
Somatic Mutational Profile of High-Grade Serous Ovarian Carcinoma and Triple-Negative Breast Carcinoma in Young and Elderly Patients: Similarities and Divergences

 and
and
Abstract
:1. Introduction
2. Methods
3. Results
3.1. Base Substitutions
3.2. Mutational Signatures
3.3. Number of (Non-Synonymous) Coding Variants
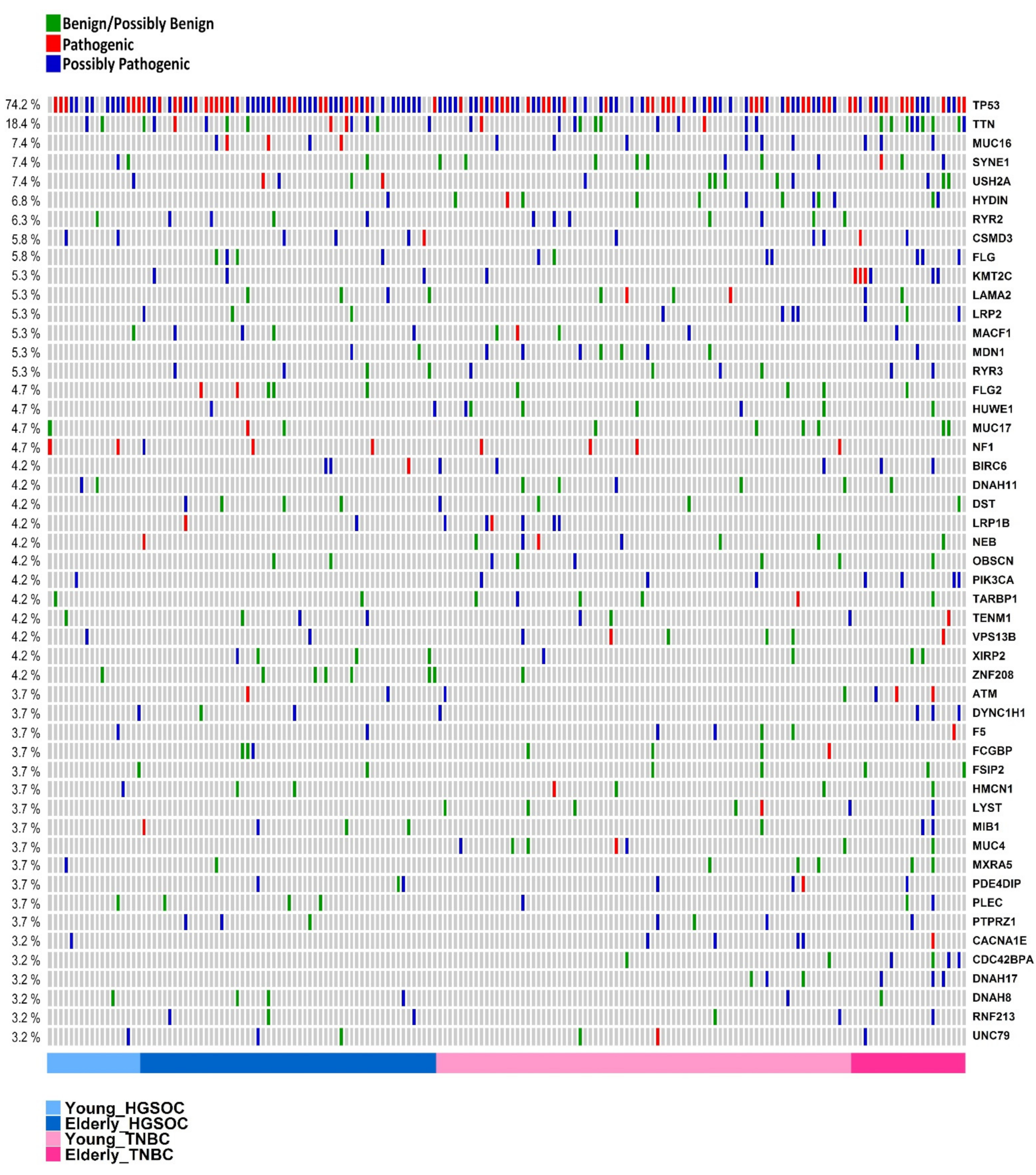
3.4. Genes Most Commonly Affected
3.5. Cancer Driver Genes
4. DNA Repair
5. Discussion
6. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Howlader, N.; Noone, A.M.; Krapcho, M.; Miller, D.; Brest, A.; Yu, M.; Ruhl, J.; Tatalovich, Z.; Mariotto, A.; Lewis, D.R.; et al. Cancer Statistics Review, 1975–2016—SEER Statistics. Available online: https://seer.cancer.gov/csr/1975_2016/ (accessed on 9 March 2020).
- Cancello, G.; Maisonneuve, P.; Rotmensz, N.; Viale, G.; Mastropasqua, M.G.; Pruneri, G.; Veronesi, P.; Torrisi, R.; Montagna, E.; Luini, A.; et al. Prognosis and Adjuvant Treatment Effects in Selected Breast Cancer Subtypes of Very Young Women (<35 Years) with Operable Breast Cancer. Ann. Oncol. 2010, 21, 1974–1981. [Google Scholar] [CrossRef] [PubMed]
- Liedtke, C.; Rody, A.; Gluz, O.; Baumann, K.; Beyer, D.; Kohls, E.-B.; Lausen, K.; Hanker, L.; Holtrich, U.; Becker, S.; et al. The Prognostic Impact of Age in Different Molecular Subtypes of Breast Cancer. Breast Cancer Res. Treat. 2015, 152, 667–673. [Google Scholar] [CrossRef] [PubMed]
- Torre, L.A.; Trabert, B.; DeSantis, C.E.; Miller, K.D.; Samimi, G.; Runowicz, C.D.; Gaudet, M.M.; Jemal, A.; Siegel, R.L. Ovarian Cancer Statistics, 2018. CA Cancer J. Clin. 2018, 68, 284–296. [Google Scholar] [CrossRef] [PubMed]
- Lockley, M.; Stoneham, S.J.; Olson, T.A. Ovarian Cancer in Adolescents and Young Adults. Pediatr. Blood Cancer 2019, 66, e27512. [Google Scholar] [CrossRef] [PubMed]
- Gockley, A.; Melamed, A.; Bregar, A.J.; Clemmer, J.T.; Birrer, M.; Schorge, J.O.; Del Carmen, M.G.; Rauh-Hain, J.A. Outcomes of Women with High-Grade and Low-Grade Advanced-Stage Serous Epithelial Ovarian Cancer. Obstet. Gynecol. 2017, 129, 439–447. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Elzakkers, J.C.J.; van der Aa, M.A.; van Altena, A.M.; de Hullu, J.A.; Harmsen, M.G. Further Insights into the Role of Tumour Characteristics in Survival of Young Women with Epithelial Ovarian Cancer. Gynecol. Oncol. 2019, 155, 213–219. [Google Scholar] [CrossRef] [PubMed]
- Yoshikawa, N.; Kajiyama, H.; Mizuno, M.; Shibata, K.; Kawai, M.; Nagasaka, T.; Kikkawa, F. Clinicopathologic Features of Epithelial Ovarian Carcinoma in Younger vs. Older Patients: Analysis in Japanese Women. J. Gynecol. Oncol. 2014, 25, 118–123. [Google Scholar] [CrossRef] [Green Version]
- Duska, L.R.; Chang, Y.C.; Flynn, C.E.; Chen, A.H.; Goodman, A.; Fuller, A.F.; Nikrui, N. Epithelial Ovarian Carcinoma in the Reproductive Age Group. Cancer 1999, 85, 2623–2629. [Google Scholar] [CrossRef]
- Lalrinpuii, E.; Bhageerathy, P.S.; Sebastian, A.; Jeyaseelan, L.; Thomas, A.; Chandy, R.; Peedicayil, A. Ovarian Cancer in Young Women. Indian J. Surg. Oncol. 2017, 8, 540–547. [Google Scholar] [CrossRef]
- Chan, J.K.; Urban, R.; Cheung, M.K.; Osann, K.; Shin, J.Y.; Husain, A.; Teng, N.N.; Kapp, D.S.; Berek, J.S.; Leiserowitz, G.S. Ovarian Cancer in Younger vs Older Women: A Population-Based Analysis. Br. J. Cancer 2006, 95, 1314–1320. [Google Scholar] [CrossRef] [Green Version]
- Copson, E.R.; Maishman, T.C.; Tapper, W.J.; Cutress, R.I.; Greville-Heygate, S.; Altman, D.G.; Eccles, B.; Gerty, S.; Durcan, L.T.; Jones, L.; et al. Germline BRCA Mutation and Outcome in Young-Onset Breast Cancer (POSH): A Prospective Cohort Study. Lancet Oncol. 2018, 19, 169–180. [Google Scholar] [CrossRef] [Green Version]
- Carraro, D.M.; Koike Folgueira, M.A.A.; Garcia Lisboa, B.C.; Ribeiro Olivieri, E.H.; Vitorino Krepischi, A.C.; de Carvalho, A.F.; de Carvalho Mota, L.D.; Puga, R.D.; do Socorro Maciel, M.; Michelli, R.A.D.; et al. Comprehensive Analysis of BRCA1, BRCA2 and TP53 Germline Mutation and Tumor Characterization: A Portrait of Early-Onset Breast Cancer in Brazil. PLoS ONE 2013, 8, e57581. [Google Scholar] [CrossRef] [PubMed]
- Hahnen, E.; Hauke, J.; Engel, C.; Neidhardt, G.; Rhiem, K.; Schmutzler, R.K. Germline Mutations in Triple-Negative Breast Cancer. Breast Care 2017, 12, 15–19. [Google Scholar] [CrossRef] [PubMed]
- Brianese, R.C.; de Mello Nakamura, K.D.; Ramalho, R.F.; de Figueiredo Barros, B.D.; e Ferreira, E.N.; da Cruz Formiga, M.N.; de Andrade, V.P.; de Lima, V.C.C.; Carraro, D.M. BRCA1 Deficiency Is a Recurrent Event in Early-Onset Triple-Negative Breast Cancer: A Comprehensive Analysis of Germline Mutations and Somatic Promoter Methylation. Breast Cancer Res. Treat. 2018, 167, 803–814. [Google Scholar] [CrossRef] [PubMed]
