
Genotoxic Bystander Signals from Irradiated Human Mesenchymal Stromal Cells Mainly Localize in the 10–100 kDa Fraction of Conditioned Medium

 ,
, 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Collection of Human Femoral Heads
2.2. Isolation of Human MSC
2.3. Isolation of Human CD34+ Cells
2.4. Preparation of Fractions of MSC Conditioned Medium
2.5. Heat Inactivation of MSC Conditioned and Control Medium
2.6. RIBE Analysis
2.7. Immunofluorescence Staining of γH2AX
2.8. Cytogenetic Analysis
2.9. Statistical Analysis
3. Results
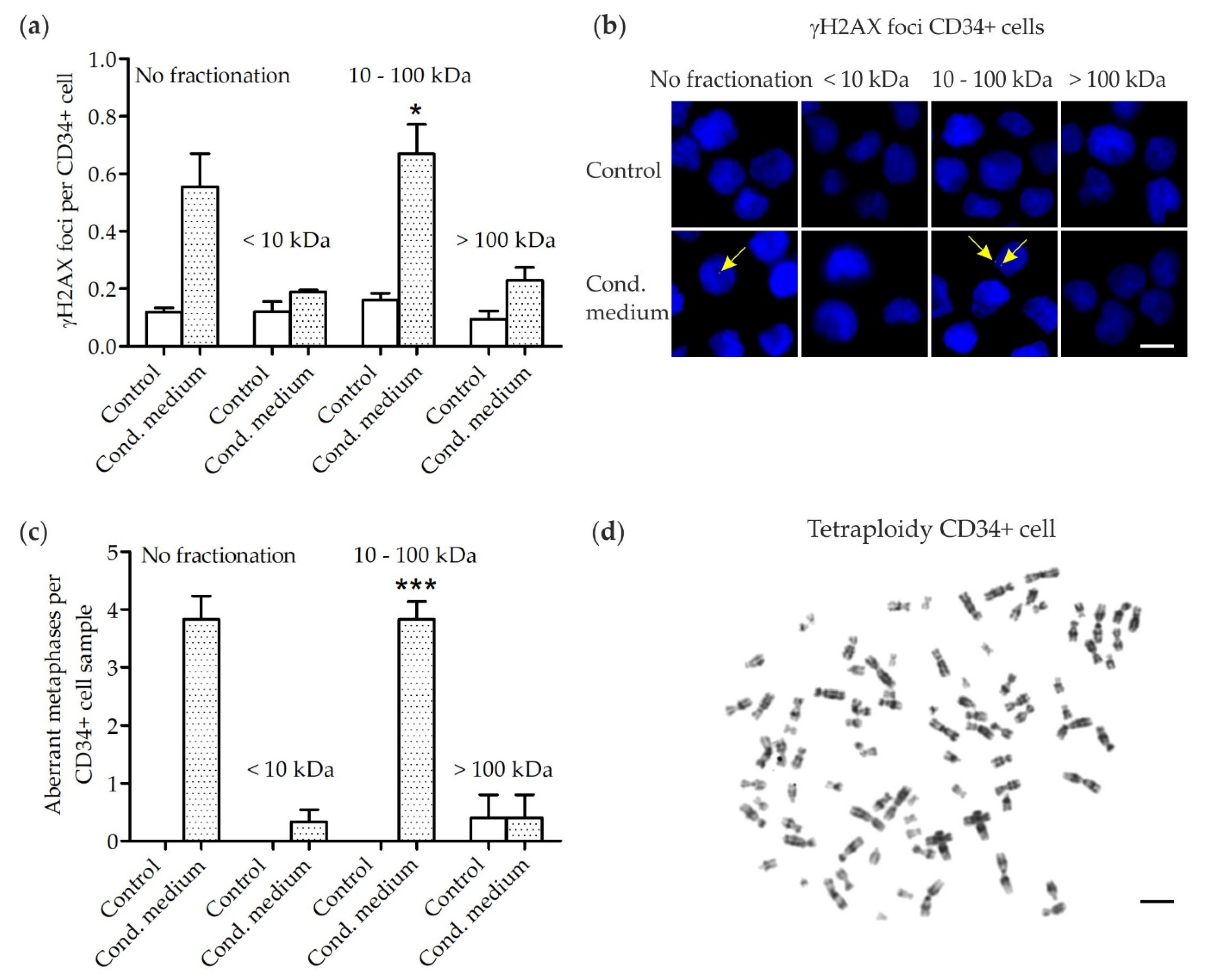
3.1. DNA Damage in Human CD34+ Cells
3.2. Chromosomal Instability in Human CD34+ Cells
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Swerdlow, S.H.; Campo, E.; Harris, N.L.; Jaffe, E.S.; Pileri, S.A.; Stein, H.; Thiele, J. WHO Classification of Tumours of Haematopoietic and Lymphoid Tissues, 4th ed.; IARC: Lyon, France, 2017; pp. 153–155. [Google Scholar]
- Ok, C.Y.; Patel, K.P.; Garcia-Manero, G.; Routbort, M.J.; Fu, B.; Tang, G.; Goswami, M.; Singh, R.; Kanagal-Shamanna, R.; Pierce, S.A.; et al. Mutational profiling of therapy-related myelodysplastic syndromes and acute myeloid leukemia by next generation sequencing, a comparison with de novo diseases. Leuk Res. 2015, 39, 348–354. [Google Scholar] [CrossRef] [Green Version]
- Wong, T.N.; Ramsingh, G.; Young, A.L.; Miller, C.A.; Touma, W.; Welch, J.S.; Lamprecht, T.L.; Shen, D.; Hundal, J.; Fulton, R.S.; et al. Role of TP53 mutations in the origin and evolution of therapy-related acute myeloid leukaemia. Nature 2015, 518, 552–555. [Google Scholar] [CrossRef] [Green Version]
- Fianchi, L.; Pagano, L.; Piciocchi, A.; Candoni, A.; Gaidano, G.; Breccia, M.; Criscuolo, M.; Specchia, G.; Maria Pogliani, E.; Maurillo, L.; et al. Characteristics and outcome of therapy-related myeloid neoplasms: Report from the Italian network on secondary leukemias. Am. J. Hematol. 2015, 90, E80–E85. [Google Scholar] [CrossRef]
- Granfeldt Ostgard, L.S.; Medeiros, B.C.; Sengelov, H.; Norgaard, M.; Andersen, M.K.; Dufva, I.H.; Friis, L.S.; Kjeldsen, E.; Marcher, C.W.; Preiss, B.; et al. Epidemiology and Clinical Significance of Secondary and Therapy-Related Acute Myeloid Leukemia: A National Population-Based Cohort Study. J. Clin. Oncol. 2015, 33, 3641–3649. [Google Scholar] [CrossRef] [PubMed]
