
No Evidence for Seed Transmission of Tomato Yellow Leaf Curl Sardinia Virus in Tomato



Abstract
:
1. Introduction
2. Materials and Methods
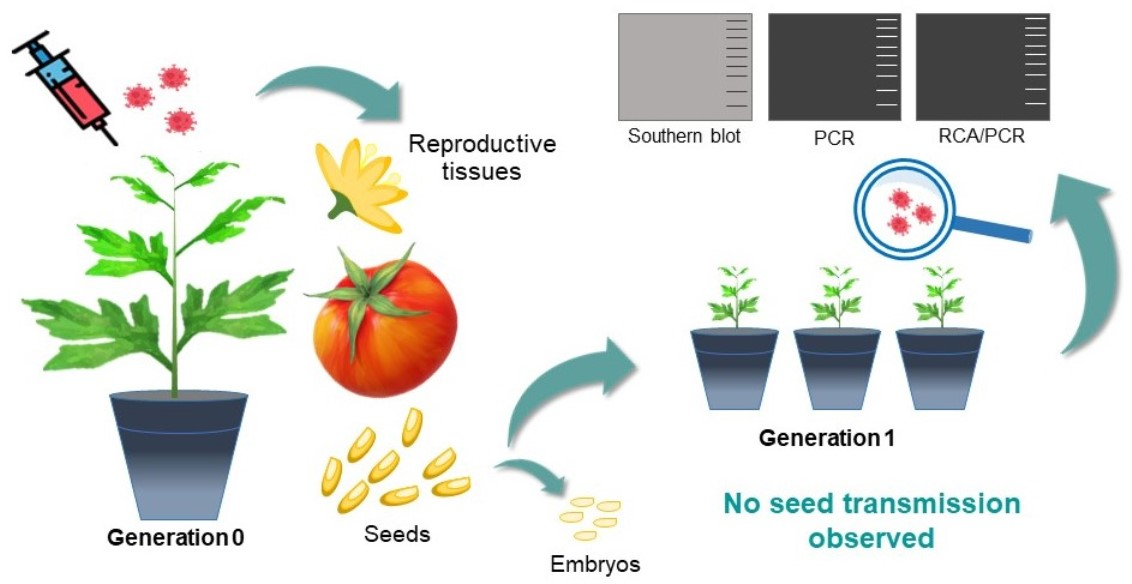
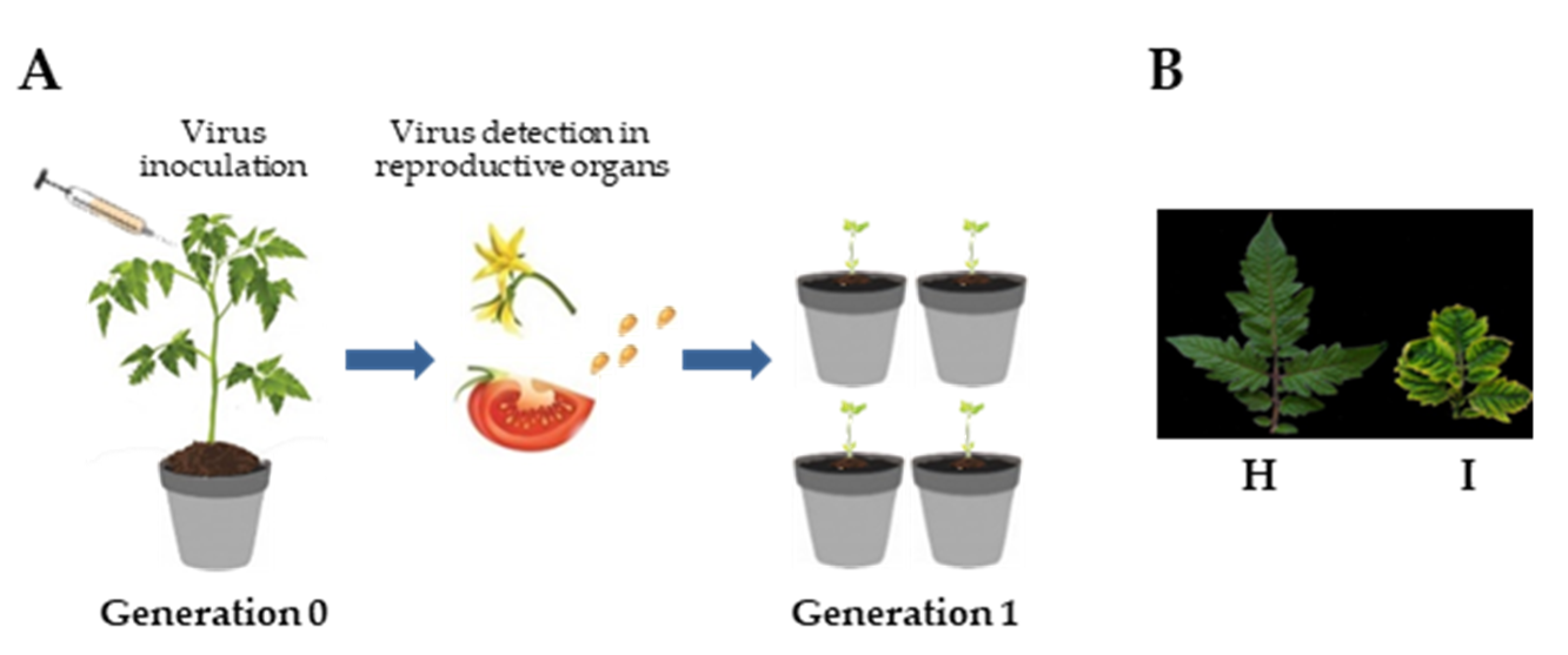
2.1. Preparation of Biological Material
2.2. Nucleic Acid Extraction
2.3. Virus Detection and Quantification
2.3.1. End-Point PCR
2.3.2. Quantitative Real-Time PCR
2.3.3. Southern Blot Assay
2.3.4. Rolling Circle Amplification (RCA)
3. Results
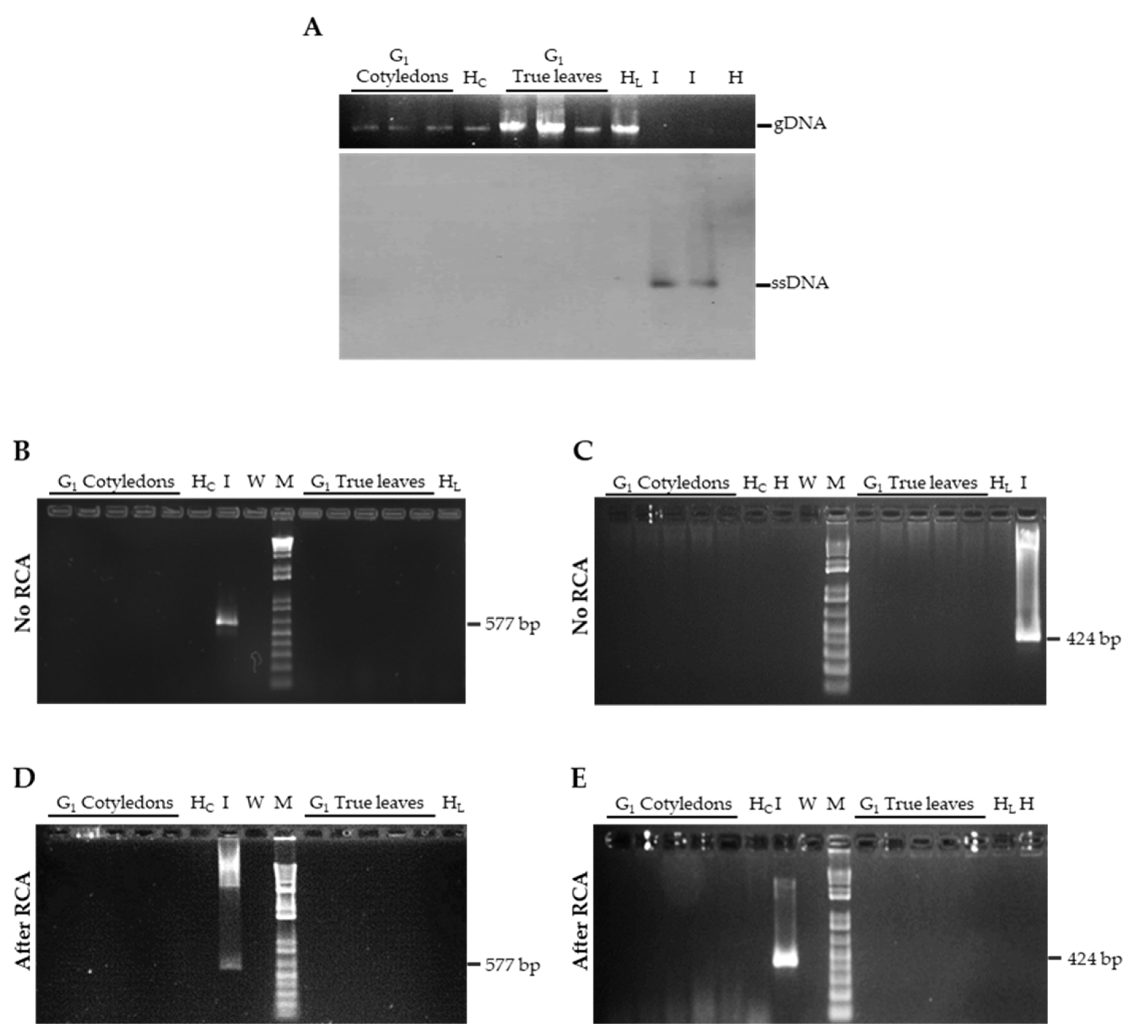
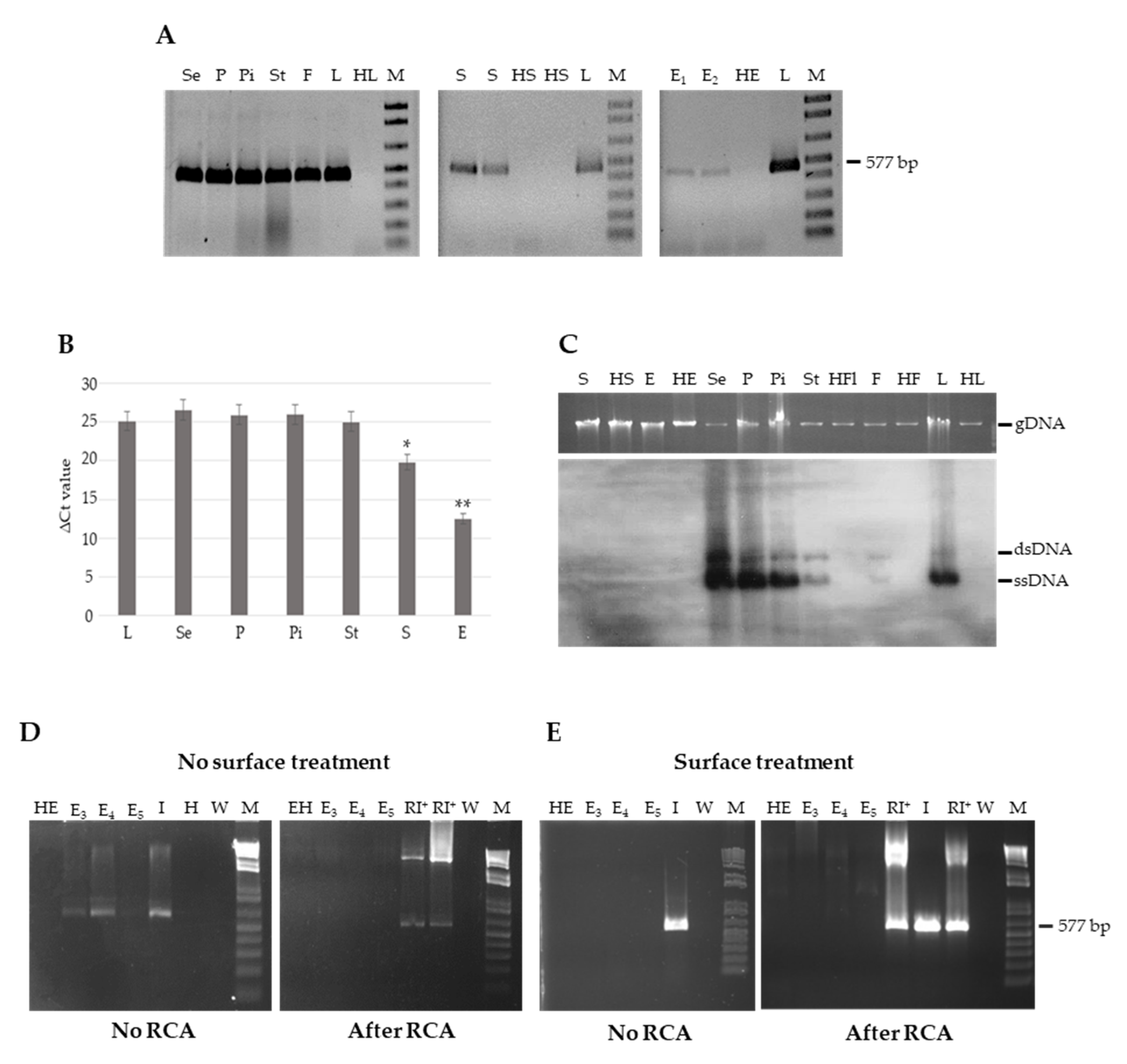
3.1. TYLCSV DNA Is not Seed-Transmitted to the Progeny of Infected Plants
3.2. Genomic TYLCSV DNA Reaches Reproductive Organs of Tomato Plants, but Is Unable to Invade Embryos
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Zerbini, F.M.; Briddon, R.W.; Idris, A.; Martin, D.P.; Moriones, E.; Navas-Castillo, J.; Rivera-Bustamante, R.; Roumagnac, P.; Varsani, A.; ICTV Report Consortium. ICTV Virus Taxonomy Profile: Geminiviridae. J. Gen. Virol. 2017, 98, 131–133. [Google Scholar] [CrossRef] [PubMed]
- Jeske, H. Geminiviruses. Curr. Top. Microbiol. Immunol. 2009, 331, 185–226. [Google Scholar]
- Navas-Castillo, J.; Fiallo-Olivé, E.; Sánchez-Campos, S. Emerging virus diseases transmitted by whiteflies. Annu. Rev. Phytopathol. 2011, 49, 219–248. [Google Scholar] [CrossRef]
- Kim, J.; Kil, E.J.; Kim, S.; Seo, H.; Byun, H.S.; Park, J.; Chung, M.N.; Kwak, H.R.; Kim, M.K.; Kim, C.S.; et al. Seed transmission of Sweet potato leaf curl virus in sweet potato (Ipomoea batatas). Plant Pathol. 2015, 64, 1284–1291. [Google Scholar] [CrossRef]
