
Impact of the Main Cardiovascular Risk Factors on Plasma Extracellular Vesicles and Their Influence on the Heart’s Vulnerability to Ischemia-Reperfusion Injury



Abstract
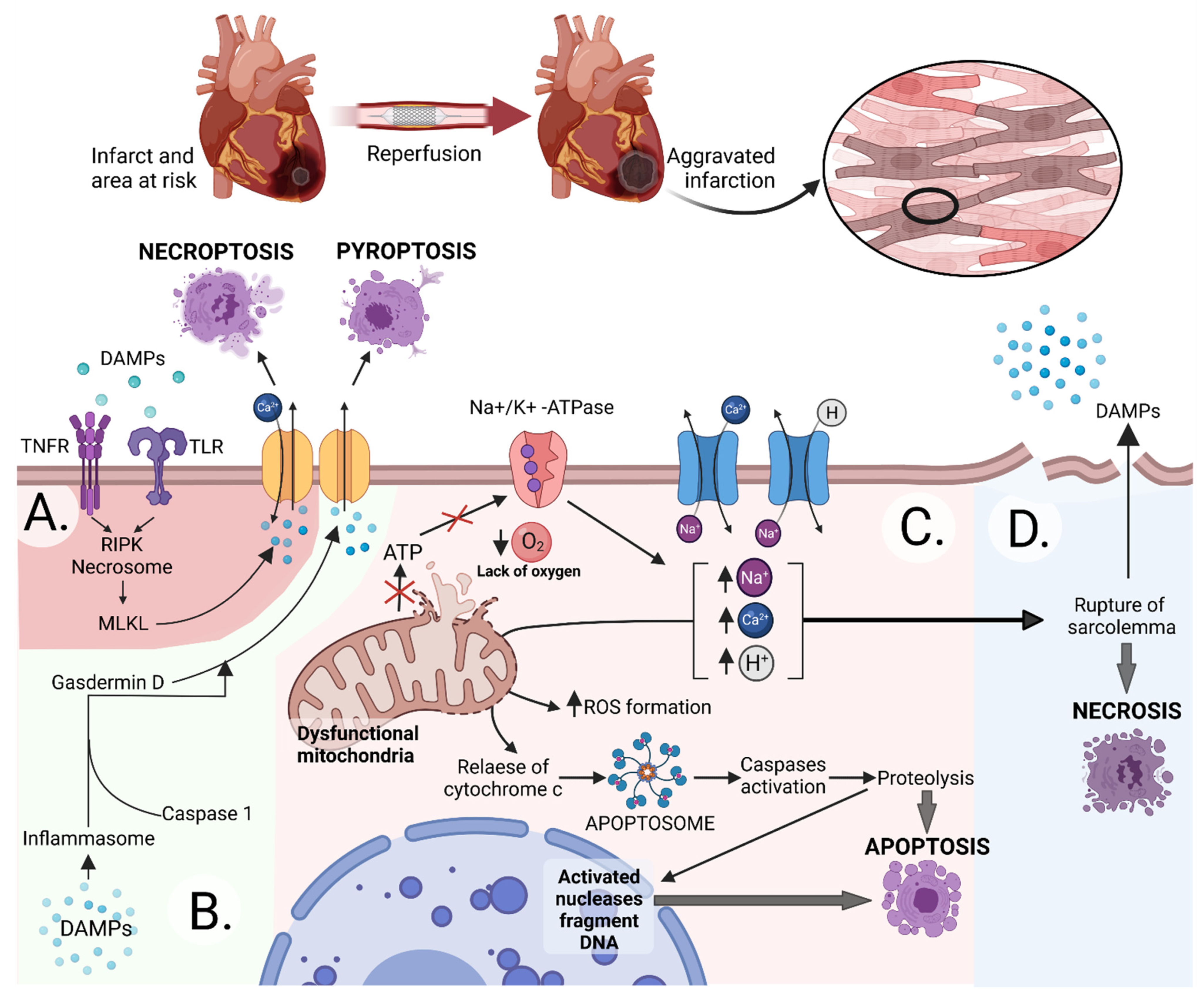
:1. Introduction
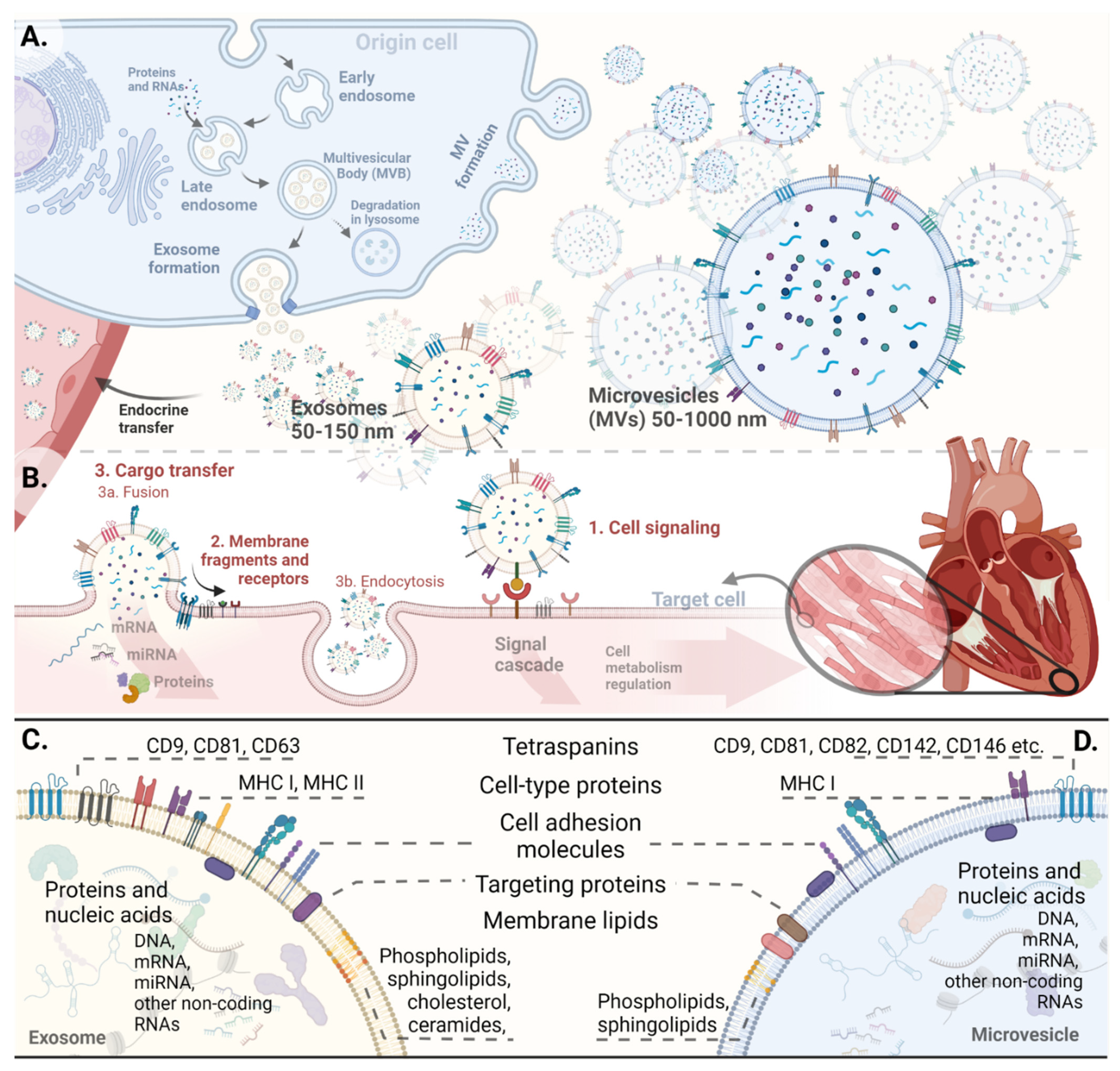
2. Extracellular Vesicles
2.1. Nomenclature and Biogenesis
2.2. EV Characterization
2.3. Uptake and Function
2.4. Vs and MI
3. Cardiovascular Risk Factors
3.1. Diabetes Mellitus and Obesity
3.1.1. DM Type 1
3.1.2. DM Type 2 and Obesity

{kind=link}
{kind=link}
{kind=link}
| Increased | |||||
|---|---|---|---|---|---|
| miRNA | Ob | DM2 | Ref. | Effect | The Potential Regulatory Mechanism in IRI |
| miR-15b | Up | - | [136,173,174,175] | Aggravating | Increases apoptosis via downregulation of Bcl-2, MAPK3, and KCNJ2 [176,177,178]; inhibits the activity of the JAK2-STAT3 pathway, and promotes ROS production [179]. |
| miR-23 | Up | Up | [132,160] | Aggravating | Regulates glutamine metabolism, promotes the transformation of BMSCs into myocardial cells, suppresses expression of Manganese superoxide dismutase, enhances mitophagy, and inhibits connexin 43 expression [161,163,180,181]. |
| miR-34 | Up | Up | [91,136,173,182,183] | Aggravating | Increases apoptosis, infarct size, and suppresses angiogenesis silencing sirtuin-1 and Wnt/β-catenin signaling pathway, suppresses cardiomyocyte proliferation and cardiac recovery post-MI regulating cell cycle activity and death via modulation of its targets, including Bcl2, Cyclin D1, and SIRT1 [90,91,184,185,186,187,188]. |
| miR-122 | Up | Up | [132,136,173,189,190] | Aggravating | Promotes cardiomyocyte apoptosis via regulation of caspase 8 [191], inhibits the expression of Bcl-2, and upregulates the expression of HIF-1α, Bax, and caspase 9 via suppression of FOXP2 [192], downregulates the expression of the AKT/mTOR pathway, and upregulates the JNK/p38MAPK pathway [193]. |
| miR-130 | Up * | Up | [68,105,132,136,173,174,194,195,196] | Aggravating | Promotes worse cardiac function recovery, larger infarct size, and greater cardiomyocyte apoptosis targeting AMPKα1/α2, Birc6, and Ucp3 [68], increases NFκB-mediated inflammation and TGF-β1-mediated fibrosis via inhibition of PPAR-γ expression [197]. |
| miR-155 | Up | Up | [38,102,131,135,136,173] | Aggravating | Increases cardiomyocyte apoptosis in vitro via the downregulation of HIF-1α, RNA-binding protein Quaking, and SIRT1 [153,154,198], enhances the inflammatory response through the activation of the JAK2/STAT1 pathway [155], increases ROS generation during IRI [199]. |
| miR-192 | Up | Up | [132,136,160,173,200,201] | Aggravating | Induces apoptosis targeting FABP3, regulates oxidative stress in IRI [162,202] |
| miR-320 | Up | Up | [37,132,135,200] | Aggravating | Increases infarct size and promotes apoptosis via the inhibition of AKIP1, IGF-1, HSP20, and AKT3 [137,138,139,203]. |
| miR-483 | Up | Up | [132,204,205] | Aggravating | Decreases cell viability and increases apoptosis by targeting the MDM4/p53 pathway [206], promotes apoptosis via the IGF-1 signaling pathway [207,208]. |
| miR-21 | Up * | Up * | [38,91,132,136,173,209,210] | Attenuating | Increases angiogenesis via silencing the cell death-inducing p53 target protein 1 [211], inhibits apoptosis via p38 downregulation and PI3k/Akt activation, decreases autophagy in the myocardial tissue via the AKT/mTOR pathway [91,152,212,213,214,215,216,217]. |
| miR-26b | Up | Up | [132,136,174,196,218] | Attenuating | Reduces inflammation in IRI and improves myocardial remodeling via MAPK pathway activation [219], targets High Mobility Group AT-Hook 2 suppressing MI-induced fibrosis [220,221]. |
| miR-142-3p | Up | Up | [105,136,195,200,201,222] | Attenuating | Inhibits IRI-induced cell apoptosis, autophagy, and fibrosis of the cardiomyocytes by targeting high mobility group box 1 and Rac Family Small GTPase 1 [223,224], improves cardiac function, and attenuates the myocardial inflammatory response targeting IRAK-1 [225]. |
| miR-146 | Up * | Up | [91,135,136,144,182,195,200,209,226,227] | Attenuating | Protects the myocardium from IRI by inhibition of NF-kB and TRAF6/p-p38/caspase-3 signaling pathways, targeting SMAD4, EGR1, and MED1; suppresses inflammatory cytokine production via IRAK-1 and TRAF6 [228,229,230,231,232], inhibits mitochondrial dysfunction in myocardial infarction by targeting cyclophilin D [233], regulates VEGF expression in the IRI heart [234]. |
| miR-150 | Up * | Up | [132,136,173,182,200] | Attenuating | Attenuates apoptosis and improves cardiac function via targeting Bax [235,236], reduces myocardial remodeling by downregulating Thioredoxin Interacting Protein [237]. |
| miR-183 | Up | - | [132] | Attenuating | Reduces infarct size and attenuates apoptosis through repressing voltage-dependent anion channel 1 expression, regulation of NF-κB signaling pathway, and suppression of p27 which activates the PI3K/AKT/FOXO3a signaling pathway [238,239,240]. |
| miR-486 | Up | Up | [136,174,241,242,243] | Attenuating | Inhibits apoptosis and improves cardiac function by suppressing PTEN expression, activating the PI3K/AKT signaling pathway [244,245], and targeting NDRG2 to inactivate the JNK/c-jun and NF-κB signaling pathways [246], promotes cardiac angiogenesis via fibroblastic MMP19-VEGFA cleavage signaling [247]. |
| miR-29 | Up | Up * | [132,136,173,200,201] | Ambiguous | Aggravating: Increasing apoptosis and fibrosis by suppression of Mcl-2 (Bcl-2 family), IGF-1, follistatin-like 1 protein, JAK2/STAT3 pathway, and the SIRT1/AMPK/PGC1α pathway [148,149,150,151,152,248];Attenuating: Inhibition of oxidative stress, apoptosis, and promotes the viability of cardiomyocytes with IRI in vivo model via downregulation of Cyclin T2 [249]. |
| miR-143 | Up | - | [136,189,195] | Ambiguous | Aggravating: Promotes cardiac ischemia-mediated mitochondrial impairment by the inhibition of protein kinase C epsilon [250], inhibits the mitosis of cardiomyocytes [251], promotes fibrosis via targeting sprouty3 [252];Attenuating: Promotes post-MI cell proliferation and reduced cell apoptosis in vitro via cyclooxygenase-2 [253] |
| miR-223 | Up * | Up | [136,173,196,254,255,256] | Ambiguous | Aggravating: Increases cardiomyocyte apoptosis and oxidative stress by targeting KLF1 [257], enhances cardiac fibrosis after MI partially through targeting RASA1 [258], inhibits the angiogenesis of coronary microvascular endothelial cells in the ischemic heart [259]; Attenuating: Protects in vitro cells from hypoxia-induced apoptosis and excessive autophagy via the AKT/mTOR pathway by targeting PARP-1 [260], inhibits I/R-induced cardiac necroptosis at multiple layers [261]. |
| Decreased | |||||
|---|---|---|---|---|---|
| miRNA | Ob | DM2 | Ref. | Effect | The Potential Regulatory Mechanism in IRI |
| miR-17-5p | Down | Down | [136,189,200,201] | Aggravating | Promotes apoptosis induced by ER-stress and oxidative stress injury targeting TSG101 and STAT3 [262,263], increases apoptosis and vascular injury by suppressing the ERK pathway, and via downregulation of Bcl-2 in endothelium cells [264]. |
| miR-24 | - | Down | [77,200,265] | Attenuating | Inhibits apoptosis and excessive O-GlcNAcylation [61,77]. |
| miR-126 | Down | Down | [132,136,142,143,182,200,222,265] | Attenuating | Regulates oxidative stress and apoptosis via downregulation of ERRFI1 expression, and decline of PI3K/AKT pathway activity [140,141,266], decreases angiogenesis [152]. |
| miR-132 | Down | - | [136,189] | Attenuating | Inhibits apoptosis and ROS production via regulation of the TUG1/miR-132-3p/HDAC3 axis and through IL-1β downregulation [267,268,269], protects against oxygen and glucose deprivation via the inhibition of FOXO3a [270], enhances neovascularization [271]. |
| miR-145 | Down | Down | [136,189,272] | Attenuating | Inhibits IRI-induced apoptosis via regulation of the AKT3/mTOR and CaMKII-mediated ASK1 antiapoptotic signaling pathways, ameliorates inflammation by the NF-κB p65 pathway, and the negative regulation of CD40, protects the heart through induction of autophagy [273,274,275,276,277]; absence of miR-145 results in greater infarct thinning and dilatation [278]. |
| miR-206 | Down | - | [136,279] | Attenuating | Reduces IRI-induced apoptosis, targeting protein tyrosine phosphatase 1B, Gadd45β, and ATG3 (activating PI3K/Akt/mTOR pathway); reduces infarct size and improves cardiac function [236,280,281,282]. |
| miR-221 | Down | - | [104,105,136,144,194,241] | Attenuating | Reduces infarct size and prevents IRI-induced apoptosis via the PUMA/ETS-1 pathway and others [145,146]. |
| miR-26a | - | Down | [142] | Ambiguous | Attenuating: Improves viability and inhibits apoptosis via regulation of the PTEN/PI3K/AKT signaling pathway, inhibition of high mobility group box 1 protein expression, inflammatory cell infiltration, and cytokine expression [156,158]. Aggravating: Possible proapoptotic mechanism via targeted regulation of the GSK3β/β-catenin signaling pathway [159] |
| miR-138 | Down | - | [136,175,283] | Ambiguous | Attenuating: Reduces infarct size and myocardial I/R-induced mitochondrial apoptosis by targeting HIF-1α [284], increases the cardiac cells’ viability under hypoxia through targeting PDK1 [285]; Aggravating: Downregulation of miR-138-5p can regulate SIRT1 to inhibit cell pyroptosis and attenuate MI progression [286], miR-138 may mediate inhibition of hypoxia-induced proliferation of endothelial progenitor cells [287]. |
3.2. Smoking
3.3. Total Cholesterol Level
3.4. Systolic Blood Pressure
3.5. Physical Effort
3.6. Sex
3.7. Age
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- Jones, D.S.; Podolsky, S.H.; Greene, J.A. The burden of disease and the changing task of medicine. N. Engl. J. Med. 2012, 366, 2333–2338. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dawber, T.R.; Meadors, G.F.; Moore, F.E., Jr. Epidemiological approaches to heart disease: The framingham study. Am. J. Public Health 1951, 41, 279–286. [Google Scholar] [CrossRef] [PubMed]
- Piepoli, M.F.; Hoes, A.W.; Agewall, S.; Albus, C.; Brotond, C.; Capatano, A.L.; Cooney, M.-T.; Cosyns, B.; Deaton, C.; Graham, I.; et al. European guidelines on cardiovascular disease prevention in clinical practice: The sixth joint task force of the european society of cardiology and other societies on cardiovascular disease prevention in clinical practice (constituted by representatives of 10 societies and by invited experts)developed with the special contribution of the european association for cardiovascular prevention & rehabilitation (EACPR). Eur. Heart J. 2016, 37, 2315–2381. [Google Scholar] [CrossRef] [PubMed]
- Pooling Project Research Group. Relationship of blood pressure, serum cholesterol, smoking habit, relative weight and ECG abnormalities to incidence of major coronary events: Final report of the pooling project. J. Chronic Dis. 1978, 31, 201–306. [Google Scholar] [CrossRef]
- Morris, J.; Heady, J.; Raffle, P.; Roberts, C.; Parks, J. Coronary heart-disease and physical activity of work. Lancet 1953, 262, 1053–1057. [Google Scholar] [CrossRef] [Green Version]
- Wilson, P.W.; D’Agostino, R.B.; Sullivan, L.; Parise, H.; Kannel, W.B. Overweight and obesity as determinants of cardiovascular risk: The Framingham experience. Arch. Intern. Med. 2002, 162, 1867–1872. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Goldschmid, M.G.; Barrett-Connor, E.; Edelstein, S.L.; Wingard, D.L.; Cohn, B.A.; Herman, W.H. Dyslipidemia and ischemic heart disease mortality among men and women with diabetes. Circulation 1994, 89, 991–997. [Google Scholar] [CrossRef] [Green Version]
- Ibanez, B.; James, S.; Agewall, S.; Antunes, M.J.; Bucciarelli-Ducci, C.; Bueno, H.; Caforio, A.L.P.; Crea, F.; Goudevenos, J.A.; Halvorsen, S.; et al. 2017 ESC Guidelines for the management of acute myocardial infarction in patients presenting with ST-segment elevation: The Task Force for the management of acute myocardial infarction in patients presenting with ST-segment elevation of the European Society of Cardiology (ESC). Eur. Heart J. 2018, 39, 119–177. [Google Scholar] [CrossRef] [Green Version]
- Hofmann, R.; Witt, N.; Lagerqvist, B.; Jernberg, T.; Lindahl, B.; Erlinge, D.; Herlitz, J.; Alfredsson, J.; Linder, R.; Omerovic, E.; et al. Oxygen therapy in ST-elevation myocardial infarction. Eur. Heart J. 2018, 39, 2730–2739. [Google Scholar] [CrossRef]
- Abe, H.; Sakurai, A.; Ono, H.; Hayashi, S.; Yoshimoto, S.; Ochi, A.; Ueda, S.; Nishimura, K.; Shibata, E.; Tamaki, M.; et al. Urinary exosomal mRNA of WT1 as diagnostic and prognostic biomarker for diabetic nephropathy. J. Med. Investig. 2018, 65, 208–215. [Google Scholar] [CrossRef] [Green Version]
- Hausenloy, D.; Botker, H.E.; Engstrom, T.; Erlinge, D.; Heusch, G.; Ibanez, B.; Kloner, R.A.; Ovize, M.; Yellon, D.; Garcia-Dorado, D. Targeting reperfusion injury in patients with ST-segment elevation myocardial infarction: Trials and tribulations. Eur. Heart J. 2017, 38, 935–941. [Google Scholar] [CrossRef] [Green Version]
- Heusch, G. Myocardial ischaemia–reperfusion injury and cardioprotection in perspective. Nat. Rev. Cardiol. 2020, 17, 773–789. [Google Scholar] [CrossRef]
- El Andaloussi, S.; Mäger, I.; Breakefield, X.O.; Wood, M.J.A. Extracellular vesicles: Biology and emerging therapeutic opportunities. Nat. Rev. Drug Discov. 2013, 12, 347–357. [Google Scholar] [CrossRef]
- Ludwig, A.-K.; Giebel, B. Exosomes: Small vesicles participating in intercellular communication. Int. J. Biochem. Cell Biol. 2012, 44, 11–15. [Google Scholar] [CrossRef] [PubMed]
- Oerlemans, M.I.F.J.; Liu, J.; Arslan, F.; Ouden, K.D.; van Middelaar, B.J.; Doevendans, P.A.; Sluijter, J.P.G. Inhibition of RIP1-dependent necrosis prevents adverse cardiac remodeling after myocardial ischemia-reperfusion in vivo. Basic Res. Cardiol. 2012, 107, 1–13. [Google Scholar] [CrossRef]
- Tani, M.; Neely, J.R. Role of intracellular Na+ in Ca2+ overload and depressed recovery of ventricular function of reperfused ischemic rat hearts. Possible involvement of H+-Na+ and Na+-Ca2+ exchange. Circ. Res. 1989, 65, 1045–1056. [Google Scholar] [CrossRef] [Green Version]
- Liebert, A.; Krause, A.; Goonetilleke, N.; Bicknell, B.; Kiat, H. A Role for photobiomodulation in the prevention of myocardial ischemic reperfusion injury: A systematic review and potential molecular mechanisms. Sci. Rep. 2017, 7, 42386. [Google Scholar] [CrossRef] [Green Version]
- Bernardi, P.; Rasola, A.; Forte, M.; Lippe, G. The mitochondrial permeability transition pore: Channel formation by f-atp synthase, integration in signal transduction, and role in pathophysiology. Physiol. Rev. 2015, 95, 1111–1155. [Google Scholar] [CrossRef]
- Bernardi, P.; Di Lisa, F. The mitochondrial permeability transition pore: Molecular nature and role as a target in cardioprotection. J. Mol. Cell. Cardiol. 2015, 78, 100–106. [Google Scholar] [CrossRef] [PubMed]
