google Scholar] [PubMed] [Green Version]
- Dominici, M.; Le Blanc, K.; Mueller, I.; Slaper-Cortenbach, I.; Marini, F.; Krause, D.; Deans, R.; Keating, A.; Prockop, D.; Horwitz, E. Minimal criteria for defining multipotent mesenchymal stromal cells. The International Society for Cellular Therapy position statement. Cytotherapy 2006, 8, 315–317. [Google Scholar] [CrossRef] [PubMed]
- Jin, H.J.; Bae, Y.K.; Kim, M.; Kwon, S.J.; Jeon, H.B.; Choi, S.J.; Kim, S.W.; Yang, Y.S.; Oh, W.; Chang, J.W. Comparative analysis of human mesenchymal stem cells from bone marrow, adipose tissue, and umbilical cord blood as sources of cell therapy. Int. J. Mol. Sci. 2013, 14, 17986–18001. [Google Scholar] [CrossRef]
- Rasini, V.; Dominici, M.; Kluba, T.; Siegel, G.; Lusenti, G.; Northoff, H.; Horwitz, E.M.; Schäfer, R. Mesenchymal stromal/stem cells markers in the human bone marrow. Cytotherapy 2013, 15, 292–306. [Google Scholar] [CrossRef] [PubMed]
- Harkness, L.; Zaher, W.; Ditzel, N.; Isa, A.; Kassem, M. CD146/MCAM defines functionality of human bone marrow stromal stem cell populations. Stem Cell Res. Ther. 2016, 7, 4. [Google Scholar] [CrossRef] [Green Version]
- Bourin, P.; Bunnell, B.A.; Casteilla, L.; Dominici, M.; Katz, A.J.; March, K.L.; Redl, H.; Rubin, J.P.; Yoshimura, K.; Gimble, J.M. Stromal cells from the adipose tissue-derived stromal vascular fraction and culture expanded adipose tissue-derived stromal/stem cells: A joint statement of the International Federation for Adipose Therapeutics and Science (IFATS) and the International Society for Cellular Therapy (ISCT). Cytotherapy 2013, 15, 641–648. [Google Scholar]
- Topoluk, N.; Hawkins, R.; Tokish, J.; Mercuri, J. Amniotic Mesenchymal Stromal Cells Exhibit Preferential Osteogenic and Chondrogenic Differentiation and Enhanced Matrix Production Compared With Adipose Mesenchymal Stromal Cells. Am. J. Sports Med. 2017, 45, 2637–2646. [Google Scholar] [CrossRef] [PubMed]
- Shen, C.; Yang, C.; Xu, S.; Zhao, H. Comparison of osteogenic differentiation capacity in mesenchymal stem cells derived from human amniotic membrane (AM), umbilical cord (UC), chorionic membrane (CM), and decidua (DC). Cell Biosci. 2019, 9, 17. [Google Scholar] [CrossRef] [PubMed]
- Murphy, M.B.; Moncivais, K.; Caplan, A.I. Mesenchymal stem cells: Environmentally responsive therapeutics for regenerative medicine. Exp. Mol. Med. 2013, 45, e54. [Google Scholar] [CrossRef] [Green Version]
- Kraskiewicz, H.; Paprocka, M.; Bielawska-Pohl, A.; Krawczenko, A.; Panek, K.; Kaczyńska, J.; Szyposzyńska, A.; Psurski, M.; Kuropka, P.; Klimczak, A. Can supernatant from immortalized adipose tissue MSC replace cell therapy? An in vitro study in chronic wounds model. Stem Cell Res. 2020, 11, 29. [Google Scholar] [CrossRef]
- Krawczenko, A.; Bielawska-Pohl, A.; Paprocka, M.; Kraskiewicz, H.; Szyposzynska, A.; Wojdat, E.; Klimczak, A. Microvesicles from Human Immortalized Cell Lines of Endothelial Progenitor Cells and Mesenchymal Stem/Stromal Cells of Adipose Tissue Origin as Carriers of Bioactive Factors Facilitating Angiogenesis. Stem Cells Int. 2020, 2020, 1289380. [Google Scholar] [CrossRef]
- Ratushnyy, A.; Ezdakova, M.; Yakubets, D.; Buravkova, L. Angiogenic Activity of Human Adipose-Derived Mesenchymal Stem Cells Under Simulated Microgravity. Stem Cells Dev. 2018, 27, 831–837. [Google Scholar] [CrossRef]
- Pevsner-Fischer, M.; Levin, S.; Zipori, D. The origins of mesenchymal stromal cell heterogeneity. Stem Cell Rev. Rep. 2011, 7, 560–568. [Google Scholar] [CrossRef] [PubMed]
- Kolf, C.M.; Cho, E.; Tuan, R.S. Mesenchymal stromal cells. Biology of adult mesenchymal stem cells: Regulation of niche, self-renewal and differentiation. Arthritis Res. 2007, 9, 204. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Klingemann, H.; Matzilevich, D.; Marchand, J. Mesenchymal Stem Cells—Sources and Clinical Applications. Transfus. Med. Hemother. 2008, 35, 272–277. [Google Scholar] [CrossRef] [PubMed]
- Strioga, M.; Viswanathan, S.; Darinskas, A.; Slaby, O.; Michalek, J. Same or not the same? Comparison of adipose tissue-derived versus bone marrow-derived mesenchymal stem and stromal cells. Stem Cells Dev. 2012, 21, 2724–2752. [Google Scholar] [CrossRef] [PubMed]
- Musial-Wysocka, A.; Kot, M.; Majka, M. The Pros and Cons of Mesenchymal Stem Cell-Based Therapies. Cell Transpl. 2019, 28, 801–812. [Google Scholar] [CrossRef] [Green Version]
- Richardson, S.M.; Kalamegam, G.; Pushparaj, P.N.; Matta, C.; Memic, A.; Khademhosseini, A.; Mobasheri, R.; Poletti, F.L.; Hoyland, J.A.; Mobasheri, A. Mesenchymal stem cells in regenerative medicine: Focus on articular cartilage and intervertebral disc regeneration. Methods 2016, 99, 69–80. [Google Scholar] [CrossRef]
- Marolt Presen, D.; Traweger, A.; Gimona, M.; Redl, H. Mesenchymal Stromal Cell-Based Bone Regeneration Therapies: From Cell Transplantation and Tissue Engineering to Therapeutic Secretomes and Extracellular Vesicles. Front. Bioeng. Biotechnol. 2019, 7, 352. [Google Scholar] [CrossRef]