- Kil, E.J.; Kim, S.; Lee, Y.J.; Byuan, H.S.; Park, J.; Seo, H.; Kim, C.S.; Shim, J.K.; Lee, J.H.; Kim, J.K.; et al. Tomato yellow leaf curl virus (TYLCV-IL): A seed transmissible geminivirus in tomatoes. Sci. Rep. 2016, 6, 19013. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kil, E.J.; Park, J.; Choi, H.S.; Kim, C.S.; Lee, S. Seed Transmission of Tomato yellow leaf curl virus in White Soybean (Glycine max). Plant Pathol. J. 2017, 33, 424–428. [Google Scholar] [CrossRef] [Green Version]
- Sangeetha, B.; Malathi, V.G.; Alice, D.; Suganthy, M.; Renukadevi, P. A distinct seed-transmissible strain of Tomato leaf curl New Delhi virus infecting Chayote in India. Virus Res. 2018, 258, 81–91. [Google Scholar] [CrossRef] [PubMed]
- Kothandaraman, S.V.; Alice, D.; Malathi, V.G. Seed-borne nature of a begomovirus, Mung bean yellow mosaic virus in black gram. Appl. Microbiol. Biotechnol. 2015, 100, 1925–1933. [Google Scholar] [CrossRef] [PubMed]
- Manivannan, K.; Renukadevi, P.; Malathi, V.G.; Karthikeyan, G.; Balakrishnan, N. A new seed-transmissible begomovirus in bitter gourd (Momordica charantia L.). Microb. Pathog. 2019, 128, 82–89. [Google Scholar] [CrossRef]
- Fadhila, C.; Lal, A.; Vo, T.; Ho, P.T.; Hidayat, S.H.; Lee, J.; Kil, E.J.; Lee, S. The threat of seed-transmissible pepper yellow leaf curl Indonesia virus in chili pepper. Microb. Pathog. 2020, 143, 104–132. [Google Scholar] [CrossRef]