- Watson, G.E.; Lorimore, S.A.; Macdonald, D.A.; Wright, E.G. Chromosomal instability in unirradiated cells induced in vivo by a bystander effect of ionizing radiation. Cancer Res. 2000, 60, 5608–5611. [Google Scholar]
- Nagasawa, H.; Little, J.B. Induction of sister chromatid exchanges by extremely low doses of alpha-particles. Cancer Res. 1992, 52, 6394–6396. [Google Scholar] [PubMed]
- Zhou, H.; Randers-Pehrson, G.; Waldren, C.A.; Vannais, D.; Hall, E.J.; Hei, T.K. Induction of a bystander mutagenic effect of alpha particles in mammalian cells. Proc. Natl. Acad. Sci. USA 2000, 97, 2099–2104. [Google Scholar] [CrossRef] [Green Version]
- Azzam, E.I.; de Toledo, S.M.; Raaphorst, G.P.; Mitchel, R.E. Low-dose ionizing radiation decreases the frequency of neoplastic transformation to a level below the spontaneous rate in C3H 10T1/2 cells. Radiat. Res. 1996, 146, 369–373. [Google Scholar] [CrossRef] [PubMed]
- Shao, C.; Lyng, F.M.; Folkard, M.; Prise, K.M. Calcium fluxes modulate the radiation-induced bystander responses in targeted glioma and fibroblast cells. Radiat. Res. 2006, 166, 479–487. [Google Scholar] [CrossRef] [PubMed]
- Chen, S.; Zhao, Y.; Han, W.; Zhao, G.; Zhu, L.; Wang, J.; Bao, L.; Jiang, E.; Xu, A.; Hei, T.K.; et al. Mitochondria-dependent signalling pathway are involved in the early process of radiation-induced bystander effects. Br. J. Cancer 2008, 98, 1839–1844. [Google Scholar] [CrossRef] [Green Version]
- Tartier, L.; Gilchrist, S.; Burdak-Rothkamm, S.; Folkard, M.; Prise, K.M. Cytoplasmic irradiation induces mitochondrial-dependent 53BP1 protein relocalization in irradiated and bystander cells. Cancer Res. 2007, 67, 5872–5879. [Google Scholar] [CrossRef] [Green Version]
- Zhou, H.; Ivanov, V.N.; Lien, Y.C.; Davidson, M.; Hei, T.K. Mitochondrial function and nuclear factor-kappaB-mediated signaling in radiation-induced bystander effects. Cancer Res. 2008, 68, 2233–2240. [Google Scholar] [CrossRef] [Green Version]
- Shao, C.; Stewart, V.; Folkard, M.; Michael, B.D.; Prise, K.M. Nitric oxide-mediated signaling in the bystander response of individually targeted glioma cells. Cancer Res. 2003, 63, 8437–8442. [Google Scholar]
- Li, J.; He, M.; Shen, B.; Yuan, D.; Shao, C. Alpha particle-induced bystander effect is mediated by ROS via a p53-dependent SCO2 pathway in hepatoma cells. Int. J. Radiat. Biol. 2013, 89, 1028–1034. [Google Scholar] [CrossRef]
- Shao, C.; Folkard, M.; Prise, K.M. Role of TGF-beta1 and nitric oxide in the bystander response of irradiated glioma cells. Oncogene 2008, 27, 434–440. [Google Scholar] [CrossRef] [Green Version]