- Labedz-Maslowska, A.; Bryniarska, N.; Kubiak, A.; Kaczmarzyk, T.; Sekula-Stryjewska, M.; Noga, S.; Boruczkowski, D.; Madeja, Z.; Zuba-Surma, E. Multilineage Differentiation Potential of Human Dental Pulp Stem Cells-Impact of 3D and Hypoxic Environment on Osteogenesis In Vitro. Int. J. Mol. Sci. 2020, 21, 6172. [Google Scholar] [CrossRef]
- Toyserkani, N.M.; Jørgensen, M.G.; Tabatabaeifar, S.; Jensen, C.H.; Sheikh, S.P.; Sørensen, J.A. Concise Review: A Safety Assessment of Adipose-Derived Cell Therapy in Clinical Trials: A Systematic Review of Reported Adverse Events. Stem Cells Transl. Med. 2017, 6, 1786–1794. [Google Scholar] [CrossRef] [Green Version]
- Mäkelä, T.; Takalo, R.; Arvola, O.; Haapanen, H.; Yannopoulos, F.; Blanco, R.; Ahvenjärvi, L.; Kiviluoma, K.; Kerkelä, E.; Nystedt, J.; et al. Safety and biodistribution study of bone marrow-derived mesenchymal stromal cells and mononuclear cells and the impact of the administration route in an intact porcine model. Cytotherapy 2015, 17, 392–402. [Google Scholar] [CrossRef]
- Lutton, B.V.; Cho, P.S.; Hirsh, E.L.; Ferguson, K.K.; Teague, A.G.; Hanekamp, J.S.; Chi, N.; Goldman, S.N.; Messina, D.J.; Houser, S.; et al. Approaches to avoid immune responses induced by repeated subcutaneous injections of allogeneic umbilical cord tissue-derived cells. Transplantation 2010, 90, 494–501. [Google Scholar] [CrossRef] [Green Version]
- Rather, H.A.; Jhala, D.; Vasita, R. Dual functional approaches for osteogenesis coupled angiogenesis in bone tissue engineering. Mater. Sci. Eng. C 2019, 103, 109761. [Google Scholar] [CrossRef] [PubMed]
- Gerstenfeld, L.C.; Edgar, C.M.; Kakar, S.; Jacobsen, K.A.; Einhorn, T.A. Osteogenic growth factors and cytokines and their role in bone repair. In Engineering of Functional Skeletal Tissues; Springer: Cham, Switzerland, 2007; pp. 17–45. [Google Scholar]
- Amarasekara, D.S.; Kim, S.; Rho, J. Regulation of Osteoblast Differentiation by Cytokine Networks. Int. J. Mol. Sci. 2021, 22, 2851. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.; Remark, L.H.; Josephson, A.M.; Leclerc, K.; Lopez, E.M.; Kirby, D.J.; Mehta, D.; Litwa, H.P.; Wong, M.Z.; Shin, S.Y.; et al. Notch-Wnt signal crosstalk regulates proliferation and differentiation of osteoprogenitor cells during intramembranous bone healing. NPJ Regen. Med. 2021, 6, 29. [Google Scholar] [CrossRef] [PubMed]
- Wozney, J.M. The bone morphogenetic protein family and osteogenesis. Mol. Reprod. Dev. 1992, 32, 160–167. [Google Scholar] [CrossRef]
- Zhang, J.; Li, L. BMP signaling and stem cell regulation. Dev. Biol. 2005, 284, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Urist, M.R. Bone: Formation by autoinduction. Science 1965, 150, 893–899. [Google Scholar] [CrossRef]
- Beederman, M.; Lamplot, J.D.; Nan, G.; Wang, J.; Liu, X.; Yin, L.; Li, R.; Shui, W.; Zhang, H.; Kim, S.H.; et al. BMP signaling in mesenchymal stem cell differentiation and bone formation. J. Biomed. Sci. Eng. 2013, 6, 32–52. [Google Scholar] [CrossRef] [Green Version]
- Karsenty, G. The genetic transformation of bone biology. Genes Dev. 1999, 13, 3037–3051. [Google Scholar] [CrossRef] [Green Version]
- Marupanthorn, K.; Tantrawatpan, C.; Kheolamai, P.; Tantikanlayaporn, D.; Manochantr, S. Bone morphogenetic protein-2 enhances the osteogenic differentiation capacity of mesenchymal stromal cells derived from human bone marrow and umbilical cord. Int. J. Mol. Med. 2017, 39, 654–662. [Google Scholar] [CrossRef] [Green Version]
- Gromolak, S.; Krawczenko, A.; Antonczyk, A.; Buczak, K.; Kielbowicz, Z.; Klimczak, A. Biological Characteristics and Osteogenic Differentiation of Ovine Bone Marrow Derived Mesenchymal Stem Cells Stimulated with FGF-2 and BMP-2. Int. J. Mol. Sci. 2020, 21, 9726. [Google Scholar] [CrossRef]
- Chen, G.; Deng, C.; Li, Y.P. TGF-beta and BMP signaling in osteoblast differentiation and bone formation. Int. J. Biol. Sci. 2012, 8, 272–288. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ornitz, D.M.; Marie, P.J. Fibroblast growth factor signaling in skeletal development and disease. Genes Dev. 2015, 29, 1463–1486. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Charoenlarp, P.; Rajendran, A.K.; Iseki, S. Role of fibroblast growth factors in bone regeneration. Inflamm. Regen. 2017, 37, 10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kuhn, L.T.; Ou, G.; Charles, L.; Hurley, M.M.; Rodner, C.M.; Gronowicz, G. Fibroblast growth factor-2 and bone morphogenetic protein-2 have a synergistic stimulatory effect on bone formation in cell cultures from elderly mouse and human bone. J. Gerontol. A Biol. Sci. Med. Sci. 2013, 68, 1170–1180. [Google Scholar] [CrossRef] [Green Version]
- Song, R.; Wang, D.; Zeng, R.; Wang, J. Synergistic effects of fibroblast growth factor-2 and bone morphogenetic protein-2 on bone induction. Mol. Med. Rep. 2017, 16, 4483–4492. [Google Scholar] [CrossRef]
- Derynck, R.; Akhurst, R.J. Differentiation plasticity regulated by TGF-beta family proteins in development and disease. Nat. Cell Biol. 2007, 9, 1000–1004. [Google Scholar] [CrossRef]
- Sasaki, T.; Ito, Y.; Bringas, P., Jr.; Chou, S.; Urata, M.M.; Slavkin, H.; Chai, Y. TGFbeta-mediated FGF signaling is crucial for regulating cranial neural crest cell proliferation during frontal bone development. Development 2006, 133, 371–381. [Google Scholar] [CrossRef] [Green Version]