- Anabestani, A.; Behjatnia, S.A.A.; Izadpanah, K.; Tabein, S.; Accotto, G.P. Seed Transmission of Beet Curly Top Virus and Beet Curly Top Iran Virus in a Local Cultivar of Petunia in Iran. Viruses 2017, 9, 299. [Google Scholar] [CrossRef]
- Rosas-Diaz, T.; Zhang, D.; Lozano-Duran, R. No evidence of seed transmissibility of Tomato yellow leaf curl virus in Nicotiana benthamiana. J. Zhejiang Univ. Sci. B 2017, 18, 437–440. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rajabu, C.A.; Kennedy, G.G.; Ndunguru, J.; Ateka, E.M.; Tairo, F.; Hanley-Bowdoin, L.; Ascencio-Ibáñez, T. Lanai: A small, fast growing tomato variety is an excellent model system for studying geminiviruses. J. Virol. Methods 2018, 256, 89–99. [Google Scholar] [CrossRef]
- Pérez-Padilla, V.; Fortes, I.M.; Romero-Rodríguez, B.; Arroyo-Mateos, M.; Castillo, A.G.; Moyano, C.; De León, L.; Moriones, E. Revisiting Seed Transmission of the Type Strain of Tomato yellow leaf curl virus in Tomato Plants. Phytopathology 2020, 110, 121–129. [Google Scholar] [CrossRef] [PubMed]
- Andreason, S.A.; Olaniyi, O.G.; Gilliard, A.C.; Wadl, P.A.; Williams, L.H.; Jackson, D.M.; Simmons, A.M.; Ling, K.S. Large-Scale Seedling Grow-Out Experiments Do Not Support Seed Transmission of Sweet Potato Leaf Curl Virus in Sweet Potato. Plants 2021, 10, 139. [Google Scholar] [CrossRef] [PubMed]
- Accotto, G.P.; Bragaloni, M.; Luison, D.; Davino, S.; Davino, M. First report of Tomato yellow leaf curl virus (TYLCV) in Italy. Plant Pathol. 2003, 52, 799. [Google Scholar] [CrossRef]
