
Analysis of the Role of Bradysia impatiens (Diptera: Sciaridae) as a Vector Transmitting Peanut Stunt Virus on the Model Plant Nicotiana benthamiana


Abstract
:1. Introduction
2. Materials and Methods
2.1. Virus-Free Insect Cultivation
2.2. The Insect Species Identification Using PCR
2.3. Viral Strain and Plant Material
2.4. Studies on the Fungus Gnats Ability to Acquire a Virus
2.5. Virus Transmission Assay
2.6. Total RNA Isolation
2.7. RT-PCR
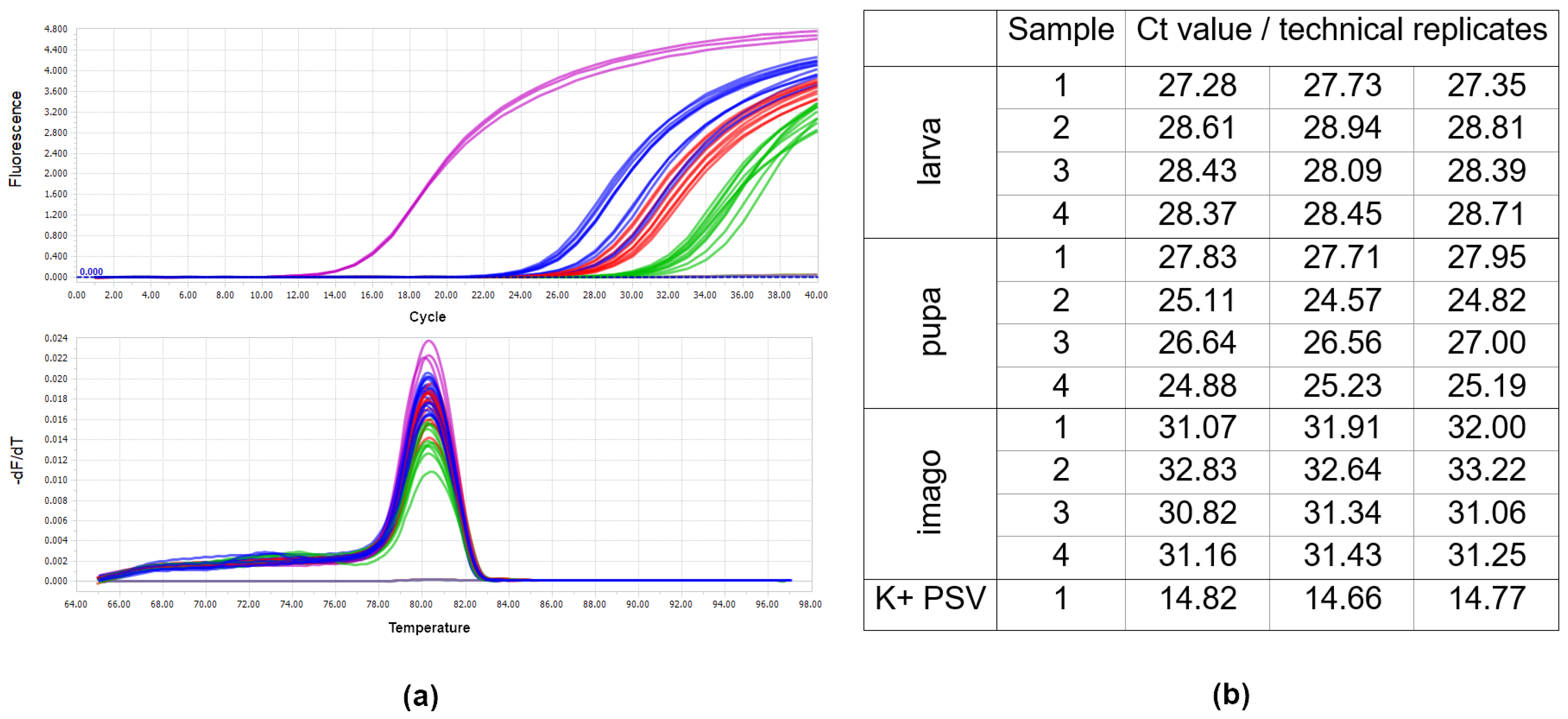
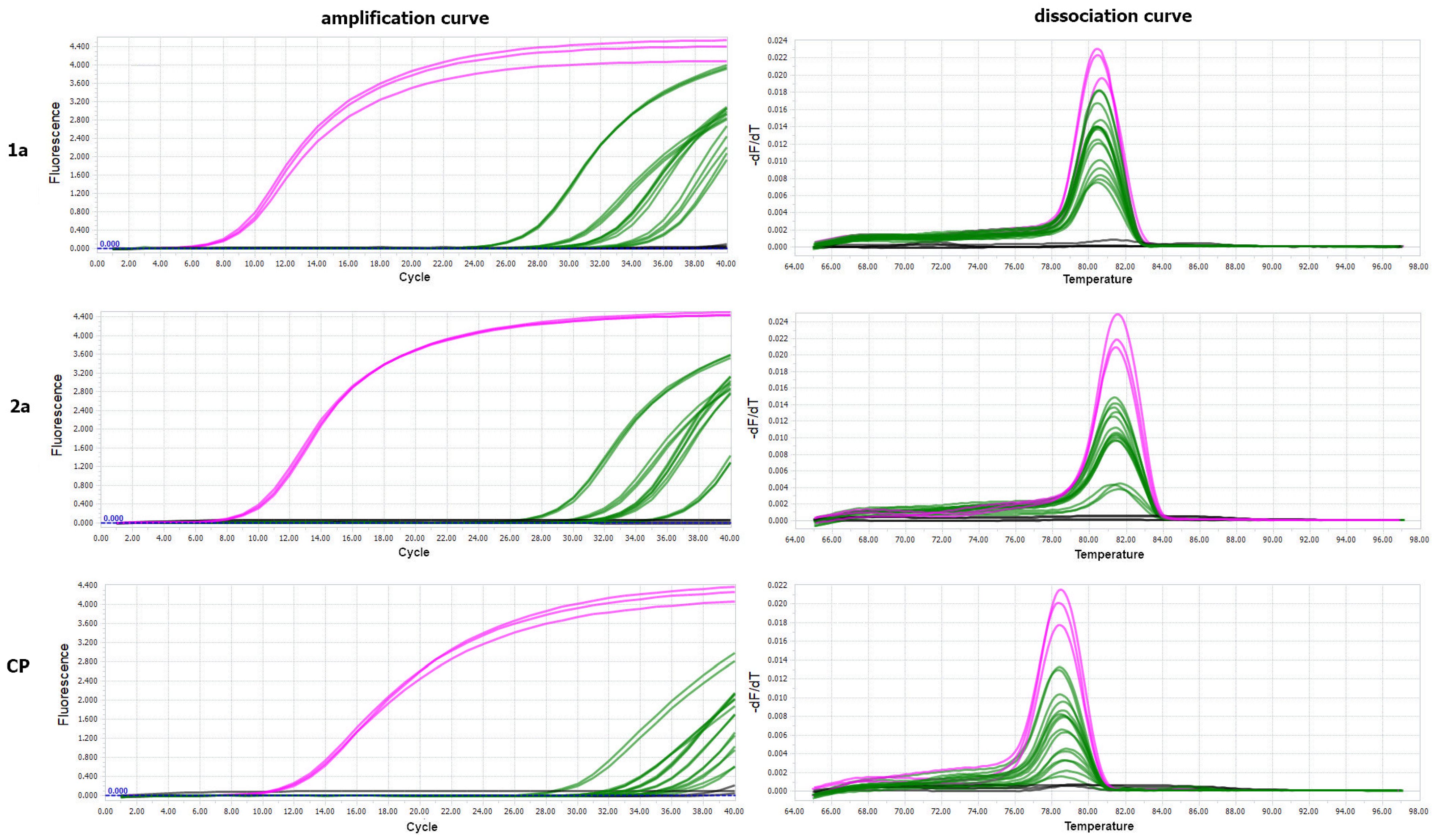
2.8. Virus Detection in Insect Individuals Using Real-Time PCR
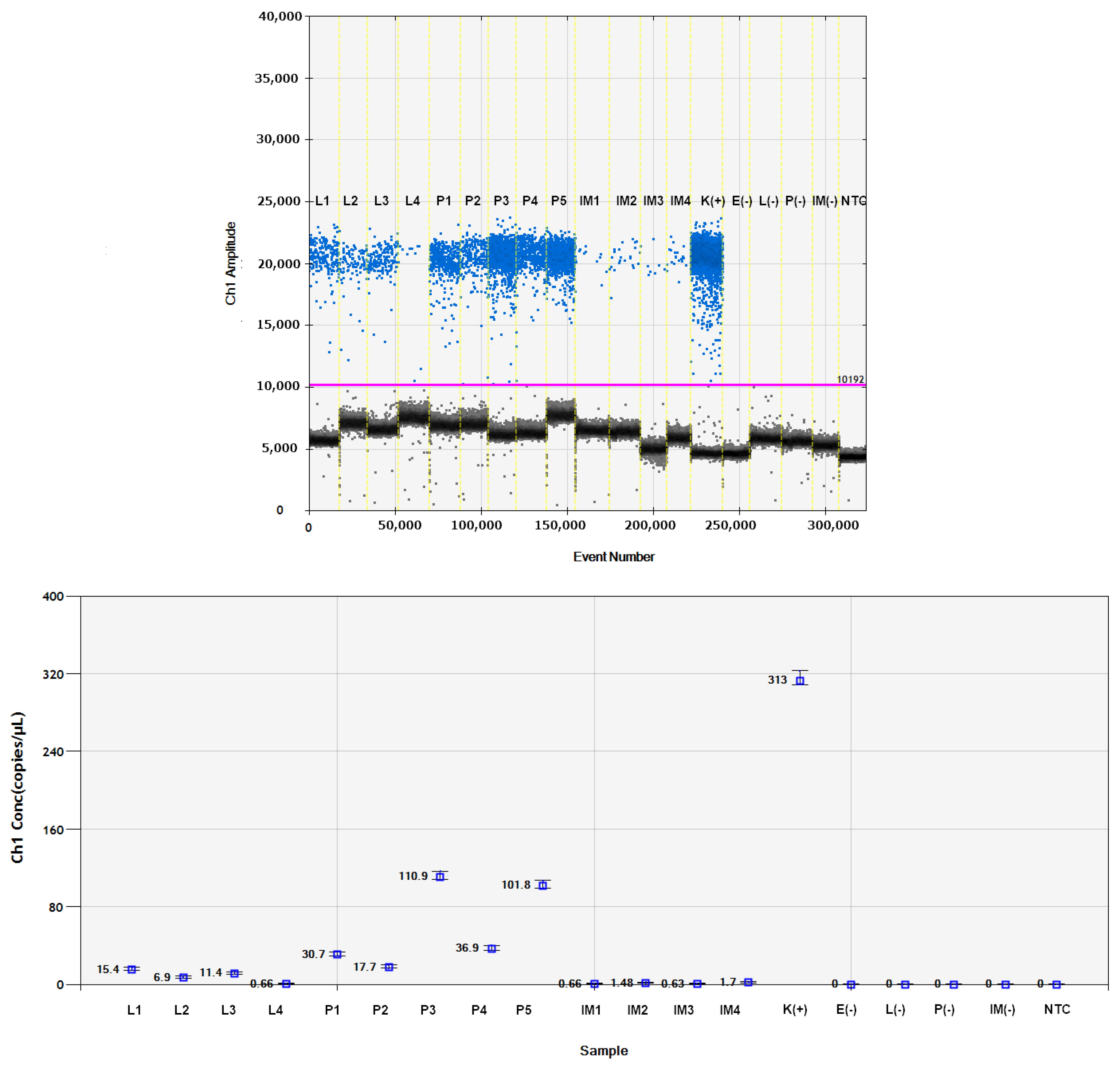
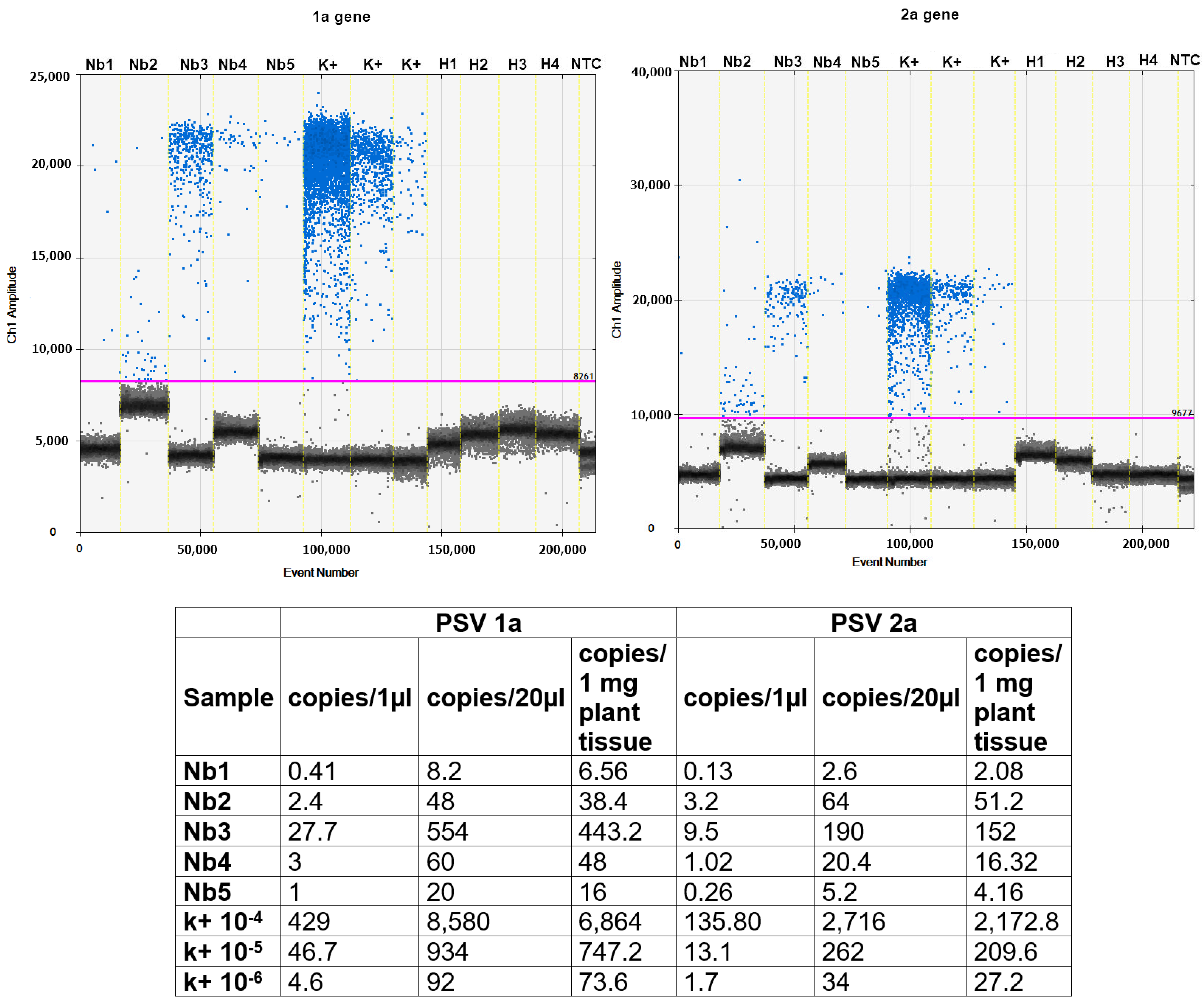
2.9. Quantitative Analysis of Viral RNAs Using Digital Droplet PCR (RT-ddPCR)
2.10. Western Blot Assay
3. Results
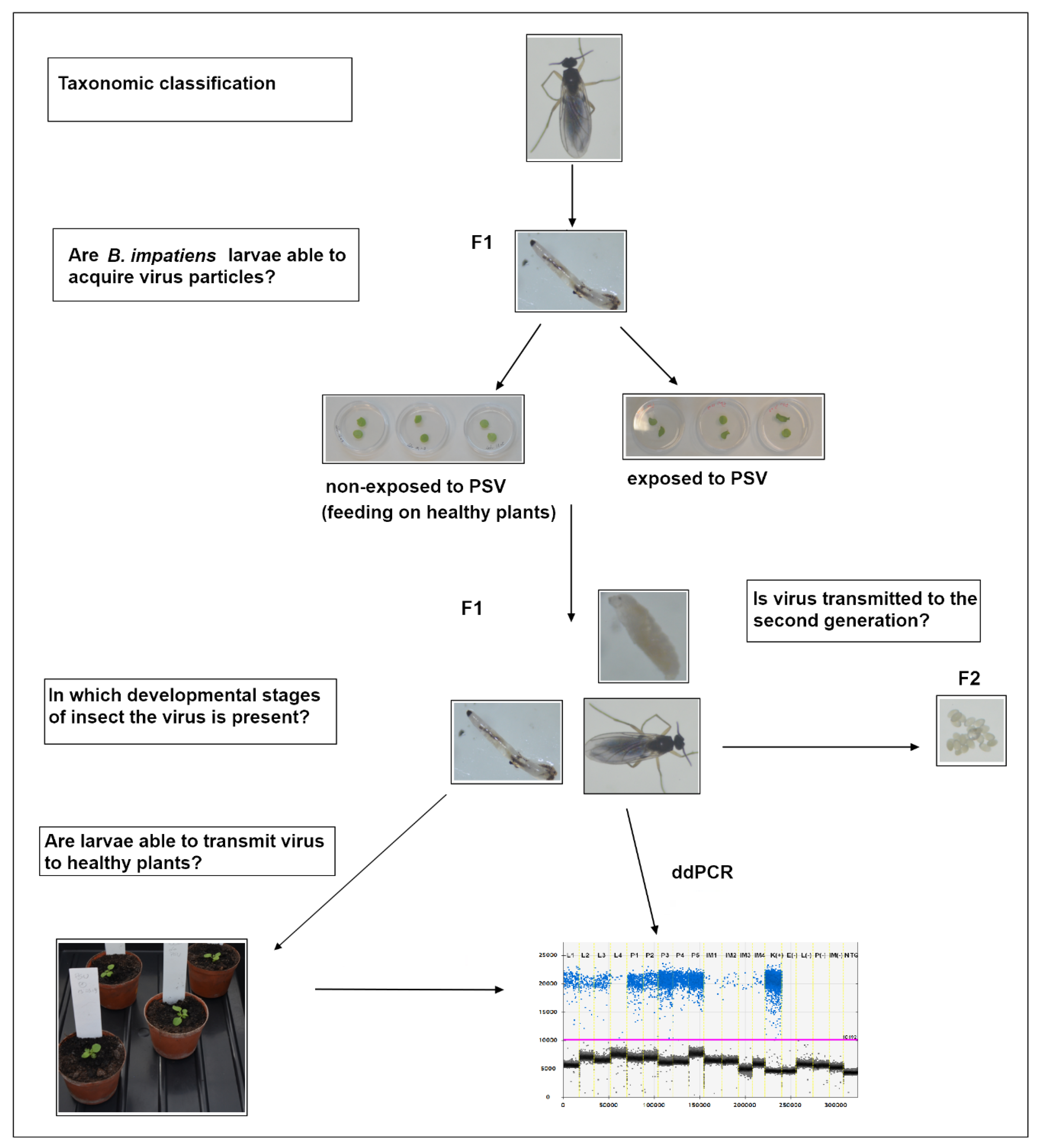
3.1. Insect Taxonomic Classification
3.2. PSV Acquisition by B. impatiens Larvae
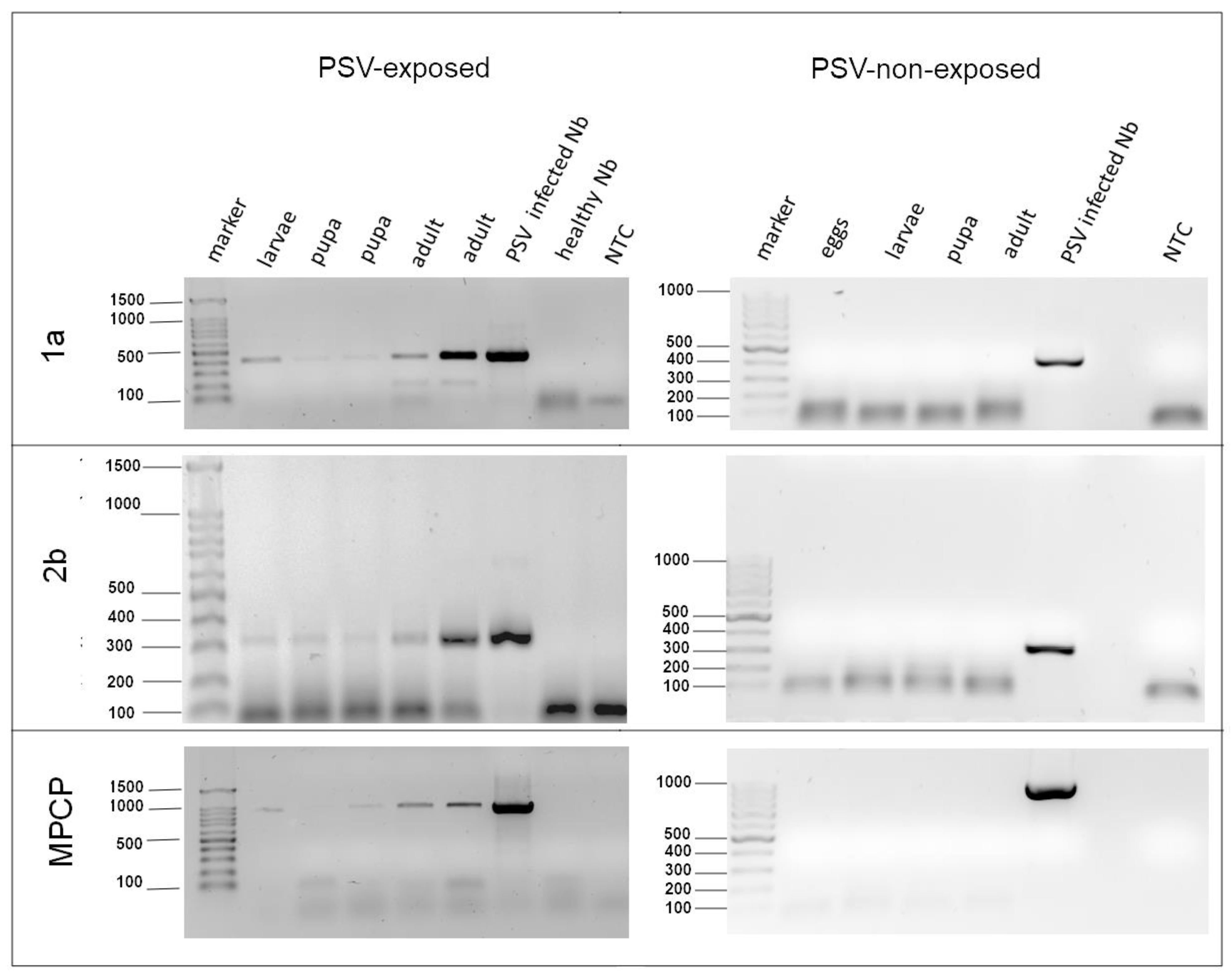
3.3. Analysis of Virus RNA Accumulation in Individuals of Fungus Gnats at Various Developmental Stages
3.4. Bradysia impatiens Ability to Vector PSV to Healthy Plants
3.5. Detection of PSV by Western Blotting
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Katumanyane, A.; Ferreira, T.; Malan, A.P. A Review of Bradysia spp. (Diptera: Sciaridae) as Pests in Nursery and Glasshouse Crops, with Special Reference to Biological Control Using Entomopathogenic Nematodes. Afr. Entomol. 2018, 26, 1–13. [Google Scholar] [CrossRef]