- Barati, D.; Shariati, S.R.P.; Moeinzadeh, S.; Melero-Martin, J.M.; Khademhosseini, A.; Jabbari, E. Spatiotemporal release of BMP-2 and VEGF enhances osteogenic and vasculogenic differentiation of human mesenchymal stem cells and endothelial colony-forming cells co-encapsulated in a patterned hydrogel. J. Control. Release 2016, 223, 126–136. [Google Scholar] [CrossRef] [Green Version]
- Bao, X.; Zhu, L.; Huang, X.; Tang, D.; He, D.; Shi, J.; Xu, G. 3D biomimetic artificial bone scaffolds with dual-cytokines spatiotemporal delivery for large weight-bearing bone defect repair. Sci. Rep. 2017, 7, 7814. [Google Scholar] [CrossRef]
- Regard, J.B.; Zhong, Z.; Williams, B.O.; Yang, Y. Wnt signaling in bone development and disease: Making stronger bone with Wnts. Cold Spring Harb. Perspect. Biol. 2012, 4, a007997. [Google Scholar] [CrossRef] [Green Version]
- Hojo, H.; Ohba, S.; Chung, U.I. Signaling pathways regulating the specification and differentiation of the osteoblast lineage. Regen. Ther. 2015, 1, 57–62. [Google Scholar] [CrossRef] [Green Version]
- Gordon, M.D.; Nusse, R. Wnt signaling: Multiple pathways, multiple receptors, and multiple transcription factors. J. Biol. Chem. 2006, 281, 22429–22433. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schlickewei, C.W.; Kleinertz, H.; Thiesen, D.M.; Mader, K.; Priemel, M.; Frosch, K.-H.; Keller, J. Current and Future Concepts for the Treatment of Impaired Fracture Healing. Int. J. Mol. Sci. 2019, 20, 5805. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Latres, E.; Chiaur, D.S.; Pagano, M. The human F box protein beta-Trcp associates with the Cul1/Skp1 complex and regulates the stability of beta-catenin. Oncogene 1999, 18, 849–854. [Google Scholar] [CrossRef] [Green Version]
- Yan, Y.; Tang, D.; Chen, M.; Huang, J.; Xie, R.; Jonason, J.H.; Tan, X.; Hou, W.; Reynolds, D.; Hsu, W.; et al. Axin2 controls bone remodeling through the beta-catenin-BMP signaling pathway in adult mice. J. Cell Sci. 2009, 122(Pt. 19), 3566–3578. [Google Scholar] [CrossRef] [Green Version]
- De, A. Wnt/Ca2+ signaling pathway: A brief overview. Acta Biochim. Biophys. Sin. 2011, 43, 745–756. [Google Scholar] [CrossRef]
- Baksh, D.; Tuan, R.S. Canonical and non-canonical Wnts differentially affect the development potential of primary isolate of human bone marrow mesenchymal stem cells. J. Cell. Physiol. 2007, 212, 817–826. [Google Scholar] [CrossRef] [PubMed]
- Luo, Z.; Shang, X.; Zhang, H.; Wang, G.; Massey, P.A.; Barton, S.R.; Kevil, C.G.; Dong, Y. Notch Signaling in Osteogenesis, Osteoclastogenesis, and Angiogenesis. Am. J. Pathol. 2019, 189, 1495–1500. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Majidinia, M.; Alizadeh, E.; Yousefi, B.; Akbarzadeh, M.; Zarghami, N. Downregulation of Notch Signaling Pathway as an Effective Chemosensitizer for Cancer Treatment. Drug Res. 2016, 66, 571–579. [Google Scholar] [CrossRef] [PubMed]
- Tu, X.; Chen, J.; Lim, J.; Karner, C.M.; Lee, S.Y.; Heisig, J.; Wiese, C.; Surendran, K.; Kopan, R.; Gessler, M.; et al. Physiological notch signaling maintains bone homeostasis via RBPjk and Hey upstream of NFATc1. PLoS Genet. 2012, 8, e1002577. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhu, F.; Sweetwyne, M.T.; Hankenson, K.D. PKCdelta is required for Jagged-1 induction of human mesenchymal stem cell osteogenic differentiation. Stem Cells 2013, 31, 1181–1192. [Google Scholar] [CrossRef] [PubMed]
- Cao, J.; Wei, Y.; Lian, J.; Yang, L.; Zhang, X.; Xie, J.; Liu, Q.; Luo, J.; He, B.; Tang, M. Notch signaling pathway promotes osteogenic differentiation of mesenchymal stem cells by enhancing BMP9/Smad signaling. Int. J. Mol. Med. 2017, 40, 378–388. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bhattarai, H.K.; Shrestha, S.; Rokka, K.; Shakya, R. Vitamin D, Calcium, Parathyroid Hormone, and Sex Steroids in Bone Health and Effects of Aging. J. Osteoporos. 2020, 2020, 9324505. [Google Scholar] [CrossRef]
- Zhang, R.; Edwards, J.R.; Ko, S.Y.; Dong, S.; Liu, H.; Oyajobi, B.O.; Papasian, C.; Deng, H.W.; Zhao, M. Transcriptional regulation of BMP2 expression by the PTH-CREB signaling pathway in osteoblasts. PLoS ONE 2011, 6, e20780. [Google Scholar] [CrossRef] [Green Version]
- Xiao, L.; Fei, Y.; Hurley, M.M. FGF2 crosstalk with Wnt signaling in mediating the anabolic action of PTH on bone formation. Bone Rep. 2018, 9, 136–144. [Google Scholar] [CrossRef]
- Luo, M.; Huang, H.X.; Huang, H.; Li, Z.T.; Lai, Y.Y. Hedgehog signaling pathway and osteoporosis. Zhongguo Gu Shang 2014, 27, 169–172. [Google Scholar]
- Lv, W.T.; Du, D.H.; Gao, R.J.; Yu, C.W.; Jia, Y.; Jia, Z.F.; Wang, C.J. Regulation of Hedgehog signaling Offers A Novel Perspective for Bone Homeostasis Disorder Treatment. Int. J. Mol. Sci. 2019, 20, 3981. [Google Scholar] [CrossRef] [Green Version]
- Long, F.; Chung, U.I.; Ohba, S.; McMahon, J.; Kronenberg, H.M.; McMahon, A.P. Ihh signaling is directly required for the osteoblast lineage in the endochondral skeleton. Development 2004, 131, 1309–1318. [Google Scholar] [CrossRef] [Green Version]
