
Utilizing Developmentally Essential Secreted Peptides Such as Thymosin Beta-4 to Remind the Adult Organs of Their Embryonic State—New Directions in Anti-Aging Regenerative Therapies

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Hallmarks of Aging
3. Pathological Alterations and Recent Therapeutic Approaches to Aid the Adult Heart Following Hypoxia
4. Beta Thymosins
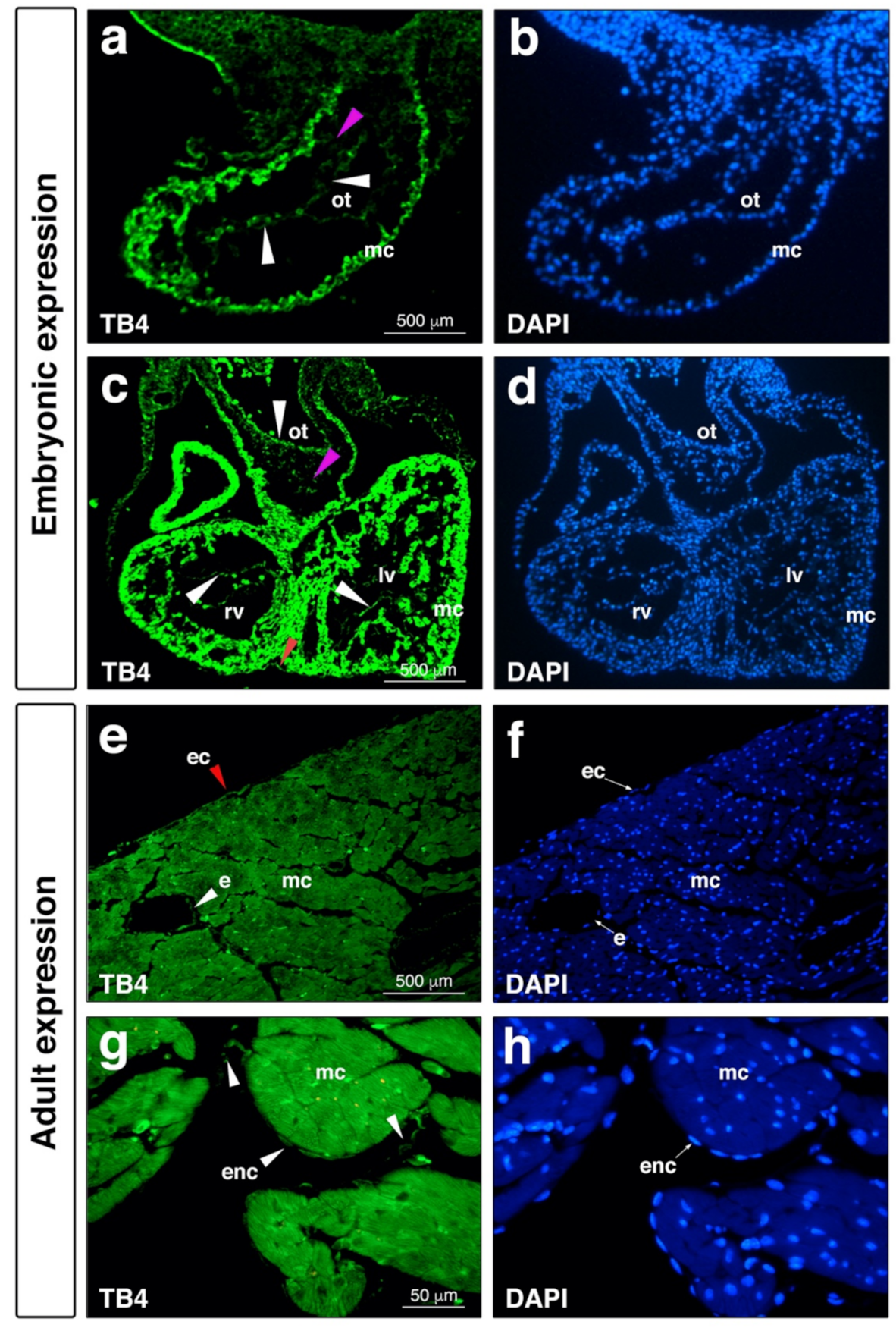
5. The Expression of Thymosin Beta-4 during Embryonic Development
6. Thymosin Beta-4 Holds Significant Potential to Initiate Organ Regeneration and Repair by Influencing Multiple Senescence Related Biological Platforms in Adult Mammals
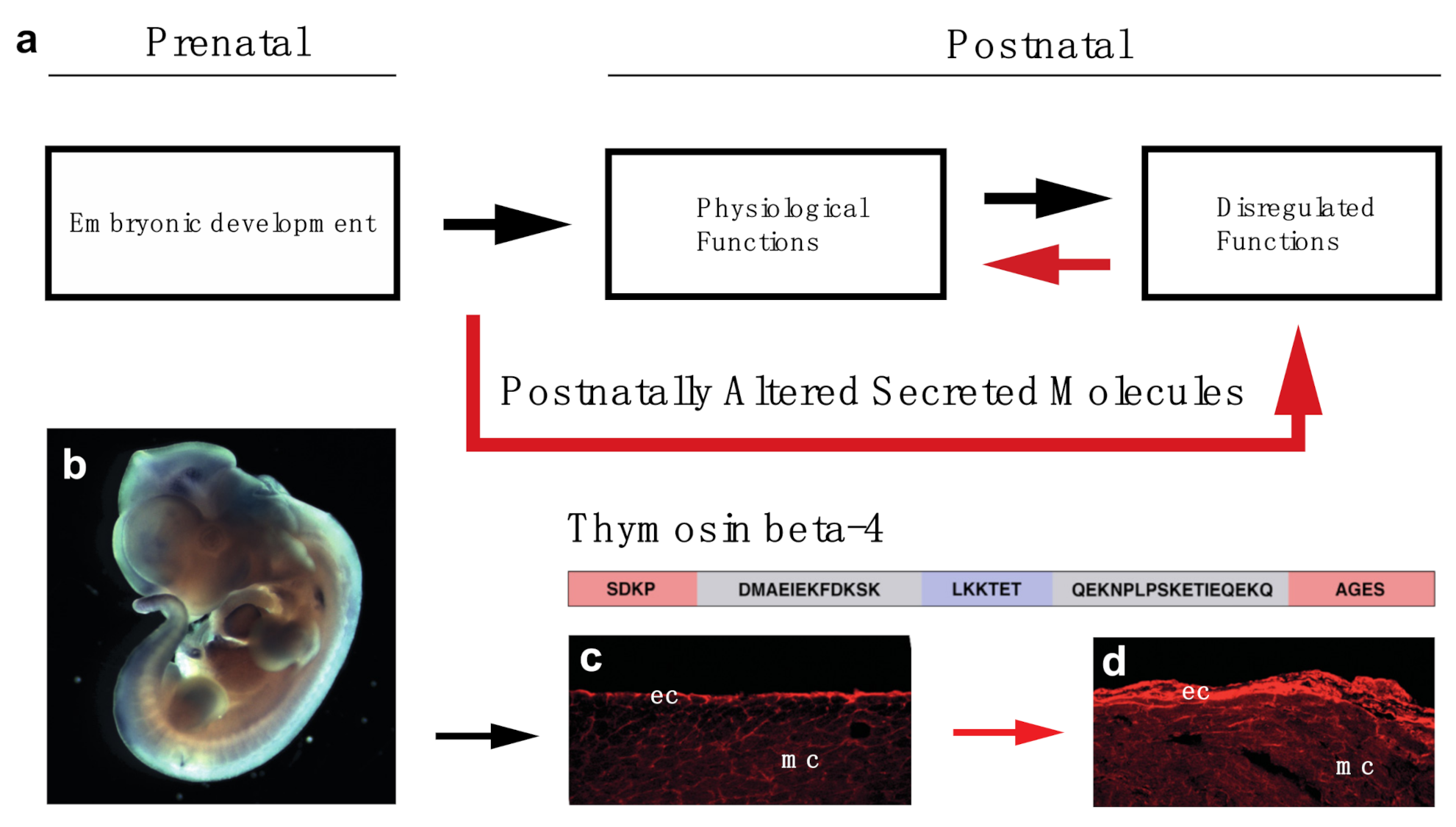
7. Activating the Embryonic Developmental Program May Be the Key for Thymosin Beta-4′s Regenerative Potential in the Heart
8. Future Perspectives
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- López-Otín, C.; Blasco, M.A.; Partridge, L.; Serrano, M.; Kroemer, G. The Hallmarks of Aging. Cell 2013, 153, 1194–1217. [Google Scholar] [CrossRef] [Green Version]
- United Nations. World Population Prospects 2019; United Nations: New York, NY, USA, 2019. [Google Scholar]
- GBD 2019 Diseases and Injuries Collaborators. Global burden of 369 diseases and injuries in 204 countries and territories, 1990–2019: A systematic analysis for the Global Burden of Disease Study 2019. Lancet 2020, 396, 1204–1222. [Google Scholar] [CrossRef]
- JafariNasabian, P.; Inglis, J.; Reilly, W.; Kelly, O.J.; Ilich, J.Z. Aging human body: Changes in bone, muscle and body fat with consequent changes in nutrient intake. J. Endocrinol. 2017, 234, R37–R51. [Google Scholar] [CrossRef] [Green Version]
- Bouaziz, W.; Lang, P.O.; Schmitt, E.; Kaltenbach, G.; Geny, B.; Vogel, T. Health benefits of multicomponent training programmes in seniors: A systematic review. Int. J. Clin. Pract. 2016, 70, 520–536. [Google Scholar] [CrossRef] [PubMed]
- Jenkin, C.R.; Eime, R.M.; Westerbeek, H.; O’Sullivan, G.; Van Uffelen, J.G.Z. Sport and ageing: A systematic review of the determinants and trends of participation in sport for older adults. BMC Public Health 2017, 17, 9780. [Google Scholar] [CrossRef]
- Khan, S.S.; Singer, B.D.; Vaughan, D.E. Molecular and physiological manifestations and measurement of aging in humans. Aging Cell 2017, 16, 624–633. [Google Scholar] [CrossRef] [PubMed]
- Hernandez-Segura, A.; Nehme, J.; Demaria, M. Hallmarks of Cellular Senescence. Trends Cell Biol. 2018, 28, 436–453. [Google Scholar] [CrossRef]
- Partridge, L.; Deelen, J.; Slagboom, P.E. Facing up to the global challenges of ageing. Nat. Cell Biol. 2018, 561, 45–56. [Google Scholar] [CrossRef]
- Niedernhofer, L.; Kirkland, J.; Ladiges, W. Molecular pathology endpoints useful for aging studies. Ageing Res. Rev. 2017, 35, 241–249. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rose, M.R.; Charlesworth, B. A test of evolutionary theories of senescence. Nat. Cell Biol. 1980, 287, 141–142. [Google Scholar] [CrossRef] [PubMed]
- Campisi, J.; Kapahi, P.; Lithgow, G.J.; Melov, S.; Newman, J.C.; Verdin, E. From discoveries in ageing research to therapeutics for healthy ageing. Nat. Cell Biol. 2019, 571, 183–192. [Google Scholar] [CrossRef] [Green Version]
- Sun, N.; Youle, R.J.; Finkel, T. The Mitochondrial Basis of Aging. Mol. Cell 2016, 61, 654–666. [Google Scholar] [CrossRef] [Green Version]
- Trifunovic, A.; Wredenberg, A.; Falkenberg, M.; Spelbrink, J.N.; Rovio, A.T.; Bruder, C.E.; Bohlooly-Y, M.; Gidlöf, S.; Oldfors, A.; Wibom, R.; et al. Premature ageing in mice expressing defective mitochondrial DNA polymerase. Nat. Cell Biol. 2004, 429, 417–423. [Google Scholar] [CrossRef]
- Katajisto, P.; Döhla, J.; Chaffer, C.L.; Pentinmikko, N.; Marjanovic, N.; Iqbal, S.; Zoncu, R.; Chen, W.W.; Weinberg, R.A.; Sabatini, D.M. Asymmetric apportioning of aged mitochondria between daughter cells is required for stemness. Science 2015, 348, 340–343. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rodrigues, L.P.; Teixeira, V.R.; Alencar-Silva, T.; Simonassi-Paiva, B.; Pereira, R.W.; Pogue, R.; Carvalho, J.L. Hallmarks of aging and immunosenescence: Connecting the dots. Cytokine Growth Factor Rev. 2021, 59, 9–21. [Google Scholar] [CrossRef] [PubMed]