- Menzel, F.; Smith, J.E.; Colauto, N.B. Bradysia difformis Frey and Bradysia ocellaris (Comstock): Two additional neotropical species of black fungus gnats (Diptera: Sciaridae) of economic importance: A redescription and review. Ann. Entomol. Soc. Am. 2003, 96, 448–457. [Google Scholar] [CrossRef]
- Rebora, M.; Salerno, G.; Piersanti, S.; Gorb, E.; Gorb, S. Entrapment of Bradysia paupera (Diptera: Sciaridae) by Phaseolus vulgaris (Fabaceae) plant leaf. Arthropod-Plant Interact. 2020, 14, 499–509. [Google Scholar] [CrossRef]
- Cloyd, R.A. Ecology of fungus gnats (Bradysia spp.) in greenhouse production systems associated with disease-interactions and alternative management strategies. Insects 2015, 6, 325–332. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wilkinson, J.D.; Daugherty, D.M. The biology and immature stages of Bradysia impatiens (Diptera: Sciaridae). Ann. Entomol. Soc. Am. 1970, 63, 656–660. [Google Scholar] [CrossRef]
- Woelke, J.B.; Pham, K.; Humala, A.E. New species of Stenomacrus (Hymenoptera: Ichneumonidae: Orthocentrinae) reared from Bradysia impatiens (Diptera: Sciaridae) in The Netherlands. J. Nat. Hist. 2020, 54, 1603–1616. [Google Scholar] [CrossRef]
- Hall, R.D.; Gerhardt, R.R. Medical and Veterinary Entomology, 2nd ed.; Mullen, G.R., Durden, L.A., Eds.; Academic Press: Cambridge, MA, USA, 2009. [Google Scholar]