- Mak, K.K.; Chen, M.H.; Day, T.F.; Chuang, P.T.; Yang, Y. Wnt/beta-catenin signaling interacts differentially with Ihh signaling in controlling endochondral bone and synovial joint formation. Development 2006, 133, 3695–3707. [Google Scholar] [CrossRef] [Green Version]
- Caddeo, S.; Boffito, M.; Sartori, S. Tissue Engineering Approaches in the Design of Healthy and Pathological In Vitro Tissue Models. Front. Bioeng. Biotechnol. 2017, 5, 40. [Google Scholar] [CrossRef] [Green Version]
- Liu, Y.; Lim, J.; Teoh, S.H. Review: Development of clinically relevant scaffolds for vascularised bone tissue engineering. Biotechnol. Adv. 2013, 31, 688–705. [Google Scholar] [CrossRef]
- Skoog, S.A.; Kumar, G.; Narayan, R.J.; Goering, P.L. Biological responses to immobilized microscale and nanoscale surface topographies. Pharm. Ther. 2018, 182, 33–55. [Google Scholar] [CrossRef]
- Osathanon, T.; Bespinyowong, K.; Arksornnukit, M.; Takahashi, H.; Pavasant, P. Human osteoblast-like cell spreading and proliferation on Ti-6Al-7Nb surfaces of varying roughness. J. Oral. Sci. 2011, 53, 23–30. [Google Scholar] [CrossRef] [Green Version]
- Nantavisai, S.; Egusa, H.; Osathanon, T.; Sawangmake, C. Mesenchymal stem cell-based bone tissue engineering for veterinary practice. Heliyon 2019, 5, e02808. [Google Scholar] [CrossRef]
- Gao, C.; Peng, S.; Feng, P.; Shuai, C. Bone biomaterials and interactions with stem cells. Bone Res. 2017, 5, 17059. [Google Scholar] [CrossRef]
- Eliaz, N.; Metoki, N. Calcium Phosphate Bioceramics: A Review of Their History, Structure, Properties, Coating Technologies and Biomedical Applications. Materials 2017, 10, 334. [Google Scholar] [CrossRef] [Green Version]
- Pan, Y.K.; Chen, C.Z.; Wang, D.G.; Lin, Z.Q. Preparation and bioactivity of micro-arc oxidized calcium phosphate coatings. Mater. Chem. Phys. 2013, 141, 842–849. [Google Scholar] [CrossRef]
- Shuai, C.; Li, P.; Liu, J.; Peng, S. Optimization of TCP/HAP ratio for better properties of calcium phosphate scaffold via selective laser sintering. Mater. Charact. 2013, 77, 23–31. [Google Scholar] [CrossRef]
- Maehira, F.; Miyagi, I.; Eguchi, Y. Effects of calcium sources and soluble silicate on bone metabolism and the related gene expression in mice. Nutrition 2009, 25, 581–589. [Google Scholar] [CrossRef] [PubMed]
- Zhou, X.; Zhang, N.; Mankoci, S.; Sahai, N. Silicates in orthopedics and bone tissue engineering materials. J. Biomed. Mater. Res. Part. A 2017, 105, 2090–2102. [Google Scholar] [CrossRef] [PubMed]
- Filipowska, J.; Pawlik, J.; Cholewa-Kowalska, K.; Tylko, G.; Pamula, E.; Niedzwiedzki, L.; Szuta, M.; Laczka, M.; Osyczka, A.M. Incorporation of sol–gel bioactive glass into PLGA improves mechanical properties and bioactivity of composite scaffolds and results in their osteoinductive properties. Biomed. Mater. 2014, 9, 065001. [Google Scholar] [CrossRef]
- Hammouche, S.; Hammouche, D.; McNicholas, M. Biodegradable bone regeneration synthetic scaffolds: In tissue engineering. Curr. Stem Cell Res. 2012, 7, 134–142. [Google Scholar] [CrossRef]
- Sonia, T.A.; Sharma, C.P. An overview of natural polymers for oral insulin delivery. Drug Discov. Today 2012, 17, 784–792. [Google Scholar] [CrossRef]
- Tian, H.; Tang, Z.; Zhuang, X.; Chen, X.; Jing, X. Biodegradable synthetic polymers: Preparation, functionalization and biomedical application. Prog. Polym. Sci. 2012, 37, 237–280. [Google Scholar] [CrossRef]
- Orchel, A.; Jelonek, K.; Kasperczyk, J.; Dobrzynski, P.; Marcinkowski, A.; Pamula, E.; Orchel, J.; Bielecki, I.; Kulczycka, A. The influence of chain microstructure of biodegradable copolyesters obtained with low-toxic zirconium initiator to in vitro biocompatibility. Biomed. Res. Int. 2013, 2013, 176946. [Google Scholar] [CrossRef]
- Yang, X.B.; Roach, H.I.; Clarke, N.M.; Howdle, S.M.; Quirk, R.; Shakesheff, K.M.; Oreffo, R.O. Human osteoprogenitor growth and differentiation on synthetic biodegradable structures after surface modification. Bone 2001, 29, 523–531. [Google Scholar] [CrossRef]
- Asti, A.; Gioglio, L. Natural and synthetic biodegradable polymers: Different scaffolds for cell expansion and tissue formation. Int. J. Artif. Organs 2014, 37, 187–205. [Google Scholar] [CrossRef]
- Guo, Z.; Lee, S.E.; Kim, H.; Park, S.; Hahn, H.T.; Karki, A.B.; Young, D.P. Fabrication, characterization and microwave properties of polyurethane nanocomposites reinforced with iron oxide and barium titanate nanoparticles. Acta Mater. 2009, 57, 267–277. [Google Scholar] [CrossRef]
- Niemeyer, P.; Krause, U.; Fellenberg, J.; Kasten, P.; Seckinger, A.; Ho, A.D.; Simank, H.G. Evaluation of mineralized collagen and alpha-tricalcium phosphate as scaffolds for tissue engineering of bone using human mesenchymal stem cells. Cells Tissues Organs 2004, 177, 68–78. [Google Scholar] [CrossRef]
- De Santis, R.; Guarino, V.; Ambrosio, L. Composite Biomaterials for Bone Repair; Woodhead Publishing: Sawston, UK, 2019; pp. 273–299. [Google Scholar]
- Wang, C.; Huang, W.; Zhou, Y.; He, L.; He, Z.; Chen, Z.; He, X.; Tian, S.; Liao, J.; Lu, B.; et al. 3D printing of bone tissue engineering scaffolds. Bioact. Mater. 2020, 5, 82–91. [Google Scholar] [CrossRef]