- Gonzalez-Angulo, A.M.; Timms, K.M.; Liu, S.; Chen, H.; Litton, J.K.; Potter, J.; Lanchbury, J.S.; Stemke-Hale, K.; Hennessy, B.T.; Arun, B.K.; et al. Incidence and Outcome of BRCA Mutations in Unselected Patients with Triple Receptor-Negative Breast Cancer. Clin. Cancer Res. 2011, 17, 1082–1089. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, M.; Zhang, J.; Ouyang, T.; Li, J.; Wang, T.; Fan, Z.; Fan, T.; Lin, B.; Xie, Y. Incidence of BRCA1 Somatic Mutations and Response to Neoadjuvant Chemotherapy in Chinese Women with Triple-Negative Breast Cancer. Gene 2016, 584, 26–30. [Google Scholar] [CrossRef]
- Xie, Y.; Gou, Q.; Wang, Q.; Zhong, X.; Zheng, H. The Role of BRCA Status on Prognosis in Patients with Triple-Negative Breast Cancer. Oncotarget 2017, 8, 87151–87162. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sugino, K.; Tamura, R.; Nakaoka, H.; Yachida, N.; Yamaguchi, M.; Mori, Y.; Yamawaki, K.; Suda, K.; Ishiguro, T.; Adachi, S.; et al. Germline and Somatic Mutations of Homologous Recombination-Associated Genes in Japanese Ovarian Cancer Patients. Sci. Rep. 2019, 9, 17808. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Krivokuca, A.; Boljevic, I.; Jovandic, S.; Magic, Z.; Mandic, A.; Tomasevic, Z.; Brankovic-Magic, M. Germline Mutations in Cancer Susceptibility Genes in High Grade Serous Ovarian Cancer in Serbia. J. Hum. Genet. 2019, 64, 281–290. [Google Scholar] [CrossRef] [PubMed]
- Maistro, S.; Teixeira, N.; Encinas, G.; Katayama, M.L.H.; Niewiadonski, V.D.T.; Cabral, L.G.; Ribeiro, R.M.; Gaburo Junior, N.; de Gouvêa, A.C.R.C.; Carraro, D.M.; et al. Germline Mutations in BRCA1 and BRCA2 in Epithelial Ovarian Cancer Patients in Brazil. BMC Cancer 2016, 16, 934. [Google Scholar] [CrossRef]
- Bernards, S.S.; Norquist, B.M.; Harrell, M.I.; Agnew, K.J.; Lee, M.K.; Walsh, T.; Swisher, E.M. Genetic Characterization of Early Onset Ovarian Carcinoma. Gynecol. Oncol. 2016, 140, 221–225. [Google Scholar] [CrossRef] [Green Version]
- Eoh, K.J.; Kim, H.M.; Lee, J.-Y.; Kim, S.; Kim, S.W.; Kim, Y.T.; Nam, E.J. Mutation Landscape of Germline and Somatic BRCA1/2 in Patients with High-Grade Serous Ovarian Cancer. BMC Cancer 2020, 20, 204. [Google Scholar] [CrossRef]
- Lord, C.J.; Ashworth, A. BRCAness Revisited. Nat. Rev. Cancer 2016, 16, 110–120. [Google Scholar] [CrossRef] [PubMed]
- The Cancer Genome Atlas Network. Comprehensive Molecular Portraits of Human Breast Tumours. Nature 2012, 490, 61–70. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bowtell, D.D.; Böhm, S.; Ahmed, A.A.; Aspuria, P.-J.; Bast, R.C.; Beral, V.; Berek, J.S.; Birrer, M.J.; Blagden, S.; Bookman, M.A.; et al. Rethinking Ovarian Cancer II: Reducing Mortality from High-Grade Serous Ovarian Cancer. Nat. Rev. Cancer 2015, 15, 668–679. [Google Scholar] [CrossRef] [PubMed]
- Sæther, N.H.; Skuja, E.; Irmejs, A.; Maksimenko, J.; Miklasevics, E.; Purkalne, G.; Gardovskis, J. Platinum-Based Neoadjuvant Chemotherapy in BRCA1-Positive Breast Cancer: A Retrospective Cohort Analysis and Literature Review. Hered. Cancer Clin. Pract. 2018, 16, 9. [Google Scholar] [CrossRef] [PubMed]
- Muggia, F.; Safra, T.; Dubeau, L. BRCA Genes: Lessons Learned from Experimental and Clinical Cancer. Ann. Oncol. 2011, 22 (Suppl. 1), i7–i10. [Google Scholar] [CrossRef]
- O’Shaughnessy, J.; Osborne, C.; Pippen, J.; Yoffe, M.; Patt, D.; Monaghan, G.; Rocha, C.; Ossovskaya, V.; Sherman, B.; Bradley, C. Efficacy of BSI-201, a Poly (ADP-Ribose) Polymerase-1 (PARP1) Inhibitor, in Combination with Gemcitabine/Carboplatin (G/C) in Patients with Metastatic Triple-Negative Breast Cancer (TNBC): Results of a Randomized Phase II Trial. J. Clin. Oncol. 2009, 27, 3. [Google Scholar] [CrossRef]
- Ledermann, J.; Harter, P.; Gourley, C.; Friedlander, M.; Vergote, I.; Rustin, G.; Scott, C.; Meier, W.; Shapira-Frommer, R.; Safra, T.; et al. Olaparib Maintenance Therapy in Platinum-Sensitive Relapsed Ovarian Cancer. N. Engl. J. Med. 2012, 366, 1382–1392. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dent, R.A.; Lindeman, G.J.; Clemons, M.; Wildiers, H.; Chan, A.; McCarthy, N.J.; Singer, C.F.; Lowe, E.S.; Watkins, C.L.; Carmichael, J. Phase I Trial of the Oral PARP Inhibitor Olaparib in Combination with Paclitaxel for First- or Second-Line Treatment of Patients with Metastatic Triple-Negative Breast Cancer. Breast Cancer Res. 2013, 15, R88. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shah, S.P.; Roth, A.; Goya, R.; Oloumi, A.; Ha, G.; Zhao, Y.; Turashvili, G.; Ding, J.; Tse, K.; Haffari, G.; et al. The Clonal and Mutational Evolution Spectrum of Primary Triple-Negative Breast Cancers. Nature 2012, 486, 395–399. [Google Scholar] [CrossRef] [PubMed]