- Chabannes, M.; Hatt, G.; Thébaud, G.; Bedford, I.D.; Lamb, C. Establishment of an in vitro sciarid fly larvae assay to study plant resistance. Ann. Appl. Biol. 2009, 155, 293–296. [Google Scholar] [CrossRef]
- James, R.L.; Dumroese, R.K.; Wenny, D.L. Botrytis cinerea carried by adult fungus gnats (Diptera: Sciaridae) in container nurseries. Tree Plant Notes 1995, 46, 48–53. [Google Scholar]
- Jagdale, G.B.; Casey, M.L.; Cañas, L.; Grewal, P.S. Effect of entomopathogenic nematode species, split application and potting medium on the control of the fungus gnat, Bradysia difformis (Diptera: Sciaridae), in the greenhouse at alternating cold and warm temperatures. Biol. Control 2007, 43, 23–30. [Google Scholar] [CrossRef]
- Leath, K.T.; Newton, R.C. Interaction of a fungus gnat Bradysia sp (Sciaridae) with Fusaium spp on alfaalfa and red clover. Phytopathology 1969, 59, 257. [Google Scholar]
- Kalb, D.W. Dispersal of Verticillium albo-atrum by the Fungus Gnat (Bradysia impatiens). Plant Dis. 1986, 70, 752. [Google Scholar] [CrossRef]
- Jarvis, W.R.; Shipp, J.L.; Gardiner, R.B. Transmission of Pythium aphanidermatum to greenhouse cucumber by the fungus gnat Bradysia impatiens (Diptera: Sciaridae). Ann. Appl. Biol. 1993, 122, 23–29. [Google Scholar] [CrossRef]