- Mota, R.; Silva, E.O.d.; Lima, F.F.; Menezes, L.; Thiele, A.C.S. 3D Printed Scaffolds as a New Perspective for Bone Tissue Regeneration: Literature Review. Mater. Sci. Appl. 2016, 7, 430–452. [Google Scholar]
- Leong, K.F.; Cheah, C.M.; Chua, C.K. Solid freeform fabrication of three-dimensional scaffolds for engineering replacement tissues and organs. Biomaterials 2003, 24, 2363–2378. [Google Scholar] [CrossRef]
- Curran, J.M.; Chen, R.; Hunt, J.A. The guidance of human mesenchymal stem cell differentiation in vitro by controlled modifications to the cell substrate. Biomaterials 2006, 27, 4783–4793. [Google Scholar] [CrossRef] [PubMed]
- Kommareddy, K.P.; Lange, C.; Rumpler, M.; Dunlop, J.W.; Manjubala, I.; Cui, J.; Kratz, K.; Lendlein, A.; Fratzl, P. Two stages in three-dimensional in vitro growth of tissue generated by osteoblastlike cells. Biointerphases 2010, 5, 45–52. [Google Scholar] [CrossRef] [Green Version]
- Kietzmann, J.; Pitt, L.; Berthon, P. Disruptions, decisions, and destinations: Enter the age of 3-D printing and additive manufacturing. Bus. Horiz. 2015, 58, 209–215. [Google Scholar] [CrossRef]
- De Mori, A.; Peña Fernández, M.; Blunn, G.; Tozzi, G.; Roldo, M. 3D Printing and Electrospinning of Composite Hydrogels for Cartilage and Bone Tissue Engineering. Polymers 2018, 10, 285. [Google Scholar] [CrossRef] [Green Version]
- Bahraminasab, M. Challenges on optimization of 3D-printed bone scaffolds. BioMed. Eng. OnLine 2020, 19, 69. [Google Scholar] [CrossRef]
- Salmi, M.; Tuomi, J.; Paloheimo, K.S.; Björkstrand, R.; Paloheimo, M.; Salo, J.; Kontio, R.; Mesimäki, K.; Mäkitie, A.A. Patient-specific reconstruction with 3D modeling and DMLS additive manufacturing. Rapid Prototyp. J. 2012, 18, 209–214. [Google Scholar] [CrossRef]
- Ghilan, A.; Chiriac, A.P.; Nita, L.E.; Rusu, A.G.; Neamtu, I.; Chiriac, V.M. Trends in 3D Printing Processes for Biomedical Field: Opportunities and Challenges. J. Polym. Environ. 2020, 28, 1345–1367. [Google Scholar] [CrossRef]
- Gopinathan, J.; Noh, I. Recent trends in bioinks for 3D printing. Biomater. Res. 2018, 22, 11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, J.-J.; Huang, L.-M.; Zhao, Q.; Xie, T. 4D Printing: History and Recent Progress. Chin. J. Polym. Sci. 2018, 36, 563–575. [Google Scholar] [CrossRef]
- Ferguson, J.C.; Tangl, S.; Barnewitz, D.; Genzel, A.; Heimel, P.; Hruschka, V.; Redl, H.; Nau, T. A large animal model for standardized testing of bone regeneration strategies. BMC Vet. Res. 2018, 14, 330. [Google Scholar] [CrossRef]
- Houdebine, L.M. Transgenic animal models in biomedical research. Methods Mol. Biol. 2007, 360, 163–202. [Google Scholar] [PubMed]
- Simpson, A.H.; Murray, I.R. Osteoporotic fracture models. Curr. Osteoporos. Rep. 2015, 13, 9–15. [Google Scholar] [CrossRef] [PubMed]
- Muschler, G.F.; Raut, V.P.; Patterson, T.E.; Wenke, J.C.; Hollinger, J.O. The design and use of animal models for translational research in bone tissue engineering and regenerative medicine. Tissue Eng. Part B Rev. 2010, 16, 123–145. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Muschler, G.F.; Nakamoto, C.; Griffith, L.G. Engineering principles of clinical cell-based tissue engineering. J. Bone Jt. Surg. Am. 2004, 86, 1541–1558. [Google Scholar] [CrossRef] [PubMed]
- Cibelli, J.; Emborg, M.E.; Prockop, D.J.; Roberts, M.; Schatten, G.; Rao, M.; Harding, J.; Mirochnitchenko, O. Strategies for improving animal models for regenerative medicine. Cell Stem Cell 2013, 12, 271–274. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Grün, N.; Hahn, D.; Okutan, B.; Marek, R.; Weinberg, A.-M. Animal Models in Orthopedic Research: The Proper Animal Model to Answer Fundamental Questions on Bone Healing Depending on Pathology and Implant Material; IntechOpen: London, UK, 2019. [Google Scholar]
- Thompson, D.D.; Simmons, H.A.; Pirie, C.M.; Ke, H.Z. FDA Guidelines and animal models for osteoporosis. Bone 1995, 17 (Suppl. S4), 125S–133S. [Google Scholar] [CrossRef]
- Einhorn, T.A. Clinically applied models of bone regeneration in tissue engineering research. Clin. Orthop. Relat. Res. 1999, 367, S59–S67. [Google Scholar] [CrossRef] [PubMed]
- Assis-Ribas, T.; Forni, M.F.; Winnischofer, S.M.B.; Sogayar, M.C.; Trombetta-Lima, M. Extracellular matrix dynamics during mesenchymal stem cells differentiation. Dev. Biol. 2018, 437, 63–74. [Google Scholar] [CrossRef] [PubMed]
- Baba, S.; Inoue, T.; Hashimoto, Y.; Kimura, D.; Ueda, M.; Sakai, K.; Matsumoto, N.; Hiwa, C.; Adachi, T.; Hojo, M. Effectiveness of scaffolds with pre-seeded mesenchymal stem cells in bone regeneration—Assessment of osteogenic ability of scaffolds implanted under the periosteum of the cranial bone of rats. Dent. Mater. J. 2010, 29, 673–681. [Google Scholar] [CrossRef] [Green Version]
- Ni, P.; Fu, S.; Fan, M.; Guo, G.; Shi, S.; Peng, J.; Luo, F.; Qian, Z. Preparation of poly(ethylene glycol)/polylactide hybrid fibrous scaffolds for bone tissue engineering. Int. J. Nanomed. 2011, 6, 3065–3075. [Google Scholar]
- Yokoya, S.; Mochizuki, Y.; Natsu, K.; Omae, H.; Nagata, Y.; Ochi, M. Rotator cuff regeneration using a bioabsorbable material with bone marrow-derived mesenchymal stem cells in a rabbit model. Am. J. Sports Med. 2012, 40, 1259–1268. [Google Scholar] [CrossRef]