- Zhou, D.; Borsa, M.; Simon, A.K. Hallmarks and detection techniques of cellular senescence and cellular ageing in immune cells. Aging Cell 2021, 20, e13316. [Google Scholar] [CrossRef] [PubMed]
- Jayaraj, G.G.; Hipp, M.S.; Hartl, F.U. Functional Modules of the Proteostasis Network. Cold Spring Harb. Perspect. Biol. 2020, 12, a033951. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hipp, M.S.; Kasturi, P.; Hartl, F.U. The proteostasis network and its decline in ageing. Nat. Rev. Mol. Cell Biol. 2019, 20, 421–435. [Google Scholar] [CrossRef]
- Burch, J.B.; Augustine, A.D.; Frieden, L.A.; Hadley, E.; Howcroft, T.K.; Johnson, R.; Khalsa, P.S.; Kohanski, R.A.; Li, X.L.; Macchiarini, F.; et al. Advances in Geroscience: Impact on Healthspan and Chronic Disease. J. Gerontol. Ser. A 2014, 69, S1–S3. [Google Scholar] [CrossRef] [Green Version]
- Sierra, F. Moving Geroscience Into Uncharted Waters. J. Gerontol. Ser. A 2016, 71, 1385–1387. [Google Scholar] [CrossRef]
- Fulop, T.; Larbi, A.; Khalil, A.; Cohen, A.A.; Witkowski, J.M. Are We Ill Because We Age? Front. Physiol. 2019, 10, 1508. [Google Scholar] [CrossRef] [Green Version]
- Bergmann, O.; Bhardwaj, R.D.; Bernard, S.; Zdunek, S.; Barnabé-Heider, F.; Walsh, S.; Zupicich, J.; Alkass, K.; Buchholz, B.A.; Druid, H.; et al. Evidence for Cardiomyocyte Renewal in Humans. Science 2009, 324, 98–102. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Senyo, S.E.; Steinhauser, M.L.; Pizzimenti, C.L.; Yang, V.K.; Cai, L.; Wang, M.; Wu, T.-D.; Guerquin-Kern, J.-L.; Lechene, C.P.; Lee, R.T. Mammalian heart renewal by pre-existing cardiomyocytes. Nat. Cell Biol. 2013, 493, 433–436. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Laflamme, M.A.; Murry, C.E. Heart regeneration. Nat. Cell Biol. 2011, 473, 326–335. [Google Scholar] [CrossRef] [Green Version]
- Eschenhagen, T.; Bolli, R.; Braun, T.; Field, L.J.; Fleischmann, B.K.; Frisen, J.; Giacca, M.; Hare, J.M.; Houser, S.; Lee, R.T.; et al. Cardiomyocyte Regeneration: A Consensus Statement. Circulation 2017, 136, 680–686. [Google Scholar] [CrossRef]
- Mollova, M.; Bersell, K.; Walsh, S.; Savla, J.; Das, L.T.; Park, S.-Y.; Silberstein, L.E.; Dos Remedios, C.G.; Graham, D.; Colan, S.; et al. Cardiomyocyte proliferation contributes to heart growth in young humans. Proc. Natl. Acad. Sci. USA 2013, 110, 1446–1451. [Google Scholar] [CrossRef] [Green Version]
- Zhu, D.; Cheng, K. Cardiac Cell Therapy for Heart Repair: Should the Cells Be Left Out? Cells 2021, 10, 641. [Google Scholar] [CrossRef]
- Worku, M.G. Pluripotent and Multipotent Stem Cells and Current Therapeutic Applications: Review. Stem Cells Cloning 2021, 14, 3–7. [Google Scholar] [PubMed]
- Witman, N.; Zhou, C.; Beverborg, N.G.; Sahara, M.; Chien, K.R. Cardiac progenitors and paracrine mediators in cardiogenesis and heart regeneration. Semin. Cell Dev. Biol. 2020, 100, 29–51. [Google Scholar] [CrossRef] [PubMed]
- Sebastião, M.J.; Marcos-Silva, L.; Gomes-Alves, P.; Alves, P.M. Proteomic and Glyco(proteo)mic tools in the profiling of cardiac progenitors and pluripotent stem cell derived cardiomyocytes: Accelerating translation into therapy. Biotechnol. Adv. 2021, 49, 107755. [Google Scholar] [CrossRef]
- Riching, A.S.; Song, K. Cardiac Regeneration: New Insights into the Frontier of Ischemic Heart Failure Therapy. Front. Bioeng. Biotechnol. 2021, 8, 637538. [Google Scholar] [CrossRef]
- James, E.; Tomaskovic-Crook, E.; Crook, J. Bioengineering Clinically Relevant Cardiomyocytes and Cardiac Tissues from Pluripotent Stem Cells. Int. J. Mol. Sci. 2021, 22, 3005. [Google Scholar] [CrossRef]
- Huang, J.; Feng, Q.; Wang, L.; Zhou, B. Human Pluripotent Stem Cell-Derived Cardiac Cells: Application in Disease Modeling, Cell Therapy, and Drug Discovery. Front. Cell Dev. Biol. 2021, 9, 655161. [Google Scholar] [CrossRef] [PubMed]
- Hsia, G.S.P.; Esposito, J.; da Rocha, L.A.; Ramos, S.L.G.; Okamoto, O.K. Clinical Application of Human Induced Pluripotent Stem Cell-Derived Organoids as an Alternative to Organ Transplantation. Stem Cells Int. 2021, 2021, 1–16. [Google Scholar] [CrossRef]
- Hashemzadeh, M.R.; Yazdi, M.E.T.; Amiri, M.S.; Mousavi, S.H. Stem cell therapy in the heart: Biomaterials as a key route. Tissue Cell 2021, 71, 101504. [Google Scholar] [CrossRef]