- Doyle, L.M.; Wang, M.Z. Overview of extracellular vesicles, their origin, composition, purpose, and methods for exosome isolation and analysis. Cells 2019, 8, 727. [Google Scholar] [CrossRef] [Green Version]
- Raposo, G.; Stoorvogel, W. Extracellular vesicles: Exosomes, microvesicles, and friends. J. Cell Biol. 2013, 200, 373–383. [Google Scholar] [CrossRef] [Green Version]
- Ostrowski, M.; Carmo, N.B.; Krumeich, S.; Fanget, I.; Raposo, G.; Savina, A.; Moita, C.F.; Schauer, K.; Hume, A.N.; Freitas, R.P.; et al. Rab27a and Rab27b control different steps of the exosome secretion pathway. Nat. Cell Biol. 2010, 12, 19–30. [Google Scholar] [CrossRef] [Green Version]
- Wollert, T.; Hurley, J.H. Molecular mechanism of multivesicular body biogenesis by ESCRT complexes. Nature 2010, 464, 864–869. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Van Niel, G.; D’Angelo, G.; Raposo, G. Shedding light on the cell biology of extracellular vesicles. Nat. Rev. Mol. Cell Biol. 2018, 19, 213–228. [Google Scholar] [CrossRef]
- Record, M.; Carayon, K.; Poirot, M.; Silvente-Poirot, S. Exosomes as new vesicular lipid transporters involved in cell–cell communication and various pathophysiologies. Biochim. Biophys. Acta Mol. Cell Biol. Lipids 2014, 1841, 108–120. [Google Scholar] [CrossRef]
- Zaborowski, M.P.; Balaj, L.; Breakefield, X.O.; Lai, C.P. Extracellular vesicles: Composition, biological relevance, and methods of study. Bioscience 2015, 65, 783–797. [Google Scholar] [CrossRef] [Green Version]
- Valadi, H.; Ekstrom, K.; Bossios, A.; Sjöstrand, M.; Lee, J.J.; Lötvall, J.O. Exosome-mediated transfer of mRNAs and microRNAs is a novel mechanism of genetic exchange between cells. Nat. Cell Biol. 2007, 9, 654–659. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nolte-’t Hoen, E.N.; Buermans, H.P.J.; Waasdorp, M.; Stoorvogel, W.; Wauben, M.H.M.; ’t Hoen, P.A. Deep sequencing of RNA from immune cell-derived vesicles uncovers the selective incorporation of small non-coding RNA biotypes with potential regulatory functions. Nucleic Acids Res. 2012, 40, 9272–9285. [Google Scholar] [CrossRef] [Green Version]
- Ratajczak, J.; Miękus, K.; Kucia, M.; Zhang, J.; Reca, R.; Dvorak, P.; Ratajczak, M.Z. Embryonic stem cell-derived microvesicles reprogram hematopoietic progenitors: Evidence for horizontal transfer of mRNA and protein delivery. Leukemia 2006, 20, 847–856. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, L.; Yang, B.-F.; Ai, J. MicroRNA transport: A new way in cell communication. J. Cell. Physiol. 2013, 228, 1713–1719. [Google Scholar] [CrossRef]
- Villarroya-Beltri, C.; Gutierrez-Vazquez, C.; Sanchez-Cabo, F.; Pérez-Hernández, D.; Vázquez, J.; Martin-Cofreces, N.; Martinez-Herrera, D.J.; Pascual-Montano, A.; Mittelbrunn, M.; Sánchez-Madrid, F. Sumoylated hnRNPA2B1 controls the sorting of miRNAs into exosomes through binding to specific motifs. Nat. Commun. 2013, 4, 2980. [Google Scholar] [CrossRef] [Green Version]
- Ibrahim, A.; Marbán, E. Exosomes: Fundamental biology and roles in cardiovascular physiology. Annu. Rev. Physiol. 2016, 78, 67–83. [Google Scholar] [CrossRef] [Green Version]
- Groot, M.; Lee, H. Sorting mechanisms for microRNAs into extracellular vesicles and their associated diseases. Cells 2020, 9, 1044. [Google Scholar] [CrossRef] [PubMed]
- Maas, S.L.N.; Breakefield, X.O.; Weaver, A.M. Extracellular vesicles: Unique intercellular delivery vehicles. Trends Cell Biol. 2017, 27, 172–188. [Google Scholar] [CrossRef] [Green Version]
- Gartz, M.; Strande, J.L. Examining the paracrine effects of exosomes in cardiovascular disease and repair. J. Am. Heart Assoc. 2018, 7, e007954. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Akbar, N.; Azzimato, V.; Choudhury, R.P.; Aouadi, M. Extracellular vesicles in metabolic disease. Diabetologia 2019, 62, 2179–2187. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ferrante, S.C.; Nadler, E.P.; Pillai, D.K.; Hubal, M.; Wang, Z.; Wang, J.M.; Gordish-Dressman, H.; Koeck, E.; Sevilla, S.; Wiles, A.A.; et al. Adipocyte-derived exosomal miRNAs: A novel mechanism for obesity-related disease. Pediatr. Res. 2015, 77, 447–454. [Google Scholar] [CrossRef]
- Eguchi, A.; Lazic, M.; Armando, A.M.; Phillips, S.A.; Katebian, R.; Maraka, S.; Quehenberger, O.; Sears, D.D.; Feldstein, A.E. Circulating adipocyte-derived extracellular vesicles are novel markers of metabolic stress. J. Mol. Med. 2016, 94, 1241–1253. [Google Scholar] [CrossRef]
- Shah, R.; Patel, T.; Freedman, J.E. Circulating extracellular vesicles in human disease. N. Engl. J. Med. 2018, 379, 958–966. [Google Scholar] [CrossRef] [PubMed]
- Mathieu, M.; Martin-Jaular, L.; Lavieu, G.; Théry, C. Specificities of secretion and uptake of exosomes and other extracellular vesicles for cell-to-cell communication. Nat. Cell Biol. 2019, 21, 9–17. [Google Scholar] [CrossRef] [PubMed]
- Yáñez-Mó, M.; Siljander, P.R.-M.; Andreu, Z.; Zavec, A.B.; Borràs, F.E.; Buzas, E.I.; Buzas, K.; Casal, E.; Cappello, F.; Carvalho, J.; et al. Biological properties of extracellular vesicles and their physiological functions. J. Extracell. Vesicles 2015, 4, 27066. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Raposo, G.; Nijman, H.W.; Stoorvogel, W.; Liejendekker, R.; Harding, C.V.; Melief, C.J.; Geuze, H.J. B lymphocytes secrete antigen-presenting vesicles. J. Exp. Med. 1996, 183, 1161–1172. [Google Scholar] [CrossRef]
- Zitvogel, L.; Regnault, A.; Lozier, A.; Wolfers, J.; Flament, C.; Tenza, D.; Ricciardi-Castagnoli, P.; Raposo, G.; Amigorena, S. Eradication of established murine tumors using a novel cell-free vaccine: Dendritic cell derived exosomes. Nat. Med. 1998, 4, 594–600. [Google Scholar] [CrossRef]
- Bellingham, S.A.; Guo, B.B.; Coleman, B.M.; Hill, A.F. Exosomes: Vehicles for the transfer of toxic proteins associated with neurodegenerative diseases? Front. Physiol. 2012, 3, 124. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, Y.; El Andaloussi, S.; Wood, M.J. Exosomes and microvesicles: Extracellular vesicles for genetic information transfer and gene therapy. Hum. Mol. Genet. 2012, 21, R125–R134. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, J.; Cui, X.; Guo, J.; Cao, C.; Zhang, Z.; Wang, B.; Zhang, L.; Shen, D.; Lim, K.; Tang, J.; et al. Small but significant: Insights and new perspectives of exosomes in cardiovascular disease. J. Cell. Mol. Med. 2020, 24, 8291–8303. [Google Scholar] [CrossRef] [PubMed]
- Latifkar, A.; Hur, Y.H.; Sanchez, J.C.; Cerione, R.A.; Antonyak, M.A. New insights into extracellular vesicle biogenesis and function. J. Cell Sci. 2019, 132, jcs222406. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Arslan, F.; Lai, R.C.; Smeets, M.B.; Akeroyd, L.; Choo, A.; Aguor, E.N.E.; Timmers, L.; Van Rijen, H.V.; Doevendans, P.A.; Pasterkamp, G.; et al. Mesenchymal stem cell-derived exosomes increase ATP levels, decrease oxidative stress and activate PI3K/Akt pathway to enhance myocardial viability and prevent adverse remodeling after myocardial ischemia/reperfusion injury. Stem Cell Res. 2013, 10, 301–312. [Google Scholar] [CrossRef] [Green Version]
- Teng, X.; Chen, L.; Chen, W.; Yang, J.; Yang, Z.; Shen, Z. Mesenchymal stem cell-derived exosomes improve the microenvironment of infarcted myocardium contributing to angiogenesis and anti-inflammation. Cell. Physiol. Biochem. 2015, 37, 2415–2424. [Google Scholar] [CrossRef] [PubMed]
- Qiu, G.; Zheng, G.; Ge, M.; Wang, J.; Huang, R.; Shu, Q.; Xu, J. Mesenchymal stem cell-derived extracellular vesicles affect disease outcomes via transfer of microRNAs. Stem Cell Res. Ther. 2018, 9, 320. [Google Scholar] [CrossRef]
- Lai, R.C.; Arslan, F.; Lee, M.M.; Sze, N.S.K.; Choo, A.; Chen, T.S.; Salto-Tellez, M.; Timmers, L.; Lee, C.N.; El Oakley, R.M.; et al. Exosome secreted by MSC reduces myocardial ischemia/reperfusion injury. Stem Cell Res. 2010, 4, 214–222. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ibrahim, A.G.-E.; Cheng, K.; Marbán, E. Exosomes as critical agents of cardiac regeneration triggered by cell therapy. Stem Cell Rep. 2014, 2, 606–619. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.; Zhang, L.; Li, Y.; Chen, L.; Wang, X.; Guo, W.; Zhang, X.; Qin, G.; He, S.-H.; Zimmerman, A.; et al. Exosomes/microvesicles from induced pluripotent stem cells deliver cardioprotective miRNAs and prevent cardiomyocyte apoptosis in the ischemic myocardium. Int. J. Cardiol. 2015, 192, 61–69. [Google Scholar] [CrossRef] [Green Version]
- Miyahara, Y.; Nagaya, N.; Kataoka, M.; Yanagawa, B.; Tanaka, K.; Hao, H.; Ishino, K.; Ishida, H.; Shimizu, T.; Kangawa, K.; et al. Monolayered mesenchymal stem cells repair scarred myocardium after myocardial infarction. Nat. Med. 2006, 12, 459–465. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, G.H.; Xu, J.; Yang, Y.J. Exosomes: Promising sacks for treating ischemic heart disease? Am. J. Physiol. Heart Circ. Physiol. 2017, 313, H508–H523. [Google Scholar] [CrossRef] [PubMed]
- Sluijter, J.P.G.; Verhage, V.; Deddens, J.C.; Akker, F.V.D.; Doevendans, P.A. Microvesicles and exosomes for intracardiac communication. Cardiovasc. Res. 2014, 102, 302–311. [Google Scholar] [CrossRef] [Green Version]
- Yu, X.; Deng, L.; Wang, D.; Li, N.; Chen, X.; Cheng, X.; Yuan, J.; Gao, X.; Liao, M.; Wang, M.; et al. Mechanism of TNF-alpha autocrine effects in hypoxic cardiomyocytes: Initiated by hypoxia inducible factor 1alpha, presented by exosomes. J. Mol. Cell. Cardiol. 2012, 53, 848–857. [Google Scholar] [CrossRef] [PubMed]
- Bang, C.; Batkai, S.; Dangwal, S.; Gupta, S.K.; Foinquinos, A.; Holzmann, A.; Just, A.; Remke, J.; Zimmer, K.; Zeug, A.; et al. Cardiac fibroblast–derived microRNA passenger strand-enriched exosomes mediate cardiomyocyte hypertrophy. J. Clin. Investig. 2014, 124, 2136–2146. [Google Scholar] [CrossRef]
- Vicencio, J.M.; Yellon, D.M.; Sivaraman, V.; Das, D.; Boi-Doku, C.; Arjun, S.; Zheng, Y.; Riquelme, J.A.; Kearney, J.; Sharma, V.; et al. Plasma exosomes protect the myocardium from ischemia-reperfusion injury. J. Am. Coll. Cardiol. 2015, 65, 1525–1536. [Google Scholar] [CrossRef] [Green Version]
- Giricz, Z.; Varga, Z.V.; Baranyai, T.; Sipos, P.; Pálóczi, K.; Kittel, Á.; Buzás, E.I.; Ferdinandy, P. Cardioprotection by remote ischemic preconditioning of the rat heart is mediated by extracellular vesicles. J. Mol. Cell. Cardiol. 2014, 68, 75–78. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Minghua, W.; Zhijian, G.; Chahua, H.; Qiang, L.; Minxuan, X.; Luqiao, W.; Weifang, Z.; Peng, L.; Biming, Z.; Lingling, Y.; et al. Plasma exosomes induced by remote ischaemic preconditioning attenuate myocardial ischaemia/reperfusion injury by transferring miR-24. Cell Death Dis. 2018, 9, 320. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Zhang, X. Ischaemic preconditioning-induced serum exosomes protect against myocardial ischaemia/reperfusion injury in rats by activating the PI3K/AKT signalling pathway. Cell Biochem. Funct. 2020, 39, 287–295. [Google Scholar] [CrossRef] [PubMed]
- Chen, Q.; Huang, M.; Wu, J.; Jiang, Q.; Zheng, X. Exosomes isolated from the plasma of remote ischemic conditioning rats improved cardiac function and angiogenesis after myocardial infarction through targeting Hsp70. Aging 2020, 12, 3682–3693. [Google Scholar] [CrossRef] [PubMed]
- Haller, P.M.; Jäger, B.; Piackova, E.; Sztulman, L.; Wegberger, C.; Wojta, J.; Gyöngyösi, M.; Kiss, A.; Podesser, B.K.; Spittler, A.; et al. Changes in circulating extracellular vesicles in patients with st-elevation myocardial infarction and potential effects of remote ischemic conditioning—A randomized controlled trial. Biomedicines 2020, 8, 218. [Google Scholar] [CrossRef] [PubMed]
- Frey, U.H.; Klaassen, M.; Ochsenfarth, C.; Murke, F.; Thielmann, M.; Kottenberg, E.; Kleinbongard, P.; Klenke, S.; Engler, A.; Heusch, G.; et al. Remote ischaemic preconditioning increases serum extracellular vesicle concentrations with altered micro-RNA signature in CABG patients. Acta Anaesthesiol. Scand. 2019, 63, 483–492. [Google Scholar] [CrossRef]
- Abel, F.; Murke, F.; Gaida, M.; Garnier, N.; Ochsenfarth, C.; Theiss, C.; Thielmann, M.; Kleinbongard, P.; Giebel, B.; Peters, J.; et al. Extracellular vesicles isolated from patients undergoing remote ischemic preconditioning decrease hypoxia-evoked apoptosis of cardiomyoblasts after isoflurane but not propofol exposure. PLoS ONE 2020, 15, e0228948. [Google Scholar] [CrossRef] [PubMed]
- Davidson, S.M.; Riquelme, J.A.; Takov, K.; Vicencio, J.M.; Boi-Doku, C.; Khoo, V.; Doreth, C.; Radenkovic, D.; Lavandero, S.; Yellon, D.M. Cardioprotection mediated by exosomes is impaired in the setting of type II diabetes but can be rescued by the use of non-diabetic exosomes in vitro. J. Cell. Mol. Med. 2018, 22, 141–151. [Google Scholar] [CrossRef]
- Gan, L.; Xie, D.; Liu, J.; Lau, W.B.; Christopher, T.A.; Lopez, B.; Zhang, L.; Gao, E.; Koch, W.; Ma, X.-L.; et al. Small extracellular microvesicles mediated pathological communications between dysfunctional adipocytes and cardiomyocytes as a novel mechanism exacerbating ischemia/reperfusion injury in diabetic mice. Circulation 2020, 141, 968–983. [Google Scholar] [CrossRef] [PubMed]
- Wider, J.; Undyala, V.V.R.; Whittaker, P.; Woods, J.; Chen, X.; Przyklenk, K. Remote ischemic preconditioning fails to reduce infarct size in the Zucker fatty rat model of type-2 diabetes: Role of defective humoral communication. Basic Res. Cardiol. 2018, 113, 16. [Google Scholar] [CrossRef] [PubMed]
- Davidson, S.M.; Takov, K.; Yellon, D. Exosomes and cardiovascular protection. Cardiovasc. Drugs Ther. 2017, 31, 77–86. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Egom, E.E.; Mamas, M.A.; Clark, A.L. The potential role of sphingolipid-mediated cell signaling in the interaction between hyperglycemia, acute myocardial infarction and heart failure. Expert Opin. Ther. Targets 2012, 16, 791–800. [Google Scholar] [CrossRef] [PubMed]
- Ricci, C.; Jong, C.J.; Schaffer, S.W. Proapoptotic and antiapoptotic effects of hyperglycemia: Role of insulin signaling. Can. J. Physiol. Pharmacol. 2008, 86, 166–172. [Google Scholar] [CrossRef]
- Miki, T.; Itoh, T.; Sunaga, D.; Miura, T. Effects of diabetes on myocardial infarct size and cardioprotection by preconditioning and postconditioning. Cardiovasc. Diabetol. 2012, 11, 67. [Google Scholar] [CrossRef] [Green Version]
- Donahoe, S.M.; Stewart, G.C.; McCabe, C.H.; Mohanavelu, S.; Murphy, S.A.; Cannon, C.P.; Antman, E.M. Diabetes and mortality following acute coronary syndromes. JAMA 2007, 298, 765–775. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xue, R.; Lei, S.; Xia, Z.-Y.; Wu, Y.; Meng, Q.; Zhan, L.; Su, W.; Liu, H.; Xu, J.; Liu, Z.; et al. Selective inhibition of PTEN preserves ischaemic post-conditioning cardioprotection in STZ-induced Type 1 diabetic rats: Role of the PI3K/Akt and JAK2/STAT3 pathways. Clin. Sci. 2016, 130, 377–392. [Google Scholar] [CrossRef]