- Gow, M.D.; Seymour, C.B.; Ryan, L.A.; Mothersill, C.E. Induction of bystander response in human glioma cells using high-energy electrons: A role for TGF-beta1. Radiat. Res. 2010, 173, 769–778. [Google Scholar] [CrossRef] [PubMed]
- Shao, C.; Furusawa, Y.; Aoki, M.; Ando, K. Role of gap junctional intercellular communication in radiation-induced bystander effects in human fibroblasts. Radiat. Res. 2003, 160, 318–323. [Google Scholar] [CrossRef] [PubMed]
- Azzam, E.I.; de Toledo, S.M.; Little, J.B. Direct evidence for the participation of gap junction-mediated intercellular communication in the transmission of damage signals from alpha -particle irradiated to nonirradiated cells. Proc. Natl. Acad. Sci. USA 2001, 98, 473–478. [Google Scholar] [CrossRef] [PubMed]
- Peng, Y.; Zhang, M.; Zheng, L.; Liang, Q.; Li, H.; Chen, J.T.; Guo, H.; Yoshina, S.; Chen, Y.Z.; Zhao, X.; et al. Cysteine protease cathepsin B mediates radiation-induced bystander effects. Nature 2017, 547, 458–462. [Google Scholar] [CrossRef] [Green Version]
- Xu, S.; Wang, J.; Ding, N.; Hu, W.; Zhang, X.; Wang, B.; Hua, J.; Wei, W.; Zhu, Q. Exosome-mediated microRNA transfer plays a role in radiation-induced bystander effect. RNA Biol. 2015, 12, 1355–1363. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ariyoshi, K.; Miura, T.; Kasai, K.; Fujishima, Y.; Nakata, A.; Yoshida, M. Radiation-Induced Bystander Effect is Mediated by Mitochondrial DNA in Exosome-Like Vesicles. Sci. Rep. 2019, 9, 9103. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jella, K.K.; Moriarty, R.; McClean, B.; Byrne, H.J.; Lyng, F.M. Reactive oxygen species and nitric oxide signaling in bystander cells. PLoS ONE 2018, 13, e0195371. [Google Scholar] [CrossRef]
- Lyng, F.M.; Howe, O.L.; McClean, B. Reactive oxygen species-induced release of signalling factors in irradiated cells triggers membrane signalling and calcium influx in bystander cells. Int. J. Radiat. Biol. 2011, 87, 683–695. [Google Scholar] [CrossRef]
- Lyng, F.M.; Maguire, P.; McClean, B.; Seymour, C.; Mothersill, C. The involvement of calcium and MAP kinase signaling pathways in the production of radiation-induced bystander effects. Radiat. Res. 2006, 165, 400–409. [Google Scholar] [CrossRef]