- Edwardson, J.R.; Christie, R.G. CRC Handbook of Viruses Infecting Legumes; CRC Press: Boca Raton, FL, USA, 1991. [Google Scholar]
- Braun, S.E.; Castrillo, L.A.; Sanderson, J.P.; Daughtrey, M.L.; Wraight, S.P.; Braun, S.E. Transstadial Transmission of Pythium in Bradysia impatiens and Lack of Adult Vectoring Capacity. Phytopathology 2010, 100, 1307. [Google Scholar] [CrossRef]
- Park, J.M.; You, Y.H.; Park, J.H.; Kim, H.H.; Ghim, S.Y.; Back, C.G. Cutaneous microflora from geographically isolated groups of bradysia agrestis, an insect vector of diverse plant pathogens. Mycobiology 2017, 45, 160–171. [Google Scholar] [CrossRef] [Green Version]
- Obrepalska-Steplowska, A.; Nowaczyk, K.; Budziszewska, M.; Czerwoniec, A.; Pospieszny, H. The sequence and model structure analysis of three Polish peanut stunt virus strains. Virus Genes 2008, 36, 221–229. [Google Scholar] [CrossRef] [PubMed]
- Obrȩpalska-Stȩplowska, A.; Budziszewska, M.; Wieczorek, P.; Czerwoniec, A. Analysis of two strains of Peanut stunt virus: SatRNA-associated and satRNA free. Virus Genes 2012, 44, 513–521. [Google Scholar] [CrossRef]
- Obrepalska-Steplowska, A.; Budziszewska, M.; Pospieszny, H. Complete nucleotide sequence of a Polish strain of Peanut stunt virus (PSV-P) that is related to but not a typical member of subgroup I. Acta Biochim. Pol. 2008, 55, 731–739. [Google Scholar] [CrossRef]
