
Hypoxia Inducible Factor-1α Attenuates Ischemic Brain Damage by Modulating Inflammatory Response and Glial Activity

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
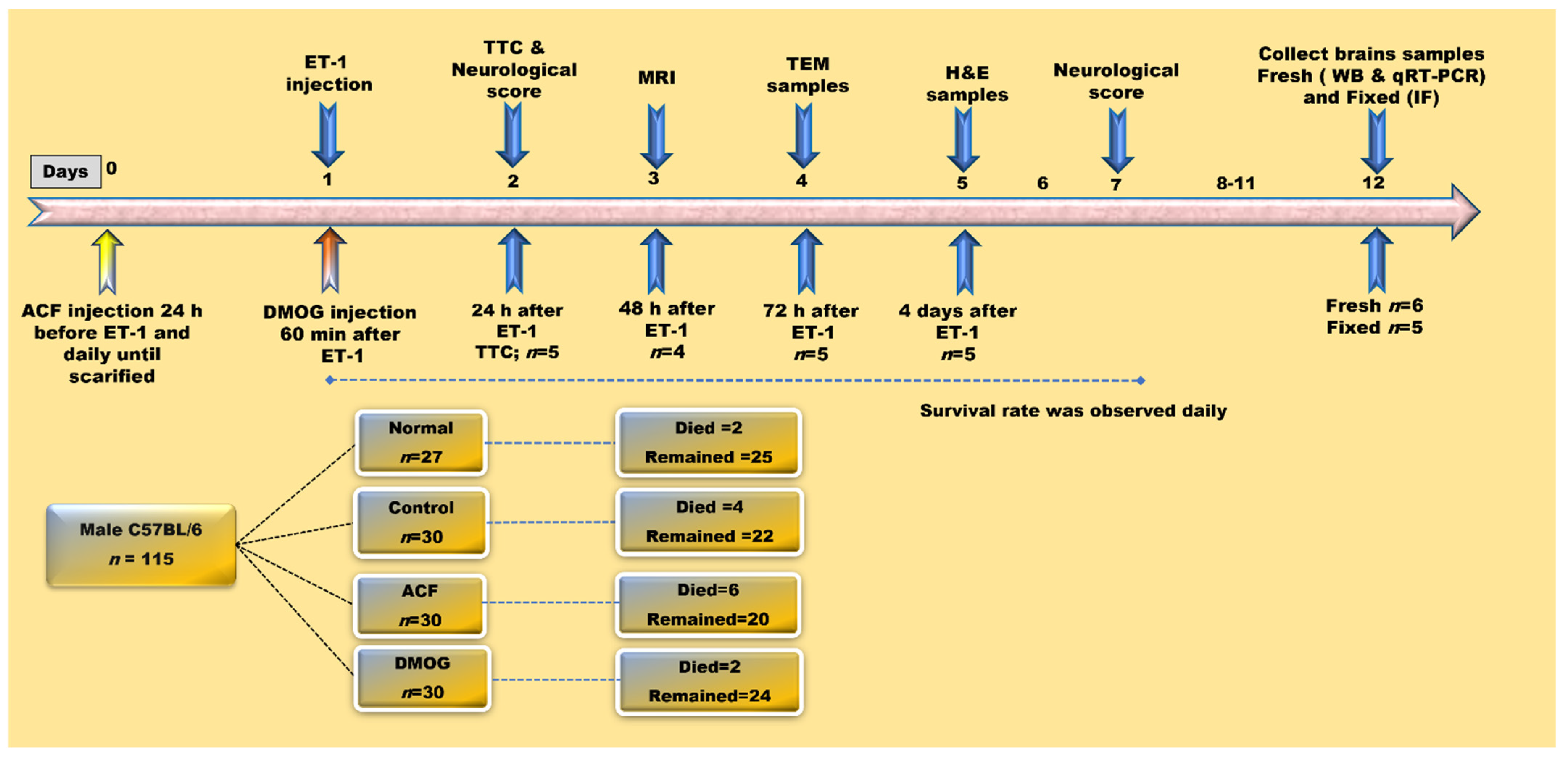
2.1. Animal Classification and Drug Administration
2.2. Induction of Focal Cerebral Ischemia by Endothelin-1
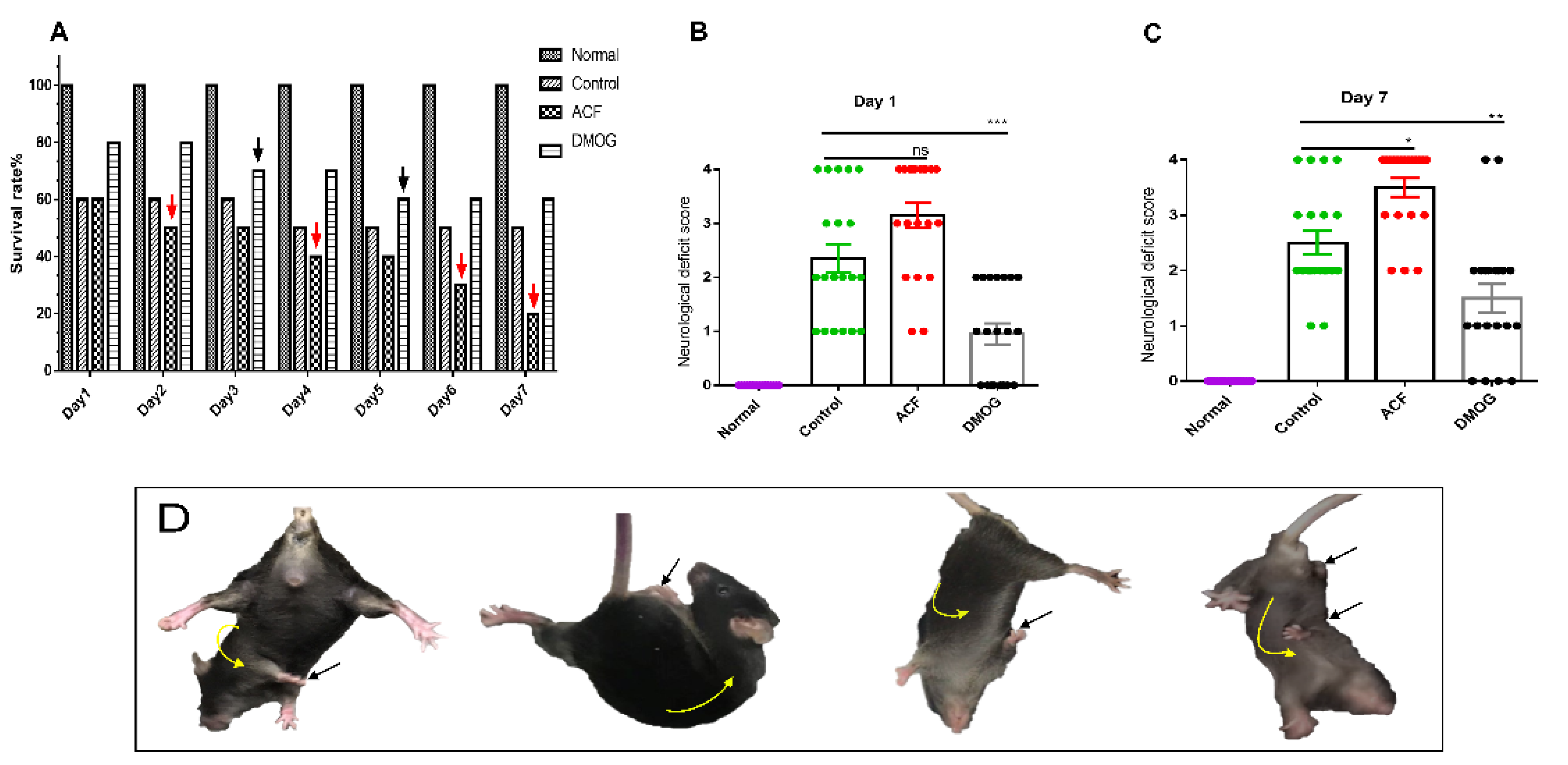
2.3. Neurological Deficit Score and Survival Rate
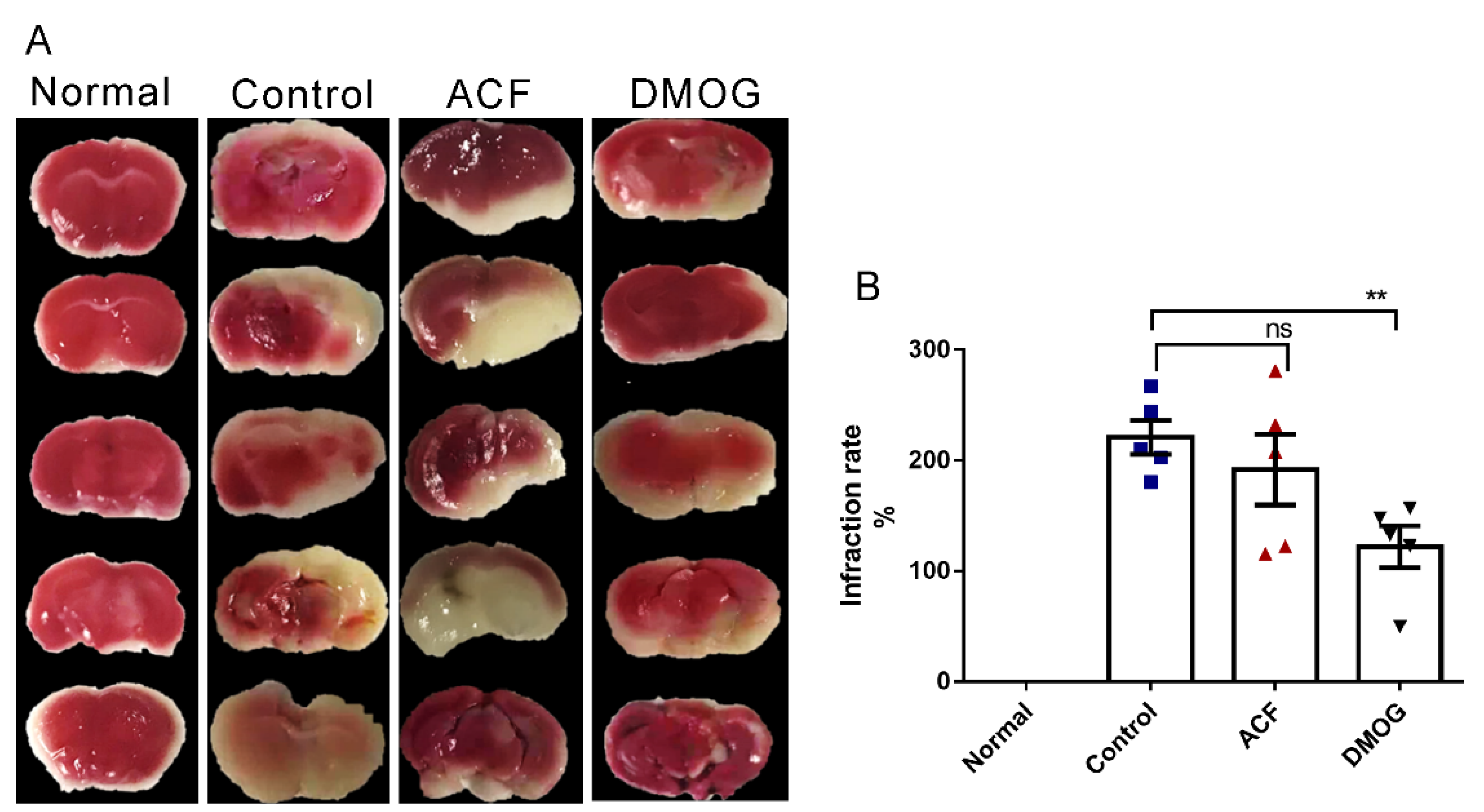
2.4. 2,3,5-. Triphenyl Tetrazolium Chloride [TTC] Staining
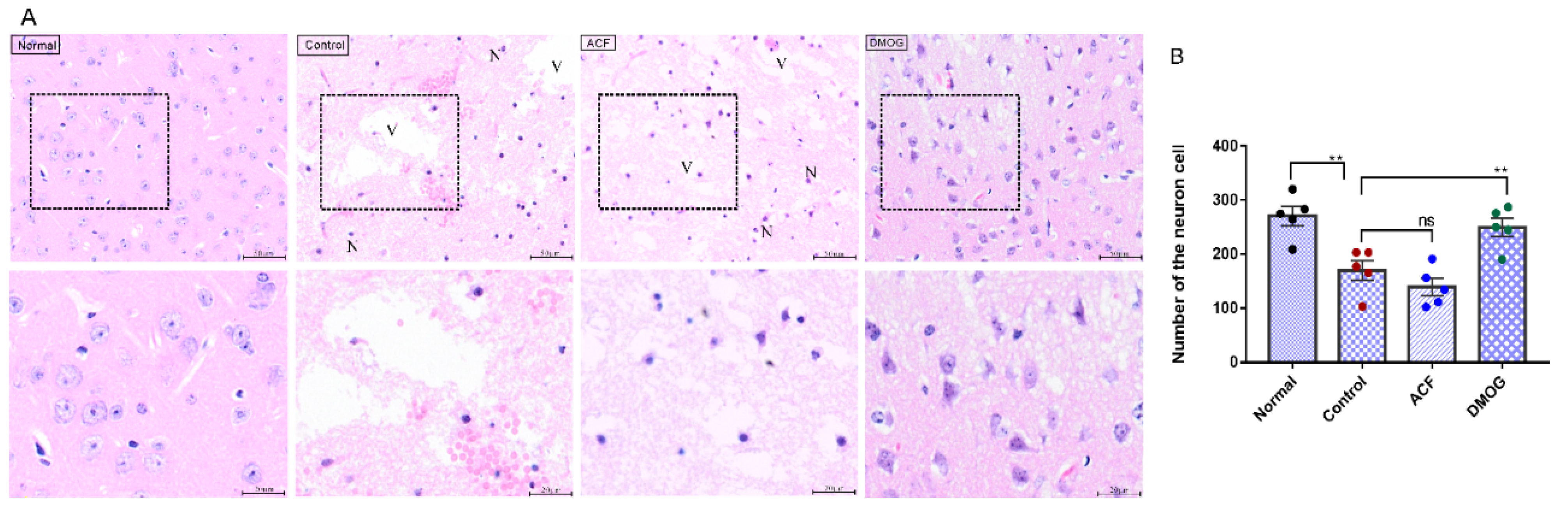
2.5. Hematoxylin-Eosin [HE]
2.6. Western Blotting
2.7. Immunofluorescence
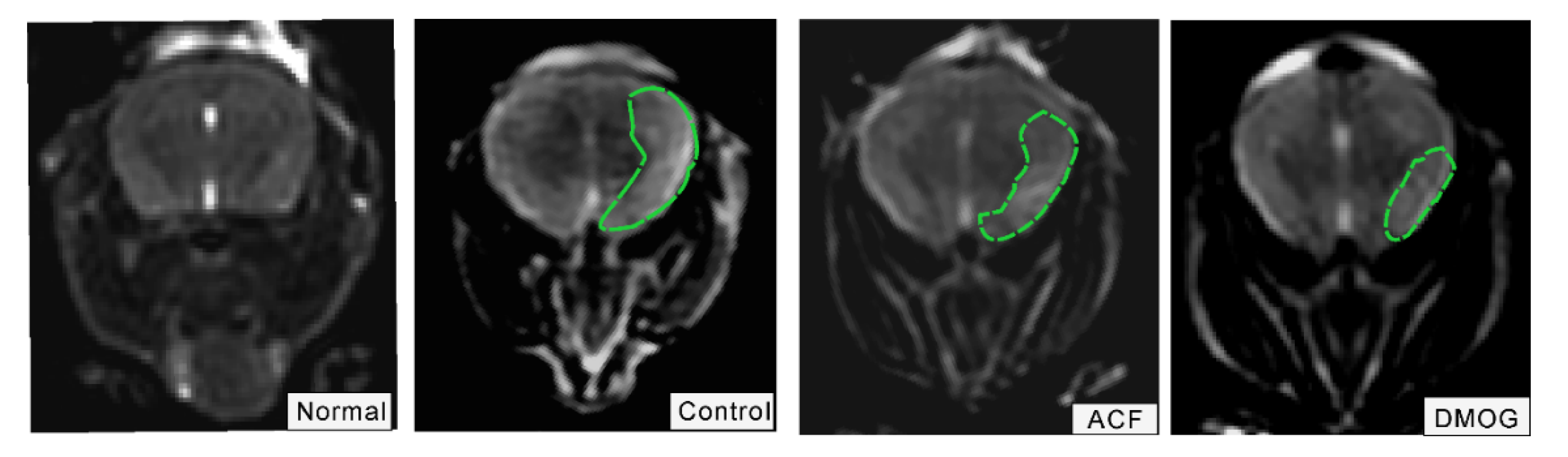
2.8. T2-Weighted MRI
2.9. Transmission Electron Microscopy
2.10. Real-Time PCR
2.11. Statistical Analysis
3. Results
3.1. HIF-1α Improved the Neurological Deficit Score
3.2. HIF-1 Reduces the Infarct Area Volume Resulting from ET-1 Injection
3.3. HIF-1α Activation Recovered the Brain Damage Following ET-1 Injection
3.4. HIF-1α Activation Possesses a Neuroprotective Effect Post-Stroke
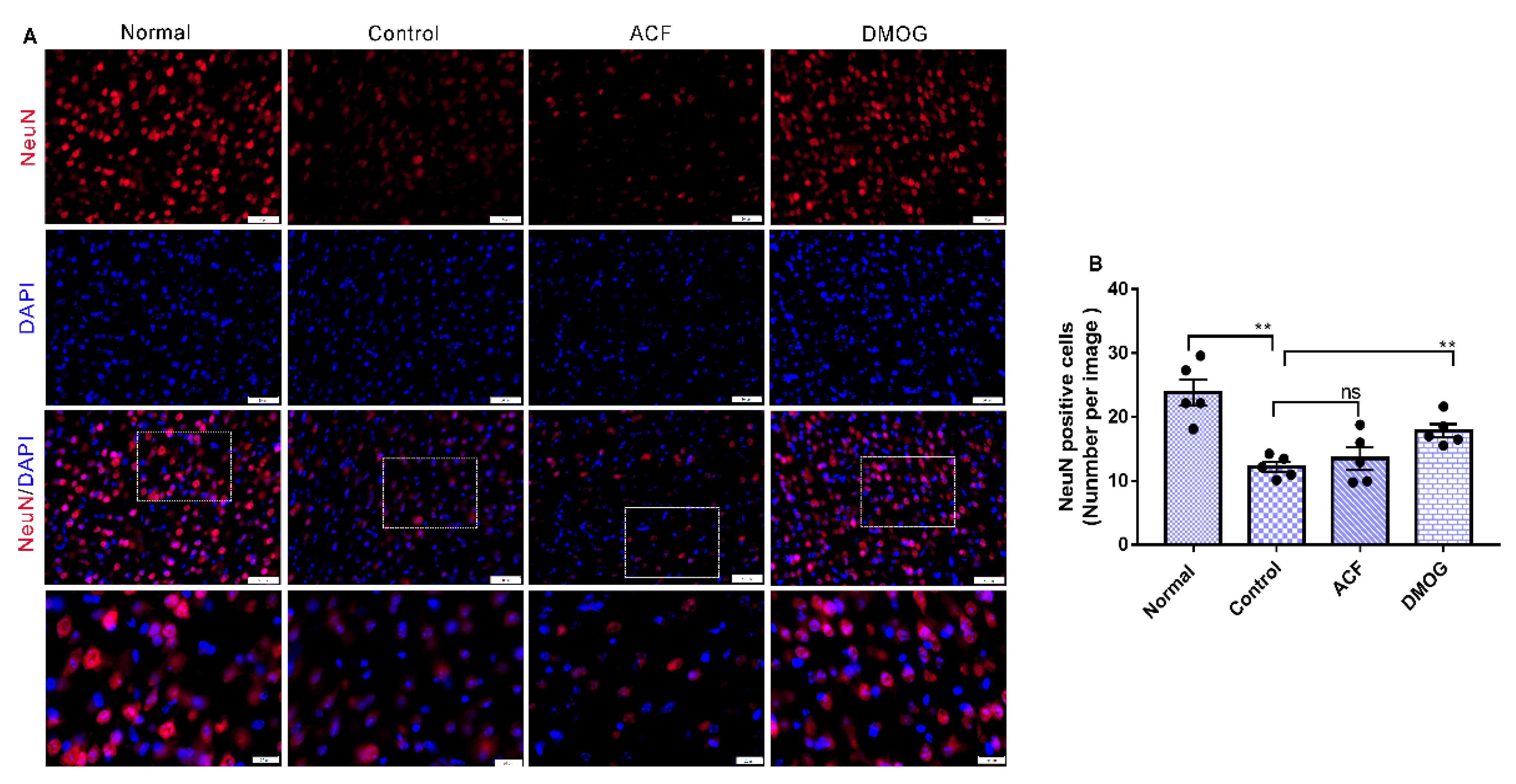
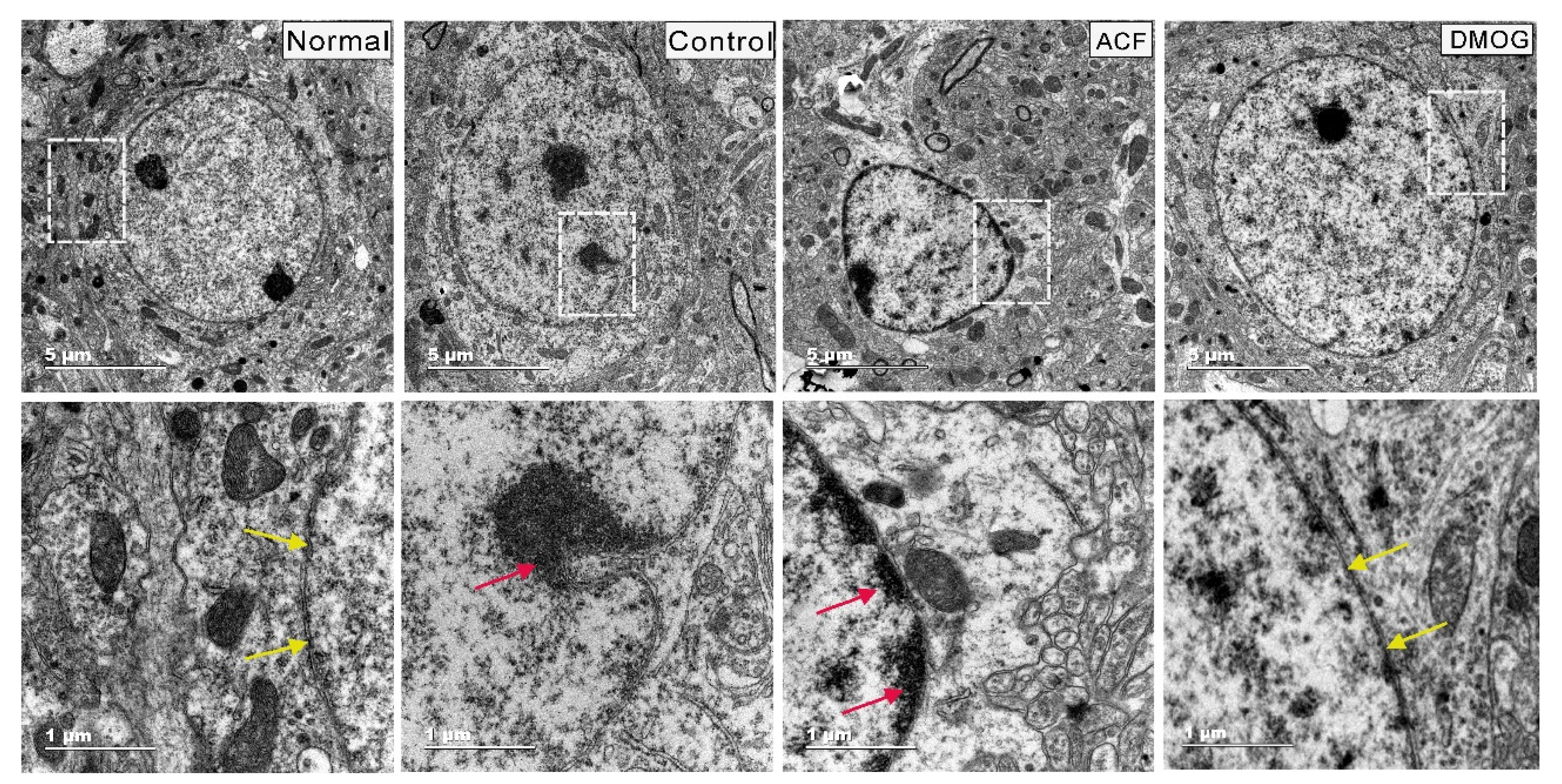
3.5. HIF-1α Activation Can Modulate Brain Damage via Neuronal Soma Protection
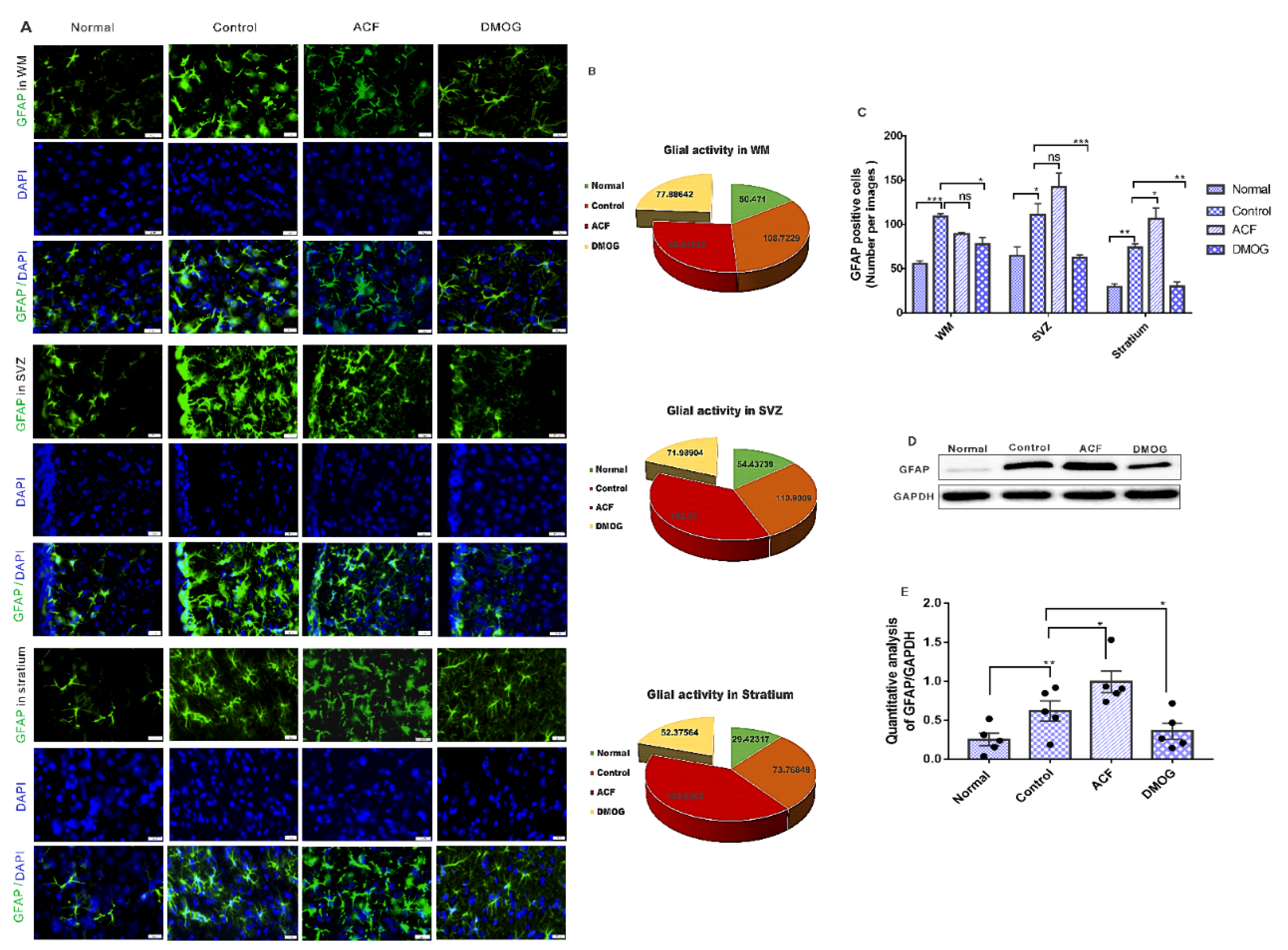
3.6. HIF-1α Reduces the Glial Activity Following ET-1 Injection
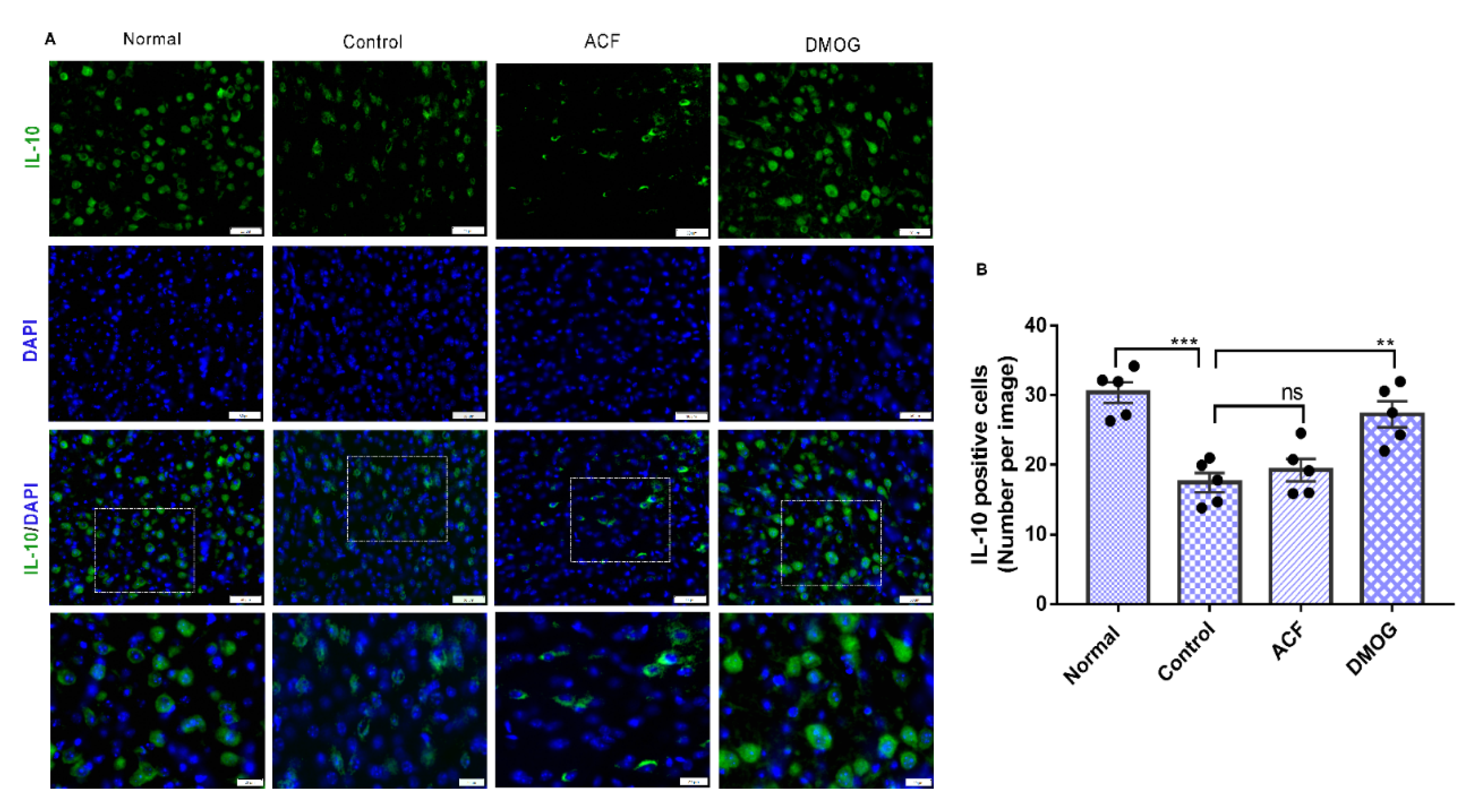
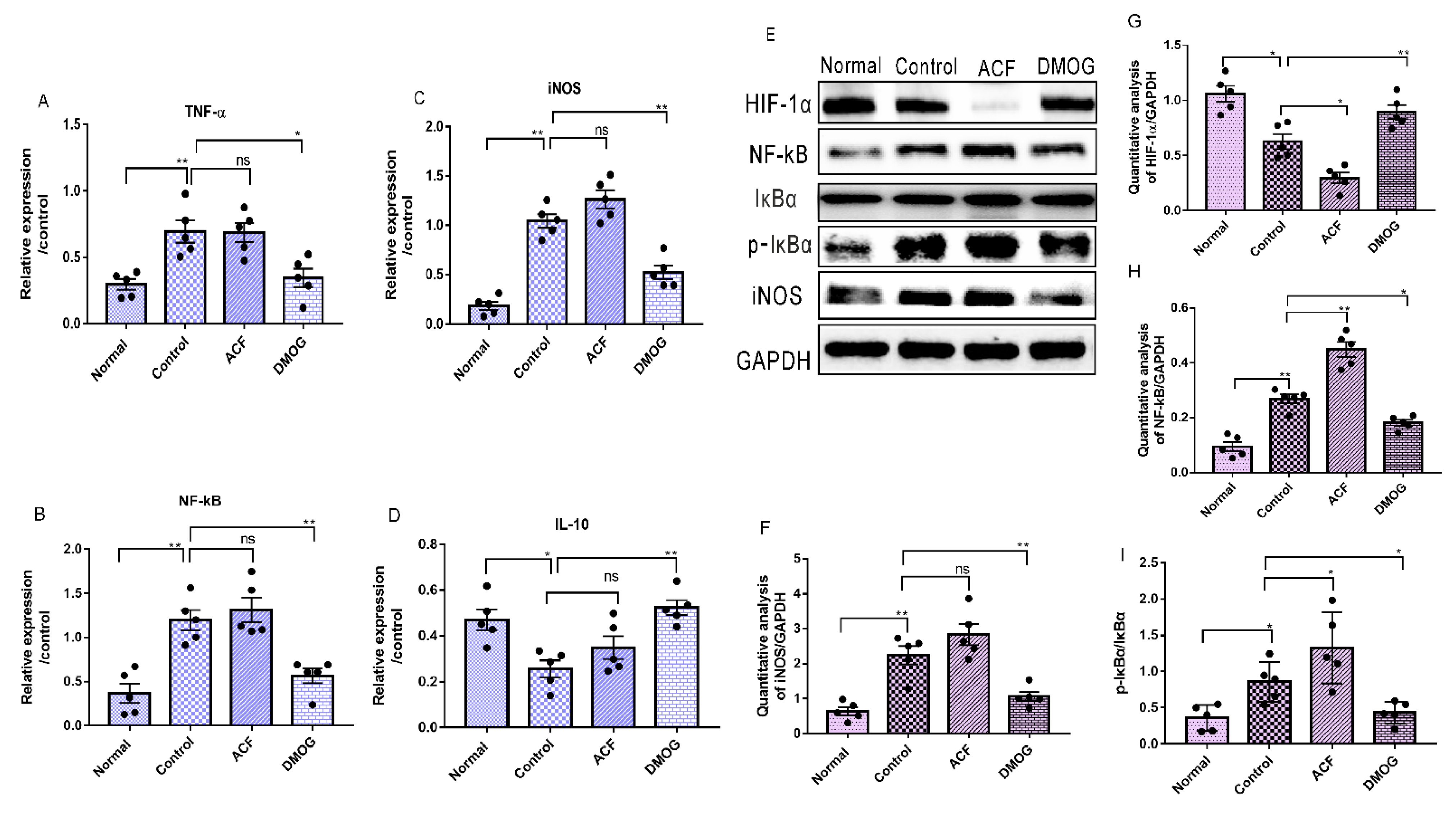
3.7. HIF-1α Activation Down-Regulates the Pro-Inflammatory Cytokines iNOS, NF-kB, and Upregulates the Anti-Inflammatory Marker IL-10
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Panel; Mohr, J.P.; Albers, G.W.; Amarenco, P.; Babikian, V.L.; Biller, J.; Brey, R.L.; Coull, B.; Easton, J.D.; Gomez, C.R.; et al. Etiology of stroke. Stroke 1997, 28, 1501–1506. [Google Scholar] [CrossRef]