- Nik-Zainal, S.; Davies, H.; Staaf, J.; Ramakrishna, M.; Glodzik, D.; Zou, X.; Martincorena, I.; Alexandrov, L.B.; Martin, S.; Wedge, D.C.; et al. Landscape of Somatic Mutations in 560 Breast Cancer Whole-Genome Sequences. Nature 2016, 534, 47–54. [Google Scholar] [CrossRef] [PubMed]
- Kan, Z.; Ding, Y.; Kim, J.; Jung, H.H.; Chung, W.; Lal, S.; Cho, S.; Fernandez-Banet, J.; Lee, S.K.; Kim, S.W.; et al. Multi-Omics Profiling of Younger Asian Breast Cancers Reveals Distinctive Molecular Signatures. Nat. Commun. 2018, 9, 1725. [Google Scholar] [CrossRef]
- Banerji, S.; Cibulskis, K.; Rangel-Escareno, C.; Brown, K.K.; Carter, S.L.; Frederick, A.M.; Lawrence, M.S.; Sivachenko, A.Y.; Sougnez, C.; Zou, L.; et al. Sequence Analysis of Mutations and Translocations across Breast Cancer Subtypes. Nature 2012, 486, 405–409. [Google Scholar] [CrossRef]
- Cancer Genome Atlas Research Network. Integrated Genomic Analyses of Ovarian Carcinoma. Nature 2011, 474, 609–615. [Google Scholar] [CrossRef]
- Sukhai, M.A.; Craddock, K.J.; Thomas, M.; Hansen, A.R.; Zhang, T.; Siu, L.; Bedard, P.; Stockley, T.L.; Kamel-Reid, S. A classification system for clinical relevance of somatic variants identified in molecular profiling of cancer. Genet. Med. 2016, 18, 128–136. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, M.M.; Datto, M.; Duncavage, E.J.; Kulkarni, S.; Lindeman, N.I.; Roy, S.; Tsimberidou, A.M.; Vnencak-Jones, C.L.; Wolff, D.J.; Younes, A.; et al. Standards and guidelines for the interpretation and reporting of sequence variants in cancer: A joint consensus recommendation of the association for molecular pathology, american society of clinical oncology, and college of american pathologists. J. Mol. Diagn. 2017, 19, 4–23. [Google Scholar] [CrossRef] [Green Version]
- Abbott, K.L.; Nyre, E.T.; Abrahante, J.; Ho, Y.-Y.; Isaksson Vogel, R.; Starr, T.K. The Candidate Cancer Gene Database: A Database of Cancer Driver Genes from Forward Genetic Screens in Mice. Nucleic Acids Res. 2015, 43, D844–D848. [Google Scholar] [CrossRef] [Green Version]
- Chakravarty, D.; Gao, J.; Phillips, S.M.; Kundra, R.; Zhang, H.; Wang, J.; Rudolph, J.E.; Yaeger, R.; Soumerai, T.; Nissan, M.H.; et al. Oncokb: A precision oncology knowledge base. JCO Precis. Oncol. 2017, 1, 1–16. [Google Scholar] [CrossRef] [PubMed]
- Lange, S.S.; Takata, K.; Wood, R.D. DNA Polymerases and Cancer. Nat. Rev. Cancer 2011, 11, 96–110. [Google Scholar] [CrossRef] [Green Version]
- Wood, R.D.; Mitchell, M.; Sgouros, J.; Lindahl, T. Human DNA Repair Genes. Science 2001, 291, 1284–1289. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chae, Y.K.; Anker, J.F.; Carneiro, B.A.; Chandra, S.; Kaplan, J.; Kalyan, A.; Santa-Maria, C.A.; Platanias, L.C.; Giles, F.J. Genomic Landscape of DNA Repair Genes in Cancer. Oncotarget 2016, 7, 23312–23321. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mayakonda, A.; Lin, D.-C.; Assenov, Y.; Plass, C.; Koeffler, H.P. Maftools: Efficient and Comprehensive Analysis of Somatic Variants in Cancer. Genome Res. 2018, 28, 1747–1756. [Google Scholar] [CrossRef] [Green Version]
- Alexandrov, L.B.; Nik-Zainal, S.; Wedge, D.C.; Aparicio, S.A.J.R.; Behjati, S.; Biankin, A.V.; Bignell, G.R.; Bolli, N.; Borg, A.; Børresen-Dale, A.-L.; et al. Signatures of Mutational Processes in Human Cancer. Nature 2013, 500, 415–421. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Reimand, J.; Kull, M.; Peterson, H.; Hansen, J.; Vilo, J. g: Profiler—A Web-Based Toolset for Functional Profiling of Gene Lists from Large-Scale Experiments. Nucleic Acids Res. 2007, 35, W193–W200. [Google Scholar] [CrossRef]
- Supek, F.; Bošnjak, M.; Škunca, N.; Šmuc, T. REVIGO Summarizes and Visualizes Long Lists of Gene Ontology Terms. PLoS ONE 2011, 6, e21800. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shyr, C.; Tarailo-Graovac, M.; Gottlieb, M.; Lee, J.J.Y.; van Karnebeek, C.; Wasserman, W.W. FLAGS, Frequently Mutated Genes in Public Exomes. BMC Med. Genom. 2014, 7, 64. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stratton, M.R.; Campbell, P.J.; Futreal, P.A. The Cancer Genome. Nature 2009, 458, 719–724. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Giannakouros, P.; Matte, I.; Rancourt, C.; Piché, A. Transformation of NIH3T3 Mouse Fibroblast Cells by MUC16 Mucin (CA125) Is Driven by Its Cytoplasmic Tail. Int. J. Oncol. 2015, 46, 91–98. [Google Scholar] [CrossRef] [Green Version]