- Moriones, E.; Navas-Castillo, J. Tomato yellow leaf curl virus, an emerging virus complex causing epidemics worldwide. Virus Res. 2000, 71, 123–134. [Google Scholar] [CrossRef]
- Kheyr-Pour, A.; Bendahmane, M.; Matzeit, V.; Accotto, G.P.; Crespi, S.; Gronenborn, B. Tomato yellow leaf curl virus from Sardinia is a whitefly-transmitted monopartite geminivirus. Nucleic Acids Res. 1991, 19, 6763–6769. [Google Scholar] [CrossRef] [Green Version]
- Mason, G.; Caciagli, P.; Accotto, G.P.; Noris, E. Real-time PCR for the quantitation of Tomato yellow leaf curl Sardinia virus in tomato plants and in Bemisia tabaci. J. Virol. Methods 2008, 147, 282–289. [Google Scholar] [CrossRef] [PubMed]
- Accotto, G.P.; Navas-Castillo, J.; Noris, E.; Moriones, E.; Louro, D. Typing of Tomato Yellow Leaf Curl Viruses in Europe. Eur. J. Plant Pathol. 2000, 106, 179–186. [Google Scholar] [CrossRef]
- Maffei, G.; Miozzi, L.; Fiorilli, V.; Novero, M.; Lanfranco, L.; Accotto, G.P. The arbuscular mycorrhizal symbiosis attenuates symptom severity and reduces virus concentration in tomato infected by Tomato yellow leaf curl Sardinia virus (TYLCSV). Mycorrhiza 2014, 24, 179–186. [Google Scholar] [CrossRef]
- Tabein, S.; Behjatnia, S.A.A.; Laviano, L.; Pecchioni, N.; Accotto, G.P.; Noris, E.; Miozzi, L. Pyramiding Ty-1/Ty-3 and Ty-2 in tomato hybrids dramatically inhibits symptom expression and accumulation of tomato yellow leaf curl disease inducing viruses. Arch. Phytopathol. Pflanzenschutz 2017, 50, 213–227. [Google Scholar] [CrossRef]