- Popp, H.D.; Naumann, N.; Brendel, S.; Henzler, T.; Weiss, C.; Hofmann, W.K.; Fabarius, A. Increase of DNA damage and alteration of the DNA damage response in myelodysplastic syndromes and acute myeloid leukemias. Leuk Res. 2017, 57, 112–118. [Google Scholar] [CrossRef]
- Popp, H.D.; Brendel, S.; Hofmann, W.K.; Fabarius, A. Immunofluorescence Microscopy of gammaH2AX and 53BP1 for Analyzing the Formation and Repair of DNA Double-strand Breaks. J. Vis. Exp. 2017, 129, 56617. [Google Scholar]
- Heim, S.; Mitelman, F. Cancer Cytogenetics, 3rd ed.; Wiley-Blackwell: Hoboken, NJ, USA, 2009; pp. 9–16. [Google Scholar]
- McGowan-Jordan, J.; Simons, A.; Schmid, M. ISCN 2016 An International System for Human Cytogenetic Nomenclature (2016); Karger: Basel, Switzerland, 2016. [Google Scholar]
- Jacobs, P.A.; Brunton, M.; Court Brown, W.M.; Doll, R.; Goldstein, H. Change of human chromosome count distribution with age: Evidence for a sex differences. Nature 1963, 197, 1080–1081. [Google Scholar] [CrossRef]
- Pierre, R.V.; Hoagland, H.C. Age-associated aneuploidy: Loss of Y chromosome from human bone marrow cells with aging. Cancer 1972, 30, 889–894. [Google Scholar] [CrossRef]
- Sokolov, M.V.; Dickey, J.S.; Bonner, W.M.; Sedelnikova, O.A. gamma-H2AX in bystander cells: Not just a radiation-triggered event, a cellular response to stress mediated by intercellular communication. Cell Cycle 2007, 6, 2210–2212. [Google Scholar] [CrossRef] [Green Version]
- Sedelnikova, O.A.; Nakamura, A.; Kovalchuk, O.; Koturbash, I.; Mitchell, S.A.; Marino, S.A.; Brenner, D.J.; Bonner, W.M. DNA double-strand breaks form in bystander cells after microbeam irradiation of three-dimensional human tissue models. Cancer Res. 2007, 67, 4295–4302. [Google Scholar] [CrossRef] [Green Version]
- Dickey, J.S.; Zemp, F.J.; Altamirano, A.; Sedelnikova, O.A.; Bonner, W.M.; Kovalchuk, O. H2AX phosphorylation in response to DNA double-strand break formation during bystander signalling: Effect of microRNA knockdown. Radiat. Prot. Dosim. 2011, 143, 264–269. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lorimore, S.A.; McIlrath, J.M.; Coates, P.J.; Wright, E.G. Chromosomal instability in unirradiated hemopoietic cells resulting from a delayed in vivo bystander effect of gamma radiation. Cancer Res. 2005, 65, 5668–5673. [Google Scholar] [CrossRef] [Green Version]
- Lorimore, S.A.; Chrystal, J.A.; Robinson, J.I.; Coates, P.J.; Wright, E.G. Chromosomal instability in unirradiated hemaopoietic cells induced by macrophages exposed in vivo to ionizing radiation. Cancer Res. 2008, 68, 8122–8126. [Google Scholar] [CrossRef] [Green Version]