- Fereres, A.; Raccah, B. Plant Virus Transmission by Insects. eLS 2015, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Gardiner, R.B.; Jarvis, W.R.; Shipp, J.L. Ingestion of Pythium spp. by larvae of the fungus gnat Bradysia impatiens (Diptera: Sciaridae). Ann. Appl. Biol. 1990, 116, 205–212. [Google Scholar] [CrossRef]
- Obrepalska-Steplowska, A.; Renaut, J.; Planchon, S.; Przybylska, A.; Wieczorek, P.; Barylski, J.; Palukaitis, P. Effect of temperature on the pathogenesis, accumulation of viral and satellite RNAs and on plant proteome in peanut stunt virus and satellite RNA-infected plants. Front. Plant Sci. 2015, 6, 903. [Google Scholar] [CrossRef] [Green Version]
- Vilkamaa, P. Checklist of the family Sciaridae (Diptera) of Finland. Zookeys 2014, 441, 151–164. [Google Scholar] [CrossRef] [Green Version]
- Tungadi, T.; Donnelly, R.; Qing, L.; Iqbal, J.; Murphy, A.M.; Pate, A.E.; Cunniffe, N.J.; Carr, J.P. Cucumber mosaic virus 2b proteins inhibit virus-induced aphid resistance in tobacco. Mol. Plant Pathol. 2020, 21, 250–257. [Google Scholar] [CrossRef] [Green Version]
- Kameya-Iwaki, M.; Murakami, K.; Ito, S.; Hanada, K.; Tanaka, S. Semipersistency of Myzus persicae transmission of cucumoviruses systemically infecting leguminous plants. J. Gen. Plant Pathol. 2000, 66, 64–67. [Google Scholar] [CrossRef]
- Hebert, T.T. Epidemiology of the peanut stunt virus in North Carolina. Phytopathology 1967, 57, 461. [Google Scholar]