- Campostrini, G.; Windt, L.M.; van Meer, B.J.; Bellin, M.; Mummery, C.L. Cardiac Tissues from Stem Cells: New Routes to Maturation and Cardiac Regeneration. Circ. Res. 2021, 128, 775–801. [Google Scholar] [CrossRef] [PubMed]
- Bolli, R.; Solankhi, M.; Tang, X.-L.; Kahlon, A. Cell Therapy in Patients with Heart Failure: A Comprehensive Review and Emerging Concepts. Cardiovasc. Res. 2021. [Google Scholar] [CrossRef] [PubMed]
- Rajabi, M.; Kassiotis, C.; Razeghi, P.; Taegtmeyer, H. Return to the fetal gene program protects the stressed heart: A strong hypothesis. Heart Fail. Rev. 2007, 12, 331–343. [Google Scholar] [CrossRef] [PubMed]
- Taegtmeyer, H.; Sen, S.; Vela, D. Return to the fetal gene program: A suggested metabolic link to gene expression in the heart. Ann. N.Y. Acad. Sci. 2010, 1188, 191–198. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Talman, V.; Teppo, J.; Pöhö, P.; Movahedi, P.; Vaikkinen, A.; Karhu, S.T.; Trošt, K.; Suvitaival, T.; Heikkonen, J.; Pahikkala, T.; et al. Molecular Atlas of Postnatal Mouse Heart Development. J. Am. Heart Assoc. 2018, 7, e010378. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Locatelli, P.; Belaich, M.N.; Lopez, A.; Olea, F.D.; Urnaga Vega, M.; Gimenez, C.S.; Simonin, J.A.; Bauza, M.D.; Castillo, M.G.; Cuniberti, L.A.; et al. Novel insights into cardiac regeneration based on differential fetal and adult ovine heart transcriptomic analysis. Am. J. Physiol. Circ. Physiol. 2020, 318, H994–H1007. [Google Scholar] [CrossRef]
- Katz, M.G.; Fargnoli, A.S.; Kendle, A.P.; Hajjar, R.J.; Bridges, C.R. The role of microRNAs in cardiac development and regenerative capacity. Am. J. Physiol. Circ. Physiol. 2016, 310, H528–H541. [Google Scholar] [CrossRef] [Green Version]
- Wu, Y.H.; Zhao, H.; Zhou, L.P.; Zhao, C.X.; Wu, Y.F.; Zhen, L.X.; Li, J.; Ge, D.X.; Xu, L.; Lin, L.; et al. miR-134 Modulates the Proliferation of Human Cardiomyocyte Progenitor Cells by Targeting Meis2. Int. J. Mol. Sci. 2015, 16, 5199. [Google Scholar] [CrossRef] [Green Version]
- Ponnusamy, M.; Liu, F.; Zhang, Y.-H.; Li, R.-B.; Zhai, M.; Liu, F.; Zhou, L.-Y.; Liu, C.-Y.; Yan, K.-W.; Dong, Y.-H.; et al. Long Noncoding RNA CPR (Cardiomyocyte Proliferation Regulator) Regulates Cardiomyocyte Proliferation and Cardiac Repair. Circulation 2019, 139, 2668–2684. [Google Scholar] [CrossRef]
- Porrello, E.R.; Mahmoud, A.I.; Simpson, E.; Hill, J.A.; Richardson, J.A.; Olson, E.N.; Sadek, H. Transient Regenerative Potential of the Neonatal Mouse Heart. Science 2011, 331, 1078–1080. [Google Scholar] [CrossRef] [Green Version]
- Porrello, E.R.; Mahmoud, A.I.; Simpson, E.; Johnson, B.A.; Grinsfelder, D.; Canseco, D.; Mammen, P.P.; Rothermel, B.A.; Olson, E.N.; Sadek, H.A. Regulation of neonatal and adult mammalian heart regeneration by the miR-15 family. Proc. Natl. Acad. Sci. USA 2013, 110, 187–192. [Google Scholar] [CrossRef] [Green Version]
- Quaife-Ryan, G.A.; Sim, C.B.; Ziemann, M.; Kaspi, A.; Rafehi, H.; Ramialison, M.; El-Osta, A.; Hudson, J.E.; Porrello, E.R. Multicellular Transcriptional Analysis of Mammalian Heart Regeneration. Circulation 2017, 136, 1123–1139. [Google Scholar] [CrossRef]
- Hu, P.; Liu, J.; Zhao, J.; Wilkins, B.J.; Lupino, K.; Wu, H.; Pei, L. Single-nucleus transcriptomic survey of cell diversity and functional maturation in postnatal mammalian hearts. Genes Dev. 2018, 32, 1344–1357. [Google Scholar] [CrossRef] [PubMed]
- Gan, J.; Sonntag, H.-J.; Tang, M.K.; Cai, D.; Lee, K. Integrative Analysis of the Developing Postnatal Mouse Heart Transcriptome. PLoS ONE 2015, 10, e0133288. [Google Scholar] [CrossRef] [PubMed]
- De Laughter, D.M.; Bick, A.G.; Wakimoto, H.; McKean, D.; Gorham, J.M.; Kathiriya, I.S.; Hinson, J.T.; Homsy, J.; Gray, J.; Pu, W.; et al. Single-Cell Resolution of Temporal Gene Expression during Heart Development. Dev. Cell 2016, 39, 480–490. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Anversa, P.; Nadal-Ginard, B. Myocyte renewal and ventricular remodelling. Nat. Cell Biol. 2002, 415, 240–243. [Google Scholar] [CrossRef]
- Alvarado-Ramy, F.; Beltrami, E.M. New guidelines for occupational exposure to blood-borne viruses. Clevel. Clin. J. Med. 2003, 70, 457–465. [Google Scholar] [CrossRef]