- Tanaka, K.; Goto, H.; Nishimura, Y.; Kasahara, K.; Mizoguchi, A.; Inagaki, M. Tetraploidy in cancer and its possible link to aging. Cancer Sci. 2018, 109, 2632–2640. [Google Scholar] [CrossRef] [PubMed]
- Huang, L.; Wang, S.A.; DiNardo, C.; Li, S.; Hu, S.; Xu, J.; Zhou, W.; Goswami, M.; Medeiros, L.J.; Tang, G. Tetraploidy/near-tetraploidy acute myeloid leukemia. Leuk Res. 2017, 53, 20–27. [Google Scholar] [CrossRef] [PubMed]
- Mothersill, C.; Seymour, C.B. Cell-cell contact during gamma irradiation is not required to induce a bystander effect in normal human keratinocytes: Evidence for release during irradiation of a signal controlling survival into the medium. Radiat. Res. 1998, 149, 256–262. [Google Scholar] [CrossRef]
- Lepleux, C.; Marie-Brasset, A.; Temelie, M.; Boulanger, M.; Brotin, E.; Goldring, M.B.; Hirtz, C.; Vares, G.; Nakajima, T.; Saintigny, Y.; et al. Bystander effectors of chondrosarcoma cells irradiated at different LET impair proliferation of chondrocytes. J. Cell Commun. Signal 2019, 13, 343–356. [Google Scholar] [CrossRef] [Green Version]
- Hu, B.; Wu, L.; Han, W.; Zhang, L.; Chen, S.; Xu, A.; Hei, T.K.; Yu, Z. The time and spatial effects of bystander response in mammalian cells induced by low dose radiation. Carcinogenesis 2006, 27, 245–251. [Google Scholar] [CrossRef]


{kind=link}
| Pt | Age/ | Cytogenetics CD34+ Cells | Cytogenetics CD34+ Cells | Cytogenetics CD34+ Cells | Cytogenetics CD34+ Cells | ||||
|---|---|---|---|---|---|---|---|---|---|
| Sex | No Fractionation | <10 kDa | 10–100 kDa | >100 kDa | |||||
| Control | CM | Control | CM | Control | CM | Control | CM | ||
| #1 | 84/♀ | 46,XX[25] | 46,XX[20] 53,XX,+1,+2,+5,+6,+14,+21,+22[1] 92,XXXX[4] | 46,XX[20] | 46,XX[25] | 46,XX[25] | 46,XX[22] 92,XXXX[3] | NA | NA |
| #2 | 65/♂ | 46,XY[25] | 46,XY[20] 92,XXXX[1] 184,XXXXYYYY,chtb(11)(q23)[1] 46,XY,dup(13q)[1] 47,XY,+21,chtb(11)(p12)[1] 46,XY,chtb(9)(12)[1] | 46,XY[25] | 46,XY[22] | 46,XY[25] | 46,XY[21] 92,XXXX[2] 69,XXY[1] 47,XY,+3[1] | 46,XY[25] | 46,XY[25] |
| #3 | 62/♂ | 46,XY[25] | 46,XY[22] 92,XXYY[3] | 46,XY[25] | 46,XY[25] | 46,XY[25] | 46,XY[20] 92,XXYY[3] 46,XY,chtb(5)(q33)[1] 46,XY,+f[1] | 46,XY[25] | 46,XY[25] |
| #4 | 62/♂ | 46,XY[25] | 46,XY[21] 92,XXYY[3] 92,XXYY,chtb(2p)[1] | 46,XY[25] | 46,XY[23] 46,XY,chtb(14q)[1] | 46,XY[25] | 46,XY[22] 92,XXYY[1] 184,XXXXYYYY[1] 46,XY,chtb(7p)[1] | 46,XY[23] 184,XXXXYYYY[2] | 46,XY[25] |