- Przyklenk, K.; Maynard, M.; Greiner, D.L.; Whittaker, P. Cardioprotection with postconditioning: Loss of efficacy in murine models of type-2 and type-1 diabetes. Antioxid. Redox Signal. 2011, 14, 781–790. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, D.; Hu, X.; Lee, S.H.; Chen, F.; Jiang, K.; Tu, Z.; Liu, Z.; Du, J.; Wang, L.; Yin, C.; et al. Diabetes exacerbates myocardial ischemia/reperfusion injury by down-regulation of microRNA and up-regulation of o-glcnacylation. JACC Basic Transl. Sci. 2018, 3, 350–362. [Google Scholar] [CrossRef]
- Negi, S.; Rutman, A.K.; Paraskevas, S. Extracellular vesicles in type 1 diabetes: Messengers and regulators. Curr. Diabetes Rep. 2019, 19, 69. [Google Scholar] [CrossRef] [PubMed]
- Cianciaruso, C.; Phelps, E.A.; Pasquier, M.; Hamelin, R.; Demurtas, D.; Ahmed, M.A.; Piemonti, L.; Hirosue, S.; Swartz, M.A.; De Palma, M.; et al. Primary human and rat β-cells release the intracellular autoantigens gad65, ia-2, and proinsulin in exosomes together with cytokine-induced enhancers of immunity. Diabetes 2017, 66, 460–473. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Garcia-Contreras, M.; Brooks, R.W.; Boccuzzi, L.; Robbins, P.D.; Ricordi, C. Exosomes as biomarkers and therapeutic tools for type 1 diabetes mellitus. Eur. Rev. Med. Pharmacol. Sci. 2017, 21, 2940–2956. [Google Scholar] [PubMed]
- Barutta, F.; Tricarico, M.; Corbelli, A.; Annaratone, L.; Pinach, S.; Grimaldi, S.; Bruno, G.; Cimino, D.; Taverna, D.; Deregibus, M.C.; et al. Urinary exosomal microRNAs in incipient diabetic nephropathy. PLoS ONE 2013, 8, e73798. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Florijn, B.W.; Duijs, J.M.; Levels, J.H.; Dallinga-Thie, G.M.; Wang, Y.; Boing, A.N.; Yuana, Y.; Stam, W.; Limpens, R.W.; Au, Y.W.; et al. Diabetic nephropathy alters the distribution of circulating angiogenic micrornas among extracellular vesicles, HDL, and Ago-2. Diabetes 2019, 68, 2287–2300. [Google Scholar] [CrossRef]
- Bergen, K.; Mobarrez, F.; Jörneskog, G.; Wallén, H.; Tehrani, S. Phosphatidylserine expressing microvesicles in relation to microvascular complications in type 1 diabetes. Thromb. Res. 2018, 172, 158–164. [Google Scholar] [CrossRef]
- Zahran, A.M.; Mohamed, I.L.; El Asheer, O.M.; Tamer, D.M.; Abo-Elela, M.G.M.; Abdel-Rahim, M.H.; El-Badawy, O.H.B.; Elsayh, K.I. Circulating endothelial cells, circulating endothelial progenitor cells, and circulating microparticles in type 1 diabetes mellitus. Clin. Appl. Thromb. Hemost. 2019, 25, 1076029618825311. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, M.; Wang, Y.; Zhu, Q.; Zhao, J.; Wang, Y.; Shang, M.; Liu, M.; Wu, Y.; Song, J.; Liu, Y. Protective effects of circulating microvesicles derived from ischemic preconditioning on myocardial ischemia/reperfusion injury in rats by inhibiting endoplasmic reticulum stress. Apoptosis 2018, 23, 436–448. [Google Scholar] [CrossRef]
- Chiva-Blanch, G.; Suades, R.; Padro, T.; Vilahur, G.; Pena, E.; Ybarra, J.; Pou, J.M.; Badimon, L. Microparticle shedding by erythrocytes, monocytes and vascular smooth muscular cells is reduced by aspirin in diabetic patients. Rev. Esp. Cardiol. 2016, 69, 672–680. [Google Scholar] [CrossRef] [PubMed]
- Chang, W.; Wang, J. Exosomes and their noncoding RNA cargo are emerging as new modulators for diabetes mellitus. Cells 2019, 8, 853. [Google Scholar] [CrossRef] [Green Version]
- Garcia-Contreras, M.; Shah, S.H.; Tamayo, A.; Robbins, P.; Golberg, R.B.; Mendez, A.J.; Ricordi, C. Plasma-derived exosome characterization reveals a distinct microRNA signature in long duration Type 1 diabetes. Sci. Rep. 2017, 7, 5998. [Google Scholar] [CrossRef]
- Costantino, S.; Paneni, F.; Lüscher, T.F.; Cosentino, F. MicroRNA profiling unveils hyperglycaemic memory in the diabetic heart. Eur. Heart J. 2016, 37, 572–576. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fu, B.-C.; Lang, J.-L.; Zhang, D.-Y.; Sun, L.; Chen, W.; Liu, W.; Liu, K.-Y.; Ma, C.-Y.; Jiang, S.-L.; Li, R.-K.; et al. Suppression of miR-34a expression in the myocardium protects against ischemia–reperfusion injury through SIRT1 protective pathway. Stem Cells Dev. 2017, 26, 1270–1282. [Google Scholar] [CrossRef]
- Dehaini, H.; Awada, H.; El-Yazbi, A.; Zouein, F.A.; Issa, K.; Eid, A.A.; Ibrahim, M.; Badran, A.; Baydoun, E.; Pintus, G.; et al. MicroRNAs as potential pharmaco-targets in ischemia-reperfusion injury compounded by diabetes. Cells 2019, 8, 152. [Google Scholar] [CrossRef] [Green Version]
- Lakhter, A.J.; Pratt, R.E.; Moore, R.E.; Doucette, K.K.; Maier, B.F.; DiMeglio, L.A.; Sims, E.K. Beta cell extracellular vesicle miR-21-5p cargo is increased in response to inflammatory cytokines and serves as a biomarker of type 1 diabetes. Diabetologia 2018, 61, 1124–1134. [Google Scholar] [CrossRef] [Green Version]
- Salama, A.; Fichou, N.; Allard, M.; Dubreil, L.; De Beaurepaire, L.; Viel, A.; Jégou, D.; Bosch, S.; Bach, J.-M. MicroRNA-29b modulates innate and antigen-specific immune responses in mouse models of autoimmunity. PLoS ONE 2014, 9, e106153. [Google Scholar] [CrossRef] [PubMed]
- Rutman, A.K.; Negi, S.; Gasparrini, M.; Hasilo, C.P.; Tchervenkov, J.; Paraskevas, S. Immune response to extracellular vesicles from human islets of langerhans in patients with type 1 diabetes. Endocrinology 2018, 159, 3834–3847. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, L.; Fang, C.; Fu, W.; Jiang, B.; Li, G.; Qin, L.; Rosenbluth, J.; Gong, G.; Xie, C.B.; Yoo, P.; et al. Endothelial cell–derived interleukin-18 released during ischemia reperfusion injury selectively expands t peripheral helper cells to promote alloantibody production. Circulation 2020, 141, 464–478. [Google Scholar] [CrossRef]
- Andreadou, I.; Cabrera-Fuentes, H.A.; Devaux, Y.; Frangogiannis, N.G.; Frantz, S.; Guzik, T.; Liehn, E.A.; Gomes, C.P.C.; Schulz, R.; Hausenloy, D.J. Immune cells as targets for cardioprotection: New players and novel therapeutic opportunities. Cardiovasc. Res. 2019, 115, 1117–1130. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, L.; Liu, W.; He, X.; Chen, Y.; Lu, J.; Liu, K.; Cao, K.; Yin, P. Association of overweight and obesity with patient mortality after acute myocardial infarction: A meta-analysis of prospective studies. Int. J. Obes. 2016, 40, 220–228. [Google Scholar] [CrossRef]
- Van Gaal, L.F.; Mertens, I.L.; De Block, C.E. Mechanisms linking obesity with cardiovascular disease. Nature 2006, 444, 875–880. [Google Scholar] [CrossRef]
- Mollard, R.; Sénéchal, M.; MacIntosh, A.C.; Hay, J.; Wicklow, B.A.; Wittmeier, K.D.M.; Sellers, E.A.C.; Dean, H.J.; Ryner, L.; Berard, L.; et al. Dietary determinants of hepatic steatosis and visceral adiposity in overweight and obese youth at risk of type 2 diabetes. Am. J. Clin. Nutr. 2014, 99, 804–812. [Google Scholar] [CrossRef] [Green Version]
- Longo, M.; Zatterale, F.; Naderi, J.; Parrilo, L.; Formisano, P.; Raciti, G.A.; Beguinot, F.; Miele, C. Adipose tissue dysfunction as determinant of obesity-associated metabolic complications. Int. J. Mol. Sci. 2019, 20, 2358. [Google Scholar] [CrossRef] [Green Version]
- Alcalá, M.; Calderon-Dominguez, M.; Bustos, E.; Ramos, P.; Casals, N.; Serra, D.; Viana, M.; Herrero, L. Increased inflammation, oxidative stress and mitochondrial respiration in brown adipose tissue from obese mice. Sci. Rep. 2017, 7, 16082. [Google Scholar] [CrossRef] [Green Version]
- Deng, Z.-B.; Poliakov, A.; Hardy, R.W.; Clements, R.; Liu, C.; Liu, Y.; Wang, J.; Xiang, X.; Zhang, S.; Zhuang, X.; et al. Adipose tissue exosome-like vesicles mediate activation of macrophage-induced insulin resistance. Diabetes 2009, 58, 2498–2505. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Crewe, C.; Scherer, P.E. Intercellular and interorgan crosstalk through adipocyte extracellular vesicles. Rev. Endocr. Metab. Disord. 2021, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Thomou, T.; Mori, M.A.; Dreyfuss, J.M.; Konishi, M.; Sakaguchi, M.; Wolfrum, C.; Rao, T.N.; Winnay, J.N.; Garcia-Martin, R.; Grinspoon, S.K.; et al. Adipose-derived circulating miRNAs regulate gene expression in other tissues. Nature 2017, 542, 450–455. [Google Scholar] [CrossRef] [PubMed]
- Ortega, F.J.; Mercader, J.M.; Catalan, V.; Moreno-Navarrete, J.M.; Pueyo, N.; Sabater-Masdeu, M.; Gomez-Ambrosi, J.; Anglada, R.; Formoso, J.A.F.; Ricart, W.; et al. Targeting the circulating MicroRNA signature of obesity. Clin. Chem. 2013, 59, 781–792. [Google Scholar] [CrossRef] [Green Version]
- Zhang, B.; Yang, Y.; Xiang, L.; Zhao, Z.; Ye, R. Adipose-derived exosomes: A novel adipokine in obesity-associated diabetes. J. Cell. Physiol. 2019, 234, 16692–16702. [Google Scholar] [CrossRef]
- Giannella, A.; Radu, C.M.; Franco, L.; Campello, E.; Simioni, P.; Avogaro, A.; De Kreutzenberg, S.V.; Ceolotto, G. Circulating levels and characterization of microparticles in patients with different degrees of glucose tolerance. Cardiovasc. Diabetol. 2017, 16, 118. [Google Scholar] [CrossRef]
- Freeman, D.W.; Hooten, N.N.; Eitan, E.; Green, J.; Mode, N.A.; Bodogai, M.; Zhang, Y.; Lehrmann, E.; Zonderman, A.B.; Biragyn, A.; et al. Altered extracellular vesicle concentration, cargo, and function in diabetes. Diabetes 2018, 67, 2377–2388. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, Y.; Feng, B.; Li, X.; Ni, Y.; Luo, Y. Plasma endothelial microparticles and their correlation with the presence of hypertension and arterial stiffness in patients with type 2 diabetes. J. Clin. Hypertens. 2012, 14, 455–460. [Google Scholar] [CrossRef] [PubMed]
- Rodrigues, K.F.; Pietrani, N.T.; Fernandes, A.P.; Bosco, A.A.; De Sousa, M.C.R.; Silva, I.D.F.O.; Silveira, J.N.; Campos, F.M.F.; Gomes, K.B. Circulating microparticles levels are increased in patients with diabetic kidney disease: A case-control research. Clin. Chim. Acta 2018, 479, 48–55. [Google Scholar] [CrossRef]
- Zhang, X.; McGeoch, S.C.; Johnstone, A.M.; Holtrop, G.; Sneddon, A.; MacRury, S.M.; Megson, I.L.; Pearson, D.W.M.; Abraham, P.; De Roos, B.; et al. Platelet-derived microparticle count and surface molecule expression differ between subjects with and without type 2 diabetes, independently of obesity status. J. Thromb. Thrombol. 2014, 37, 455–463. [Google Scholar] [CrossRef]
- Landers-Ramos, R.Q.; Serra, M.C.; Blumenthal, J.B.; Ryan, A.S.; Hafer-Macko, C.E.; Prior, S.J. Type 2 diabetes and older age contribute to elevated plasma microparticle concentrations independent of chronic stroke. Exp. Physiol. 2018, 103, 1560–1570. [Google Scholar] [CrossRef] [Green Version]
- Jung, K.-H.; Chu, K.; Lee, S.-T.; Bahn, J.-J.; Kim, J.-H.; Kim, M.; Lee, S.K.; Roh, J.-K. Risk of macrovascular complications in type 2 diabetes mellitus: Endothelial microparticle profiles. Cerebrovasc. Dis. 2011, 31, 485–493. [Google Scholar] [CrossRef]
- Gkaliagkousi, E.; Nikolaidou, B.; Gavriilaki, E.; Lazaridis, A.; Yiannaki, E.; Anyfanti, P.; Zografou, I.; Markala, D.; Douma, S. Increased erythrocyte- and platelet-derived microvesicles in newly diagnosed type 2 diabetes mellitus. Diabetes Vasc. Dis. Res. 2019, 16, 458–465. [Google Scholar] [CrossRef]
- Nomura, S. Dynamic role of microparticles in type 2 diabetes mellitus. Curr. Diabetes Rev. 2009, 5, 245–251. [Google Scholar] [CrossRef]
- Dec-Gilowska, M.; Trojnar, M.; Makaruk, B.; Czop, M.; Przybylska-Kuc, S.; Mosiewicz-Madejska, B.; Dzida, G.; Mosiewicz, J. Circulating endothelial microparticles and aortic stiffness in patients with type 2 diabetes mellitus. Medicina 2019, 55, 596. [Google Scholar] [CrossRef] [Green Version]
- Cheng, V.; Kashyap, S.R.; Schauer, P.R.; Kirwan, J.P.; McCrae, K.R. Restoration of glycemic control in patients with type 2 diabetes mellitus after bariatric surgery is associated with reduction in microparticles. Surg. Obes. Relat. Dis. 2013, 9, 207–212. [Google Scholar] [CrossRef] [Green Version]
- Hubal, M.; Nadler, E.P.; Ferrante, S.C.; Barberio, M.; Suh, J.-H.; Wang, J.; Dohm, G.L.; Pories, W.J.; Mietus-Snyder, M.; Freishtat, R. Circulating adipocyte-derived exosomal MicroRNAs associated with decreased insulin resistance after gastric bypass. Obesity 2017, 25, 102–110. [Google Scholar] [CrossRef]
- Ji, C.; Guo, X. The clinical potential of circulating microRNAs in obesity. Nat. Rev. Endocrinol. 2019, 15, 731–743. [Google Scholar] [CrossRef]
- Kumar, A.; Sundaram, K.; Mu, J.; Dryden, G.W.; Sriwastva, M.K.; Lei, C.; Zhang, L.; Qiu, X.; Xu, F.; Yan, J.; et al. High-fat diet-induced upregulation of exosomal phosphatidylcholine contributes to insulin resistance. Nat. Commun. 2021, 12, 213. [Google Scholar] [CrossRef]
- Santilli, F.; Marchisio, M.; Lanuti, P.; Boccatonda, A.; Miscia, S.; Davì, G. Microparticles as new markers of cardiovascular risk in diabetes and beyond. Thromb. Haemost. 2016, 116, 220–234. [Google Scholar] [CrossRef]
- Kranendonk, M.E.G.; De Kleijn, D.P.V.; Kalkhoven, E.; Kanhai, D.A.; Uiterwaal, C.S.P.M.; Van Der Graaf, Y.; Pasterkamp, G.; Visseren, F.L.J.; Doevendans, P.A.; Algra, A.; et al. Extracellular vesicle markers in relation to obesity and metabolic complications in patients with manifest cardiovascular disease. Cardiovasc. Diabetol. 2014, 13, 37. [Google Scholar] [CrossRef] [Green Version]
- Prattichizzo, F.; De Nigris, V.; Sabbatinelli, J.; Giuliani, A.; Castaño, C.; Párrizas, M.; Crespo, I.; Grimaldi, A.; Baranzini, N.; Spiga, R.; et al. CD31+ extracellular vesicles from patients with type 2 diabetes shuttle a mirna signature associated with cardiovascular complications. Diabetes 2021, 70, 240–254. [Google Scholar] [CrossRef]
- Ge, Q.; Xie, X.X.; Xiao, X.; Li, X. Exosome-like vesicles as new mediators and therapeutic targets for treating insulin resistance and beta-cell mass failure in type 2 diabetes mellitus. J. Diabetes Res. 2019, 2019, 3256060. [Google Scholar] [CrossRef] [Green Version]
- Sáez, T.; Toledo, F.; Sobrevia, L. Impaired signalling pathways mediated by extracellular vesicles in diabesity. Mol. Asp. Med. 2019, 66, 13–20. [Google Scholar] [CrossRef]
- Nie, H.; Pan, Y.; Zhou, Y. Exosomal microRNA-194 causes cardiac injury and mitochondrial dysfunction in obese mice. Biochem. Biophys. Res. Commun. 2018, 503, 3174–3179. [Google Scholar] [CrossRef]
- Li, F.; Zhang, K.; Xu, T.; Du, W.; Yu, B.; Liu, Y.; Nie, H. Exosomal microRNA-29a mediates cardiac dysfunction and mitochondrial inactivity in obesity-related cardiomyopathy. Endocrine 2019, 63, 480–488. [Google Scholar] [CrossRef]
- Zou, T.; Zhu, M.; Ma, Y.-C.; Xiao, F.; Yu, X.; Xu, L.; Ma, L.-Q.; Yang, J.; Dong, J.-Z. MicroRNA-410-5p exacerbates high-fat diet-induced cardiac remodeling in mice in an endocrine fashion. Sci. Rep. 2018, 8, 8780. [Google Scholar] [CrossRef]
- Vander Heide, R.S. Increased expression of HSP27 protects canine myocytes from simulated ischemia-reperfusion injury. Am. J. Physiol. Heart Circ. Physiol. 2002, 282, H935–H941. [Google Scholar] [CrossRef]
- Stępień, E.; Durak-Kozica, M.; Kamińska, A.; Targosz-Korecka, M.; Libera, M.; Tylko, G.; Opalińska, A.; Kapusta, M.; Solnica, B.; Georgescu, A.; et al. Circulating ectosomes: Determination of angiogenic microRNAs in type 2 diabetes. Theranostics 2018, 8, 3874–3890. [Google Scholar] [CrossRef]