- Beltrami, A.P.; Barlucchi, L.; Torella, D.; Baker, M.; Limana, F.; Chimenti, S.; Kasahara, H.; Rota, M.; Musso, E.; Urbanek, K.; et al. Adult Cardiac Stem Cells Are Multipotent and Support Myocardial Regeneration. Cell 2003, 114, 763–776. [Google Scholar] [CrossRef] [Green Version]
- Orlic, D.; Kajstura, J.; Chimenti, S.; Jakoniuk, I.; Anderson, S.M.; Li, B.; Pickel, J.; McKay, R.D.G.; Nadal-Ginard, B.; Bodine, D.M.; et al. Bone marrow cells regenerate infarcted myocardium. Nat. Cell Biol. 2001, 410, 701–705. [Google Scholar] [CrossRef]
- Murry, C.E.; Soonpaa, M.H.; Reinecke, H.; Nakajima, H.; Nakajima, H.O.; Rubart, M.; Pasumarthi, K.B.S.; Virag, J.I.; Bartelmez, S.H.; Poppa, V.; et al. Haematopoietic stem cells do not transdifferentiate into cardiac myocytes in myocardial infarcts. Nat. Cell Biol. 2004, 428, 664–668. [Google Scholar] [CrossRef] [PubMed]
- Ieda, M.; Fu, J.-D.; Delgado-Olguin, P.; Vedantham, V.; Hayashi, Y.; Bruneau, B.G.; Srivastava, D. Direct Reprogramming of Fibroblasts into Functional Cardiomyocytes by Defined Factors. Cell 2010, 142, 375–386. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Balsam, L.B.; Wagers, A.J.; Christensen, J.L.; Kofidis, T.; Weissman, I.L.; Robbins, R.C. Haematopoietic stem cells adopt mature haematopoietic fates in ischaemic myocardium. Nat. Cell Biol. 2004, 428, 668–673. [Google Scholar] [CrossRef] [PubMed]
- Heusch, G. Molecular basis of cardioprotection: Signal transduction in ischemic pre-, post-, and remote conditioning. Circ. Res. 2015, 116, 674–699. [Google Scholar] [CrossRef]
- Heusch, G.; Bøtker, H.E.; Przyklenk, K.; Redington, A.; Yellon, D. Remote Ischemic Conditioning. J. Am. Coll. Cardiol. 2015, 65, 177–195. [Google Scholar] [CrossRef] [Green Version]
- Sprick, J.D.; Mallet, R.T.; Przyklenk, K.; Rickards, C.A. Ischaemic and hypoxic conditioning: Potential for protection of vital organs. Exp. Physiol. 2019, 104, 278–294. [Google Scholar] [CrossRef]
- Tsibulnikov, S.Y.; Maslov, L.N.; Gorbunov, A.S.; Voronkov, N.S.; Boshchenko, A.A.; Popov, S.V.; Prokudina, E.S.; Singh, N.; Downey, J.M. A Review of Humoral Factors in Remote Preconditioning of the Heart. J. Cardiovasc. Pharmacol. Ther. 2019, 24, 403–421. [Google Scholar] [CrossRef]
- Shrivastava, S.; Srivastava, D.; Olson, E.N.; DiMaio, J.M.; Bock-Marquette, I. Thymosin β4 and cardiac repair. Ann. N.Y. Acad. Sci. 2010, 1194, 87–96. [Google Scholar] [CrossRef] [PubMed]
- Magadum, A.; Kaur, K.; Zangi, L. mRNA-Based Protein Replacement Therapy for the Heart. Mol. Ther. 2019, 27, 785–793. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Goldstein, A.L.; Slater, F.D.; White, A. Preparation, assay, and partial purification of a thymic lymphocytopoietic factor (thymosin). Proc. Natl. Acad. Sci. USA 1966, 56, 1010–1017. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huff, T.; Müller, C.S.; Otto, A.M.; Netzker, R.; Hannappel, E. β-Thymosins, small acidic peptides with multiple functions. Int. J. Biochem. Cell Biol. 2001, 33, 205–220. [Google Scholar] [CrossRef]
- Hannappel, E. Beta-Thymosins. Ann. N.Y. Acad. Sci. 2007, 1112, 21–37. [Google Scholar] [CrossRef] [PubMed]
- Hoch, K.; Volk, D.E. Chapter One—Structures of Thymosin Proteins. In Vitamins and Hormones; Litwack, G., Ed.; Academic Press: London, UK, 2016; pp. 1–24. [Google Scholar]
- Mora, C.A.; Baumann, C.A.; Paino, J.E.; Goldstein, A.L.; Badamchian, M. Biodistribution of synthetic thymosin beta 4 in the serum, urine, and major organs of mice. Int. J. Immunopharmacol. 1997, 19, 1–8. [Google Scholar] [CrossRef]
- Hannappel, E.; Xu, G.J.; Morgan, J.; Hempstead, J.; Horecker, B.L. Thymosin beta 4: A ubiquitous peptide in rat and mouse tissues. Proc. Natl. Acad. Sci. USA 1982, 79, 2172–2175. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, G.J.; Hannappel, E.; Morgan, J.; Hempstead, J.; Horecker, B.L. Synthesis of thymosin beta 4 by peritoneal macrophages and adherent spleen cells. Proc. Natl. Acad. Sci. USA 1982, 79, 4006–4009. [Google Scholar] [CrossRef] [Green Version]
- Hannappel, E.; Leibold, W. Biosynthesis rates and content of thymosin beta 4 in cell lines. Arch. Biochem. Biophys. 1985, 240, 236–241. [Google Scholar] [CrossRef]