| #5 | 85/♂ | 46,XY[13] 45,X,-Y[12] | 46,XY[10] 45,X,-Y[12] 90,XX,-Y,-Y[1] 92,XXYY[1] 184,XXXXYYYY[1] | 46,XY[5] 45,X,-Y[20] | 46,XY[7] 45,X,-Y[18] | 46,XY[21] 45,X,-Y[4] | 46,XY[18] 45,X,-Y[3] 92,XXYY[2] 47,XY,+2[1] 50,XY,+1,+7,+9,+14[1] | 46,XY[10] 45,X,-Y[15] | 46,XY[13] 45,X,-Y[7] 92,XXYY[1] 90,XX,-Y,-Y[1] |
| #6 | 52/♂ | 46,XY[25] | 46,XY[22] 92,XXYY[2] 184,XXXXYYYY[1] | 46,XY[25] | 46,XY[24] 46,XY,+f[1] | 46,XY[21] | 46,XY[21] 92,XXYY[3] 184,XXXXYYYY[1] | 46,XY[25] | 46,XY[25] |
| Pt | Age/ | Cytogenetics CD34+ Cells | Cytogenetics CD34+ Cells | Cell Doubling Time (Days) | Cell Doubling Time (Days) | ||||
|---|---|---|---|---|---|---|---|---|---|
| Sex | Control | CM | Control | CM | Control | CM | Control | CM | |
| No Heat Inactivation | +Heat Inactivation | No Heat Inactivation | +Heat Inactivation | ||||||
| #5 | 85/♂ | 46,XY[13] 45,X,-Y[12] | 46,XY[10] 45,X,-Y[12] 90,XX,-Y,-Y[1] 92,XXYY[1] 184,XXXXYYYY[1] | 46,XY[15] | 46,XY[14] | 1.0 | 1.0 | 1.9 | 1.5 |
| #6 | 52/♂ | 46,XY[25] | 46,XY[22] 92,XXYY[2] 184,XXXXYYYY[1] | 46,XY[25] | 46,XY[24] | 1.2 | 1.0 | 1.5 | 1.7 |
| #7 | 86/♀ | 46,XX[25] | 46,XX[20] 46,XX,chtb(10q)[1] 46,XX,del(19)(p10)[1] 92,XXXX[2] 184,XXXXXXXX[1] | 46,XX[23] 92,XXXX[2] | 46,XX[22] 92,XXXX[2] 46,XX,chtb(4q)[1] | 1.3 | 1.2 | 1.8 | 1.4 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kohl, V.; Fabarius, A.; Drews, O.; Bierbaum, M.; Jawhar, A.; Darwich, A.; Weiss, C.; Flach, J.; Brendel, S.; Kleiner, H.; et al. Genotoxic Bystander Signals from Irradiated Human Mesenchymal Stromal Cells Mainly Localize in the 10–100 kDa Fraction of Conditioned Medium. Cells 2021, 10, 827. https://doi.org/10.3390/cells10040827
Kohl V, Fabarius A, Drews O, Bierbaum M, Jawhar A, Darwich A, Weiss C, Flach J, Brendel S, Kleiner H, et al. Genotoxic Bystander Signals from Irradiated Human Mesenchymal Stromal Cells Mainly Localize in the 10–100 kDa Fraction of Conditioned Medium. Cells. 2021; 10(4):827. https://doi.org/10.3390/cells10040827
Chicago/Turabian StyleKohl, Vanessa, Alice Fabarius, Oliver Drews, Miriam Bierbaum, Ahmed Jawhar, Ali Darwich, Christel Weiss, Johanna Flach, Susanne Brendel, Helga Kleiner, and et al. 2021. "Genotoxic Bystander Signals from Irradiated Human Mesenchymal Stromal Cells Mainly Localize in the 10–100 kDa Fraction of Conditioned Medium" Cells 10, no. 4: 827. https://doi.org/10.3390/cells10040827