- Yamamoto, Y.; Abe, A.; Emi, N. Clarifying the Impact of Polycomb Complex Component Disruption in Human Cancers. Mol. Cancer Res. 2014, 12, 479–484. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, X.; Zhang, G.; Chen, B.; Wang, Y.; Guo, L.; Cao, L.; Ren, C.; Wen, L.; Liao, N. Association between Histone Lysine Methyltransferase KMT2C Mutation and Clinicopathological Factors in Breast Cancer. Biomed. Pharmacother. 2019, 116, 108997. [Google Scholar] [CrossRef] [PubMed]
- Gala, K.; Li, Q.; Sinha, A.; Razavi, P.; Dorso, M.; Sanchez-Vega, F.; Chung, Y.R.; Hendrickson, R.; Hsieh, J.J.; Berger, M.; et al. KMT2C Mediates the Estrogen Dependence of Breast Cancer through Regulation of ERα Enhancer Function. Oncogene 2018, 37, 4692–4710. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bertucci, F.; Ng, C.K.Y.; Patsouris, A.; Droin, N.; Piscuoglio, S.; Carbuccia, N.; Soria, J.C.; Dien, A.T.; Adnani, Y.; Kamal, M.; et al. Genomic Characterization of Metastatic Breast Cancers. Nature 2019, 569, 560–564. [Google Scholar] [CrossRef] [PubMed]
- Niyomnaitham, S.; Parinyanitikul, N.; Roothumnong, E.; Jinda, W.; Samarnthai, N.; Atikankul, T.; Suktitipat, B.; Thongnoppakhun, W.; Limwongse, C.; Pithukpakorn, M. Tumor Mutational Profile of Triple Negative Breast Cancer Patients in Thailand Revealed Distinctive Genetic Alteration in Chromatin Remodeling Gene. PeerJ 2019, 7, e6501. [Google Scholar] [CrossRef] [PubMed]
- Sato, K.; Akimoto, K. Expression Levels of KMT2C and SLC20A1 Identified by Information-Theoretical Analysis Are Powerful Prognostic Biomarkers in Estrogen Receptor-Positive Breast Cancer. Clin. Breast Cancer 2017, 17, e135–e142. [Google Scholar] [CrossRef] [Green Version]
- Rummel, S.K.; Lovejoy, L.; Shriver, C.D.; Ellsworth, R.E. Contribution of Germline Mutations in Cancer Predisposition Genes to Tumor Etiology in Young Women Diagnosed with Invasive Breast Cancer. Breast Cancer Res. Treat. 2017, 164, 593–601. [Google Scholar] [CrossRef] [PubMed]
- Bakhuizen, J.J.; Hogervorst, F.B.; Velthuizen, M.E.; Ruijs, M.W.; van Engelen, K.; van Os, T.A.; Gille, J.J.; Collée, M.; van den Ouweland, A.M.; van Asperen, C.J.; et al. TP53 Germline Mutation Testing in Early-Onset Breast Cancer: Findings from a Nationwide Cohort. Fam. Cancer 2019, 18, 273–280. [Google Scholar] [CrossRef] [PubMed]
- Olivier, M.; Langerød, A.; Carrieri, P.; Bergh, J.; Klaar, S.; Eyfjord, J.; Theillet, C.; Rodriguez, C.; Lidereau, R.; Bièche, I.; et al. The Clinical Value of Somatic TP53 Gene Mutations in 1794 Patients with Breast Cancer. Clin. Cancer Res. 2006, 12, 1157–1167. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Soussi, T.; Wiman, K.G. TP53: An Oncogene in Disguise. Cell Death Differ. 2015, 22, 1239–1249. [Google Scholar] [CrossRef]
- Li, W.; Shao, D.; Li, L.; Wu, M.; Ma, S.; Tan, X.; Zhong, S.; Guo, F.; Wang, Z.; Ye, M. Germline and Somatic Mutations of Multi-Gene Panel in Chinese Patients with Epithelial Ovarian Cancer: A Prospective Cohort Study. J. Ovarian Res. 2019, 12, 80. [Google Scholar] [CrossRef]
- Zhang, Y.; Shi, X.; Zhang, J.; Chen, X.; Zhang, P.; Liu, A.; Zhu, T. A Comprehensive Analysis of Somatic Alterations in Chinese Ovarian Cancer Patients. Sci. Rep. 2021, 11, 387. [Google Scholar] [CrossRef]
- Chen, H.; Chong, W.; Yang, X.; Zhang, Y.; Sang, S.; Li, X.; Lu, M. Age-Related Mutational Signature Negatively Associated with Immune Activity and Survival Outcome in Triple-Negative Breast Cancer. Oncoimmunology 2020, 9, 1788252. [Google Scholar] [CrossRef]
- Mealey, N.E.; O’Sullivan, D.E.; Pader, J.; Ruan, Y.; Wang, E.; Quan, M.L.; Brenner, D.R. Mutational Landscape Differences between Young-Onset and Older-Onset Breast Cancer Patients. BMC Cancer 2020, 20, 212. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Azim, H.A.; Nguyen, B.; Brohée, S.; Zoppoli, G.; Sotiriou, C. Genomic Aberrations in Young and Elderly Breast Cancer Patients. BMC Med. 2015, 13, 266. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Patch, A.-M.; Christie, E.L.; Etemadmoghadam, D.; Garsed, D.W.; George, J.; Fereday, S.; Nones, K.; Cowin, P.; Alsop, K.; Bailey, P.J.; et al. Whole-Genome Characterization of Chemoresistant Ovarian Cancer. Nature 2015, 521, 489–494. [Google Scholar] [CrossRef]
- Ashley, C.W.; Da Cruz Paula, A.; Kumar, R.; Mandelker, D.; Pei, X.; Riaz, N.; Reis-Filho, J.S.; Weigelt, B. Analysis of Mutational Signatures in Primary and Metastatic Endometrial Cancer Reveals Distinct Patterns of DNA Repair Defects and Shifts during Tumor Progression. Gynecol. Oncol. 2019, 152, 11–19. [Google Scholar] [CrossRef] [PubMed]
- Masoodi, T.; Siraj, S.; Siraj, A.K.; Azam, S.; Qadri, Z.; Parvathareddy, S.K.; Tulbah, A.; Al-Dayel, F.; AlHusaini, H.; AlOmar, O.; et al. Genetic Heterogeneity and Evolutionary History of High-Grade Ovarian Carcinoma and Matched Distant Metastases. Br. J. Cancer 2020, 122, 1219–1230. [Google Scholar] [CrossRef] [Green Version]