- Marti, H.J.; Bernaudin, M.; Bellail, A.; Schoch, H.; Euler, M.; Petit, E.; Risau, W. Hypoxia-Induced Vascular Endothelial Growth Factor Expression Precedes Neovascularization after Cerebral Ischemia. Am. J. Pathol. 2000, 156, 965–976. [Google Scholar] [CrossRef] [Green Version]
- Shi, H. Hypoxia inducible factor 1 as a therapeutic target in ischemic stroke. Curr. Med. Chem. 2009, 16, 4593–4600. [Google Scholar] [CrossRef] [Green Version]
- Bernaudin, M.; Nedelec, A.S.; Divoux, D.; MacKenzie, E.T.; Petit, E.; Schumann-Bard, P. Normobaric hypoxia induces tolerance to focal permanent cerebral ischemia in association with an increased expression of hypoxia-inducible factor-1 and its target genes, erythropoietin, and VEGF, in the adult mouse brain. J. Cereb. Blood Flow Metab. 2002, 4, 393–403. [Google Scholar] [CrossRef] [Green Version]
- Jones, N.M.; Bergeron, M. Hypoxic preconditioning induces changes in HIF-1A target genes in neonatal rat brain. J. Cereb. Blood Flow Metab. 2001, 9, 1105–1114. [Google Scholar] [CrossRef] [Green Version]
- Bergeron, M.; Yu, A.Y.; Solway, K.E.; Semenza, G.L.; Sharp, F.R. Induction of hypoxia-inducible factor-1[HIF-1A] and its target genes following focal ischemia in rat brain. Eur. J. Neurosci. 1999, 12, 4159–4170. [Google Scholar] [CrossRef]
- Baranova, O.; Miranda, L.F.; Pichiule, P.; Dragatsis, I.; Johnson, R.S.; Chavez, J.C. Neuron-specific inactivation of the hypoxia inducible factor 1α increases brain injury in a mouse model of transient focal cerebral ischemia. J. Neurosci. 2007, 23, 6320–6332. [Google Scholar] [CrossRef] [PubMed]