- Lin, S.C.; Morrison-Bogorad, M. Developmental expression of mRNAs encoding thymosins beta 4 and beta 10 in rat brain and other tissues. J. Mol. Neurosci. 1990, 2, 35–44. [Google Scholar] [CrossRef] [PubMed]
- Nemolato, S.; Cabras, T.; Fanari, M.; Cau, F.; Fanni, D.; Gerosa, C.; Manconi, B.; Messana, I.; Castagnola, M.; Faa, G. Immunoreactivity of thymosin beta 4 in human foetal and adult genitourinary tract. Eur. J. Histochem. 2010, 54, e43. [Google Scholar] [CrossRef] [Green Version]
- Salhab, M.; Papillier, P.; Perreau, C.; Guyader-Joly, C.; Dupont, J.; Mermillod, P.; Uzbekova, S. Thymosins β-4 and β-10 are expressed in bovine ovarian follicles and upregulated in cumulus cells during meiotic maturation. Reprod. Fertil. Dev. 2010, 22, 1206. [Google Scholar] [CrossRef] [PubMed]
- Gerosa, C.; Fanni, D.; Nemolato, S.; Locci, A.; Marinelli, V.; Cabras, T.; Messana, I.; Castagnola, M.; Monga, G.; Fanos, V.; et al. Thymosin beta-10 expression in developing human kidney. J. Matern. Neonatal Med. 2010, 23, 125–128. [Google Scholar] [CrossRef] [PubMed]
- Nemolato, S.; Cabras, T.; Cau, F.; Fanari, M.U.; Fanni, D.; Manconi, B.; Messana, I.; Castagnola, M.; Faa, G. Different Thymosin Beta 4 Immunoreactivity in Foetal and Adult Gastrointestinal Tract. PLoS ONE 2010, 5, e9111. [Google Scholar] [CrossRef] [Green Version]
- Fanni, D.; Gerosa, C.; Nemolato, S.; Locci, A.; Marinelli, V.; Cabras, T.; Messana, I.; Fanos, V.; Castagnola, M.; Faa, G. Thymosin beta 10 expression in developing human salivary glands. Early Hum. Dev. 2011, 87, 779–783. [Google Scholar] [CrossRef]
- Rossdeutsch, A.; Smart, N.; Dubé, K.N.; Turner, M.; Riley, P.R. Essential role for thymosin beta4 in regulating vascular smooth muscle cell development and vessel wall stability. Circ. Res. 2012, 111, e89–e102. [Google Scholar] [CrossRef] [Green Version]
- Shin, S.-H.; Lee, S.; Bae, J.-S.; Jee, J.-G.; Cha, H.-J.; Lee, A.Y.M. Thymosin Beta4 Regulates Cardiac Valve Formation Via Endothelial-Mesenchymal Transformation in Zebrafish Embryos. Mol. Cells 2014, 37, 330–336. [Google Scholar] [CrossRef] [Green Version]
- Saunders, V.; Dewing, J.M.; Sanchez-Elsner, T.; Wilson, D.I. Expression and localisation of thymosin beta-4 in the developing human early fetal heart. PLoS ONE 2018, 13, e0207248. [Google Scholar] [CrossRef]
- Yang, H.M.; Kang, S.W.; Sung, J.; Kim, K.; Kleinman, H. Purinergic Signaling Involvement in Thymosin beta4-mediated Corneal Epithelial Cell Migration. Curr. Eye Res. 2020, 1–7. [Google Scholar] [CrossRef]
- Padmanabhan, K.; Grobe, H.; Cohen, J.; Soffer, A.; Mahly, A.; Adir, O.; Zaidel-Bar, R.; Luxenburg, C. Thymosin β4 is essential for adherens junction stability and epidermal planar cell polarity. Development 2020, 147. [Google Scholar] [CrossRef]
- Mantri, M.; Scuderi, G.J.; Abedini-Nassab, R.; Wang, M.F.Z.; McKellar, D.; Shi, H.; Grodner, B.; Butcher, J.T.; De Vlaminck, I. Spatiotemporal single-cell RNA sequencing of developing chicken hearts identifies interplay between cellular differentiation and morphogenesis. Nat. Commun. 2021, 12, 1–13. [Google Scholar] [CrossRef]
- Dathe, V.; Brand-Saberi, B. Expression of thymosin beta4 during chick development. Brain Struct. Funct. 2004, 208, 27–32. [Google Scholar] [CrossRef] [PubMed]
- Gómez-Márquez, J.; del Amo, F.F.; Carpintero, P.; Anadón, R. High levels of mouse thymosin beta4 mRNA in differentiating P19 embryonic cells and during development of cardiovascular tissues. Biochim. Biophys. Acta 1996, 1306, 187–193. [Google Scholar] [CrossRef]
- Bock-Marquette, I.; Saxena, A.; White, M.D.; DiMaio, J.M.; Srivastava, D. Thymosin beta4 activates integrin-linked kinase and promotes cardiac cell migration, survival and cardiac repair. Nature 2004, 432, 466–472. [Google Scholar] [CrossRef]
- Hoff, M.J.V.D.; Moorman, A.F.; Ruijter, J.M.; Lamers, W.H.; Bennington, R.W.; Markwald, R.R.; Wessels, A. Myocardialization of the Cardiac Outflow Tract. Dev. Biol. 1999, 212, 477–490. [Google Scholar] [CrossRef] [Green Version]
- Smart, N.; Risebro, C.A.; Melville, A.A.D.; Moses, K.; Schwartz, R.J.; Chien, K.R.; Riley, P.R. Thymosin beta-4 Is Essential for Coronary Vessel Development and Promotes Neovascularization via Adult Epicardium. Ann. N.Y. Acad. Sci. 2007, 1112, 171–188. [Google Scholar] [CrossRef] [PubMed]