- Yang, S.Y.C.; Lheureux, S.; Karakasis, K.; Burnier, J.V.; Bruce, J.P.; Clouthier, D.L.; Danesh, A.; Quevedo, R.; Dowar, M.; Hanna, Y.; et al. Landscape of Genomic Alterations in High-Grade Serous Ovarian Cancer from Exceptional Long- and Short-Term Survivors. Genome Med. 2018, 10, 81. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gupta, G.K.; Collier, A.L.; Lee, D.; Hoefer, R.A.; Zheleva, V.; Siewertsz van Reesema, L.L.; Tang-Tan, A.M.; Guye, M.L.; Chang, D.Z.; Winston, J.S.; et al. Perspectives on Triple-Negative Breast Cancer: Current Treatment Strategies, Unmet Needs, and Potential Targets for Future Therapies. Cancers (Basel) 2020, 12, 2392. [Google Scholar] [CrossRef] [PubMed]
- Zhu, S.; Xu, Y.; Wang, L.; Liao, S.; Wang, Y.; Shi, M.; Tu, Y.; Zhou, Y.; Wei, W. Ceramide Kinase Mediates Intrinsic Resistance and Inferior Response to Chemotherapy in Triple-Negative Breast Cancer by Upregulating Ras/ERK and PI3K/Akt Pathways. Cancer Cell Int. 2021, 21, 42. [Google Scholar] [CrossRef]
- Tsang, Y.T.; Deavers, M.T.; Sun, C.C.; Kwan, S.-Y.; Kuo, E.; Malpica, A.; Mok, S.C.; Gershenson, D.M.; Wong, K.-K. KRAS (but Not BRAF) Mutations in Ovarian Serous Borderline Tumour Are Associated with Recurrent Low-Grade Serous Carcinoma. J. Pathol. 2013, 231, 449–456. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Meinhold-Heerlein, I.; Fotopoulou, C.; Harter, P.; Kurzeder, C.; Mustea, A.; Wimberger, P.; Hauptmann, S.; Sehouli, J. The New WHO Classification of Ovarian, Fallopian Tube, and Primary Peritoneal Cancer and Its Clinical Implications. Arch. Gynecol. Obstet. 2016, 293, 695–700. [Google Scholar] [CrossRef] [PubMed]
- Rojas, V.; Hirshfield, K.M.; Ganesan, S.; Rodriguez-Rodriguez, L. Molecular Characterization of Epithelial Ovarian Cancer: Implications for Diagnosis and Treatment. Int. J. Mol. Sci. 2016, 17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhong, F.; Zhu, T.; Pan, X.; Zhang, Y.; Yang, H.; Wang, X.; Hu, J.; Han, H.; Mei, L.; Chen, D.; et al. Comprehensive Genomic Profiling of High-Grade Serous Ovarian Carcinoma from Chinese Patients Identifies Co-Occurring Mutations in the Ras/Raf Pathway with TP53. Cancer Med. 2019, 8, 3928–3935. [Google Scholar] [CrossRef] [Green Version]
- Ma, D.; Jiang, Y.-Z.; Xiao, Y.; Xie, M.-D.; Zhao, S.; Jin, X.; Xu, X.-E.; Shao, Z.-M. Integrated Molecular Profiling of Young and Elderly Patients with Triple-Negative Breast Cancer Indicates Different Biological Bases and Clinical Management Strategies. Cancer 2020, 126, 3209–3218. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.; Zhao, L.; Rojas, C.; Bateman, N.W.; Yao, H.; Lara, O.D.; Celestino, J.; Morgan, M.B.; Nguyen, T.V.; Conrads, K.A.; et al. Molecular Analysis of Clinically Defined Subsets of High-Grade Serous Ovarian Cancer. Cell Rep. 2020, 31, 107502. [Google Scholar] [CrossRef] [PubMed]
- Yamulla, R.J.; Nalubola, S.; Flesken-Nikitin, A.; Nikitin, A.Y.; Schimenti, J.C. Most Commonly Mutated Genes in High-Grade Serous Ovarian Carcinoma Are Nonessential for Ovarian Surface Epithelial Stem Cell Transformation. Cell Rep. 2020, 32, 108086. [Google Scholar] [CrossRef]
- Aine, M.; Boyaci, C.; Hartman, J.; Häkkinen, J.; Mitra, S.; Campos, A.B.; Nimeus, E.; Ehinger, A.; Vallon-Christersson, J.; Borg, Å.; et al. Molecular Analyses of Triple-Negative Breast Cancer in the Young and Elderly. Breast Cancer Res. 2021, 23, 20. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| HGSOC | TNBC | Young Patients: TNBC vs. HGSOC | Elderly Patients: TNBC vs. HGSOC | ||||||
|---|---|---|---|---|---|---|---|---|---|
| Young | Elderly | p-Value | Young | Elderly | p-Value | p-Value | p-Value | ||
| Median C>T substitutions (%) | 37 | 41 | ns | 27 | 35 | 0.00055 | 0.00084 | ns | |
| Median n. of coding variants/sample | 37 | 44 | ns | 50.5 | 61 | ns | 0.014 | 0.046 | |
| Median n. of MS variants/sample | 32 | 37.5 | ns | 42.5 | 53 | ns | 0.011 | 0.046 | |
| Median n. of potentially pathogenic (FS/SS/NS)/possibly pathogenic variants/sample | 14 | 13.5 | ns | 17.5 | 21 | ns | ns | 0.016 | |
| Median n. CGC/sample | 3 | 4 | ns | 3 | 6 | 0.00013 | ns | 0.001 | |
| Median n. CCGD/sample | 4 | 6 | ns | 6 | 7 | 0.044 | ns | 0.025 | |
| Mutational Signatures (%) | 1—Age | 33.3 | 41.7 | ns | 23.3 | - | - | - | - |
| 2—APOBEC (C>T) | - | - | 47.8 | - | - | - | |||
| 3—HR | 42.9 | 25 | 0.011 | 50 | 17.4 | 1.63 × 10−6 | ns | ns | |
| 5—Unknown | 23.8 | 33.3 | ns | - | - | - | - | - | |
| 6—MMR | - | - | - | - | 34.8 | - | - | - | |
| 13—APOBEC (C>G) | - | - | - | 26.7 | - | - | - | - | |
| CGC | CGC combinations (OG and TSG) | 14/21 (66.6%) | 51/60 (85%) | ns | 62/86 (72.1%) | 23/23 (100%) | 0.0032 | ns | ns |