- Sharp, F.R.; Bernaudin, M. HIF-I and oxygen sensing in the brain. Nat. Rev. Neurosci. 2004, 6, 437–448. [Google Scholar] [CrossRef]
- Pichiule, P.; Agani, F.; Chavez, J.C.; Xu, K.; LaManna, J.C. HIF-1Aα and VEGF expression after transient global cerebral ischemia. Adv. Exp. Med. Biol. 2003, 530, 611–617. [Google Scholar]
- Herx, L.M.; Yong, V.W. Interleukin-1 beta is required for the early evolution of reactive astrogliosis following CNS lesion. J. Neuropathol. Exp. Neurol. 2001, 60, 961–971. [Google Scholar] [CrossRef] [Green Version]
- Brunello, A.G.; Weissenberger, J.; Kappeler, A.; Vallan, C.; Peters, M.; Rose, J.S.; Weis, J. Astrocytic alterations in interleukin-6/Soluble interleukin-6 receptor alpha double-transgenic mice. Am. J. Pathol. 2000, 157, 1485–1493. [Google Scholar] [CrossRef]
- Scarisbrick, I.A.; Radulovic, M.; Burda, J.E. Kallikrein 6 is a novel molecular trigger of reactive astrogliosis. Biol. Chem. 2012, 393, 355–367. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Choudhury, G.R.; Ding, S. Reactive astrocytes and therapeutic potential in focal ischemic stroke. Neurobiol. Dis. 2016, 85, 234–244. [Google Scholar] [CrossRef] [Green Version]
- Asano, T.; Mori, T.; Shimoda, T.; Shinagawa, R.; Satoh, S.; Yada, N.; Katsumata, S.; Matsuda, S.; Kagamiishi, Y.; Tateishi, N. Arundic acid [ONO-2506] ameliorates delayed ischemic brain damage by preventing astrocytic overproduction of S100B. Curr. Drug Targets CNS Neurol. Disord. 2005, 4, 127–142. [Google Scholar] [CrossRef]
- Sofroniew, M.V. Reactive astrocytes in neural repair and protection. Neuroscientist 2005, 11, 400–407. [Google Scholar] [CrossRef]