- Li, S.; Wei, J.; Zhang, C.; Li, X.; Meng, W.; Mo, X.; Zhang, Q.; Liu, Q.; Ren, K.; Du, R.; et al. Cell-derived microparticles in patients with type 2 diabetes mellitus: A systematic review and meta-analysis. Cell. Physiol. Biochem. 2016, 39, 2439–2450. [Google Scholar] [CrossRef]
- Kim, H.; Bae, Y.-U.; Lee, H.; Kim, H.; Jeon, J.S.; Noh, H.; Han, D.C.; Byun, D.W.; Kim, S.H.; Park, H.K.; et al. Effect of diabetes on exosomal miRNA profile in patients with obesity. BMJ Open Diabetes Res. Care 2020, 8, e001403. [Google Scholar] [CrossRef]
- Prattichizzo, F.; Matacchione, G.; Giuliani, A.; Sabbatinelli, J.; Olivieri, F.; de Candia, P.; De Nigris, V.; Ceriello, A. Extracellular vesicle-shuttled miRNAs: A critical appraisal of their potential as nano-diagnostics and nano-therapeutics in type 2 diabetes mellitus and its cardiovascular complications. Theranostics 2021, 11, 1031–1045. [Google Scholar] [CrossRef]
- Wang, X.; Huang, W.; Liu, G.; Cai, W.; Millard, R.W.; Wang, Y.; Chang, J.; Peng, T.; Fan, G.-C. Cardiomyocytes mediate anti-angiogenesis in type 2 diabetic rats through the exosomal transfer of miR-320 into endothelial cells. J. Mol. Cell. Cardiol. 2014, 74, 139–150. [Google Scholar] [CrossRef] [Green Version]
- Santamaria-Martos, F.; Benítez, I.D.; Latorre, J.; Lluch, A.; Moreno-Navarrete, J.M.; Sabater, M.; Ricart, W.; de la Torre, M.S.; Mora, S.; Fernández-Real, J.M.; et al. Comparative and functional analysis of plasma membrane-derived extracellular vesicles from obese vs. nonobese women. Clin. Nutr. 2020, 39, 1067–1076. [Google Scholar] [CrossRef]
- Withers, S.B.; Dewhurst, T.; Hammond, C.; Topham, C.H. MiRNAs as novel adipokines: Obesity-related circulating MiRNAs influence chemosensitivity in cancer patients. Non-Coding RNA 2020, 6, 5. [Google Scholar] [CrossRef] [Green Version]
- Tian, Z.-Q.; Jiang, H.; Lu, Z.-B. MiR-320 regulates cardiomyocyte apoptosis induced by ischemia–reperfusion injury by targeting AKIP1. Cell. Mol. Biol. Lett. 2018, 23, 41. [Google Scholar] [CrossRef] [Green Version]
- Song, C.; Liu, B.; Diao, H.-Y.; Shi, Y.-F.; Zhang, J.-C.; Li, Y.-X.; Liu, N.; Yu, Y.-P.; Wang, G.; Wang, J.-P.; et al. Down-regulation of microRNA-320 suppresses cardiomyocyte apoptosis and protects against myocardial ischemia and reperfusion injury by targeting IGF-1. Oncotarget 2016, 7, 39740–39757. [Google Scholar] [CrossRef]
- Ren, X.-P.; Wu, J.; Wang, X.; Sartor, M.A.; Qian, J.; Jones, K.; Nicolaou, P.; Pritchard, T.J.; Fan, G.-C. MicroRNA-320 is involved in the regulation of cardiac ischemia/reperfusion injury by targeting heat-shock protein 20. Circulation 2009, 119, 2357–2366. [Google Scholar] [CrossRef] [Green Version]
- Wang, W.; Zheng, Y.; Wang, M.; Yan, M.; Jiang, J.; Li, Z. Exosomes derived miR-126 attenuates oxidative stress and apoptosis from ischemia and reperfusion injury by targeting ERRFI1. Gene 2019, 690, 75–80. [Google Scholar] [CrossRef]
- Luo, Q.; Guo, D.; Liu, G.; Chen, G.; Hang, M.; Jin, M. Exosomes from MiR-126-Overexpressing adscs are therapeutic in relieving acute myocardial ischaemic injury. Cell. Physiol. Biochem. 2017, 44, 2105–2116. [Google Scholar] [CrossRef]
- Jansen, F.; Wang, H.; Przybilla, D.; Franklin, B.S.; Dolf, A.; Pfeifer, P.; Schmitz, T.; Flender, A.; Endl, E.; Nickenig, G.; et al. Vascular endothelial microparticles-incorporated microRNAs are altered in patients with diabetes mellitus. Cardiovasc. Diabetol. 2016, 15, 49. [Google Scholar] [CrossRef] [Green Version]
- Wu, K.; Yang, Y.; Zhong, Y.; Ammar, H.M.; Zhang, P.; Guo, R.; Liu, H.; Cheng, C.; Koroscil, T.M.; Chen, Y.; et al. The effects of microvesicles on endothelial progenitor cells are compromised in type 2 diabetic patients via downregulation of the miR-126/VEGFR2 pathway. Am. J. Physiol. Endocrinol. Metab. 2016, 310, E828–E837. [Google Scholar] [CrossRef] [Green Version]
- Ogawa, R.; Tanaka, C.; Sato, M.; Nagasaki, H.; Sugimura, K.; Okumura, K.; Nakagawa, Y.; Aoki, N. Adipocyte-derived microvesicles contain RNA that is transported into macrophages and might be secreted into blood circulation. Biochem. Biophys. Res. Commun. 2010, 398, 723–729. [Google Scholar] [CrossRef]
- Zhou, Y.; Richards, A.M.; Wang, P. MicroRNA-221 Is Cardioprotective and anti-fibrotic in a rat model of myocardial infarction. Mol. Ther. Nucleic Acids 2019, 17, 185–197. [Google Scholar] [CrossRef] [Green Version]
- Lai, T.-C.; Lee, T.-L.; Chang, Y.-C.; Chen, Y.-C.; Lin, S.-R.; Lin, S.-W.; Pu, C.-M.; Tsai, J.-S.; Chen, Y.-L. MicroRNA-221/222 mediates adsc-exosome-induced cardioprotection against ischemia/reperfusion by targeting puma and ets-1. Front. Cell Dev. Biol. 2020, 8, 1475. [Google Scholar] [CrossRef]
- Liu, T.; Sun, Y.-C.; Cheng, P.; Shao, H.-G. Adipose tissue macrophage-derived exosomal miR-29a regulates obesity-associated insulin resistance. Biochem. Biophys. Res. Commun. 2019, 515, 352–358. [Google Scholar] [CrossRef]
- Chiricosta, L.; Silvestro, S.; Gugliandolo, A.; Marconi, G.D.; Pizzicannella, J.; Bramanti, P.; Trubiani, O.; Mazzon, E. Extracellular vesicles of human periodontal ligament stem cells contain micrornas associated to proto-oncogenes: Implications in cytokinesis. Front. Genet. 2020, 11, 582. [Google Scholar] [CrossRef]
- Wang, L.; Niu, X.; Hu, J.; Xing, H.; Sun, M.; Wang, J.; Jian, Q.; Yang, H. After Myocardial Ischemia-Reperfusion, miR-29a, and Let7 Could Affect Apoptosis through Regulating IGF-1. BioMed Res. Int. 2015, 2015, 245412. [Google Scholar] [CrossRef] [Green Version]
- Niu, X.; Pu, S.; Ling, C.; Xu, J.; Wang, J.; Sun, S.; Yao, Y.; Zhang, Z. lncRNA Oip5-as1 attenuates myocardial ischaemia/reperfusion injury by sponging miR-29a to activate the SIRT1/AMPK/PGC1α pathway. Cell Prolif. 2020, 53, e12818. [Google Scholar] [CrossRef]
- Ye, Y.; Hu, Z.; Lin, Y.; Zhang, C.; Perez-Polo, J.R. Downregulation of microRNA-29 by antisense inhibitors and a PPAR-γ agonist protects against myocardial ischaemia-reperfusion injury. Cardiovasc. Res. 2010, 87, 535–544. [Google Scholar] [CrossRef] [Green Version]
- Kukreja, R.C.; Yin, C.; Salloum, F. MicroRNAs: New players in cardiac injury and protection. Mol. Pharmacol. 2011, 80, 558–564. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, J.-G.; Xu, X.-M.; Ji, H.; Sun, B. Inhibiting miR-155 protects against myocardial ischemia/reperfusion injury via targeted regulation of HIF-1α in rats. Iran. J. Basic Med. Sci. 2019, 22, 1050–1058. [Google Scholar] [CrossRef]
- Guo, J.; Liu, H.-B.; Sun, C.; Yan, X.-Q.; Hu, J.; Yu, J.; Yuan, Y.; Du, Z.-M. MicroRNA-155 Promotes Myocardial Infarction-Induced Apoptosis by Targeting RNA-binding protein QKI. Oxid. Med. Cell. Longev. 2019, 2019, 4579806–4579814. [Google Scholar] [CrossRef] [PubMed]
- Ge, X.; Meng, Q.; Wei, L.; Liu, J.; Li, M.; Liang, X.; Lin, F.; Zhang, Y.; Li, Y.; Liu, Z.; et al. Myocardial ischemia-reperfusion induced cardiac extracellular vesicles harbour proinflammatory features and aggravate heart injury. J. Extracell. Vesicles 2021, 10, e12072. [Google Scholar] [CrossRef]
- Xing, X.; Guo, S.; Zhang, G.; Liu, Y.; Bi, S.; Wang, X.; Lu, Q. miR-26a-5p protects against myocardial ischemia/reperfusion injury by regulating the PTEN/PI3K/AKT signaling pathway. Braz. J. Med. Biol. Res. 2020, 53, e9106. [Google Scholar] [CrossRef] [Green Version]
- Xu, H.; Du, X.; Xu, J.; Zhang, Y.; Tian, Y.; Liu, G.; Wang, X.; Ma, M.; Du, W.; Liu, Y.; et al. Pancreatic beta cell microRNA-26a alleviates type 2 diabetes by improving peripheral insulin sensitivity and preserving beta cell function. PLoS Biol. 2020, 18, e3000603. [Google Scholar] [CrossRef] [PubMed]
- Yao, L.; Lv, X.; Wang, X. MicroRNA 26a inhibits HMGB1 expression and attenuates cardiac ischemia-reperfusion injury. J. Pharmacol. Sci. 2016, 131, 6–12. [Google Scholar] [CrossRef] [Green Version]
- Gong, D.D.; Yu, J.; Yu, J.-C.; Diang, X.-D. Effect of miR-26a targeting GSK-3beta/beta-catenin signaling pathway on myocardial apoptosis in rats with myocardial ischemia-reperfusion. Eur. Rev. Med. Pharmacol. Sci. 2019, 23, 7073–7082. [Google Scholar] [CrossRef]
- Liu, C.; Gao, Y.; Wu, J.; Zou, J. Exosomal miR-23a and miR-192, Potential diagnostic biomarkers for type 2 diabetes. Clin. Lab. 2021, 67, 494–593. [Google Scholar] [CrossRef]
- Long, B.; Gan, T.-Y.; Zhang, R.-C.; Zhang, A.Y.-H. miR-23a regulates cardiomyocyte apoptosis by targeting manganese superoxide dismutase. Mol. Cells 2017, 40, 542–549. [Google Scholar] [CrossRef]
- Roy, S.; Benz, F.; Alder, J.; Bantel, H.; Janssen, J.; Vucur, M.; Gautheron, J.; Schneider, A.; Schüller, F.; Loosen, S.; et al. Down-regulation of miR-192-5p protects from oxidative stress-induced acute liver injury. Clin. Sci. 2016, 130, 1197–1207. [Google Scholar] [CrossRef]
- Wang, L.; Li, Q.; Diao, J.; Lin, L.; Wei, J. MiR-23a is involved in myocardial ischemia/reperfusion injury by directly targeting cx43 and regulating mitophagy. Inflammation 2021, 44, 1581–1591. [Google Scholar] [CrossRef]
- Santovito, D.; De Nardis, V.; Marcantonio, P.; Mandolini, C.; Paganelli, C.; Vitale, E.; Buttitta, F.; Bucci, M.; Mezzetti, A.; Consoli, A.; et al. Plasma exosome microrna profiling unravels a new potential modulator of adiponectin pathway in diabetes: Effect of glycemic control. J. Clin. Endocrinol. Metab. 2014, 99, E1681–E1685. [Google Scholar] [CrossRef] [PubMed]
- Al-Qaissi, A.; Papageorgiou, M.; Deshmukh, H.; Madden, L.A.; Rigby, A.; Kilpatrick, E.S.; Atkin, S.L.; Sathyapalan, T. Effects of acute insulin-induced hypoglycaemia on endothelial microparticles in adults with and without type 2 diabetes. Diabetes Obes. Metab. 2019, 21, 533–540. [Google Scholar] [CrossRef] [PubMed]
- Ghai, V.; Kim, T.-K.; Etheridge, A.; Nielsen, T.; Hansen, T.; Pedersen, O.; Galas, D.; Wang, K. Extracellular vesicle encapsulated micrornas in patients with type 2 diabetes are affected by metformin treatment. J. Clin. Med. 2019, 8, 617. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, J.; Xu, J.-P.; Zhao, X.-Z.; Sun, X.-J.; Xu, Z.-W.; Song, S.-J. Protective effect of metformin on myocardial injury in metabolic syndrome patients following percutaneous coronary intervention. Cardiology 2014, 127, 133–139. [Google Scholar] [CrossRef]
- El Messaoudi, S.; Nederlof, R.; Zuurbier, C.J.; van Swieten, H.A.; Pickkers, P.; Noyez, L.; Dieker, H.-J.; Coenen, M.; Donders, A.R.T.; Vos, A.; et al. Effect of metformin pretreatment on myocardial injury during coronary artery bypass surgery in patients without diabetes (MetCAB): A double-blind, randomised controlled trial. Lancet Diabetes Endocrinol. 2015, 3, 615–623. [Google Scholar] [CrossRef]
- El Messaoudi, S.; Schreuder, T.H.; Kengen, R.D.; Rongen, G.A.; Broek, P.H.V.D.; Thijssen, D.H.J.; Riksen, N.P. Impact of metformin on endothelial ischemia-reperfusion injury in humans in vivo: A prospective randomized open, blinded-endpoint study. PLoS ONE 2014, 9, e96062. [Google Scholar] [CrossRef] [Green Version]
- Higgins, L.; Palee, S.; Chattipakorn, S.C.; Chattipakorn, N. Effects of metformin on the heart with ischaemia-reperfusion injury: Evidence of its benefits from in vitro, in vivo and clinical reports. Eur. J. Pharmacol. 2019, 858, 172489. [Google Scholar] [CrossRef]
- Shimazu, T.; Inami, N.; Satoh, D.; Kajiura, T.; Yamada, K.; Iwasaka, T.; Nomura, S. Effect of acarbose on platelet-derived microparticles, soluble selectins, and adiponectin in diabetic patients. J. Thromb. Thrombol. 2009, 28, 429–435. [Google Scholar] [CrossRef]
- Tsimerman, G.; Roguin, A.; Bachar, A.; Melamed, E.; Brenner, B.; Aharon, A. Involvement of microparticles in diabetic vascular complications. Thromb. Haemost. 2011, 106, 310–321. [Google Scholar] [CrossRef]
- Thompson, M.D.; Cismowski, M.J.; Serpico, M.; Pusateri, A.; Brigstock, D.R. Elevation of circulating microRNA levels in obese children compared to healthy controls. Clin. Obes. 2017, 7, 216–221. [Google Scholar] [CrossRef]
- Cui, X.; You, L.; Zhu, L.; Wang, X.; Zhou, Y.; Li, Y.; Wen, J.; Xia, Y.; Wang, X.; Ji, C.; et al. Change in circulating microRNA profile of obese children indicates future risk of adult diabetes. Metabolism 2018, 78, 95–105. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pescador, N.; Pérez-Barba, M.; Ibarra, J.M.; Corbatón-Anchuelo, A.; Martínez-Larrad, M.T.; Serrano-Ríos, M. Serum circulating microrna profiling for identification of potential type 2 diabetes and obesity biomarkers. PLoS ONE 2013, 8, e77251. [Google Scholar] [CrossRef] [Green Version]
- Liu, Y.; Yang, L.; Yin, J.; Su, D.; Pan, Z.; Li, P.; Wang, X. MicroRNA-15b deteriorates hypoxia/reoxygenation-induced cardiomyocyte apoptosis by downregulating Bcl-2 and MAPK3. J. Investig. Med. 2018, 66, 39–45. [Google Scholar] [CrossRef] [PubMed]
- Liu, L.; Zhang, G.; Liang, Z.; Liu, X.; Li, T.; Fan, J.; Bai, J.; Wang, Y. MicroRNA-15b enhances hypoxia/reoxygenation-induced apoptosis of cardiomyocytes via a mitochondrial apoptotic pathway. Apoptosis 2014, 19, 19–29. [Google Scholar] [CrossRef]
- Niu, S.; Xu, L.; Yuan, Y.; Yang, S.; Ning, H.; Qin, X.; Xin, P.; Yuan, D.; Jiao, J.; Zhao, Y. Effect of down-regulated miR-15b-5p expression on arrhythmia and myocardial apoptosis after myocardial ischemia reperfusion injury in mice. Biochem. Biophys. Res. Commun. 2020, 530, 54–59. [Google Scholar] [CrossRef] [PubMed]
- Wang, P.; Sun, J.; Lv, S.; Xie, T.; Wang, X. Apigenin alleviates myocardial reperfusion injury in rats by downregulating miR-15b. Med. Sci. Monit. 2019, 25, 2764–2776. [Google Scholar] [CrossRef] [PubMed]
- Kou, Y.; Zheng, W.T.; Zhang, Y.R. Inhibition of miR-23 protects myocardial function from ischemia-reperfusion injury through restoration of glutamine metabolism. Eur. Rev. Med. Pharmacol. Sci. 2016, 20, 4286–4293. [Google Scholar]
- Lu, M.; Xu, Y.; Wang, M.; Guo, T.; Luo, F.; Su, N.; Wang, Z.; Xu, L.; Liu, Z. MicroRNA-23 inhibition protects the ischemia/reperfusion injury via inducing the differentiation of bone marrow mesenchymal stem cells into cardiomyocytes. Int. J. Clin. Exp. Pathol. 2019, 12, 1060. [Google Scholar]
- Hijmans, J.G.; Diehl, K.J.; Bammert, T.D.; Kavlich, P.J.; Lincenberg, G.M.; Greiner, J.J.; Stauffer, B.L.; DeSouza, C.A. Influence of overweight and obesity on circulating inflammation-related microRNA. MicroRNA 2018, 7, 148–154. [Google Scholar] [CrossRef] [PubMed]
- Manoel Alves, J.; Handerson Gomes Teles, R.; do Valle Gomes Gatto, C.; Muñoz, V.R.; Regina Cominetti, M.; Garcia de Oliveira Duarte, A.C. Mapping research in the obesity, adipose tissue, and microrna field: A bibliometric analysis. Cells 2019, 8, 1581. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, Y.; Cheng, H.-W.; Qiu, Y.; Dupee, D.; Noonan, M.; Lin, Y.-D.; Fisch, S.; Unno, K.; Sereti, K.-I.; Liao, R. MicroRNA-34a plays a key role in cardiac repair and regeneration following myocardial infarction. Circ. Res. 2015, 117, 450–459. [Google Scholar] [CrossRef] [PubMed]
- Zhao, T.; Li, J.; Chen, A.F. MicroRNA-34a induces endothelial progenitor cell senescence and impedes its angiogenesis via suppressing silent information regulator 1. Am. J. Physiol. Metab. 2010, 299, E110–E116. [Google Scholar] [CrossRef] [Green Version]