- Banerjee, I.; Morris, T.M.; Evans, S.M.; Chen, J. and Chen, J. Thymosin beta4 is not required for embryonic viability or vascular development. Circ. Res. 2013, 112, e25–e28. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Smart, N.; Risebro, C.A.; Melville, A.A.; Moses, K.; Schwartz, R.J.; Chien, K.R.; Riley, P.R. Thymosin beta4 induces adult epicardial progenitor mobilization and neovascularization. Nature 2007, 445, 177–182. [Google Scholar] [CrossRef] [PubMed]
- Banerjee, I.; Zhang, J.; Moore-Morris, T.; Lange, S.; Shen, T.; Dalton, N.D.; Gu, Y.; Peterson, K.L.; Evans, S.M.; Chen, J. Thymosin beta 4 is dispensable for murine cardiac development and function. Circ. Res. 2012, 110, 456–464. [Google Scholar] [CrossRef] [PubMed]
- Dubé, K.N.; Smart, N. Thymosin β4 and the vasculature: Multiple roles in development, repair and protection against disease. Expert Opin. Biol. Ther. 2018, 18, 131–139. [Google Scholar] [CrossRef] [PubMed]
- Faa, G.; Piras, M.; Mancuso, L.; Coni, P.; Pichiri, G.; Orrù, G.; Fanni, D.; Gerosa, C.; Cao, G.; Taibi, R.; et al. Thymosin beta-4 prenatal administration improves fetal development and halts side effects due to preterm delivery. Eur. Rev. Med. Pharmacol. Sci. 2021, 25, 431–437. [Google Scholar] [PubMed]
- Muñoz-Espín, D.; Serrano, M. Cellular senescence: From physiology to pathology. Nat. Rev. Mol. Cell Biol. 2014, 15, 482–496. [Google Scholar] [CrossRef] [PubMed]
- Kirkwood, T.B.; Austad, S.N. Why do we age? Nature 2000, 408, 233–238. [Google Scholar] [CrossRef]
- Hinkel, R.; Ball, H.L.; DiMaio, J.M.; Shrivastava, S.; Thatcher, J.E.; Singh, A.N.; Sun, X.; Faskerti, G.; Olson, E.N.; Kupatt, C.; et al. C-terminal variable AGES domain of Thymosin beta4: The molecule’s primary contribution in support of post-ischemic cardiac function and repair. J. Mol. Cell. Cardiol. 2015, 87, 113–125. [Google Scholar] [CrossRef] [PubMed]
- Young, J.D.; Lawrence, A.J.; MacLean, A.G.; Leung, B.P.; McInnes, I.B.; Canas, B.; Pappin, D.J.C.; Stevenson, R.D. Thymosin beta 4 sulfoxide is an anti-inflammatory agent generated by monocytes in the presence of glucocorticoids. Nat. Med. 1999, 5, 1424–1427. [Google Scholar] [CrossRef]
- Belsky, J.B.; Rivers, E.P.; Filbin, M.R.; Lee, P.J.; Morris, D.C. Thymosin beta 4 regulation of actin in sepsis. Expert Opin. Biol. Ther. 2018, 18, 193–197. [Google Scholar] [CrossRef] [PubMed]
- Carion, T.W.; Ebrahim, A.S.; Kracht, D.; Agrawal, A.; Strand, E.; Kaddurah, O.; McWhirter, C.R.; Sosne, G.; Berger, E.A. Thymosin Beta-4 and Ciprofloxacin Adjunctive Therapy Improves Pseudomonas Aeruginosa-Induced Keratitis. Cells 2018, 7, 145. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sosne, G.; Qiu, P.; Kurpakus-Wheater, M. Thymosin beta 4: A novel corneal wound healing and anti-inflammatory agent. Clin. Ophthalmol. 2007, 1, 201–207. [Google Scholar]
- Ho, E.N.; Kwok, W.H.; Lau, M.Y.; Wong, A.S.; Wan, T.S.; Lam, K.K.; Schiff, P.J.; Stewart, B.D. Doping control analysis of TB-500, a synthetic version of an active region of thymosin beta(4), in equine urine and plasma by liquid chromatography-mass spectrometry. J. Chromatogr. A 2012, 1265, 57–69. [Google Scholar] [CrossRef] [PubMed]
- Kleinman, H.K.; Sosne, G. Thymosin beta4 Promotes Dermal Healing. Vitam. Horm. 2016, 102, 251–275. [Google Scholar]
- Yang, W.S.; Kang, S.; Sung, J.; Kleinman, H.K. Thymosin beta4: Potential to treat epidermolysis bullosa and other severe dermal injuries. Eur. J. Dermatol. 2019, 29, 459–467. [Google Scholar]
- Hinkel, R.; Klett, K.; Bähr, A.; Kupatt, C. Thymosin beta4-mediated protective effects in the heart. Expert Opin. Biol. Ther. 2018, 18, 121–129. [Google Scholar] [CrossRef]
- Srivastava, D.; Ieda, M.; Fu, J.; Qian, L. Cardiac repair with thymosin β4 and cardiac reprogramming factors. Ann. N.Y. Acad. Sci. 2012, 1270, 66–72. [Google Scholar] [CrossRef]