- Mabvakure, B.; Martin, D.P.; Karberger, S.; Cloete, L.; Van Brunschot, S.; Geering, A.D.; Thomas, J.E.; Bananej, K.; Lett, J.M.; Lefeuvre, P.; et al. Ongoing geographical spread of Tomato yellow leaf curl virus. Virology 2016, 498, 257–264. [Google Scholar] [CrossRef] [PubMed]
- Yan, Z.; Wolters, A.-M.A.; Navas-Castillo, J.; Bai, Y. The Global Dimension of Tomato Yellow Leaf Curl Disease: Current Status and Breeding Perspectives. Microorganisms 2021, 9, 740. [Google Scholar] [CrossRef] [PubMed]
- Navas-Castillo, J.; Sánchez-Campos, S.; Díaz, J.A.; Sáez-Alonso, E.; Moriones, E. Tomato yellow leaf curl virus-Is causes a novel disease of common bean and severe epidemics in tomato in Spain. Plant Dis. 1999, 83, 29–32. [Google Scholar] [CrossRef] [Green Version]
- Sánchez-Campos, S.; Navas-Castillo, J.; Camero, R.; Soria, C.; Díaz, J.A.; Moriones, E. Displacement of Tomato Yellow Leaf Curl Virus (TYLCV)-Sr by TYLCV-Is in Tomato Epidemics in Spain. Phytopathology 1999, 89, 1038–1043. [Google Scholar] [CrossRef] [Green Version]
- Sánchez-Campos, S.; Navas-Castillo, J.; Monci, F.; Díaz, J.A.; Moriones, E. Mercurialis ambigua and Solanum luteum: Two Newly Discovered Natural Hosts of Tomato Yellow Leaf Curl Geminiviruses. Eur. J. Plant Pathol. 2000, 106, 391–394. [Google Scholar] [CrossRef]
- Barbieri, M.; Acciarri, N.; Sabatini, E.; Sardo, L.; Accotto, G.P.; Pecchioni, N. Introgression of resistance to two Mediterranean virus species causing tomato yellow leaf curl into a valuable traditional tomato variety. J. Plant. Pathol. 2014, 92, 485–493. [Google Scholar]