- Li, J.H.; Dai, J.; Han, B.; Wu, G.-H.; Wang, C.-H. MiR-34a regulates cell apoptosis after myocardial infarction in rats through the Wnt/beta-catenin signaling pathway. Eur. Rev. Med. Pharmacol. Sci. 2019, 23, 2555–2562. [Google Scholar] [CrossRef]
- Wang, X.; Yuan, B.; Cheng, B.; Liu, Y.; Zhang, B.; Wang, X.; Lin, X.; Yang, B.; Gong, G. Crocin alleviates myocardial ischemia/reperfusion-induced endoplasmic reticulum stress via regulation of mir-34a/sirt1/nrf2 pathway. Shock 2019, 51, 123–130. [Google Scholar] [CrossRef] [PubMed]
- Dong, F.-F.; Dong, S.-H.; Liang, Y.; Wang, K.; Qin, Y.-W.; Zhao, X.-X. MiR-34a promotes myocardial infarction in rats by inhibiting the activity of SIRT1. Eur. Rev. Med. Pharmacol. Sci. 2019, 23, 7059–7065. [Google Scholar]
- Heneghan, H.; Miller, N.; McAnena, O.J.; O’Brien, T.; Kerin, M.J. Differential miRNA Expression in omental adipose tissue and in the circulation of obese patients identifies novel metabolic biomarkers. J. Clin. Endocrinol. Metab. 2011, 96, E846–E850. [Google Scholar] [CrossRef]
- Wang, R.; Hong, J.; Cao, Y.; Shi, J.; Gu, W.; Ning, G.; Zhang, Y.; Wang, W. Elevated circulating microRNA-122 is associated with obesity and insulin resistance in young adults. Eur. J. Endocrinol. 2015, 172, 291–300. [Google Scholar] [CrossRef]
- Zhang, Z.; Li, H.; Chen, S.; Li, Y.; Cui, Z.; Ma, J. MicroRNA-122 regulates caspase-8 and promotes the apoptosis of mouse cardiomyocytes. Braz. J. Med. Biol. Res. 2017, 50, e5760. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Peng, H.; Luo, Y.; Ying, Y. lncRNA XIST attenuates hypoxia-induced H9c2 cardiomyocyte injury by targeting the miR-122-5p/FOXP2 axis. Mol. Cell. Probes 2020, 50, 101500. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.; Li, H.; Cui, Z.; Zhou, Z.; Chen, S.; Ma, J.; Hou, L.; Pan, X.; Li, Q. Long non-coding RNA UCA1 relieves cardiomyocytes H9c2 injury aroused by oxygen-glucose deprivation via declining miR-122. Artif. Cells Nanomed. Biotechnol. 2019, 47, 3492–3499. [Google Scholar] [CrossRef] [PubMed]
- Thomé, J.G.; Mendoza, M.R.; Cheuiche, A.V.; La Porta, V.L.; Silvello, D.; dos Santos, K.G.; Andrades, M.E.; Clausell, N.; Rohde, L.E.; Biolo, A. Circulating microRNAs in obese and lean heart failure patients: A case–control study with computational target prediction analysis. Gene 2015, 574, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Al-Rawaf, H.A. Circulating microRNAs and adipokines as markers of metabolic syndrome in adolescents with obesity. Clin. Nutr. 2019, 38, 2231–2238. [Google Scholar] [CrossRef]
- Liang, Y.; Li, J.; Xiao, H.; He, Y.; Zhang, L.; Yan, Y. Identification of stress-related microRNA biomarkers in type 2 diabetes mellitus: A systematic review and meta-analysis. J. Diabetes 2020, 12, 633–644. [Google Scholar] [CrossRef] [Green Version]
- Chu, X.; Wang, Y.; Pang, L.; Huang, J.; Sun, X.; Chen, X. miR-130 aggravates acute myocardial infarction-induced myocardial injury by targeting PPAR-gamma. J. Cell. Biochem. 2018, 119, 7235–7244. [Google Scholar] [CrossRef]
- Huang, G.; Hao, F.; Hu, X. Downregulation of microRNA-155 stimulates sevoflurane-mediated cardioprotection against myocardial ischemia/reperfusion injury by binding to SIRT1 in mice. J. Cell. Biochem. 2019, 120, 15494–15505. [Google Scholar] [CrossRef] [PubMed]
- Eisenhardt, S.U.; Weiss, J.B.W.; Smolka, C.; Maxeiner, J.; Pankratz, F.; Bemtgen, X.; Kustermann, M.; Thiele, J.R.; Schmidt, Y.; Stark, G.B.; et al. MicroRNA-155 aggravates ischemia–reperfusion injury by modulation of inflammatory cell recruitment and the respiratory oxidative burst. Basic Res. Cardiol. 2015, 110, 32. [Google Scholar] [CrossRef]
- Villard, A.; Marchand, L. Diagnostic value of cell-free circulating micrornas for obesity and type 2 diabetes: A meta-analysis. J. Mol. Biomark. Diagn. 2015, 6, 1–9. [Google Scholar] [CrossRef] [Green Version]
- Karolina, D.S.; Tavintharan, S.; Armugam, A.; Sepramaniam, S.; Pek, S.L.T.; Wong, M.T.K.; Lim, S.C.; Sum, C.F.; Jeyaseelan, K. Circulating miRNA profiles in patients with metabolic syndrome. J. Clin. Endocrinol. Metab. 2012, 97, E2271–E2276. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Y.; Huang, R.; Zhou, W.; Zhao, Q.; Lü, Z. miR-192-5p mediates hypoxia/reoxygenation-induced apoptosis in H9c2 cardiomyocytes via targeting of FABP3. J. Biochem. Mol. Toxicol. 2017, 31, e21873. [Google Scholar] [CrossRef] [PubMed]
- Cao, L.; Chai, S. miR-320-3p is involved in morphine pre-conditioning to protect rat cardiomyocytes from ischemia/reperfusion injury through targeting Akt3. Mol. Med. Rep. 2020, 22, 1480–1488. [Google Scholar] [CrossRef] [PubMed]
- Gallo, W.; Esguerra, J.L.S.; Eliasson, L.; Melander, O. miR-483-5p associates with obesity and insulin resistance and independently associates with new onset diabetes mellitus and cardiovascular disease. PLoS ONE 2018, 13, e0206974. [Google Scholar] [CrossRef]
- Gallo, W.; Ottosson, F.; Kennbäck, C.; Jujic, A.; Esguerra, J.L.S.; Eliasson, L.; Melander, O. Replication study reveals miR-483-5p as an important target in prevention of cardiometabolic disease. BMC Cardiovasc. Disord. 2021, 21, 162. [Google Scholar] [CrossRef]
- Zhang, H.; Wang, J.; Du, A.; Li, Y. MiR-483-3p inhibition ameliorates myocardial ischemia/reperfusion injury by targeting the MDM4/p53 pathway. Mol. Immunol. 2020, 125, 9–14. [Google Scholar] [CrossRef]
- Sun, H.; Cai, J.; Xu, L.; Liu, J.; Chen, M.; Zheng, M.; Wang, L.; Yang, X. miR-483-3p regulates acute myocardial infarction by transcriptionally repressing insulin growth factor 1 expression. Mol. Med. Rep. 2018, 17, 4785–4790. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Qiao, Y.; Zhao, Y.; Liu, Y.; Ma, N.; Wang, C.; Zou, J.; Liu, Z.; Zhou, Z.; Han, D.; He, J.; et al. miR-483-3p regulates hyperglycaemia-induced cardiomyocyte apoptosis in transgenic mice. Biochem. Biophys. Res. Commun. 2016, 477, 541–547. [Google Scholar] [CrossRef]
- Chien, H.Y.; Lee, T.-P.; Chen, C.-Y.; Chiu, Y.-H.; Lin, Y.-C.; Lee, L.-S.; Li, W.-C. Circulating microRNA as a diagnostic marker in populations with type 2 diabetes mellitus and diabetic complications. J. Chin. Med. Assoc. 2015, 78, 204–211. [Google Scholar] [CrossRef] [Green Version]
- Sekar, D.; Venugopal, B.; Sekar, P.; Ramalingam, K. Role of microRNA 21 in diabetes and associated/related diseases. Gene 2016, 582, 14–18. [Google Scholar] [CrossRef]
- Liao, Z.; Chen, Y.; Duan, C.; Zhu, K.; Huang, R.; Zhao, H.; Hintze, M.; Pu, Q.; Yuan, Z.; Lv, L.; et al. Cardiac telocytes inhibit cardiac microvascular endothelial cell apoptosis through exosomal miRNA-21-5p-targeted cdip1 silencing to improve angiogenesis following myocardial infarction. Theranostics 2021, 11, 268–291. [Google Scholar] [CrossRef] [PubMed]
- Cheng, Y.; Liu, X.; Zhang, S.; Lin, Y.; Yang, J.; Zhang, C. MicroRNA-21 protects against the H2O2-induced injury on cardiac myocytes via its target gene PDCD4. J. Mol. Cell. Cardiol. 2009, 47, 5–14. [Google Scholar] [CrossRef] [Green Version]
- Cheng, Y.; Zhu, P.; Yang, J.; Liu, X.; Dong, S.; Wang, X.; Chun, B.; Zhuang, J.; Zhang, C. Ischaemic preconditioning-regulated miR-21 protects heart against ischaemia/reperfusion injury via anti-apoptosis through its target PDCD4. Cardiovasc. Res. 2010, 87, 431–439. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ma, N.; Bai, J.; Zhang, W.; Luo, H.; Zhang, X.; Liu, D.; Qiao, C. Trimetazidine protects against cardiac ischemia/reperfusion injury via effects on cardiac miRNA-21 expression, Akt and the Bcl-2/Bax pathway. Mol. Med. Rep. 2016, 14, 4216–4222. [Google Scholar] [CrossRef] [Green Version]
- Qiao, S.; Olson, J.; Paterson, M.; Yan, Y.; Zaja, I.; Liu, Y.; Riess, M.L.; Kersten, J.R.; Liang, M.; Warltier, D.C.; et al. MicroRNA-21 mediates isoflurane-induced cardioprotection against ischemia–reperfusion injury via akt/nitric oxide synthase/mitochondrial permeability transition pore pathway. Anesthesiology 2015, 123, 786–798. [Google Scholar] [CrossRef] [Green Version]
- Yang, L.; Wang, B.; Zhou, Q.; Wang, Y.; Liu, X.; Liu, Z.; Zhan, Z. MicroRNA-21 prevents excessive inflammation and cardiac dysfunction after myocardial infarction through targeting KBTBD7. Cell Death Dis. 2018, 9, 769. [Google Scholar] [CrossRef] [Green Version]
- Salloum, F.; Yin, C.; Kukreja, R.C. Role of microRNAs in cardiac preconditioning. J. Cardiovasc. Pharmacol. 2010, 56, 581–588. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Masi, L.N.; Lotufo, P.A.; Ferreira, F.M.; Rodrigues, A.C.; Serdan, T.D.A.; Braga, A.A.; Cury, D.P.; Hirata, M.J.; Curi, R.; Bensenor, I.J.M.; et al. Profiling plasma-extracellular vesicle proteins and microRNAs in diabetes onset in middle-aged male participants in the ELSA-Brasil study. Physiol. Rep. 2021, 9, e14731. [Google Scholar] [CrossRef] [PubMed]
- Ruan, Z.; Wang, S.; Yu, W.; Deng, F. LncRNA MALAT1 aggravates inflammation response through regulating PTGS2 by targeting miR-26b in myocardial ischemia-reperfusion injury. Int. J. Cardiol. 2019, 288, 122. [Google Scholar] [CrossRef]
- Chen, X.; Ding, Z.; Li, T.; Jiang, W.; Zhang, J.; Deng, X. MicroR-26b Targets High Mobility Group, AT-hook 2 to Ameliorate Myocardial Infarction-induced Fibrosis by Suppression of Cardiac Fibroblasts Activation. Curr. Neurovasc. Res. 2020, 17, 204–213. [Google Scholar] [CrossRef]
- Jiang, T.; Liu, Y.; Chen, B.; Si, L. Identification of potential molecular mechanisms and small molecule drugs in myocardial ischemia/reperfusion injury. Braz. J. Med. Biol. Res. 2020, 53. [Google Scholar] [CrossRef]
- Ortega, F.J.; Mercader, J.M.; Moreno-Navarrete, J.M.; Rovira, O.; Guerra, E.; Esteve, E.; Xifra, G.; Martínez, C.; Ricart, W.; Rieusset, J.; et al. Profiling of circulating micrornas reveals common micrornas linked to type 2 diabetes that change with insulin sensitization. Diabetes Care 2014, 37, 1375–1383. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Su, Q.; Liu, Y.; Lv, X.-W.; Ye, Z.-L.; Sun, Y.-H.; Kong, B.-H.; Qin, Z.-B. Inhibition of lncRNA TUG1 upregulates miR-142-3p to ameliorate myocardial injury during ischemia and reperfusion via targeting HMGB1- and Rac1-induced autophagy. J. Mol. Cell. Cardiol. 2019, 133, 12–25. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Ouyang, M.; Wang, Q.; Jian, Z. MicroRNA-142-3p inhibits hypoxia/reoxygenation-induced apoptosis and fibrosis of cardiomyocytes by targeting high mobility group box 1. Int. J. Mol. Med. 2016, 38, 1377–1386. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Su, Q.; Lv, X.; Ye, Z.; Sun, Y.; Kong, B.; Qin, Z.; Li, L. The mechanism of miR-142-3p in coronary microembolization-induced myocardiac injury via regulating target gene IRAK-1. Cell Death Dis. 2019, 10, 61. [Google Scholar] [CrossRef] [Green Version]
- Cui, X.; He, Z.; Liang, Z.; Chen, Z.; Wang, H.; Zhang, J. Exosomes from adipose-derived mesenchymal stem cells protect the myocardium against ischemia/reperfusion injury through wnt/beta-catenin signaling pathway. J. Cardiovasc. Pharmacol. 2017, 70, 225–231. [Google Scholar] [CrossRef] [Green Version]
- Rong, Y.; Bao, W.; Shan, Z.; Liu, J.; Yu, X.; Xia, S.; Gao, H.; Wang, X.; Yao, P.; Hu, F.B.; et al. Increased microRNA-146a levels in plasma of patients with newly diagnosed type 2 diabetes mellitus. PLoS ONE 2013, 8, e73272. [Google Scholar] [CrossRef] [Green Version]
- Wang, X.; Ha, T.; Liu, L.; Zou, J.; Zhang, X.; Kalbfleisch, J.; Gao, X.; Williams, D.; Li, C. Increased expression of microRNA-146a decreases myocardial ischaemia/reperfusion injury. Cardiovasc. Res. 2013, 97, 432–442. [Google Scholar] [CrossRef] [Green Version]
- Zhang, T.; Ma, Y.; Gao, L.; Mao, C.; Zeng, H.; Wang, X.; Sun, Y.; Gu, J.; Wang, Y.; Chen, K.; et al. MicroRNA-146a protects against myocardial ischaemia reperfusion injury by targeting Med1. Cell. Mol. Biol. Lett. 2019, 24, 62. [Google Scholar] [CrossRef]
- Zhang, W.; Shao, M.; He, X.; Wang, B.; Li, Y.; Guo, X. Overexpression of microRNA-146 protects against oxygen-glucose deprivation/recovery-induced cardiomyocyte apoptosis by inhibiting the NF-κB/TNF-α signaling pathway. Mol. Med. Rep. 2017, 17, 1913–1918. [Google Scholar] [CrossRef]
- Di, Y.-F.; Li, D.-C.; Shen, Y.-Q.; Wang, C.-L.; Zhang, D.-Y.; Shang, A.; Hu, T. MiR-146b protects cardiomyocytes injury in myocardial ischemia/reperfusion by targeting Smad4. Am. J. Transl. Res. 2017, 9, 656–663. [Google Scholar] [PubMed]
- Pan, J.; Alimujiang, M.; Chen, Q.; Shi, H.; Luo, X. Exosomes derived from miR-146a-modified adipose-derived stem cells attenuate acute myocardial infarction−induced myocardial damage via downregulation of early growth response factor 1. J. Cell. Biochem. 2019, 120, 4433–4443. [Google Scholar] [CrossRef]
- Su, Q.; Xu, Y.; Cai, R.; Dai, R.; Yang, X.; Liu, Y.; Kong, B. miR-146a inhibits mitochondrial dysfunction and myocardial infarction by targeting cyclophilin D. Mol. Ther. Nucleic Acids 2021, 23, 1258–1271. [Google Scholar] [CrossRef] [PubMed]
- Seo, H.-H.; Lee, S.-Y.; Lee, C.Y.; Kim, R.; Kim, P.; Oh, S.; Lee, H.; Lee, M.Y.; Kim, J.; Kim, L.K.; et al. Exogenous miRNA-146a enhances the therapeutic efficacy of human mesenchymal stem cells by increasing vascular endothelial growth factor secretion in the ischemia/reperfusion-injured heart. J. Vasc. Res. 2017, 54, 100–108. [Google Scholar] [CrossRef] [PubMed]
- Wu, Z.; Cheng, S.; Wang, S.; Li, W.; Liu, J. BMSCs-derived exosomal microRNA-150-5p attenuates myocardial infarction in mice. Int. Immunopharmacol. 2021, 93, 107389. [Google Scholar] [CrossRef]
- Zhou, Y.; Chen, Q.; Lew, K.S.; Richards, A.M.; Wang, P. Discovery of potential therapeutic mirna targets in cardiac ischemia–reperfusion injury. J. Cardiovasc. Pharmacol. Ther. 2016, 21, 296–309. [Google Scholar] [CrossRef] [PubMed]
- Ou, H.; Teng, H.; Qin, Y.; Luo, X.; Yang, P.; Zhang, W.; Chen, W.; Lv, D.; Tang, H. Extracellular vesicles derived from microRNA-150-5p-overexpressing mesenchymal stem cells protect rat hearts against ischemia/reperfusion. Aging 2020, 12, 12669–12683. [Google Scholar] [CrossRef]
- Lin, D.; Cui, B.; Ma, J.; Ren, J. MiR-183-5p protects rat hearts against myocardial ischemia/reperfusion injury through targeting VDAC1. BioFactors 2020, 46, 83–93. [Google Scholar] [CrossRef]
- Xing, J.; Xie, T.; Tan, W.; Li, R.; Yu, C.; Han, X. microRNA-183 improve myocardial damager via NF-kb pathway: In vitro and in vivo study. J. Cell. Biochem. 2018, 120, 10145–10154. [Google Scholar] [CrossRef]
- Gong, L.; Xu, H.; Chang, H.; Tong, Y.; Zhang, T.; Guo, G. Knockdown of long non-coding RNA MEG3 protects H9c2 cells from hypoxia-induced injury by targeting microRNA-183. J. Cell. Biochem. 2018, 119, 1429–1440. [Google Scholar] [CrossRef]
- Prats-Puig, A.; Ortega, F.J.; Mercader, J.M.; Moreno-Navarrete, J.M.; Moreno, M.; Bonet, N.; Ricart, W.; López-Bermejo, A.; Fernández-Real, J.M. Changes in circulating micrornas are associated with childhood obesity. J. Clin. Endocrinol. Metab. 2013, 98, E1655–E1660. [Google Scholar] [CrossRef]