| CGC alone (OG or TSG) | 5/21 (23.8%) | 6/60 (10%) | ns | 12/86 (14%) | 0/23 (0%) | ns | ns | ns | |
| DNA repair potentially/possibly | 4/21 (19%) | 19/60 (32%) | ns | 13/86 (15%) | 12/23 (52%) | 0.0005 | ns | ns | |
| Ras | 3/21 (14.3%) | 9/60 (15%) | ns | 17/86 (19.7%) | 8/23 (34.8%) | ns | ns | ns | |
| HGSOC | TNBC | ||||||||
|---|---|---|---|---|---|---|---|---|---|
| Gene | Role in Cancer | Y (n = 21) | E (n = 60) | p-value | Y (n = 86) | E (n = 23) | p-Value | Y-HGSOC vs. Y-TNBC (p-value) | E-HGSOC vs. E-TNBC (p-value) |
| TP53 | OG/TSG | 14 | 50 | ns | 60 | 17 | ns | ns | ns |
| MUC16 | OG | - | 5 | ns | 9 | 6 | ns | ns | ns |
| CSMD3 | TSG | 2 | 4 | ns | 3 | 2 | ns | ns | ns |
| PIK3CA | OG | - | 1 | ns | 4 | 6 | ns | ns | ns |
| KMT2C | TSG | - | 3 | ns | 1 | 6 | 0.006 | ns | ns |
| NF1 | TSG | 2 | 3 | ns | 4 | - | ns | ns | ns |
| ATM | TSG | - | 3 | ns | 2 | 3 | ns | ns | ns |
| BIRC6 | OG | - | 3 | ns | 3 | 2 | ns | ns | ns |
| LRP1B | TSG | - | 2 | ns | 6 | - | ns | ns | ns |
| MUC4 | OG | - | - | ns | 6 | 1 | ns | ns | ns |
| CLTCL1 | TSG | 1 | 2 | ns | 2 | 1 | ns | ns | ns |
| BRAF | OG | - | 1 | ns | 2 | 2 | ns | ns | ns |
| BRCA2 | TSG | - | 2 | ns | 2 | 1 | ns | ns | ns |
| CREBBP | OG/TSG | - | 3 | ns | 1 | 1 | ns | ns | ns |
| GRM3 | OG | - | 2 | ns | 3 | - | ns | ns | ns |
| MED12 | TSG | - | 1 | ns | 2 | 2 | ns | ns | ns |
| MET | OG | - | 1 | ns | 3 | 1 | ns | ns | ns |
| NOTCH1 | OG/TSG | - | 1 | ns | 2 | 2 | ns | ns | ns |
| RB1 | TSG | 1 | 3 | ns | 1 | - | ns | ns | ns |
| ZFHX3 | TSG | - | 2 | ns | 2 | 1 | ns | ns | ns |
| a—Young HGSOC | ||||
|---|---|---|---|---|
| Sample_ID | OG | TSG | OG/TSG | |
| TCGA-09–1664-01 | KRAS ++ | NF1 - | - | |
| TCGA-13–0792-01 | CTNND2 ? | CIITA ? | TP53 - | |
| TCGA-13–0793-01 | MECOM ? | BAZ1A ? | TP53 -, DAXX - | |
| TCGA-13–0884-01 | ACVR1 ?, TRRAP ? | CSMD3 ?, SLC34A2 ? | BIRC3 *, TP53 - | |
| TCGA-24–1105-01 | - | - | TP53 - | |
| TCGA-24–1416-01 | PIK3CA ++ | - | TP53 - | |
| TCGA-25–1328-01 | - | - | - | |
| TCGA-25–2404-01 | - | ZMYM3 ? | TP53 - | |
| TCGA-25–2408-01 | - | CDKN1B ? | - | |
| TCGA-29–1688-01 | HIP1 | - | TP53 - | |
| TCGA-29–1769-01 | SETDB1 ? | - | RAD21 ? | |
| TCGA-29–2436-01 | - | KAT6B ? | - | |
| TCGA-36–2530-01 | ERBB2 ? | DNMT3A ?, IKBK | - | |
| TCGA-36–2537-01 | - | - | TP53 - | |
| TCGA-36–2538-01 | - | ERCC3 ?, RB1- | TP53 - | |
| TCGA-36–2540-01 | - | - | - | |
| TCGA-59–2363-01 | - | CLTCL1 ?, CSMD3 ?, NF1 - | TP53 - | |
| TCGA-61–1725-01 | JAK3 ? | RAD17 ? | TP53 - | |
| TCGA-61–2008-01 | - | BLM ?, CNTNAP2 ? | TP53 - | |
| TCGA-61–2109-01 | CHD4 ? | FAT1 ? | TP53 - | |
| TCGA-61–2611-02 | ACKR3 ? | - | TP53 - | |
| b—Elderly HGSOC | ||||
| Sample_ID | OG | TSG | OG/TSG | |
| TCGA-04–1331-01 | DDIT3 ? | BRCA2-, KDM5C ?, LATS1 ?, NF1 ? | TBL1XR1 ?, TP53 - | |
| TCGA-04–1337-01 | - | CDH1 ?, FBXW7 -- | ELF4 ?, TP53 - | |
| TCGA-04–1338-01 | MTOR ? | CDH11 ?, FAT1, KMT2C ? | CREBBP -, PABPC1 ?, TP53 - | |
| TCGA-04–1341-01 | CTNND2 ?, GRM3 ?, PDGFRA ?, PREX2 ?, TCF7L2 ? | CDH10 ?, FANCA ?, ATR ? | TP53 - | |
| TCGA-04–1342-01 | KIT ? | LZTR1 ?, SMARCA4 ? | - | |
| TCGA-04–1347-01 | BRAF ?, GRM3 ? | DROSHA ?, FAT1 ? | TP53 - | |
| TCGA-04–1351-01 | - | - | - | |
| TCGA-04–1365-01 | AFF3 ? | ZFHX3 - | TP53 - | |
| TCGA-04–1517-01 | - | - | TP53 - | |
| TCGA-04–1652-01 | - | LRP1B ?, PTPRT ? | TP53 - | |
| TCGA-09–0364-01 | - | CNOT3 ?, CNTNAP2 ? | TP53 - | |
| TCGA-09–1661-01 | DDIT3 ? | PBMR1 ?, TNFAIP3 ? | TP53 - | |
| TCGA-09–1672-01 | KRAS ++ | - | - | |
| TCGA-09–1674-01 | LPP ?, PIK3CB ? | HNF1A -, PALB2, RB1 -, SETD2 ? | TP53 - | |
| TCGA-09–2044-01 | CTNNB1 ?, MET ? | PTPN13 ?, RMI2 ?, TSC1 - | TP53 - | |
| TCGA-10–0933-01 | MUC16 ? | RB1- | RHOA ?, TP53 - | |
| TCGA-10–0938-01 | NUP98 ?, SND1 ? | - | TP53 - | |
| TCGA-13–0755-01 | MUC16 ? | ARID2 ?, FEN1 ?, GRIN2A ?, KMT2C ? | TP53 - | |
| TCGA-13–0802-01 | - | - | - | |
| TCGA-13–0888-01 | CRTC1 ?, MYCN ?, RAP1GDS1 ?, SOX2 ? | BARD1 ?, EBF1 ?, EP300 ?, USP44 ?, ZFHX3 ? | TP53 - | |
| TCGA-13–0889-01 | ERBB2 ? | AMER1 ?, ATRX ? | TP53 - | |
| TCGA-13–1411-01 | - | CLTCL1 ?, DICER1 ?, SMARCB1 ? | TP53 - | |
| TCGA-13–1481-01 | ARHGAP5 ?, GLI1 ?, MUC16 ?, RARA ? | BRCA2 - | TP53 - | |
| TCGA-13–1507-01 | ATF1 ?, SETDB1 ? | MLH1 ?, PTPRT ? | TP53 - | |
| TCGA-20–0991-01 | MACC1 ? | - | TP53 - | |
| TCGA-20–1686-01 | WAS ? | CSMD3 ?, SLC34A2 ? | IRS4 ?, TP53 - | |
| TCGA-23–1116-01 | - | - | TP53 - | |
| TCGA-23–2641-01 | - | - | NOTCH2 ?, TP53 - | |
| TCGA-24–0966-01 | - | LZTR1 ?, N4BP2 ? | CREBBP ?, TET1 ?, TP53 - | |
| TCGA-24–0982-01 | - | SMC1A ? | TP53 - | |
| TCGA-24–1422-01 | CHD4 ?, MUC16 ? | XPC ? | TP53 - | |