- Shen, X.; Yan, Z.; Wang, X.; Wang, Y.; Arens, M.; Du, Y.; Visser, R.; Kormelink, R.; Bai, Y.; Wolters, A.A. The NLR Protein Encoded by the Resistance Gene Ty-2 Is Triggered by the Replication-Associated Protein Rep/C1 of Tomato Yellow Leaf Curl Virus. Front. Plant Sci. 2020, 11, 545306. [Google Scholar] [CrossRef] [PubMed]
- Luna, A.P.; Morilla, G.; Voinnet, O.; Bejarano, E.R. Functional analysis of gene-silencing suppressors from tomato yellow leaf curl disease viruses. Mol. Plant. Microbe Interact. 2012, 25, 1294–1306. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Matić, S.; Pegoraro, M.; Noris, E. The C2 protein of tomato yellow leaf curl Sardinia virus acts as a pathogenicity determinant and a 16-amino acid domain is responsible for inducing a hypersensitive response in plants. Virus Res. 2016, 215, 12–19. [Google Scholar] [CrossRef] [PubMed]
- Morilla, G.; Krenz, B.; Jeske, H.; Bejarano, E.R.; Wege, C. Tête à tête of tomato yellow leaf curl virus and tomato yellow leaf curl Sardinia virus in single nuclei. J. Virol. 2004, 78, 10715–10723. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sardo, L.; Wege, C.; Kober, S.; Kocher, C.; Accotto, G.P.; Noris, E. RNA viruses and their silencing suppressors boost Abutilon mosaic virus, but not the Old World Tomato yellow leaf curl Sardinia virus. Virus Res. 2011, 161, 170–180. [Google Scholar] [CrossRef] [PubMed]
- Kil, E.-J.; Park, J.; Choi, E.-Y.; Byun, H.-S.; Lee, K.-Y.; An, C.G.; Lee, J.-H.; Lee, G.-S.; Choi, H.-S.; Kim, C.-S.; et al. Seed transmission of Tomato yellow leaf curl virus in sweet pepper (Capsicum annuum). Eur. J. Plant. Pathol. 2017, 150, 759–764. [Google Scholar] [CrossRef]