- El Sadiq, E.O.; Ahmed, A.H. Comparative studies on aphid transmission of the Sudanese strain of peanut stunt virus. J. Phytopathol. 1986, 115, 160–164. [Google Scholar] [CrossRef]
- Liang, Y.; Gao, X.-W. The cuticle protein gene MPCP4 of Myzus persicae (Homoptera: Aphididae) plays a critical role in cucumber mosaic virus acquisition. J. Econ. Entomol. 2017, 110, 848–853. [Google Scholar] [CrossRef]
- Krenz, B.; Bronikowski, A.; Lu, X.; Ziebell, H.; Thompson, J.R.; Perry, K.L. Visual monitoring of Cucumber mosaic virus infection in Nicotiana benthamiana following transmission by the aphid vector Myzus persicae. J. Gen. Virol. 2015, 96, 2904–2912. [Google Scholar] [CrossRef] [Green Version]
- Wamonje, F.O.; Donnelly, R.; Tungadi, T.D.; Murphy, A.M.; Pate, A.E.; Woodcock, C.; Caulfield, J.; Mutuku, J.M.; Bruce, T.J.A.; Gilligan, C.A.; et al. Different Plant Viruses Induce Changes in Feeding Behavior of Specialist and Generalist Aphids on Common Bean That Are Likely to Enhance Virus Transmission. Front. Plant Sci. 2020, 10, 1811. [Google Scholar] [CrossRef] [Green Version]
- Kameya-Iwaki, M.; Shigeyoshi, M.; Ito, S.; Tanaka, S. Effects of susceptibility of test plants on modes of Cucumovirus transmission by Myzus persicae. J. Gen. Plant Pathol. 2001, 67, 69–72. [Google Scholar] [CrossRef]