- Taheri, M.; Eghtedarian, R.; Ghafouri-Fard, S.; Omrani, M.D. Non-coding RNAs and type 2 diabetes mellitus. Arch. Physiol. Biochem. 2020, 1–10. [Google Scholar] [CrossRef]
- Marzano, F.; Faienza, M.F.; Caratozzolo, M.F.; Brunetti, G.; Chiara, M.; Horner, D.S.; Annese, A.; D’Erchia, A.M.; Consiglio, A.; Pesole, G.; et al. Pilot study on circulating miRNA signature in children with obesity born small for gestational age and appropriate for gestational age. Pediatr. Obes. 2018, 13, 803–811. [Google Scholar] [CrossRef]
- Sun, X.-H.; Wang, X.; Zhang, Y.; Hui, J. Exosomes of bone-marrow stromal cells inhibit cardiomyocyte apoptosis under ischemic and hypoxic conditions via miR-486-5p targeting the PTEN/PI3K/AKT signaling pathway. Thromb. Res. 2019, 177, 23–32. [Google Scholar] [CrossRef]
- Zhu, H.-H.; Wang, X.-T.; Sun, Y.-H.; He, W.-K.; Liang, J.-B.; Mo, B.-H.; Li, L. MicroRNA-486-5p targeting PTEN Protects Against Coronary Microembolization-Induced Cardiomyocyte Apoptosis in Rats by activating the PI3K/AKT pathway. Eur. J. Pharmacol. 2019, 855, 244–251. [Google Scholar] [CrossRef]
- Zhang, X.; Zhang, C.; Wang, N.; Li, Y.; Zhang, D.; Li, Q. MicroRNA-486 alleviates hypoxia-induced damage in h9c2 cells by targeting ndrg2 to inactivate jnk/c-jun and nf-κb signaling pathways. Cell. Physiol. Biochem. 2018, 48, 2483–2492. [Google Scholar] [CrossRef]
- Li, Q.; Xu, Y.; Lv, K.; Wang, Y.; Zhong, Z.; Xiao, C.; Zhu, K.; Ni, C.; Wang, K.; Kong, M.; et al. Small extracellular vesicles containing miR-486-5p promote angiogenesis after myocardial infarction in mice and nonhuman primates. Sci. Transl. Med. 2021, 13, 584. [Google Scholar] [CrossRef]
- Ren, D.; Li, F.; Gao, A.; Cao, Q.; Liu, Y.; Zhang, J. Hypoxia-induced apoptosis of cardiomyocytes is restricted by ginkgolide B-downregulated microRNA-29. Cell Cycle 2020, 19, 1067–1076. [Google Scholar] [CrossRef]
- Tian, R.; Guan, X.; Qian, H.; Wang, L.; Shen, Z.; Fang, L.; Liu, Z. Restoration of NRF2 attenuates myocardial ischemia reperfusion injury through mediating microRNA -29a-3p/ CCNT2 axis. BioFactors 2021, 47, 414–426. [Google Scholar] [CrossRef]
- Hong, H.; Tao, T.; Chen, S.; Liang, C.; Qiu, Y.; Zhou, Y.; Zhang, R. MicroRNA-143 promotes cardiac ischemia-mediated mitochondrial impairment by the inhibition of protein kinase Cepsilon. Basic Res. Cardiol. 2017, 112, 60. [Google Scholar] [CrossRef]
- Ma, W.-Y.; Song, R.-J.; Xu, B.-B.; Xu, Y.; Wang, X.-X.; Sun, H.-Y.; Li, S.-N.; Liu, S.-Z.; Yu, M.-X.; Yang, F.; et al. Melatonin promotes cardiomyocyte proliferation and heart repair in mice with myocardial infarction via miR-143-3p/Yap/Ctnnd1 signaling pathway. Acta Pharmacol. Sin. 2020, 42, 921–931. [Google Scholar] [CrossRef]
- Li, C.; Li, J.; Xue, K.; Zhang, J.; Wang, C.; Zhang, Q.; Chen, X.; Gao, C.; Yu, X.; Sun, L. MicroRNA-143-3p promotes human cardiac fibrosis via targeting sprouty3 after myocardial infarction. J. Mol. Cell. Cardiol. 2019, 129, 281–292. [Google Scholar] [CrossRef]
- Liu, K.; Zhao, D.; Wang, D. LINC00528 regulates myocardial infarction by targeting the miR-143-3p/COX-2 axis. Bioengineered 2020, 11, 11–18. [Google Scholar] [CrossRef] [Green Version]
- Wen, D.; Qiao, P.; Wang, L. Circulating microRNA-223 as a potential biomarker for obesity. Obes. Res. Clin. Pract. 2015, 9, 398–404. [Google Scholar] [CrossRef]
- Sánchez-Ceinos, J.; Rangel-Zuñiga, O.A.; Clemente-Postigo, M.; Podadera-Herreros, A.; Camargo, A.; Alcalá-Diaz, J.F.; Guzmán-Ruiz, R.; López-Miranda, J.; Malagón, M.M. miR-223-3p as a potential biomarker and player for adipose tissue dysfunction preceding type 2 diabetes onset. Mol. Ther. Nucleic Acids 2021, 23, 1035–1052. [Google Scholar] [CrossRef]
- Yan, L.-N.; Zhang, X.; Xu, F.; Fan, Y.-Y.; Ge, B.; Guo, H.; Li, Z.-L. Four-microRNA signature for detection of type 2 diabetes. World J. Clin. Cases 2020, 8, 1923–1931. [Google Scholar] [CrossRef]
- Tang, Q.; Li, M.-Y.; Su, Y.-F.; Fu, J.; Zou, Z.-Y.; Wang, Y.; Li, S.-N. Absence of miR-223-3p ameliorates hypoxia-induced injury through repressing cardiomyocyte apoptosis and oxidative stress by targeting KLF15. Eur. J. Pharmacol. 2018, 841, 67–74. [Google Scholar] [CrossRef]
- Liu, X.; Xu, Y.; Deng, Y.; Li, H. MicroRNA-223 regulates cardiac fibrosis after myocardial infarction by targeting RASA1. Cell. Physiol. Biochem. 2018, 46, 1439–1454. [Google Scholar] [CrossRef]
- Dai, G.-H.; Ma, P.-Z.; Song, X.-B.; Liu, N.; Zhang, T.; Wu, B. MicroRNA-223-3p inhibits the angiogenesis of ischemic cardiac microvascular endothelial cells via affecting rps6kb1/hif-1a signal pathway. PLoS ONE 2014, 9, e108468. [Google Scholar] [CrossRef] [Green Version]
- Liu, X.; Deng, Y.; Xu, Y.; Jin, W.; Li, H. MicroRNA-223 protects neonatal rat cardiomyocytes and H9c2 cells from hypoxia-induced apoptosis and excessive autophagy via the Akt/mTOR pathway by targeting PARP-1. J. Mol. Cell. Cardiol. 2018, 118, 133–146. [Google Scholar] [CrossRef]
- Qin, D.; Wang, X.; Li, Y.; Yang, L.; Wang, R.; Peng, J.; Essandoh, K.; Mu, X.; Peng, T.; Han, Q.; et al. MicroRNA-223-5p and -3p cooperatively suppress necroptosis in ischemic/reperfused hearts. J. Biol. Chem. 2016, 291, 20247–20259. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhao, L.; Jiang, S.; Wu, N.; Shi, E.; Yang, L.; Li, Q. MiR-17-5p-mediated endoplasmic reticulum stress promotes acute myocardial ischemia injury through targeting Tsg101. Cell Stress Chaperon 2021, 26, 77–90. [Google Scholar] [CrossRef] [PubMed]
- Du, W.; Pan, Z.; Chen, X.; Wang, L.; Zhang, Y.; Li, S.; Liang, H.; Xu, C.; Wu, Y.; Shan, H.; et al. By targeting Stat3 microRNA-17-5p promotes cardiomyocyte apoptosis in response to ischemia followed by reperfusion. Cell. Physiol. Biochem. 2014, 34, 955–965. [Google Scholar] [CrossRef] [PubMed]
- Yang, S.; Fan, T.; Hu, Q.; Xu, W.; Yang, J.; Xu, C.; Zhang, B.; Chen, J.; Jiang, H. Downregulation of microRNA-17-5p improves cardiac function after myocardial infarction via attenuation of apoptosis in endothelial cells. Mol. Genet. Genom. 2018, 293, 883–894. [Google Scholar] [CrossRef]
- Zampetaki, A.; Kiechl, S.; Drozdov, I.; Willeit, P.; Mayr, U.; Prokopi, M.; Mayr, A.; Weger, S.; Oberhollenzer, F.; Bonora, E.; et al. Plasma microrna profiling reveals loss of endothelial mir-126 and other micrornas in type 2 diabetes. Circ. Res. 2010, 107, 810–817. [Google Scholar] [CrossRef]
- Ren, Y.; Bao, R.; Guo, Z.; Kai, J.; Cai, C.G.; Li, Z. miR-126-5p regulates H9c2 cell proliferation and apoptosis under hypoxic conditions by targeting IL-17A. Exp. Ther. Med. 2020, 21, 67. [Google Scholar] [CrossRef]
- Su, Q.; Liu, Y.; Lv, X.-W.; Dai, R.-X.; Yang, X.-H.; Kong, B.-H. LncRNA TUG1 mediates ischemic myocardial injury by targeting miR-132-3p/HDAC3 axis. Am. J. Physiol. Heart Circ. Physiol. 2020, 318, H332–H344. [Google Scholar] [CrossRef]
- Zhao, Z.; Du, S.; Shen, S.; Wang, L. microRNA-132 inhibits cardiomyocyte apoptosis and myocardial remodeling in myocardial infarction by targeting IL-1beta. J. Cell. Physiol. 2020, 235, 2710–2721. [Google Scholar] [CrossRef]
- Chen, L.; Wang, G.-Y.; Dong, J.-H.; Cheng, X.-J. MicroRNA-132 improves myocardial remodeling after myocardial infarction. Eur. Rev. Med. Pharmacol. Sci. 2019, 23, 6299–6306. [Google Scholar]
- Zhang, J.; Xu, H.; Gong, L.; Liu, L. MicroRNA-132 protects H9c2 cells against oxygen and glucose deprivation-evoked injury by targeting FOXO3A. J. Cell. Physiol. 2020, 235, 176–184. [Google Scholar] [CrossRef]
- Ma, T.; Chen, Y.; Chen, Y.; Meng, Q.; Sun, J.; Shao, L.; Yu, Y.; Huang, H.; Hu, Y.; Yang, Z.; et al. MicroRNA-132, Delivered by Mesenchymal Stem Cell-Derived Exosomes, Promote Angiogenesis in Myocardial Infarction. Stem Cells Int. 2018, 2018, 3290372. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shahrokhi, S.Z.; Saeidi, L.; Sadatamini, M.; Jafarzadeh, M.; Rahimipour, A.; Kazerouni, F. Can miR-145-5p be used as a marker in diabetic patients? Arch. Physiol. Biochem. 2020, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Yan, L.; Guo, N.; Cao, Y.; Zeng, S.; Wang, J.; Lv, F.; Wang, Y.; Cao, X. miRNA-145 inhibits myocardial infarction-induced apoptosis through autophagy via Akt3/mTOR signaling pathway in vitro and in vivo. Int. J. Mol. Med. 2018, 42, 1537–1547. [Google Scholar] [CrossRef] [Green Version]
- Liu, Z.; Fan, S.; Tao, B.; Pu, Y.; Xia, H.; Xu, L. microRNA-145 Protects against Myocardial Ischemia Reperfusion Injury via CaMKII-Mediated Antiapoptotic and Anti-Inflammatory Pathways. Oxid. Med. Cell. Longev. 2019, 2019, 8948657. [Google Scholar] [CrossRef] [PubMed]
- Yuan, M.; Zhang, L.; You, F.; Zhou, J.; Ma, Y.; Yang, F.; Tao, L. MiR-145-5p regulates hypoxia-induced inflammatory response and apoptosis in cardiomyocytes by targeting CD40. Mol. Cell. Biochem. 2017, 431, 123–131. [Google Scholar] [CrossRef]
- Qi, Z.; Li, S.; Su, Y.; Zhang, J.; Kang, Y.; Huang, Y.; Jin, F.; Xing, Q. Role of microRNA-145 in protection against myocardial ischemia/reperfusion injury in mice by regulating expression of GZMK with the treatment of sevoflurane. J. Cell. Physiol. 2019, 234, 16526–16539. [Google Scholar] [CrossRef]
- Higashi, K.; Yamada, Y.; Minatoguchi, S.; Baba, S.; Iwasa, M.; Kanamori, H.; Kawasaki, M.; Nishigaki, K.; Takemura, G.; Kumazaki, M.; et al. MicroRNA-145 repairs infarcted myocardium by accelerating cardiomyocyte autophagy. Am. J. Physiol. Heart Circ. Physiol. 2015, 309, H1813–H1826. [Google Scholar] [CrossRef] [Green Version]
- Song, H.-F.; He, S.; Li, S.-H.; Wu, J.; Yin, W.; Shao, Z.; Du, G.-Q.; Wu, J.; Li, J.; Weisel, R.D.; et al. Knock-out of MicroRNA 145 impairs cardiac fibroblast function and wound healing post-myocardial infarction. J. Cell. Mol. Med. 2020, 24, 9409–9419. [Google Scholar] [CrossRef]
- Iacomino, G.; Siani, A. Role of microRNAs in obesity and obesity-related diseases. Genes Nutr. 2017, 12, 23. [Google Scholar] [CrossRef]
- Yan, Y.; Dang, H.; Zhang, X.; Wang, X.; Liu, X. The protective role of MiR-206 in regulating cardiomyocytes apoptosis induced by ischemic injury by targeting PTP1B. Biosci. Rep. 2020, 40, BSR20191000. [Google Scholar] [CrossRef]
- Kong, F.; Jin, J.; Lv, X.; Han, Y.; Liang, X.; Gao, Y.; Duan, X. RETRACTED: Long noncoding RNA RMRP upregulation aggravates myocardial ischemia-reperfusion injury by sponging miR-206 to target ATG3 expression. Biomed. Pharmacother. 2019, 109, 716–725. [Google Scholar] [CrossRef]
- Zhai, C.; Qian, Q.; Tang, G.; Han, B.; Hu, H.; Yin, D.; Pan, H.; Zhang, S. MicroRNA-206 Protects against Myocardial Ischaemia-Reperfusion Injury in Rats by Targeting Gadd45beta. Mol. Cells 2017, 40, 916–924. [Google Scholar]
- Wu, L.; Dai, X.; Zhan, J.; Zhang, Y.; Zhang, H.; Zeng, S.; Xi, W.; Zhang, H. Profiling peripheral microRNAs in obesity and type 2 diabetes mellitus. APMIS 2015, 123, 580–585. [Google Scholar] [CrossRef]
- Liu, Y.; Zou, J.; Liu, X.; Zhang, Q. MicroRNA-138 attenuates myocardial ischemia reperfusion injury through inhibiting mitochondria-mediated apoptosis by targeting HIF1-α. Exp. Ther. Med. 2019, 18, 3325–3332. [Google Scholar] [CrossRef] [Green Version]
- Zhu, H.; Xue, H.; Jin, Q.-H.; Guo, J.; Chen, Y.-D. MiR-138 protects cardiac cells against hypoxia through modulation of glucose metabolism by targetting pyruvate dehydrogenase kinase 1. Biosci. Rep. 2017, 37, BSR20170296. [Google Scholar] [CrossRef] [Green Version]
- Mao, Q.; Liang, X.-L.; Zhang, C.-L.; Pang, Y.-H.; Lu, Y.-X. LncRNA KLF3-AS1 in human mesenchymal stem cell-derived exosomes ameliorates pyroptosis of cardiomyocytes and myocardial infarction through miR-138-5p/Sirt1 axis. Stem Cell Res. Ther. 2019, 10, 393. [Google Scholar] [CrossRef] [PubMed]
- Zhou, W.; Zhou, W.; Zeng, Q.; Xiong, J. MicroRNA-138 inhibits hypoxia-induced proliferation of endothelial progenitor cells via inhibition of HIF-1α-mediated MAPK and AKT signaling. Exp. Ther. Med. 2017, 13, 1017–1024. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lisa, K.T.; Gilpin, E.; Ahnve, S.; Henning, H.; Ross, J. Smoking status at the time of acute myocardial infarction and subsequent prognosis. Am. Heart J. 1985, 110, 535–541. [Google Scholar] [CrossRef]
- Symons, R.; Masci, P.G.; Francone, M.; Claus, P.; Barison, A.; Carbone, I.; Agati, L.; Galea, N.; Janssens, S.; Bogaert, J. Impact of active smoking on myocardial infarction severity in reperfused ST-segment elevation myocardial infarction patients: The smoker’s paradox revisited. Eur. Heart J. 2016, 37, 2756–2764. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fujita, Y.; Araya, J.; Ito, S.; Kobayashi, K.; Kosaka, N.; Yoshioka, Y.; Kadota, T.; Hara, H.; Kuwano, K.; Ochiya, T. Suppression of autophagy by extracellular vesicles promotes myofibroblast differentiation in COPD pathogenesis. J. Extracell. Vesicles 2015, 4, 28388. [Google Scholar] [CrossRef]
- Xu, L.; Deng, X. Tobacco-specific nitrosamine 4-(methylnitrosamino)-1-(3-pyridyl)-1-butanone induces phosphorylation of mu- and m-calpain in association with increased secretion, cell migration, and invasion. J. Biol. Chem. 2004, 279, 53683–53690. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, F.; Yin, Z.; Yang, L.; Fan, J.; Xu, J.; Jin, Y.; Yu, J.; Zhang, D.; Yang, G. Smoking Induced Extracellular Vesicles Release and Their Distinct Properties in Non-Small Cell Lung Cancer. J. Cancer 2019, 10, 3435–3443. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Baek, R.; Varming, K.; Jorgensen, M.M. Does smoking, age or gender affect the protein phenotype of extracellular vesicles in plasma? Transfus. Apher. Sci. 2016, 55, 44–52. [Google Scholar] [CrossRef] [Green Version]
- Mobarrez, F.; Antoniewicz, L.; Bosson, J.A.; Kuhl, J.; Pisetsky, D.S.; Lundbäck, M. The Effects of Smoking on Levels of Endothelial Progenitor Cells and Microparticles in the Blood of Healthy Volunteers. PLoS ONE 2014, 9, e90314. [Google Scholar] [CrossRef] [Green Version]
- Enjeti, A.K.; Ariyarajah, A.; D’Crus, A.; Seldon, M.; Lincz, L.F. Circulating microvesicle number, function and small RNA content vary with age, gender, smoking status, lipid and hormone profiles. Thromb. Res. 2017, 156, 65–72. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Benedikter, B.J.; Volgers, C.; Van Eijck, P.H.; Wouters, E.F.; Savelkoul, P.H.; Reynaert, N.L.; Haenen, G.R.; Rohde, G.G.; Weseler, A.R.; Stassen, F.R. Cigarette smoke extract induced exosome release is mediated by depletion of exofacial thiols and can be inhibited by thiol-antioxidants. Free Radic. Biol. Med. 2017, 108, 334–344. [Google Scholar] [CrossRef]
- Serban, K.A.; Rezania, S.; Petrusca, D.N.; Poirier, C.; Cao, D.; Justice, M.J.; Patel, M.; Tsvetkova, I.; Kamocki, K.; Mikosz, A.; et al. Structural and functional characterization of endothelial microparticles released by cigarette smoke. Sci. Rep. 2016, 6, 31596. [Google Scholar] [CrossRef]
- Cordazzo, C.; Petrini, S.; Neri, T.; Lombardi, S.; Carmazzi, Y.; Pedrinelli, R.; Paggiaro, P.; Celi, A. Rapid shedding of proinflammatory microparticles by human mononuclear cells exposed to cigarette smoke is dependent on Ca2+ mobilization. Inflamm. Res. 2014, 63, 539–547. [Google Scholar] [CrossRef]