- Buffo, A.; Rolando, C.; Ceruti, S. Astrocytes in the Damaged brain: Molecular and cellular insights into their reactive response and healing potential. Biochem. Pharmacol. 2010, 79, 77–89. [Google Scholar] [CrossRef] [Green Version]
- Stoll, G.; Jander, S.; Schroeter, M. Inflammation and glial responses in ischemic brain lesions. Prog. Neurobiol. 1998, 56, 149–171. [Google Scholar] [CrossRef]
- Sukumari-Ramesh, S.; Alleyne, C.H.; Dhandapani, K.M. Astrocyte-specific expression of survivin after intracerebral hemorrhage in mice: A possible role in reactive gliosis? J. Neurotrauma 2012, 29, 2798–2804. [Google Scholar] [CrossRef]
- Bezzi, P.; Domercq, M.; Brambilla, L.; Galli, R.; Schols, D.; De Clercq, E.; Vescovi, A.; Bagetta, G.; Kollias, G.; Meldolesi, J.; et al. CXCR4-activated astrocyte glutamate release via TNF_: Amplification by microglia triggers neurotoxicity. Nat. Neurosci. 2001, 4, 702–710. [Google Scholar] [CrossRef]
- Wang, Q.; Tang, X.N.; Yenari, M.A. The inflammatory response in stroke. J. Neuroimmunol. 2007, 184, 53–68. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fuxe, K.; Cintra, A.; Andbjer, B.; Anggard, E.; Goldstein, M.; Agnati, L.F. Centrally administered endothelin-1 produces lesions in the brain of the male rat. Acta Physiol. Scand. 1989, 137, 155–156. [Google Scholar] [CrossRef]
- Agnati, L.F.; Zoli, M.; Kurosawa, M.; Benfenati, F.; Biagini, G.; Zini, I.; Hallstròm, A.; Ungerstedt, U.; Toffano, G.; Fuxe, K. A new model of focal brain ischemia based on the intracerebral injection of endothelin 1. Ital. J. Neurol. Sci. 1991, 12, 49–53. [Google Scholar]