- Garzo, E.; Moreno, A.; Plaza, M.; Fereres, A. Feeding Behavior and Virus-transmission Ability of Insect Vectors Exposed to Systemic Insecticides. Plants 2020, 9, 895. [Google Scholar] [CrossRef]
- Ng, J.C.K.; Perry, K.L. Transmission of plant viruses by aphid vectors. Mol. Plant Pathol. 2004, 5, 505–511. [Google Scholar] [CrossRef] [PubMed]
- Yadav, D.; Rana, R. Transmission of plant virus through arthropod vector. J. Entomol. Zool. Stud. 2020, 8, 1934–1939. [Google Scholar]
- Kapongo, J.-P.; Kevan, P.G.; Shipp, L.; Taki, H. Making a Pest Beneficial: Fungus Gnats [Bradysia impatiens (Diptera: Sciaridea)] as Potential Vectors of Microbial Control Agents to Suppress Pathogens they Also Spread. In Entomovectoring for Precision Biocontrol and Enhanced Pollination of Crops; Springer: Berlin/Heidelberg, Germany, 2020; pp. 239–250. [Google Scholar]
- Obrepalska-Steplowska, A.; Wieczorek, P.; Budziszewska, M.; Jeszke, A.; Renaut, J. How can plant virus satellite RNAs alter the effects of plant virus infection? A study of the changes in the Nicotiana benthamiana proteome after infection by Peanut stunt virus in the presence or absence of its satellite RNA. Proteomics 2013, 13, 2162–2175. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Genomic Strand | Primer | Sequence 5′> 3′ | Amplicon Length, Annealing Temperature Ta °C | |
|---|---|---|---|---|
| RNA1 | P1a | CACAAATCCGGCTGAGAAATG | this study | 426 bp, 57.3 |
| P1b | CAAGATACCAGCGTAGATCAC | |||