- Li, C.-J.; Liu, Y.; Chen, Y.; Yu, D.; Williams, K.J.; Liu, M.-L. Novel Proteolytic Microvesicles Released from Human Macrophages after Exposure to Tobacco Smoke. Am. J. Pathol. 2013, 182, 1552–1562. [Google Scholar] [CrossRef] [Green Version]
- Andres, J.; Smith, L.C.; Murray, A.; Jin, Y.; Businaro, R.; Laskin, J.D.; Laskin, D.L. Role of extracellular vesicles in cell-cell communication and inflammation following exposure to pulmonary toxicants. Cytokine Growth Factor Rev. 2020, 51, 12–18. [Google Scholar] [CrossRef] [PubMed]
- Benedikter, B.J.; Bouwman, F.G.; Heinzmann, A.C.A.; Vajen, T.; Mariman, E.C.; Wouters, E.F.M.; Savelkoul, P.H.M.; Koenen, R.R.; Rohde, G.G.U.; Van Oerle, R.; et al. Proteomic analysis reveals procoagulant properties of cigarette smoke-induced extracellular vesicles. J. Extracell. Vesicles 2019, 8, 1585163. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stassen, F.R.M.; van Eijck, P.H.; Savelkoul, P.H.M.; Wouters, E.F.M.; Rohde, G.G.U.; Briede, J.J.; Raynaert, N.L.; de Kok, T.M.; Benedikter, B.J. Cell type- and exposure-specific modulation of cd63/cd81-positive and tissue factor-positive extracellular vesicle release in response to respiratory toxicants. Oxid. Med. Cell. Longev. 2019, 2019, 5204218. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, M.; Yu, D.; Williams, K.J.; Liu, M.-L. Tobacco smoke induces the generation of procoagulant microvesicles from human monocytes/macrophages. Arter. Thromb. Vasc. Biol. 2010, 30, 1818–1824. [Google Scholar] [CrossRef] [PubMed]
- Haque, S.; Kodidela, S.; Sinha, N.; Kumar, P.; Cory, T.J.; Kumar, S. Differential packaging of inflammatory cytokines/ chemokines and oxidative stress modulators in U937 and U1 macrophages-derived extracellular vesicles upon exposure to tobacco constituents. PLoS ONE 2020, 15, e0233054. [Google Scholar] [CrossRef] [PubMed]
- Neri, M.; Fineschi, V.; Paolo, M.; Pomara, C.; Riezzo, I.; Turillazzi, E.; Cerretani, D. Cardiac oxidative stress and inflammatory cytokines response after myocardial infarction. Curr. Vasc. Pharmacol. 2015, 13, 26–36. [Google Scholar] [CrossRef]
- Toldo, S.; Mauro, A.G.; Cutter, Z.S.; Abbate, A. Inflammasome, pyroptosis, and cytokines in myocardial ischemia-reperfusion injury. Am. J. Physiol. Heart Circ. Physiol. 2018, 315, H1553–H1568. [Google Scholar] [CrossRef]
- Fujita, Y.; Araya, J.; Ochiya, T. Extracellular vesicles in smoking-related lung diseases. Oncotarget 2015, 6, 43144–43145. [Google Scholar] [CrossRef]
- Díaz, I.; Calderón-Sánchez, E.M.; Del Toro, R.; Avila-Medina, J.; Pedro, E.S.D.R.-D.; Domínguez-Rodríguez, A.; Rosado, J.A.; Hmadcha, A.; Ordoñez, A.; Smani, T. miR-125a, miR-139 and miR-324 contribute to Urocortin protection against myocardial ischemia-reperfusion injury. Sci. Rep. 2017, 7, 8898. [Google Scholar] [CrossRef]
- Wu, Q.; Shang, Y.; Bai, Y.; Wu, Y.; Wang, H.; Shen, T. Sufentanil preconditioning protects against myocardial ischemia/reperfusion injury via miR-125a/DRAM2 axis. Cell Cycle 2021, 20, 383–391. [Google Scholar] [CrossRef]
- Wu, X.Q.; Tian, X.-Y.; Wang, Z.-W.; Wu, X.; Wang, J.-P.; Yan, T.-Z. miR-191 secreted by platelet-derived microvesicles induced apoptosis of renal tubular epithelial cells and participated in renal ischemia-reperfusion injury via inhibiting CBS. Cell Cycle 2019, 18, 119–129. [Google Scholar] [CrossRef] [Green Version]
- Pan, W.; Wang, L.; Zhang, X.-D.; Zhang, H.; Zhang, J.; Wang, G.; Xu, P.; Zhang, Y.; Hu, P.; Du, R.-L.; et al. Hypoxia-induced microRNA-191 contributes to hepatic ischemia/reperfusion injury through the ZONAB/Cyclin D1 axis. Cell Death Differ. 2019, 26, 291–305. [Google Scholar] [CrossRef]
- Bao, M.-H.; Feng, X.; Zhang, Y.-W.; Lou, X.-Y.; Cheng, Y.; Zhou, H. Let-7 in Cardiovascular diseases, heart development and cardiovascular differentiation from stem cells. Int. J. Mol. Sci. 2013, 14, 23086–23102. [Google Scholar] [CrossRef] [Green Version]
- Badrnya, S.; Assinger, A.; Baumgartner, R. Smoking alters circulating plasma microvesicle pattern and microRNA signatures. Thromb. Haemost. 2014, 112, 128–136. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhong, Y.; Li, Y.-P.; Yin, Y.-Q.; Hu, B.-L.; Gao, H. Dexmedetomidine inhibits pyroptosis by down-regulating miR-29b in myocardial ischemia reperfusion injury in rats. Int. Immunopharmacol. 2020, 86, 106768. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Cheng, L.; Xu, L.; Zhang, Y.; Yang, Y.; Fu, Q.; Mi, W.; Li, H. The lncRNA, H19 Mediates the protective effect of hypoxia postconditioning against hypoxia-reoxygenation injury to senescent cardiomyocytes by targeting microRNA-29b-3p. Shock 2019, 52, 249–256. [Google Scholar] [CrossRef]
- Wang, P.-P.; Zhang, Y.-J.; Xie, T.; Sun, J.; Wang, X.-D. MiR-223 promotes cardiomyocyte apoptosis by inhibiting Foxo3a expression. Eur. Rev. Med. Pharmacol. Sci. 2018, 22, 6119–6126. [Google Scholar]
- Sundar, I.K.; Li, D.; Rahman, I. Small RNA-sequence analysis of plasma-derived extracellular vesicle miRNAs in smokers and patients with chronic obstructive pulmonary disease as circulating biomarkers. J. Extracell. Vesicles 2019, 8, 1684816. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McDonough, S.R.; Rahman, I.; Sundar, I.K. Recent updates on biomarkers of exposure and systemic toxicity in e-cigarette users and EVALI. Am. J. Physiol. Lung Cell. Mol. Physiol. 2021, 320, L661–L679. [Google Scholar] [CrossRef]
- Singh, K.P.; Maremanda, K.P.; Li, D.; Rahman, I. Exosomal microRNAs are novel circulating biomarkers in cigarette, waterpipe smokers, E-cigarette users and dual smokers. BMC Med. Genom. 2020, 13, 128. [Google Scholar] [CrossRef]
- Martínez, M.C.; Andriantsitohaina, R. Extracellular vesicles in metabolic syndrome. Circ. Res. 2017, 120, 1674–1686. [Google Scholar] [CrossRef]
- Georgescu, A.; Alexandru, N.; Andrei, E.; Titorencu, I.; Dragan, E.; Tarziu, C.; Ghiorghe, S.; Badila, E.; Bartos, D.; Popov, D. Circulating microparticles and endothelial progenitor cells in atherosclerosis: Pharmacological effects of irbesartan. J. Thromb. Haemost. 2012, 10, 680–691. [Google Scholar] [CrossRef]
- Nijiati, M.; Gao, Y.; Abudureheman, Z.; Yu, X.; Li, G. Relationship between the level of circulating endothelial micro-particles in the blood and blood lipid content in uyghur and han patients with acute coronary syndrome. Clin. Lab. 2015, 61, 1071–1075. [Google Scholar] [CrossRef]
- Pienimaeki-Roemer, A.; Fischer, A.; Tafelmeier, M.; Orsó, E.; Konovalova, T.; Böttcher, A.; Liebisch, G.; Reidel, A.; Schmitz, G. High-density lipoprotein 3 and apolipoprotein A-I alleviate platelet storage lesion and release of platelet extracellular vesicles. Transfusion 2014, 54, 2301–2314. [Google Scholar] [CrossRef]
- Hjuler Nielsen, M.; Irvine, H.; Vedel, S.; Raungaard, B.; Beck-Nielsen, H.; Handberg, A. Elevated atherosclerosis-related gene expression, monocyte activation and microparticle-release are related to increased lipoprotein-associated oxidative stress in familial hypercholesterolemia. PLoS ONE 2015, 10, e0121516. [Google Scholar] [CrossRef]
- Xia, J.; Qu, Y.; Yin, C.; Xu, N. Preoperative rosuvastatin protects patients with coronary artery disease undergoing noncardiac surgery. Cardiology 2015, 131, 30–37. [Google Scholar] [CrossRef]
- Mannacio, V.A.; Iorio, D.; DE Amicis, V.; Di Lello, F.; Musumeci, F. Effect of rosuvastatin pretreatment on myocardial damage after coronary surgery: A randomized trial. J. Thorac. Cardiovasc. Surg. 2008, 136, 1541–1548. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tomoda, H.; Elgendy, I.Y.; Mahmoud, A.N.; Bavry, A.A.; Wang, S.; Peng, D.-Q.; Casadei, B.; Collins, R.; Zheng, Z. Perioperative Rosuvastatin in Cardiac Surgery. N. Engl. J. Med. 2016, 375, 1744. [Google Scholar] [CrossRef]
- Suades, R.; Padró, T.; Alonso, R.; Mata, P.; Badimon, L. Lipid-lowering therapy with statins reduces microparticle shedding from endothelium, platelets and inflammatory cells. Thromb. Haemost. 2013, 110, 366–377. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nomura, S.; Inami, N.; Shouzu, A.; Omoto, S.; Kimura, Y.; Takahashi, N.; Tanaka, A.; Urase, F.; Maeda, Y.; Ohtani, H.; et al. The effects of pitavastatin, eicosapentaenoic acid and combined therapy on platelet-derived microparticles and adiponectin in hyperlipidemic, diabetic patients. Platelets 2009, 20, 16–22. [Google Scholar] [CrossRef]
- Camargo, L.; França, C.N.; Izar, M.; Bianco, H.; Lins, L.; Barbosa, S.; Pinheiro, L.; Fonseca, F. Effects of simvastatin/ezetimibe on microparticles, endothelial progenitor cells and platelet aggregation in subjects with coronary heart disease under antiplatelet therapy. Braz. J. Med. Biol. Res. 2014, 47, 432–437. [Google Scholar] [CrossRef] [PubMed]
- Huang, B.; Cheng, Y.; Xie, Q.; Lin, G.; Wu, Y.; Feng, Y.; Gao, J.; Xu, D. Effect of 40 mg versus 10 mg of atorvastatin on oxidized low-density lipoprotein, high-sensitivity c-reactive protein, circulating endothelial-derived microparticles, and endothelial progenitor cells in patients with ischemic cardiomyopathy. Clin. Cardiol. 2012, 35, 125–130. [Google Scholar] [CrossRef]
- Verbree-Willemsen, L.; Zhang, Y.-N.; Gijsberts, C.M.; Schoneveld, A.H.; Wang, J.-W.; Lam, C.S.; Vernooij, F.; Bots, M.L.; Peelen, L.M.; Grobbee, D.E.; et al. LDL extracellular vesicle coagulation protein levels change after initiation of statin therapy. Findings from the METEOR trial. Int. J. Cardiol. 2018, 271, 247–253. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kulshreshtha, A.; Singh, S.; Ahmad, M.; Khanna, K.; Ahmad, T.; Agrawal, A.; Ghosh, B. Simvastatin mediates inhibition of exosome synthesis, localization and secretion via multicomponent interventions. Sci. Rep. 2019, 9, 16373. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kocsis, G.F.; Pipis, J.; Fekete, V.; Kovács-Simon, A.; Odendaal, L.; Molnár, E.; Giricz, Z.; Janáky, T.; Van Rooyen, J.; Csont, T.; et al. Lovastatin interferes with the infarct size-limiting effect of ischemic preconditioning and postconditioning in rat hearts. Am. J. Physiol. Heart Circ. Physiol. 2008, 294, H2406–H2409. [Google Scholar] [CrossRef]
- Baietti, M.F.; Zhang, Z.; Mortier, E.; Melchior, A.; Degeest, G.; Geeraerts, A.; Ivarsson, Y.; Depoortere, F.; Coomans, C.; Vermeiren, E.; et al. Syndecan-syntenin-ALIX regulates the biogenesis of exosomes. Nat. Cell Biol. 2012, 14, 677–685. [Google Scholar] [CrossRef]
- Zhang, Y.; Liu, D.; Chen, X.; Li, J.; Li, L.; Bian, Z.; Sun, F.; Lu, J.; Yin, Y.; Cai, X.; et al. Secreted monocytic mir-150 enhances targeted endothelial cell migration. Mol. Cell 2010, 39, 133–144. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tang, Y.; Wang, Y.; Park, K.-M.; Hu, Q.; Teoh, J.-P.; Broskova, Z.; Ranganathan, P.; Jayakumar, C.; Li, J.; Su, H.; et al. MicroRNA-150 protects the mouse heart from ischaemic injury by regulating cell death. Cardiovasc. Res. 2015, 106, 387–397. [Google Scholar] [CrossRef] [Green Version]
- Carrick, D.; Haig, C.; Maznyczka, A.M.; Carberry, J.; Mangion, K.; Ahmed, N.; Petrie, M.C.; Eteiba, H.; Lindsay, M.; Hood, S.; et al. Hypertension, microvascular pathology, and prognosis after an acute myocardial infarction. Hypertension 2018, 72, 720–730. [Google Scholar] [CrossRef]
- Galiñanes, M. Role of clinical pathologies in myocardial injury following ischaemia and reperfusion. Cardiovasc. Res. 2004, 61, 512–521. [Google Scholar] [CrossRef]
- Mølgaard, S.; Faricelli, B.; Salomonsson, M.; Engstrøm, T.; Treiman, M. Increased myocardial vulnerability to ischemia–reperfusion injury in the presence of left ventricular hypertrophy. J. Hypertens. 2016, 34, 513–523. [Google Scholar] [CrossRef]
- Kang, D.G.; Jeong, M.H.; Ahn, Y.; Chae, S.C.; Hur, S.H.; Hong, T.J.; Kim, Y.J.; Seong, I.W.; Chae, J.K.; Rhew, J.Y.; et al. Clinical effects of hypertension on the mortality of patients with acute myocardial infarction. J. Korean Med. Sci. 2009, 24, 800–806. [Google Scholar] [CrossRef]
- Reinstadler, S.J.; Stiermaier, T.; Eitel, C.; Saad, M.; Metzler, B.; de Waha, S.; Fuernau, G.; Desch, s.; Thiele, H.; Eitel, I. Antecedent hypertension and myocardial injury in patients with reperfused ST-elevation myocardial infarction. J. Cardiovasc. Magn. Reson. 2017, 18, 80. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Powers, S.K.; Lennon, S.L.; Quindry, J.; Mehta, J.L. Exercise and cardioprotection. Curr. Opin. Cardiol. 2002, 17, 495–502. [Google Scholar] [CrossRef] [PubMed]
- Powers, S.K.; Quindry, J.C.; Kavazis, A.N. Exercise-induced cardioprotection against myocardial ischemia–reperfusion injury. Free Radic. Biol. Med. 2008, 44, 193–201. [Google Scholar] [CrossRef]
- Lee, Y.; Min, K.; Talbert, E.E.; Kavazis, A.N.; Smuder, A.J.; Willis, W.T.; Powers, S.K. Exercise protects cardiac mitochondria against ischemia–reperfusion injury. Med. Sci. Sports Exerc. 2012, 44, 397–405. [Google Scholar] [CrossRef]
- Guo, Y.; Chen, J.; Qiu, H. Novel mechanisms of exercise-induced cardioprotective factors in myocardial infarction. Front. Physiol. 2020, 11, 199. [Google Scholar] [CrossRef] [PubMed]
- Michelsen, M.M.; Støttrup, N.B.; Schmidt, M.R.; Løfgren, B.; Jensen, R.V.; Tropak, M.; St-Michel, E.J.; Redington, A.N.; Bøtker, H.E. Exercise-induced cardioprotection is mediated by a bloodborne, transferable factor. Basic Res. Cardiol. 2012, 107, 260. [Google Scholar] [CrossRef] [PubMed]
- Bei, Y.; Xu, T.; Lu, D.; Yu, P.; Xu, J.; Che, L.; Das, A.; Tigges, J.; Toxavidis, V.; Ghiran, I.; et al. Exercise-induced circulating extracellular vesicles protect against cardiac ischemia–reperfusion injury. Basic Res. Cardiol. 2017, 112, 38. [Google Scholar] [CrossRef]
- Oliveira, G.; Porto, W.; Palu, C.; Pereira, L.M.; Petriz, B.; Almeida, J.A.; Viana, J.; Filho, N.N.A.; Franco, O.L.; Pereira, R. Effects of acute aerobic exercise on rats serum extracellular vesicles diameter, concentration and small rnas content. Front. Physiol. 2018, 9, 532. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wiklander, O.P.B.; Nordin, J.Z.; O’Loughlin, A.; Gustafsson, Y.; Corso, G.; Mäger, I.; Vader, P.; Lee, Y.; Sork, H.; Seow, Y.; et al. Extracellular vesicle in vivo biodistribution is determined by cell source, route of administration and targeting. J. Extracell. Vesicles 2015, 4, 26316. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Whitham, M.; Parker, B.L.; Friedrichsen, M.; Hingst, J.R.; Hjorth, M.; Hughes, W.; Egan, C.L.; Cron, L.; Watt, K.; Kuchel, R.P.; et al. Extracellular vesicles provide a means for tissue crosstalk during exercise. Cell Metab. 2018, 27, 237–251.e4. [Google Scholar] [CrossRef] [Green Version]
- Wilhelm, E.N.; Mourot, L.; Rakobowchuk, M. Exercise-derived microvesicles: A review of the literature. Sports Med. 2018, 48, 2025–2039. [Google Scholar] [CrossRef] [Green Version]
- Frühbeis, C.; Helmig, S.; Tug, S.; Simon, P.; Krämer-Albers, E.-M. Physical exercise induces rapid release of small extracellular vesicles into the circulation. J. Extracell. Vesicles 2015, 4, 28239. [Google Scholar] [CrossRef] [Green Version]