- Liang, H.; Li, X.; Shimer, A.L.; Balian, G.; Shen, F.H. A novel strategy of spine defect repair with a degradable bioactive scaffold preloaded with adipose-derived stromal cells. Spine J. 2014, 14, 445–454. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Yamaoka, K.; Sonomoto, K.; Kaneko, H.; Satake, M.; Yamamoto, Y.; Kondo, M.; Zhao, J.; Miyagawa, I.; Yamagata, K.; et al. Local delivery of mesenchymal stem cells with poly-lactic-co-glycolic acid nano-fiber scaffold suppress arthritis in rats. PLoS ONE 2014, 9, e114621. [Google Scholar] [CrossRef] [Green Version]
- Berner, A.; Henkel, J.; Woodruff, M.A.; Steck, R.; Nerlich, M.; Schuetz, M.A.; Hutmacher, D.W. Delayed minimally invasive injection of allogenic bone marrow stromal cell sheets regenerates large bone defects in an ovine preclinical animal model. Stem Cells Transl. Med. 2015, 4, 503–512. [Google Scholar] [CrossRef] [PubMed]
- Yoon, D.; Kang, B.J.; Kim, Y.; Lee, S.H.; Rhew, D.; Kim, W.H.; Kweon, O.K. Effect of serum-derived albumin scaffold and canine adipose tissue-derived mesenchymal stem cells on osteogenesis in canine segmental bone defect model. J. Vet. Sci. 2015, 16, 397–404. [Google Scholar] [CrossRef] [Green Version]
- Masaoka, T.; Yoshii, T.; Yuasa, M.; Yamada, T.; Taniyama, T.; Torigoe, I.; Shinomiya, K.; Okawa, A.; Morita, S.; Sotome, S. Bone Defect Regeneration by a Combination of a beta-Tricalcium Phosphate Scaffold and Bone Marrow Stromal Cells in a Non-Human Primate Model. Open Biomed. Eng. J. 2016, 10, 2–11. [Google Scholar] [CrossRef] [Green Version]
- Decambron, A.; Fournet, A.; Bensidhoum, M.; Manassero, M.; Sailhan, F.; Petite, H.; Logeart-Avramoglou, D.; Viateau, V. Low-dose BMP-2 and MSC dual delivery onto coral scaffold for critical-size bone defect regeneration in sheep. J. Orthop. Res. 2017, 35, 2637–2645. [Google Scholar] [CrossRef]
- Smith, J.O.; Tayton, E.R.; Khan, F.; Aarvold, A.; Cook, R.B.; Goodship, A.; Bradley, M.; Oreffo, R.O. Large animal in vivo evaluation of a binary blend polymer scaffold for skeletal tissue-engineering strategies; translational issues. J. Tissue Eng. Regen. Med. 2017, 11, 1065–1076. [Google Scholar] [CrossRef] [Green Version]
- Zhang, D.; Gao, P.; Li, Q.; Li, J.; Li, X.; Liu, X.; Kang, Y.; Ren, L. Engineering biomimetic periosteum with beta-TCP scaffolds to promote bone formation in calvarial defects of rats. Stem Cell Res. 2017, 8, 134. [Google Scholar]
- Chu, W.; Gan, Y.; Zhuang, Y.; Wang, X.; Zhao, J.; Tang, T.; Dai, K. Mesenchymal stem cells and porous beta-tricalcium phosphate composites prepared through stem cell screen-enrich-combine(-biomaterials) circulating system for the repair of critical size bone defects in goat tibia. Stem Cell Res. 2018, 9, 157. [Google Scholar]
- Lin, C.C.; Lin, S.C.; Chiang, C.C.; Chang, M.C.; Lee, O.K. Reconstruction of Bone Defect Combined with Massive Loss of Periosteum Using Injectable Human Mesenchymal Stem Cells in Biocompatible Ceramic Scaffolds in a Porcine Animal Model. Stem Cells Int. 2019, 2019, 6832952. [Google Scholar] [CrossRef] [PubMed]
- Liu, Z.; Ge, Y.; Zhang, L.; Wang, Y.; Guo, C.; Feng, K.; Yang, S.; Zhai, Z.; Chi, Y.; Zhao, J.; et al. The effect of induced membranes combined with enhanced bone marrow and 3D PLA-HA on repairing long bone defects in vivo. J. Tissue Eng. Regen. Med. 2020, 14, 1403–1414. [Google Scholar] [CrossRef]
- Yea, J.H.; Bae, T.S.; Kim, B.J.; Cho, Y.W.; Jo, C.H. Regeneration of the rotator cuff tendon-to-bone interface using umbilical cord-derived mesenchymal stem cells and gradient extracellular matrix scaffolds from adipose tissue in a rat model. Acta Biomater. 2020, 114, 104–116. [Google Scholar] [CrossRef] [PubMed]
- Giannotti, S.; Trombi, L.; Bottai, V.; Ghilardi, M.; D’Alessandro, D.; Danti, S.; Dell’Osso, G.; Guido, G.; Petrini, M. Use of autologous human mesenchymal stromal cell/fibrin clot constructs in upper limb non-unions: Long-term assessment. PLoS ONE 2013, 8, e73893. [Google Scholar] [CrossRef]
- Perel, P.; Roberts, I.; Sena, E.; Wheble, P.; Briscoe, C.; Sandercock, P.; Macleod, M.; Mignini, L.E.; Jayaram, P.; Khan, K.S. Comparison of treatment effects between animal experiments and clinical trials: Systematic review. BMJ 2007, 334, 197. [Google Scholar] [CrossRef] [Green Version]
- Denayer, T.; Stöhr, T.; Van Roy, M. Animal models in translational medicine: Validation and prediction. New Horiz. Transl. Med. 2014, 2, 5–11. [Google Scholar] [CrossRef] [Green Version]
- Yamasaki, T.; Yasunaga, Y.; Ishikawa, M.; Hamaki, T.; Ochi, M. Bone-marrow-derived mononuclear cells with a porous hydroxyapatite scaffold for the treatment of osteonecrosis of the femoral head: A preliminary study. J. Bone Jt. Surg. Br. 2010, 92, 337–341. [Google Scholar] [CrossRef] [Green Version]
- Jager, M.; Herten, M.; Fochtmann, U.; Fischer, J.; Hernigou, P.; Zilkens, C.; Hendrich, C.; Krauspe, R. Bridging the gap: Bone marrow aspiration concentrate reduces autologous bone grafting in osseous defects. J. Orthop. Res. 2011, 29, 173–180. [Google Scholar] [CrossRef] [PubMed]