- Hughes, P.M.; Anthony, D.C.; Ruddin, M.; Botham, M.S.; Rankine, E.L.; Sablone, M.; Baumann, D.; Mir, A.K.; Perry, V.H. Focal lesions in the rat central nervous system induced by endothelin 1. J. Neuropathol. Exp. Neurol. 2003, 62, 1276–1286. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Delavaran, H.; Sjunnesson, H.; Arvidsson, A.; Lindvall, O.; Norrving, B.; van Westen, D.; Kokaia, Z.; Lindgren, A. Proximity of brain infarcts to regions of endogenous neurogenesis and involvement of striatum in ischemic stroke. Eur. J. Neurol. 2013, 20, 473–479. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Q.; Memezawa, H.; Smith, M.-L.; Siej¨o, B.K. Hyperthermia complicates middle cerebral artery occlusion induced by an intraluminal filament. Brain Res. 1994, 649, 253–259. [Google Scholar] [CrossRef]
- Souza-Rodrigues, R.D.; Costa, A.M.R.; Lima, R.R.; Dos, S.C.D.; Picanc, D.C.W.; Gomes-Leal, W. Inflammatory response and white matter damage after microinjections of endothelin-1 into the rat striatum. Brain Res. 2008, 1200, 78–88. [Google Scholar] [CrossRef]
- Dos Santos, C.D.; Picanc, D.C.W.; Gomes-Leal, W. Differential patterns of inflammatory response, axonal damage and myelin impairment following excitotoxic or ischemic damage to the trigeminal spinal nucleus of adult rats. Brain Res. 2007, 1172, 130–144. [Google Scholar] [CrossRef] [PubMed]
- Franco, E.C.S.; Cardoso, M.M.; Gouveia, A.; Pereira, A.; Gomes-Leal, W. Modulation of microglial activation enhances neuroprotection and functional recovery derived from bone marrow mononuclear cell transplantation after cortical ischemia. Neurosci. Res. 2012, 73, 122–132. [Google Scholar] [CrossRef]
- Jian, Y.; Cuiying, L.; Xiangnan, D.; Menglei, L.; Xunming, J.; Huishan, D.; Heng, Z. Hypoxia Inducible Factor 1a Plays a Key Role in Remote Ischemic Preconditioning Against Stroke by Modulating Inflammatory Responses in Rats. J. Am. Heart Assoc. 2018, 7, e007589. [Google Scholar]