| TCGA-24–1552-01 | - | - | TP53 - | |
| TCGA-24–1849-01 | BRD4 ? | GPC5 ?, RB1 - | BTK ?, TP53 - | |
| TCGA-24–2030-01 | - | NDRG1 ? | BIRC6 ?, TP53 - | |
| TCGA-24–2033-01 | AFF4 ?, GNAS ? | DNM2 ?, STAG1 ? | BIRC6 ?, CBLC ?, EZH2 ?, TP53 - | |
| TCGA-24–2261-01 | - | CSMD3 ? | TP53 - | |
| TCGA-25–1325-01 | MUC16 ? | FAT4 ? | TP53 - | |
| TCGA-25–1329-01 | MTOR * | - | TP53 - | |
| TCGA-25–1634-01 | EGFR ? | EIF3E ? | TBX3 - | |
| TCGA-25–2392-01 | FGFR4 ?, SRC ? | CDK12 --, USP44 ? | TP53 - | |
| TCGA-25–2393-01 | - | MED12 ?, RNF43 ? | BCORL1 ? | |
| TCGA-25–2399-01 | CACNA1D ? | ATM -, NCOR1 ? | TP53 - | |
| TCGA-25–2400-01 | WAS ? | LPR1B ? | TP53 - | |
| TCGA-29–1702-01 | KIT | SLC34A2 ? | TP53 - | |
| TCGA-29–1761-01 | - | CNTNAP2 ?, FANCA -, PRMD2, SMC1A ? | TP53 - | |
| TCGA-29–1771-01 | KMT2A ? | NF1 - | TP53 - | |
| TCGA-29–1774-01 | - | DROSHA ?, SIRPA ? | - | |
| TCGA-29–1778-01 | KMT2A ? | - | TP53 - | |
| TCGA-29–2429-01 | PSIP1 ?, ROS1 ? | ATM ?, FLCN - | - | |
| TCGA-31–1950-01 | PAX3 ? | CCD6 ?, CTCTCL1 ? | CREBBP ?, TP53 - | |
| TCGA-36–1575-01 | - | KDM5C ? | TP53 - | |
| TCGA-36–1576-01 | TSHR ? | - | - | |
| TCGA-36–2543-01 | RET, UBR5 ? | NF1 - | TP53 - | |
| TCGA-42–2587-01 | AFF4 ?, FCRL4 ?, POU2AF1 ? | CDC73 ?, WNK2 | BCL9L ?, ESR1 ?, TP53 - | |
| TCGA-59–2352-01 | A1CF ?, SIX1 ?, ZEB1 ? | CCDC6 ?, CNTNAP2 ?, CSMD3 ?, PRDM1 ? | BIRC6 ?, TP53 - | |
| TCGA-61–1730-01 | ALK ?, PLCG1 ? | FAT1 ? | TP53 - | |
| TCGA-61–1741-01 | CHD4 ? | - | DAXX -, TP53 - | |
| TCGA-61–1899-01 | CDH10 ?, PIK3CB ? | CSMD3 ?, KMT2C ? | - | |
| TCGA-61–2012-01 | BCL6 ?, SGK1 ? | PHF6 -, POLG ? | NOTCH1 ?, TBX3 ? | |
| TCGA-OY-A56Q-01 | - | - | TP53 - | |
| c—Young TNBC | ||||
| Sample_ID | OG | TSG | OG/TSG | Putative Drivers |
| BB01_044 | - | - | BIRC6 ?, KMT2D ?, TP53 - | - |
| BB01_074 | BCL9 ?, CCND3 ? | ATM ?, DICER1 ?, LRP1B ? | CREBBP -, TP53 - | - |
| BB01_109 | ETV1 | - | TP53 - | - |
| BB01_126 | RET ? | - | TP53 - | - |
| BR067 | ACK3 ?, ETV4 ?, GRM3 ?, MUC4 ? | - | STAT5B ?, TP53 - | - |
| BR078 | - | - | - | HERC1, HUWE1 |
| BR080 | BRAF ? | NRG1 ? | TP53 - | - |
| BR088 | - | - | - | KIF13B |
| BR091 | - | - | TP53 - | - |
| BR097 | NR4A3 ?, PIK3CA ++, PTPN11 ? | DICER1 --, NF1 - | FOXO3 ?, TP53 - | - |
| BR100 | MET ?, PIK3CB ? | KMT2C ?, LRP1B ? | TP53 - | - |
| BR105 | RANBP2 ?, UBR5 ? | LRP1B ? | TP53 - | - |
| BR108 | MUC16 ? | SMARCA4 - | BIRC6 ?, TP53 - | - |
| BR121 | AFF3 ?, BRAF ? | ATRX ? | - | - |
| BR145 | - | ASXL2 ? | TP53 - | - |
| BR164 | UBR5 ? | - | TP53 - | - |
| BR176 | ERBB2 ?, MUC4 | CLTCL1 ? | TP53 - | - |
| BR200 | PREX2 ?, TRRAP ? | LRIG3 ? | ELF4 ?, TP53 - | - |
| BR255 | CHST11 ?, DNM2 ? | BARD1 ?, ETV6 ?, FHIT ?, LPR1B ?, POLG ?, TET2 | - | - |
| BR301 | MUC4 | CTLCL1 ? | LEF1 ?, TP53 - | - |
| BR313 | USP6 | - | TP53 - | - |
| BR367 | STIL ? | - | TP53 - | - |
| BR393 | - | ZFHX3 ?, ZMYM3 ? | TET1 ?, TP53 - | - |
| BR395 | - | - | TP53 - | - |
| BR-M-045 | CARD11, HLF ?, MUC16 ? | LRP1B ? | TP53- | - |
| BR-V-022 | - | CDH1, LRP1B ? | TP53 - | - |
| BR-V-051 | - | - | TP53 - | - |
| BR-V-070 | - | - | NOTCH1 ? | - |
| PD11326a | - | AXIN2, KAT6B | TP53 - | - |
| PD13627a | - | PTPRC ? | - | - |
| PD18024a | - | - | TP53 - | - |
| PD22036a | - | NF1 - | - | - |
| PD22358a | CDH17 ? | POLE ?, SETD2 ? | - | - |
| PD23554a | FLI1 | BAZ1A ? | KMT2D ?, TP53 - | - |
| PD23563a | IL6ST ? | SPOP ? | TP53 - | - |
| PD23566a | HIST1H3B ?, MET ? | PTPRD - | FES ?, TP53 - | - |
| PD24182a | MUC4 ?, TRIM27 | CDX2 ?, CSMD3 ? | TP53 - | - |
| PD24186a | RAC1 ? | - | - | - |
| PD24191a | CACNA1D ?, MUC16 ?, MUC4 ? | PTEN - | - | - |
| PD24196 | - | FAT4 ?, PTPN13 | TP53 - | - |
| PD24202a | PPM1D ? | NF1 - | FOXO3 ? | - |
| PD24337a | XPO1 ? | CDH11 ?, PTPRD ? | TP53 - | - |
| PD3905a | PIK3CA ++ | - | TP53 - | - |
| PD4005a | - | ERCC5, PTPN6 ? | TBL1XR1 ?, TP53 - | - |
| PD4006a | - | - | - | MYT1, NFKB1 |
| PD4107a | ABL2, POU2AF1 ? | ARHGEF12 ? | NTRK1 ?, TP53 - | - |
| PD4833a | UBR5 ? | - | BCL9L ?, TP53 - | - |
| PD4836a | - | ARID1B ?, LARP4B ? | TP53 - | - |
| PD5930a | - | PHF6 - | RECQL4 ? | - |
| PD5945a | - | - | NFE2L2 ?, TP53 - | - |
| PD6406a | PREX2 ? | - | - | - |
| PD6411a | PREX2 | - | TP53 - | - |
| PD6413a | - | TET2 ? | - | - |
| PD6722a | CSF3R ? | FAZ ?, RB1- | TP53 - | - |
| PD9004a | SSX1, TNC | CDC73 ?, DNMT3A ?, GRIN2A ? | TP53 - | - |
| PD9595a | CSF1R ? | - | BTK ?, TP53 - | - |
| PD9696a | BCL11A ? | WNK2 ? | CUX1 ?, TP53 - | - |
| SA083 | - | ZFHX3 - | - | - |
| SA097 | BRD4 ?, PRDM16 ? | CCDC6 | TP53 - | - |
| SA208 | - | PIK3R1 - | - | - |
| SA220 | FOXP1 ?, NFATC2 ? | ATR ?, ROBO2, SMARCA4 ? | - | - |