- Cuthbert, R.J.; Churchman, S.M.; Tan, H.B.; McGonagle, D.; Jones, E.; Giannoudis, P.V. Induced periosteum a complex cellular scaffold for the treatment of large bone defects. Bone 2013, 57, 484–492. [Google Scholar] [CrossRef] [PubMed]
- Šponer, P.; Filip, S.; Kučera, T.; Brtková, J.; Urban, K.; Palička, V.; Kočí, Z.; Syka, M.; Bezrouk, A.; Syková, E. Utilizing Autologous Multipotent Mesenchymal Stromal Cells and β-Tricalcium Phosphate Scaffold in Human Bone Defects: A Prospective, Controlled Feasibility Trial. Biomed. Res. Int. 2016, 2016, 2076061. [Google Scholar] [CrossRef]
- Zhuang, Y.; Gan, Y.; Shi, D.; Zhao, J.; Tang, T.; Dai, K. A novel cytotherapy device for rapid screening, enriching and combining mesenchymal stem cells into a biomaterial for promoting bone regeneration. Sci. Rep. 2017, 7, 15463. [Google Scholar] [CrossRef] [Green Version]
- Chu, W.; Wang, X.; Gan, Y.; Zhuang, Y.; Shi, D.; Liu, F.; Sun, Y.; Zhao, J.; Tang, T.; Dai, K. Screen-enrich-combine circulating system to prepare MSC/beta-TCP for bone repair in fractures with depressed tibial plateau. Regen. Med. 2019, 14, 555–569. [Google Scholar] [CrossRef] [PubMed]
- Morrison, D.A.; Kop, A.M.; Nilasaroya, A.; Sturm, M.; Shaw, K.; Honeybul, S. Cranial reconstruction using allogeneic mesenchymal stromal cells: A phase 1 first-in-human trial. J. Tissue Eng. Regen. Med. 2018, 12, 341–348. [Google Scholar] [CrossRef]
- Gjerde, C.; Mustafa, K.; Hellem, S.; Rojewski, M.; Gjengedal, H.; Yassin, M.A.; Feng, X.; Skaale, S.; Berge, T.; Rosen, A.; et al. Cell therapy induced regeneration of severely atrophied mandibular bone in a clinical trial. Stem Cell Res. 2018, 9, 213. [Google Scholar] [CrossRef]
- Gomez-Barrena, E.; Padilla-Eguiluz, N.; Rosset, P.; Gebhard, F.; Hernigou, P.; Baldini, N.; Rouard, H.; Sensebe, L.; Gonzalo-Daganzo, R.M.; Giordano, R.; et al. Early efficacy evaluation of mesenchymal stromal cells (MSC) combined to biomaterials to treat long bone non-unions. Injury 2020, 51 (Suppl. S1), S63–S73. [Google Scholar] [CrossRef] [Green Version]


{kind=link}
| Animal | Cells (Suspension) | Scaffold | Treated Side | Results | References |
|---|---|---|---|---|---|
| Rat | Human BM-MSCs (2 × 106 cells/mL) | PLLA | Cranial bone defect | Pre-seeding an MSCs-scaffold construct leads to a higher osteogenic capacity than for MSCs injected into a scaffold during surgery. | [129] |
| Rat | BM-MSCs | PEG/PLA | Thigh muscle pouches | An MSCs-scaffold construct had an excellent osteogenic potential in vitro and a good biocompatibility in vivo. | [130] |
| Rabbit | BM-MSCs | PGA | Defect of infraspinatus tendons | 16 weeks after implantation, mechanical analysis and the tendon maturing score showed higher values in the MSC-scaffold treated group than in the PGA-only treated rabbits. | [131] |
| Rat | AT-MSCs (104 cells per scaffold) | PLGA | Vertebral body of the spine defect | Between 2 and 4 weeks after MSC-scaffold construct implantation, bone formation occurred. However, in the group treated with osteogenic-induced AT-MSCs and a scaffold, a second bone formation occurred, contrary to the non-induced group. | [132] |
| Rat | Human BM-MSCs (2 × 104 cells/cm2 or 2 × 105 cells/cm2 of scaffold) | nano-fiber PLGA | Collagen-induced arthritis | An MSCs-scaffold construct suppressed bone destruction and arthritis in rats. | [133] |
| Sheep | BM-MSCs (100 × 106 cells) | PCL-HA + CaP | Segmental tibial bone defect | For a delayed injection of BM-MSCs into a scaffold, 4 weeks after biomaterial implantation biomechanical testing and micro-CT analysis showed improved bone regeneration compared to previously-seeded PCL-HA-cell construct or scaffold-only group. | [134] |
| Canine | AT-MSCs (1 × 106 cells/50 µL PBS) | (1) autologous serum-derived albumin (ASA) scaffold, (2) ASA + β-TCP | Segmental ulna bone defect | 16 weeks post-implantation, radiograph and histomorphometric analysis showed the most extensive new bone formation in ASA with AT-MSCs compared to untreated, ASA-only, and ASA+β-TCP with or without AT-MSCs. | [135] |
| Monkey | BM-MSCs (1.3–4.1 × 106/mL) | β-TCP | Segmental femoral bone defect | 12 weeks after transplantation, β-TCP + AT-MSCs treatment led to a higher success rate of bone regeneration compared to β-TCP treatment alone. | [136] |
| Sheep | BM-MSCs (107 cells) | coral scaffold | Long metatarsal bone defect | 4 months post implantation, micro-CT and histological analysis showed better bone formation in the group treated with the construct scaffold + BMP-2 + BM-MSCs compared to scaffold + BMP-2 or scaffold + BM-MSCs. | [137] |
| Sheep | BM-MSCs (107 cells) | PLLA-PCL | Segmental tibial bone defect | 12 weeks after implantation, significant bone regeneration was confirmed with micro-CT, mechanical testing and histological analysis in the group treated with PLLA-PCL + BM-MSCs compared to PLLA-PCL-only and untreated group. | [138] |
| Rat | BM-MSCs, osteogenic and endothelial differentiated BM-MSCs (5 × 104 cells/cm2 BM-MSCs sheet)—biomimetic periosteum (BP) | β-TCP | Calvarial defect | 8 weeks post-surgery, micro-CT and histological analysis showed better new bone formation in β-TCP + BP and β-TCP + autologous periosteum groups than in the control groups. | [139] |