- Selvamani, A.; Sohrabji, F. Reproductive age modulates the impact of focal ischemia on the forebrain as well as the effects of estrogen treatment in female rats. Neurobiol. Aging 2010, 31, 1618–1628. [Google Scholar] [CrossRef] [Green Version]
- Sharkey, J.; Butcher, S.P. Characterization of an experimental model of stroke produced by intracerebral microinjection of endothelin-1 adjacent to the rat middle cerebral artery. J. Neurosci. Methods. 1995, 60, 125–131. [Google Scholar] [CrossRef]
- Xiaoxue, D.; Feng, G.; Shijia, C.; Benson, O.A.B.; Nashwa, A.; Zhiying, H.; Marong, F. Combinational Pretreatment of Colony-Stimulating Factor 1 Receptor Inhibitor and Triptolide Upregulates BDNF-Akt and Autophagic Pathways to Improve Cerebral Ischemia. Mediat. Inflamm. 2020, 2020, 8796103. [Google Scholar]
- Yan, J.; Zhou, B.; Taheri, S.; Shi, H. Differential effects of HIF-1A inhibition by YC-1 on the overall outcome and blood-brain barrier damage in a rat model of ischemic stroke. PLoS ONE 2011, 6, e27798. [Google Scholar] [CrossRef] [Green Version]
- Umschweif, G.; Alexandrovich, A.G.; Trembovler, V.; Horowitz, M.; Shohami, E. Hypoxia-inducible factor 1 is essential for spontaneous recovery from traumatic brain injury and is a key mediator of heat acclimation induced neuroprotection. J. Cereb. Blood Flow Metab. 2013, 33, 524–531. [Google Scholar] [CrossRef] [Green Version]
- Ogle, M.E.; Gu, X.; Espinera, A.R.; Wei, L. Inhibition of prolyl hydroxylases by dimethyloxalylglycine after stroke reduces ischemic brain injury and requires hypoxia inducible factor-1a. Neurobiol. Dis. 2012, 45, 733–742. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zahra, G.M.; Mohsen, R.P.; Mohammad, H.M.; Kamran, M. Cancer cells change their glucose metabolism to overcome increased ROS: One step from cancer cell to cancer stem cell? Biomed. Pharmacother. 2019, 112, 108690. [Google Scholar]