- Peng, H.; Xu, J.; Yang, X.P.; Dai, X.; Peterson, E.L.; Carretero, O.A.; Rhaleb, N.E. Thymosin-β4 prevents cardiac rupture and improves cardiac function in mice with myocardial infarction. Am. J. Physiol. Circ. Physiol. 2014, 307, H741–H751. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huang, Z.; Song, Y.; Pang, Z.; Zhang, B.; Yang, H.; Shi, H.; Chen, J.; Gong, H.; Qian, J.; Ge, J. Targeted delivery of thymosin beta 4 to the injured myocardium using CREKA-conjugated nanoparticles. Int. J. Nanomed. 2017, 12, 3023–3036. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Quan, Z.; Wang, Q.-L.; Zhou, P.; Wang, G.-D.; Tan, Y.-Z.; Wang, H.-J. Thymosin β4 promotes the survival and angiogenesis of transplanted endothelial progenitor cells in the infarcted myocardium. Int. J. Mol. Med. 2017, 39, 1347–1356. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhao, Y.; Song, J.; Bi, X.; Gao, J.; Shen, Z.; Zhu, J.; Fu, G. Thymosin β4 promotes endothelial progenitor cell angiogenesis via a vascular endothelial growth factor-dependent mechanism. Mol. Med. Rep. 2018, 18, 2314–2320. [Google Scholar] [CrossRef] [Green Version]
- Kassem, K.M.; Vaid, S.; Peng, H.; Sarkar, S.; Rhaleb, N.E. Tβ4-Ac-SDKP pathway: Any relevance for the cardiovascular system? Can. J. Physiol. Pharmacol. 2019, 97, 589–599. [Google Scholar] [CrossRef]
- Bjørklund, G.; Dadar, M.; Aaseth, J.; Chirumbolo, S. Thymosin β4: A Multi-Faceted Tissue Repair Stimulating Protein in Heart Injury. Curr. Med. Chem. 2020, 27, 6294–6305. [Google Scholar] [CrossRef]
- Wang, Y.-L.; Yu, S.-N.; Shen, H.-R.; Wang, H.-J.; Wu, X.-P.; Wang, Q.-L.; Zhou, B.; Tan, Y.-Z. Thymosin β4 released from functionalized self-assembling peptide activates epicardium and enhances repair of infarcted myocardium. Theranostics 2021, 11, 4262–4280. [Google Scholar] [CrossRef]
- Gladka, M.M.; Kohela, A.; Molenaar, B.; Versteeg, D.; Kooijman, L.; Monshouwer-Kloots, J.; Kremer, V.; Vos, H.R.; Huibers, M.M.H.; Haigh, J.J.; et al. Cardiomyocytes stimulate angiogenesis after ischemic injury in a ZEB2-dependent manner. Nat. Commun. 2021, 12, 1–16. [Google Scholar] [CrossRef]
- Bock-Marquette, I.; Shrivastava, S.; Pipes, G.T.; Thatcher, J.E.; Blystone, A.; Shelton, J.M.; Galindo, C.L.; Melegh, B.; Srivastava, D.; Olson, E.N.; et al. Thymosin beta4 mediated PKC activation is essential to initiate the embryonic coronary developmental program and epicardial progenitor cell activation in adult mice in vivo. J. Mol. Cell Cardiol. 2009, 46, 728–738. [Google Scholar] [CrossRef] [Green Version]
- Lepilina, A.; Coon, A.N.; Kikuchi, K.; Holdway, J.E.; Roberts, R.W.; Burns, C.G.; Poss, K.D. A dynamic epicardial injury response supports progenitor cell activity during zebrafish heart regeneration. Cell 2006, 127, 607–619. [Google Scholar] [CrossRef] [Green Version]
- Cao, Y.; Duca, S.; Cao, J. Epicardium in Heart Development. Cold Spring Harb. Perspect. Biol. 2019, 12, a037192. [Google Scholar] [CrossRef] [PubMed]
- Cao, J.; Poss, K.D. The epicardium as a hub for heart regeneration. Nat. Rev. Cardiol. 2018, 15, 631–647. [Google Scholar] [CrossRef] [PubMed]
- Cao, Y.; Cao, J. Covering and Re-Covering the Heart: Development and Regeneration of the Epicardium. J. Cardiovasc. Dev. Dis. 2018, 6, 3. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wessels, A.; Pérez-Pomares, J. The epicardium and epicardially derived cells (EPDCs) as cardiac stem cells. Anat. Rec. A Discov. Mol. Cell. Evol. Biol. 2004, 276A, 43–57. [Google Scholar] [CrossRef]
- Quijada, P.; Trembley, M.A.; Small, E.M. The Role of the Epicardium during Heart Development and Repair. Circ. Res. 2020, 126, 377–394. [Google Scholar] [CrossRef] [PubMed]
- Männer, J. The development of pericardial villi in the chick embryo. Brain Struct. Funct. 1992, 186, 379–385. [Google Scholar]
- Lie-Venema, H.; Akker, N.M.S.V.D.; Bax, N.A.M.; Winter, E.M.; Maas, S.; Kekarainen, T.; Hoeben, R.C.; DeRuiter, M.C.; Poelmann, R.E.; Groot, A.C.G.-D. Origin, Fate, and Function of Epicardium-Derived Cells (EPDCs) in Normal and Abnormal Cardiac Development. Sci. World, J. 2007, 7, 1777–1798. [Google Scholar] [CrossRef] [Green Version]