- Hou, Z.; Qin, X.; Hu, Y.; Zhang, X.; Li, G.; Wu, J.; Li, J.; Sha, J.; Chen, J.; Xia, J.; et al. Longterm exercise-derived exosomal mir-342-5p. Circ. Res. 2019, 124, 1386–1400. [Google Scholar] [CrossRef] [PubMed]
- Barber, J.L.; Zellars, K.N.; Barringhaus, K.G.; Bouchard, C.; Spinale, F.G.; Sarzynski, M.A. The effects of regular exercise on circulating cardiovascular-related micrornas. Sci. Rep. 2019, 9, 7527. [Google Scholar] [CrossRef] [PubMed]
- Mooren, F.C.; Viereck, J.; Krüger, K.; Thum, T. Circulating micrornas as potential biomarkers of aerobic exercise capacity. Am. J. Physiol. Heart Circ. Physiol. 2014, 306, H557–H563. [Google Scholar] [CrossRef]
- Domańska-Senderowska, D.; Laguette, M.-J.N.; Jegier, A.; Cieszczyk, P.; September, A.V.; Brzeziańska-Lasota, E. MicroRNA profile and adaptive response to exercise training: A review. Endoscopy 2019, 40, 227–235. [Google Scholar] [CrossRef] [PubMed]
- Terlecki-Zaniewicz, L.; Lämmermann, I.; Latreille, J.; Bobbili, M.R.; Pils, V.; Schosserer, M.; Weinmüllner, R.; Dellago, H.; Skalicky, S.; Pum, D.; et al. Small extracellular vesicles and their miRNA cargo are anti-apoptotic members of the senescence-associated secretory phenotype. Aging 2018, 10, 1103–1132. [Google Scholar] [CrossRef]
- Huang, X.; Yuan, T.; Tschannen, M.; Sun, Z.; Jacob, H.; Du, M.; Liang, M.; Dittmar, R.L.; Liu, Y.; Liang, M.; et al. Characterization of human plasma-derived exosomal RNAs by deep sequencing. BMC Genom. 2013, 14, 319. [Google Scholar] [CrossRef] [Green Version]
- Shi, J.; Bei, Y.; Kong, X.; Liu, X.; Lei, Z.; Xu, T.; Wang, H.; Xuan, Q.; Chen, P.; Xu, J.; et al. miR-17-3p contributes to exercise-induced cardiac growth and protects against myocardial ischemia-reperfusion injury. Theranostics 2017, 7, 664–676. [Google Scholar] [CrossRef]
- Peng, Y.; Zhao, J.-L.; Peng, Z.-Y.; Xu, W.-F.; Yu, G.-L. Exosomal miR-25-3p from mesenchymal stem cells alleviates myocardial infarction by targeting pro-apoptotic proteins and EZH2. Cell Death Dis. 2020, 11, 317. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Jiang, M.; Deng, S.; Lu, J.; Huang, H.; Zhang, Y.; Gong, P.; Shen, X.; Ruan, H.; Jin, M.; et al. miR-93-5p-containing exosomes treatment attenuates acute myocardial infarction-induced myocardial damage. Mol. Ther. Nucleic Acids 2018, 11, 103–115. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gomes, C.P.; Oliveira, G.P., Jr.; Madrid, B.; Almeida, J.A.; Franco, O.L.; Pereira, R.W. Circulating miR-1, miR-133a, and miR-206 levels are increased after a half-marathon run. Biomarkers 2014, 19, 585–589. [Google Scholar] [CrossRef]
- Clauss, S.; Wakili, R.; Hildebrand, B.; Kaab, S.; Hoster, E.; Klier, I.; Martens, E.; Hanley, A.; Hanssen, H.; Halle, M.; et al. MicroRNAs as Biomarkers for acute atrial remodeling in marathon runners (the mirathon study—A sub-study of the Munich marathon study). PLoS ONE 2016, 11, e0148599. [Google Scholar] [CrossRef] [PubMed]
- Safdar, A.; Saleem, A.; Tarnopolsky, M.A. The potential of endurance exercise-derived exosomes to treat metabolic diseases. Nat. Rev. Endocrinol. 2016, 12, 504–517. [Google Scholar] [CrossRef]
- Eichner, N.Z.M.; Erdbrügger, U.; Malin, S.K. Extracellular vesicles: A novel target for exercise-mediated reductions in type 2 diabetes and cardiovascular disease risk. J. Diabetes Res. 2018, 2018, 7807245. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Safdar, A.; Tarnopolsky, M.A. Exosomes as mediators of the systemic adaptations to endurance exercise. Cold Spring Harb. Perspect. Med. 2018, 8, a029827. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Go, A.S.; Mozaffarian, D.; Roger, V.L.; Benjamin, E.J.; Berry, J.D.; Blaha, M.J.; Dai, S.; Ford, E.S.; Fox, C.S.; Franco, S.; et al. Heart disease and stroke statistics—2014 update: A report from the American Heart Association. Circulation 2014, 129, e28–e292. [Google Scholar] [CrossRef] [Green Version]
- Gustafson, C.M.; Shepherd, A.J.; Miller, V.M.; Jayachandran, M. Age- and sex-specific differences in blood-borne microvesicles from apparently healthy humans. Biol. Sex Differ. 2015, 6, 10. [Google Scholar] [CrossRef] [Green Version]
- Kobayashi, Y.; Eguchi, A.; Tempaku, M.; Honda, T.; Togashi, K.; Iwasa, M.; Hasegawa, H.; Takei, Y.; Sumida, Y.; Taguchi, O. Circulating extracellular vesicles are associated with lipid and insulin metabolism. Am. J. Physiol. Endocrinol. Metab. 2018, 315, E574–E582. [Google Scholar] [CrossRef]
- Noulsri, E.; Palasuwan, A. Effects of donor age, donor sex, blood-component processing, and storage on cell-derived microparticle concentrations in routine blood-component preparation. Transfus. Apher. Sci. 2018, 57, 587–592. [Google Scholar] [CrossRef]
- Ma, F.; Liu, H.; Shen, Y.; Zhang, Y.; Pan, S. Platelet-derived microvesicles are involved in cardio-protective effects of remote preconditioning. Int. J. Clin. Exp. Pathol. 2015, 8, 10832–10839. [Google Scholar] [PubMed]
- Tushuizen, M.E.; Diamant, M.; Sturk, A.; Nieuwland, R. Cell-derived microparticles in the pathogenesis of cardiovascular disease: Friend or foe? Arterioscler. Thromb. Vasc. Biol. 2011, 31, 4–9. [Google Scholar] [CrossRef] [Green Version]
- Bammert, T.D.; Hijmans, J.G.; Kavlich, P.J.; Lincenberg, G.M.; Reiakvam, W.R.; Fay, R.T.; Greiner, J.J.; Stauffer, B.L.; DeSouze, C.A. Influence of sex on the number of circulating endothelial microparticles and microRNA expression in middle-aged adults. Exp. Physiol. 2017, 102, 894–900. [Google Scholar] [CrossRef] [PubMed]
- El Khoudary, S.R.; Aggarwal, B.; Beckie, T.M.; Hodis, H.N.; Johnson, A.E.; Langer, R.D.; Limacher, M.C.; Manson, J.E.; Stefanick, M.L.; Allison, M.A.; et al. Menopause Transition and cardiovascular disease risk: Implications for timing of early prevention: A scientific statement from the american heart association. Circulation 2020, 142, e506–e532. [Google Scholar] [CrossRef]
- Rank, A.; Nieuwland, R.; Roesner, S.; Nikolajek, K.; Hiller, E.; Toth, B. Climacteric lowers plasma levels of platelet-derived microparticles: A pilot study in pre- versus postmenopausal women. Acta Haematol. 2012, 128, 53–59. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, J.; Li, H.; Fan, B.; Xu, W.; Zhang, X. Extracellular vesicles in normal pregnancy and pregnancy-related diseases. J. Cell. Mol. Med. 2020, 24, 4377–4388. [Google Scholar] [CrossRef] [Green Version]
- Gill, M.; Motta-Mejia, C.; Kandzija, N.; Cooke, W.; Zhang, W.; Cerdeira, A.S.; Bastie, C.; Redman, C.; Vatish, M. Placental syncytiotrophoblast-derived extracellular vesicles carry active nep (neprilysin) and are increased in preeclampsia. Hypertension 2019, 73, 1112–1119. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Takov, K.; He, Z.; Johnston, H.; Timms, J.; Guillot, P.V.; Yellon, D.; Davidson, S.M. Small extracellular vesicles secreted from human amniotic fluid mesenchymal stromal cells possess cardioprotective and promigratory potential. Basic Res. Cardiol. 2020, 115, 26. [Google Scholar] [CrossRef]
- Li, J.; Umar, S.; Iorga, A.; Youn, J.-Y.; Wang, Y.; Regitz-Zagrosek, V.; Cai, H.; Eghbali, M. Cardiac vulnerability to ischemia/reperfusion injury drastically increases in late pregnancy. Basic Res. Cardiol. 2012, 107, 271. [Google Scholar] [CrossRef] [Green Version]
- Panagiotou, N.; Neytchev, O.; Selman, C.; Shiels, P.G. Extracellular vesicles, ageing, and therapeutic interventions. Cells 2018, 7, 110. [Google Scholar] [CrossRef] [Green Version]
- Chen, G.H.; Xu, C.-S.; Zhang, J.; Li, Q.; Cui, H.-H.; Li, X.-D.; Chang, L.; Tang, R.-J.; Xu, J.-Y.; Tian, X.-Q.; et al. Inhibition of miR-128-3p by tongxinluo protects human cardiomyocytes from ischemia/reperfusion injury via upregulation of p70s6k1/p-p70s6k1. Front. Pharmacol. 2017, 8, 775. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, T.; Luo, Y.; Wang, J.; Zhang, N.; Gu, C.; Li, L.; Qian, D.; Cai, W.; Fan, J.; Yin, G. Exosomal miRNA-128-3p from mesenchymal stem cells of aged rats regulates osteogenesis and bone fracture healing by targeting Smad5. J. Nanobiotechnol. 2020, 18, 47. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rossello, X.; Yellon, D.M. The RISK pathway and beyond. Basic Res. Cardiol. 2018, 113, 2. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Merhi-Soussi, F.; Kwak, B.; Magne, D.; Chadjichristos, C.; Berti, M.; Pelli, G.; James, R.W.; Mach, F.; Gabay, C. Interleukin-1 plays a major role in vascular inflammation and atherosclerosis in male apolipoprotein E-knockout mice. Cardiovasc. Res. 2005, 66, 583–593. [Google Scholar] [CrossRef] [Green Version]
- Gomes de Andrade, G.; Cechinel, L.R.; Bertoldi, K.; Galvão, F.; Worm, P.V.; Siqueira, I.R. The Aging Process Alters IL-1beta and CD63 levels differently in extracellular vesicles obtained from the plasma and cerebrospinal fluid. Neuroimmunomodulation 2018, 25, 18–22. [Google Scholar] [CrossRef]
- Zhu, J.; Huang, J.; Dai, D.; Wang, X.; Gao, J.; Han, W.; Zhang, R. Recombinant human interleukin-1 receptor antagonist treatment protects rats from myocardial ischemia–reperfusion injury. Biomed. Pharmacother. 2019, 111, 1–5. [Google Scholar] [CrossRef]
- Toldo, S.; Schatz, A.M.; Mezzaroma, E.; Chawla, R.; Stallard, T.W.; Stallard, W.C.; Jahangiri, A.; Van Tassell, B.W.; Abbate, A. Recombinant human interleukin-1 receptor antagonist provides cardioprotection during myocardial ischemia reperfusion in the mouse. Cardiovasc. Drugs Ther. 2012, 26, 273–276. [Google Scholar] [CrossRef]
- Grothusen, C.; Hagemann, A.; Attmann, T.; Braesen, J.; Broch, O.; Cremer, J.; Schoettler, J. Impact of an interleukin-1 receptor antagonist and erythropoietin on experimental myocardial ischemia/reperfusion injury. Sci. World J. 2012, 2012, 737585. [Google Scholar] [CrossRef] [Green Version]
- Silvain, J.; Kerneis, M.; Zeitouni, M.; Lattuca, B.; Mertens, E.; Procopi, N.; Suc, G.; Salloum, T.; Frisdal, E.; Le Goff, W.; et al. Interleukin-1Beta and risk of premature death and MACE in patients with myocardial infarction. J. Am. Coll. Cardiol. 2020, 41, ehaa946.1697. [Google Scholar] [CrossRef]
- Fafian-Labora, J.A.; Lesende-Rodriguez, I.; Fernández-Pernas, P.; Sangiao-Alvarellos, S.; Monserrat, L.; Arntz, O.J.; Van De Loo, F.A.J.; Mateos, J.; Arufe, M.C. Effect of age on pro-inflammatory miRNAs contained in mesenchymal stem cell-derived extracellular vesicles. Sci. Rep. 2017, 7, 43923. [Google Scholar] [CrossRef] [Green Version]
- Olivieri, F.; Rippo, M.R.; Prattichizzo, F.; Babini, L.; Graciotti, L.; Recchioni, R.; Procopio, A.D. Toll like receptor signaling in “inflammaging”: MicroRNA as new players. Immun. Ageing 2013, 10, 11. [Google Scholar] [CrossRef] [Green Version]
- Lopez-Otin, C.; Blasco, M.A.; Partridge, L.; Serrano, M.; Kroemer, G. The hallmarks of aging. Cell 2013, 153, 1194–1217. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- González-Montero, J.; Brito, R.; Gajardo, A.I.; Rodrigo, R. Myocardial reperfusion injury and oxidative stress: Therapeutic opportunities. World J. Cardiol. 2018, 10, 74–86. [Google Scholar] [CrossRef] [PubMed]
- Yoshida, M.; Satoh, A.; Lin, J.B.; Mills, K.F.; Sasaki, Y.; Rensing, N.; Wong, M.; Apte, R.S.; Imai, S.-I. Extracellular vesicle-contained enampt delays aging and extends lifespan in mice. Cell Metab. 2019, 30, 329–342.e5. [Google Scholar] [CrossRef] [PubMed]
- Chouchani, E.T.; Pell, V.R.; Gaude, E.; Sundier, S.Y.; Robb, E.L.; Logan, A.; Nadtochiy, S.M.; Ord, E.N.J.; Smith, A.C.; Eyassu, F.; et al. Ischaemic accumulation of succinate controls reperfusion injury through mitochondrial ROS. Nature 2014, 515, 431–435. [Google Scholar] [CrossRef] [Green Version]
- Yoshino, J.; Mills, K.F.; Yoon, M.J.; Imai, S.-I. Nicotinamide Mononucleotide, a Key NAD+ Intermediate, Treats the Pathophysiology of Diet- and Age-Induced Diabetes in Mice. Cell Metab. 2011, 14, 528–536. [Google Scholar] [CrossRef] [Green Version]
- Imai, S.-I.; Guarente, L. NAD+ and sirtuins in aging and disease. Trends Cell Biol. 2014, 24, 464–471. [Google Scholar] [CrossRef]
- Imai, S.-I. Nicotinamide phosphoribosyltransferase (nampt): A link between nad biology, metabolism, and diseases. Curr. Pharm. Des. 2009, 15, 20–28. [Google Scholar] [CrossRef] [Green Version]
- Eitan, E.; Green, J.; Bodogai, M.; Mode, N.A.; Bæk, R.; Jørgensen, M.M.; Freeman, D.W.; Witwer, K.; Zonderman, A.B.; Biragyn, A.; et al. Age-related changes in plasma extracellular vesicle characteristics and internalization by leukocytes. Sci. Rep. 2017, 7, 1342. [Google Scholar] [CrossRef] [Green Version]
- Deschamps, A.; Murphy, E.; Sun, J. Estrogen receptor activation and cardioprotection in ischemia reperfusion injury. Trends Cardiovasc. Med. 2010, 20, 73–78. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lazo, S.; Hooten, N.N.; Green, J.; Eitan, E.; Mode, N.A.; Liu, Q.; Zonderman, A.B.; Ezike, N.; Mattson, M.P.; Ghosh, P.; et al. Mitochondrial DNA in extracellular vesicles declines with age. Aging Cell 2021, 20, e13283. [Google Scholar] [CrossRef] [PubMed]
- Picca, A.; Guerra, F.; Calvani, R.; Bucci, C.; Monaco, M.R.L.; Bentivoglio, A.R.; Coelho-Júnior, H.J.; Landi, F.; Bernabei, R.; Marzetti, E. Mitochondrial dysfunction and aging: Insights from the analysis of extracellular vesicles. Int. J. Mol. Sci. 2019, 20, 805. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Weilner, S.; Keider, V.; Winter, M.; Harreither, E.; Salzer, B.; Weiss, F.; Schraml, E.; Messner, P.; Pietschmann, P.; Hildner, F.; et al. Vesicular Galectin-3 levels decrease with donor age and contribute to the reduced osteo-inductive potential of human plasma derived extracellular vesicles. Aging 2016, 8, 16–33. [Google Scholar] [CrossRef] [Green Version]
- Al-Salam, S.; Hashmi, S. Myocardial ischemia reperfusion injury: Apoptotic, inflammatory and oxidative stress role of galectin-3. Cell. Physiol. Biochem. 2018, 50, 1123–1139. [Google Scholar] [CrossRef]
- Ibarrola, J.F.; Matilla, L.; Martínez-Martínez, E.; Gueret, A.; Fernández-Celis, A.; Henry, J.-P.; Nicol, L.; Jaisser, F.; Mulder, P.; Ouvrard-Pascaud, A.; et al. Myocardial injury after ischemia/reperfusion is attenuated by pharmacological galectin-3 inhibition. Sci. Rep. 2019, 9, 607. [Google Scholar] [CrossRef] [Green Version]



Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Majka, M.; Kleibert, M.; Wojciechowska, M. Impact of the Main Cardiovascular Risk Factors on Plasma Extracellular Vesicles and Their Influence on the Heart’s Vulnerability to Ischemia-Reperfusion Injury. Cells 2021, 10, 3331. https://doi.org/10.3390/cells10123331
Majka M, Kleibert M, Wojciechowska M. Impact of the Main Cardiovascular Risk Factors on Plasma Extracellular Vesicles and Their Influence on the Heart’s Vulnerability to Ischemia-Reperfusion Injury. Cells. 2021; 10(12):3331. https://doi.org/10.3390/cells10123331
Chicago/Turabian StyleMajka, Miłosz, Marcin Kleibert, and Małgorzata Wojciechowska. 2021. "Impact of the Main Cardiovascular Risk Factors on Plasma Extracellular Vesicles and Their Influence on the Heart’s Vulnerability to Ischemia-Reperfusion Injury" Cells 10, no. 12: 3331. https://doi.org/10.3390/cells10123331