- Marxsen, J.H.; Stengel, P.; Doege, K.; Heikkinen, P.; Jokilehto, T.; Wagner, T.; Jelkmann, W.; Jaakkola, P.; Metzen, E. Hypoxia-inducible factor-1 [HIF-1A] promotes its degradation by induction of HIF-alpha-prolyl-4-hydroxylases. Biochem. J. 2004, 381, 761–767. [Google Scholar] [CrossRef] [PubMed]
- Zepeda, A.B.; Pessoa, A.; Castillo, R.L.; Figueroa, C.A.; Pulgar, V.M.; Farías, J.G. Cellular, and molecular mechanisms in the hypoxic tissue: Role of HIF-1A and ROS. Cell Biochem. Funct. 2013, 31, 451–459. [Google Scholar] [CrossRef] [PubMed]
- Alique, M.; Sánchez-López, E.; Bodega, G.; Giannarelli, C.; Carracedo, J.; Ramírez, R. Hypoxia-Inducible Factor-1α: The Master Regulator of Endothelial Cell Senescence in Vascular Aging. Cells 2020, 9, 195. [Google Scholar] [CrossRef] [Green Version]
- Ivanova, I.G.; Park, C.V.; Kenneth, N.S. Translating the Hypoxic Response—The Role of HIF Protein Translation in the Cellular Response to Low Oxygen. Cells 2019, 8, 114. [Google Scholar] [CrossRef] [Green Version]
- Wielockx, B.; Grinenko, T.; Mirtschink, P.; Chavakis, T. Hypoxia Pathway Proteins in Normal and Malignant Hematopoiesis. Cells 2019, 8, 155. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Salminen, A.; Kaarniranta, K.; Kauppinen, A. Hypoxia-Inducible Histone Lysine Demethylases: Impact on the Aging Process and Age-Related Diseases. Aging Dis. 2016, 7, 180–200. [Google Scholar]