- Katz, T.C.; Singh, M.; Degenhardt, K.; Rivera-Feliciano, J.; Johnson, R.L.; Epstein, J.A.; Tabin, C.J. Distinct Compartments of the Proepicardial Organ Give Rise to Coronary Vascular Endothelial Cells. Dev. Cell 2012, 22, 639–650. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cai, C.-L.; Martin, J.C.; Sun, Y.; Cui, L.; Wang, L.; Ouyang, K.; Yang, L.; Bu, L.; Liang, X.; Zhang, X.; et al. A myocardial lineage derives from Tbx18 epicardial cells. Nat. Cell Biol. 2008, 454, 104–108. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhou, B.; Ma, Q.; Rajagopal, S.; Wu, S.M.; Domian, I.; Rivera-Feliciano, J.; Jiang, D.; Von Gise, A.; Ikeda, S.; Chien, K.R.; et al. Epicardial progenitors contribute to the cardiomyocyte lineage in the developing heart. Nat. Cell Biol. 2008, 454, 109–113. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yamaguchi, Y.; Cavallero, S.; Patterson, M.; Shen, H.; Xu, J.; Kumar, S.R.; Sucov, H.M. Adipogenesis and epicardial adipose tissue: A novel fate of the epicardium induced by mesenchymal transformation and PPARgamma activation. Proc. Natl. Acad. Sci. USA 2015, 112, 2070–2075. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Porrello, E.R.; Olson, E.N. A neonatal blueprint for cardiac regeneration. Stem Cell Res. 2014, 13, 556–570. [Google Scholar] [CrossRef] [Green Version]
- Lu, J.; Richardson, J.A.; Olson, E.N. Capsulin: A novel bHLH transcription factor expressed in epicardial progenitors and mesenchyme of visceral organs. Mech. Dev. 1998, 73, 23–32. [Google Scholar] [CrossRef]
- Kumar, N.; Liao, T.D.; Romero, C.A.; Maheshwari, M.; Peterson, E.L.; Carretero, O.A. Thymosin beta4 Deficiency Exacerbates Renal and Cardiac Injury in Angiotensin-II-Induced Hypertension. Hypertension 2018, 71, 1133–1142. [Google Scholar] [CrossRef] [PubMed]
- Morita, T.; Hayashi, K. Tumor Progression Is Mediated by Thymosin-beta4 through a TGFbeta/MRTF Signaling Axis. Mol. Cancer Res. 2018, 16, 880–893. [Google Scholar] [CrossRef] [Green Version]
- Sribenja, S.; Wongkham, S.; Wongkham, C.; Yao, Q.; Chen, C. Roles and mechanisms of beta-thymosins in cell migration and cancer metastasis: An update. Cancer Investig. 2013, 31, 103–110. [Google Scholar] [CrossRef] [PubMed]
- Wong, H.Y.; Wang, G.M.; Croessmann, S.; Zabransky, D.J.; Chu, D.; Garay, J.P.; Cidado, J.; Cochran, R.L.; Beaver, J.A.; Aggarwal, A.; et al. TMSB4Y is a candidate tumor suppressor on the Y chromosome and is deleted in male breast cancer. Oncotarget 2015, 6, 44927–44940. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Caers, J.; Otjacques, E.; Hose, D.; Klein, B.; Vanderkerken, K. Thymosin beta4 in multiple myeloma: Friend or foe. Ann. N.Y. Acad. Sci. 2010, 1194, 125–129. [Google Scholar] [CrossRef] [PubMed]
- Caers, J.; Hose, D.; Kuipers, I.; Bos, T.J.; Van Valckenborgh, E.; Menu, E.; De Bruyne, E.; Goldschmidt, H.; Van Camp, B.; Klein, B.; et al. Thymosin beta4 has tumor suppressive effects and its decreased expression results in poor prognosis and decreased survival in multiple myeloma. Haematologica 2010, 95, 163–167. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Goldstein, A.L.; Kleinman, H.K. Advances in the basic and clinical applications of thymosin beta4. Expert Opin. Biol. Ther. 2015, 15, S139–S145. [Google Scholar] [CrossRef]
- Schneider, M.D. Regenerative medicine: Prometheus unbound. Nat. Cell Biol. 2004, 432, 451–453. [Google Scholar]





Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Maar, K.; Hetenyi, R.; Maar, S.; Faskerti, G.; Hanna, D.; Lippai, B.; Takatsy, A.; Bock-Marquette, I. Utilizing Developmentally Essential Secreted Peptides Such as Thymosin Beta-4 to Remind the Adult Organs of Their Embryonic State—New Directions in Anti-Aging Regenerative Therapies. Cells 2021, 10, 1343. https://doi.org/10.3390/cells10061343
Maar K, Hetenyi R, Maar S, Faskerti G, Hanna D, Lippai B, Takatsy A, Bock-Marquette I. Utilizing Developmentally Essential Secreted Peptides Such as Thymosin Beta-4 to Remind the Adult Organs of Their Embryonic State—New Directions in Anti-Aging Regenerative Therapies. Cells. 2021; 10(6):1343. https://doi.org/10.3390/cells10061343
Chicago/Turabian StyleMaar, Klaudia, Roland Hetenyi, Szabolcs Maar, Gabor Faskerti, Daniel Hanna, Balint Lippai, Aniko Takatsy, and Ildiko Bock-Marquette. 2021. "Utilizing Developmentally Essential Secreted Peptides Such as Thymosin Beta-4 to Remind the Adult Organs of Their Embryonic State—New Directions in Anti-Aging Regenerative Therapies" Cells 10, no. 6: 1343. https://doi.org/10.3390/cells10061343