| SA231 | - | - | - | MBD2 |
| SA235 | - | BAP1 ? | - | - |
| SA236 | FLT3 ?, MUC16 ? | ARID2 ?, IGF2BP2 ? | TP53 - | - |
| TCGA-A1-A0SP-01 | - | ARID1A ?, ZMYM3 ? | TP53 - | - |
| TCGA-A2-A04P-01 | ERBB3, GRM3 ?, PIK3CA ++, SETDB1 ? | ACVR2A ?, BRCA2 ?, ROBO2 ?, TNFAIP3 ? | IRS4 ?, TP53 - | - |
| TCGA-A2-A0CM-01 | KAT6A ? | CDH11 ?, MGMT ? | - | - |
| TCGA-A2-A3XU-01 | - | - | - | - |
| TCGA-AO-A124–01 | CCR7 ?, CSF3R ?, MAP2K2 ?, MUC16 ? | BRCA2 ?, CDKN2A-, EXT2 ?, FANCG ? | TP53 - | - |
| TCGA-AO-A129–01 | BCL11A ?, IL6ST ?, USP6 ? | DDX3X ?, KAT6B ? | MAP3K13 ?, TP53 - | - |
| TCGA-AO-A12F-01 | - | - | - | TAF1 |
| TCGA-AR-A0TU-01 | AKT3 ?, ETV1 ? | GRIN2A -, NF2 - | - | - |
| TCGA-AR-A0U1–01 | - | BCOR ?, RSPO2 ? | TP53 - | - |
| TCGA-B6-A0IQ-01 | - | SMC1A ? | TP53 - | - |
| TCGA-B6-A0RS-01 | A1CF ?, CHD4 ?, MET ?, MUC16 ? | MED12 ?, MUTYH ?, ZMYM3 ? | TP53 - | - |
| TCGA-B6-A0RT-01 | GRM3 ?, MUC16 ? | - | TP53 - | - |
| TCGA-B6-A0RU-01 | ARHGAP5 ? | - | POLQ ?, TP53 - | - |
| TCGA-B6-A0WX-01 | - | SLC34A2 ?, ZNRF3 ? | EPAS1, TP53 - | - |
| TCGA-BH-A0BL-01 | HLF ? | CSMD3 ?, LZTR1 - | TP53 - | - |
| TCGA-BH-A0E0–01 | ALK ? | - | NOTCH1 ?,RHOA -, TP53 - | - |
| TCGA-D8-A27F-01 | MDM4 | BAP1 -, CSMD3 ? | BIRC6 ?, TP53- | - |
| TCGA-E2-A14N-01 | MAPK1 ?, ROS1 ? | ABI1 ?, PTCH1 ? | TP53 - | - |
| TCGA-E2-A1L7–01 | SGK1 ? | GPC5 ?, PTPRB ? | TP53 - | - |
| TCGA-E9-A3QA-01 | - | NF1-, POT1 | - | - |
| TCGA-OL-A5RW-01 | AFF3, CTNND2 ?, HNRNPA2B1 ?, MUC4, RAF1 ? | ATM, AXIN1 ?, CBLB ?, MED12 - | IRS4 ? | - |
| TCGA-OL-A66I-01 | RANBP2 ? | CYLD ? | TP53 - | - |
| d—Elderly TNBC | ||||
| Sample_ID | OG | TSG | OG/TSG | |
| PD10011a | MPL ? | ASXL1 -, KMT2C - | TP53 - | |
| PD13298a | RET | CSMD3 ?, KMT2C -, PTPRB ? | NOTCH2 ?, TP53 - | |
| PD24333a | BRAF ++, KRAS ++, MUC16 ?, PDGFRA ?, PIK3CA ++, ROS1 ? | KMT2C - | RAD21 ? | |
| PD6047a | - | KMT2C ? | TP53 - | |
| PD7067a | FLI1 ?, TAL1 ? | ATM ?, BRCA1 - | POLQ ?, TP53 - | |
| PD8982a | CALR ?, MUC16 ?, TRRAP ? | ARID1B ?, CDH10 ?, CLTC ?, MED12 ?, SPEN ? | BIRC6 ?, MAP3K1 -, NOTCH1 ?, POLQ ?, TP53 - | |
| PD9575a | BCL3 ?, ERBB3 + | PTEN-, PTPN13 ? | TP53 - | |
| PD9584a | H3F3A ?, HRAS ++ | - | TBX3 - | |
| SA031 | BRAF ?, MET ?, NRAS ? | DDX3X ?, STAG2 ? | BCORL1 | |
| SA052 | - | ATM - | KDM6A -, NOTCH1 - | |
| SA056 | ARGHAP5 ?, ERBB4 ?, PIK3CA ++ | - | TP53 - | |
| SA106 | FCRL4, HIF1A | BRCA2-, CAMTA1, CSMD3 ?, FLCN ?, SPEN ? | BCL9L ?, TP53 - | |
| TCGA-A2-A1G1–01 | - | - | CREBBP -, TP53 -, MAP3K1 ? | |
| TCGA-AC-A2BK-01 | - | MYH9 ?, PTPRD ? | KMT2D -, TP53 - | |
| TCGA-AR-A1AJ-01 | - | PTEN -- | TP53 - | |
| TCGA-BH-A0WA-01 | IL6ST ? | AXIN1 ?, BRCA1 - | TP53 - | |
| TCGA-BH-A18G-01 | ARHGAP5 ?, BCL9 ?, ERBB3 ?, MAML2 ?, MUC16 ?, MUC4, TRIM27 ?, ZEB1 | ARHGAP26 ?, ARHGEF10L ?, ARID1B -, ASXL2 ?, ATM -, ATRX ?, BCOR, CARS ?, CCNB1IP1 ?, CDH10 ?, CLTCL1 ?, CNOT3, CYLD ?, DICER1 ?, EP300 ?, ERCC3 ?, ERCC5 ?, ETV6 ?, IGH2BP2 ?, KDM5C ?, KMT2C ?, MLF1 ?, MYH9 ?, PPP2R1A -, TET2, ZFHX3 ? | FES ?, KDM6A ?, KMT2D ?, MAP3K1 ?, QKI ? | |
| TCGA-BH-A1F0–01 | SGK1 ?, STAT3 ? | ASXL2 ?, GPC5 ?, KMT2C ? | - | |
| TCGA-BH-A1FC-01 | CTNNA2 ?, DDX5 ?, TCF7L2 ?, TEC ? | CLTC ?, NCOR1 ? | TP53 - | |
| TCGA-C8-A12K-01 | - | ATP2B3 ?, IKZF1 ?, STK11 ? | FES ?, IRS4 ?, TP53 - | |
| TCGA-C8-A131–01 | KRAS ++, PIK3CA ++ | NTHL1 ?, PTPRD ?, SETD2 ? | TP53 - | |
| TCGA-D8-A1JK-01 | CTNNA2 ?, KATA ?, PIK3CA ++, STAT3 +, TNC ? | ASXL1 ?, FANCD2 ?, FAT4 ?, NCOR1 ? | TP53 - | |
| TCGA-E2-A1LK-01 | CACNA1D ? | EP300 ?, LARP4B ?, MED12 ? | KDM6A -, TP53 - | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Serio, P.A.d.M.P.; de Lima Pereira, G.F.; Katayama, M.L.H.; Roela, R.A.; Maistro, S.; Folgueira, M.A.A.K. Somatic Mutational Profile of High-Grade Serous Ovarian Carcinoma and Triple-Negative Breast Carcinoma in Young and Elderly Patients: Similarities and Divergences. Cells 2021, 10, 3586. https://doi.org/10.3390/cells10123586
Serio PAdMP, de Lima Pereira GF, Katayama MLH, Roela RA, Maistro S, Folgueira MAAK. Somatic Mutational Profile of High-Grade Serous Ovarian Carcinoma and Triple-Negative Breast Carcinoma in Young and Elderly Patients: Similarities and Divergences. Cells. 2021; 10(12):3586. https://doi.org/10.3390/cells10123586
Chicago/Turabian StyleSerio, Pedro Adolpho de Menezes Pacheco, Gláucia Fernanda de Lima Pereira, Maria Lucia Hirata Katayama, Rosimeire Aparecida Roela, Simone Maistro, and Maria Aparecida Azevedo Koike Folgueira. 2021. "Somatic Mutational Profile of High-Grade Serous Ovarian Carcinoma and Triple-Negative Breast Carcinoma in Young and Elderly Patients: Similarities and Divergences" Cells 10, no. 12: 3586. https://doi.org/10.3390/cells10123586