| q1F | CTTCTGCCCTCGTTGATAAAG | [22] | 131bp, 57 | |
| q1R | CATACCGATTTCGAATCACTTC | |||
| RNA2 | v2b1 | TGAGAATTCAAAAAAAAAACAATGTCGAG TGTCGAGCAG | this study | 320 bp, 59 |
| v2b2 | CGCCTGCAGTTATCAGGAATAACTACCCTC | |||
| q2aF | CTTCTAGGTATCCCCGTAAG | [22] | 130 bp, 56 | |
| q2aR | CAAGCACATTGATACCCTATC | |||
| RNA3 | P3MPCP1 | GAGGTATGGTTATCTTGGACATC | [19] | 958 bp, 60 |
| P3MPCP2 | GAAGTTGAACACAGGAAACCTTC | |||
| qCP F | ACACATACACTTCGTTGGATG | [22] | 130 bp, 56 | |
| qCP R | CCTCWTCTTCGGAAATTCAG |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Budziszewska, M.; Frąckowiak, P.; Obrępalska-Stęplowska, A. Analysis of the Role of Bradysia impatiens (Diptera: Sciaridae) as a Vector Transmitting Peanut Stunt Virus on the Model Plant Nicotiana benthamiana. Cells 2021, 10, 1546. https://doi.org/10.3390/cells10061546
Budziszewska M, Frąckowiak P, Obrępalska-Stęplowska A. Analysis of the Role of Bradysia impatiens (Diptera: Sciaridae) as a Vector Transmitting Peanut Stunt Virus on the Model Plant Nicotiana benthamiana. Cells. 2021; 10(6):1546. https://doi.org/10.3390/cells10061546
Chicago/Turabian StyleBudziszewska, Marta, Patryk Frąckowiak, and Aleksandra Obrępalska-Stęplowska. 2021. "Analysis of the Role of Bradysia impatiens (Diptera: Sciaridae) as a Vector Transmitting Peanut Stunt Virus on the Model Plant Nicotiana benthamiana" Cells 10, no. 6: 1546. https://doi.org/10.3390/cells10061546