- Li, X.; Liao, Y.; Dong, Y.; Li, S.; Wang, F.; Wu, R.; Yuan, Z.; Cheng, J. Mib2 Deficiency Inhibits Microglial Activation and Alleviates Ischemia-Induced Brain Injury. Aging Dis. 2020, 11, 523–535. [Google Scholar] [CrossRef] [PubMed]
- Li, K.; Li, J.; Zheng, J.; Qin, S. Reactive Astrocytes in Neurodegenerative Diseases. Aging Dis. 2019, 10, 664–675. [Google Scholar] [CrossRef] [PubMed] [Green Version]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Forward Primer | Reverse Primer |
|---|---|---|
| β-actin | 5′-CTGTCCCTGTATGCCTCTG-3′. | 5′-ATGTCACGCACGATTTCC-3′. |
| IL-10 | 5′-CGGGAAGACAATAACTGCACCC-3′ | 5′-CGGTTAGCAGTATGTTGTCCAGC-3′ |
| NF-kB | 5′-GCTGCCAAAGAAGGACACGACA-3′ | 5′-GGCAGGCTATTGCTCATCACAG-3′ |
| iNOS | 5′-CACCTTGGAGTTCACCCAGT-3′ | 5′-ACCACTCGTACTTGGGATGC-3′ |
| TNF-ἀ | 5′-GGTGCCTATGTCTCAGCCTCTT-3′ | 5′-GCCATAGAACTGATGAGAGGGAG-3′ |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Amin, N.; Chen, S.; Ren, Q.; Tan, X.; Botchway, B.O.A.; Hu, Z.; Chen, F.; Ye, S.; Du, X.; Chen, Z.; et al. Hypoxia Inducible Factor-1α Attenuates Ischemic Brain Damage by Modulating Inflammatory Response and Glial Activity. Cells 2021, 10, 1359. https://doi.org/10.3390/cells10061359
Amin N, Chen S, Ren Q, Tan X, Botchway BOA, Hu Z, Chen F, Ye S, Du X, Chen Z, et al. Hypoxia Inducible Factor-1α Attenuates Ischemic Brain Damage by Modulating Inflammatory Response and Glial Activity. Cells. 2021; 10(6):1359. https://doi.org/10.3390/cells10061359
Chicago/Turabian StyleAmin, Nashwa, Shijia Chen, Qiannan Ren, Xiaoning Tan, Benson O. A. Botchway, Zhiying Hu, Fengpei Chen, Shan Ye, Xiaoxue Du, Zuobing Chen, and et al. 2021. "Hypoxia Inducible Factor-1α Attenuates Ischemic Brain Damage by Modulating Inflammatory Response and Glial Activity" Cells 10, no. 6: 1359. https://doi.org/10.3390/cells10061359