- Just, K.; Arif, U.; Luik, A.; Kvarnheden, A. Monitoring infection of tomato fruit by Tomato yellow leaf curl virus. Plant Pathol. 2017, 66, 522–528. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Primers | Sequence (5′ to 3′) | Size of Amplicon (bp) | Reference |
|---|---|---|---|
| TY1(+) | GCCCATGTA(T/C)CG(A/G)AAGCC | 577 | [20] |
| TY2(−) | GG(A/G)TTAGA(A/G)GCATG(A/C)GTAC | ||
| TY2458(+) | CATTTTCATGTAGTTCTCTG | 424 | This manuscript |
| TY109(−) | CACCAGCTGAACAGTTATTTAA | ||
| TY2222(+) | GTCGTTGGCTGTCTGTTGTC | 150 | [21] |
| TY2371(−) | AGGTCAGCACATTTCCATCC | ||
| SlyAPX-862(+) | CCCCTTTTGGCTTAATACTCG | 87 | [21] |
| SlyAPX-948(−) | GCAGAAATGGAAATGCGATAA |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tabein, S.; Miozzi, L.; Matić, S.; Accotto, G.P.; Noris, E. No Evidence for Seed Transmission of Tomato Yellow Leaf Curl Sardinia Virus in Tomato. Cells 2021, 10, 1673. https://doi.org/10.3390/cells10071673
Tabein S, Miozzi L, Matić S, Accotto GP, Noris E. No Evidence for Seed Transmission of Tomato Yellow Leaf Curl Sardinia Virus in Tomato. Cells. 2021; 10(7):1673. https://doi.org/10.3390/cells10071673
Chicago/Turabian StyleTabein, Saeid, Laura Miozzi, Slavica Matić, Gian Paolo Accotto, and Emanuela Noris. 2021. "No Evidence for Seed Transmission of Tomato Yellow Leaf Curl Sardinia Virus in Tomato" Cells 10, no. 7: 1673. https://doi.org/10.3390/cells10071673