| Goat | BM-MSCs | β-TCP | Critical size bone defects in tibia | 6 months after operation X-ray, micro-CT and histological analysis showed that the defect treatment using β-TCP + BM-MSCs was significantly superior to that using β-TCP alone. | [140] |
| Pig | Human AT-MSCs | TCP | Segmental long bone defect | 8 and 12 weeks after reconstruction, radiographic images and pathological sections analysis showed that TCP + human AT-MSCs promoted bone healing. | [141] |
| Rabbit | BM-MSCs | PLA-HA | Radius long bone defect | 8, 12, and 16 weeks post transplantation, micro-CT, X-ray and histological analysis showed enhanced bone reconstruction in PLA-HA + BM-MSCs combined with induced membrane group compared to the other groups. | [142] |
| Rat | Human UC-MSCs (2 × 105 cells) | HA-G | Tendon-to-bone interface | After 8 weeks, histological and biomechanical evaluation showed that the total regeneration score was significantly higher in the HA-G + UC MSC group compared to the other groups. | [143] |
| Study Number | Disease | Cells (Suspension) | Scaffold | Patients (Groups) | Results | References |
|---|---|---|---|---|---|---|
| Not reported | Osteonecrosis of the femoral head | BMMNCs (1 × 109 cells in 40 mL) | IP-CHA | 30 patients: 8 patients treated with cell-free IP-CHA (control group) and 22 patients with IP-CHA + BMMNCs | 29 weeks after surgery in the IP-CHA- and BMMNC-treated group, the osteonecrotic lesion decreased in size. In the control group, a severe collapse of the femoral head occurred in 6 patients. | [147] |
| Study #3096 Ethics Committee of the Heinrich Heine University Duesseldorf | Local bone defects larger than 1 cm × 1 cm | BMAC (8 mL) | Col or HA | 39 patients: 12 patients treated with Col + BMAC and 27 patients with HA + BMAC | New bone formation was observed in all treated patients; however, it appeared earlier in the HA group (6.8 weeks) compared to Col (13.6 weeks). | [148] |
| Not reported | Critical size bone defects | IM as a complex cellular scaffold (rich source of MSCs) | 8 patients | Cellular composition and molecular profile of IM-promoted large defect repair. | [149] | |
| 3766/2012 Comitato Etico Sperimentazione Farmaco CESF, Azienda Ospedaliero-Universitaria Pisana, Pisa, Italy | Upper limb atrophic pseudarthrosis | BM-MSCs (0.5 × 106 –2.0 × 106 cells in 2 mL of autologous plasma) | Autologous fibrin clots | 8 patients | In all patients, recovery of limb functions was observed. | [144] |
| EudraCT number 2012-005599-33 EU Clinical Trials Register | Femoral defects | BM-MSCs (15 ± 4.5 × 106 cells in 1.5 mL) | β-TCP | 18 patients: 9 patients treated with β-TCP alone (control group) and 9 patients with β-TCP + BM-MSCs | 12 months after surgery, in all 9 patients treated with β-TCP and BM-MSCs, trabecular remodeling was detected, and in the control group, only in one patient. | [150] |
| ChiCTR-ONC-17011448 Chinese Clinical Trial Registry | Non-unions and others | BM-MSCs | β-TCP | 42 patients | In all patients, radiography showed full bone healing after 9 months. | [151] |
| 2017-385-T282 Shanghai Jiao Tong University Affiliated Ninth People’s Hospital Medical Ethics Committee | Depressed tibial plateaus fractures | BM-MSCs | β-TCP | 39 patients: 23 patients treated only with β-TCP (control group) and 16 patients with β-TCP + BM-MSCs | Excellent or good recovery was observed 2 years post transplantation in 15 of 16 patients treated with MSCs/β-TCP and in 14 of 23 treated with β-TCP alone. | [152] |
| EC2012/047 Royal Perth Hospital Ethics Committee | Cranial defects | BM-MSCs (min. 0.5 × 106 cells per ml of scaffold granules) | β-TCP | 3 patients | Between 3 and 6 months post transplantation, good cranial contour restoration was maintained in all three patients. However, between 6 and 12 months, there was evidence of construct resorption. | [153] |
| EudraCT, 2012-003139-50 EU Clinical Trials Register | Severely atrophied mandibular bone | BM-MSCs (20 × 106 cells/1 cm3 of scaffold) | BCP | 11 patients | In all patients, successful ridge augmentation and new bone formation of a dental implant were observed. | [154] |
| EudraCT, 2011-005441-13 EU Clinical Trials Register | Long bone delayed and non-unions | BM-MSCs | BCP | 28 patients | 3 months after surgery, radiological consolidation amounted to 25.0% (7/28 cases), after 6 months, 67.8% (19/28 cases), and after 12 months, 92.8% (26/28 cases). | [155] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Stamnitz, S.; Klimczak, A. Mesenchymal Stem Cells, Bioactive Factors, and Scaffolds in Bone Repair: From Research Perspectives to Clinical Practice. Cells 2021, 10, 1925. https://doi.org/10.3390/cells10081925
Stamnitz S, Klimczak A. Mesenchymal Stem Cells, Bioactive Factors, and Scaffolds in Bone Repair: From Research Perspectives to Clinical Practice. Cells. 2021; 10(8):1925. https://doi.org/10.3390/cells10081925
Chicago/Turabian StyleStamnitz, Sandra, and Aleksandra Klimczak. 2021. "Mesenchymal Stem Cells, Bioactive Factors, and Scaffolds in Bone Repair: From Research Perspectives to Clinical Practice" Cells 10, no. 8: 1925. https://doi.org/10.3390/cells10081925