
Perspectives in the Cell-Based Therapies of Various Aspects of the Spinal Cord Injury-Associated Pathologies: Lessons from the Animal Models


Abstract
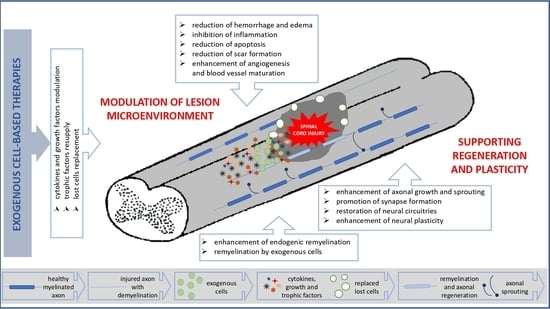
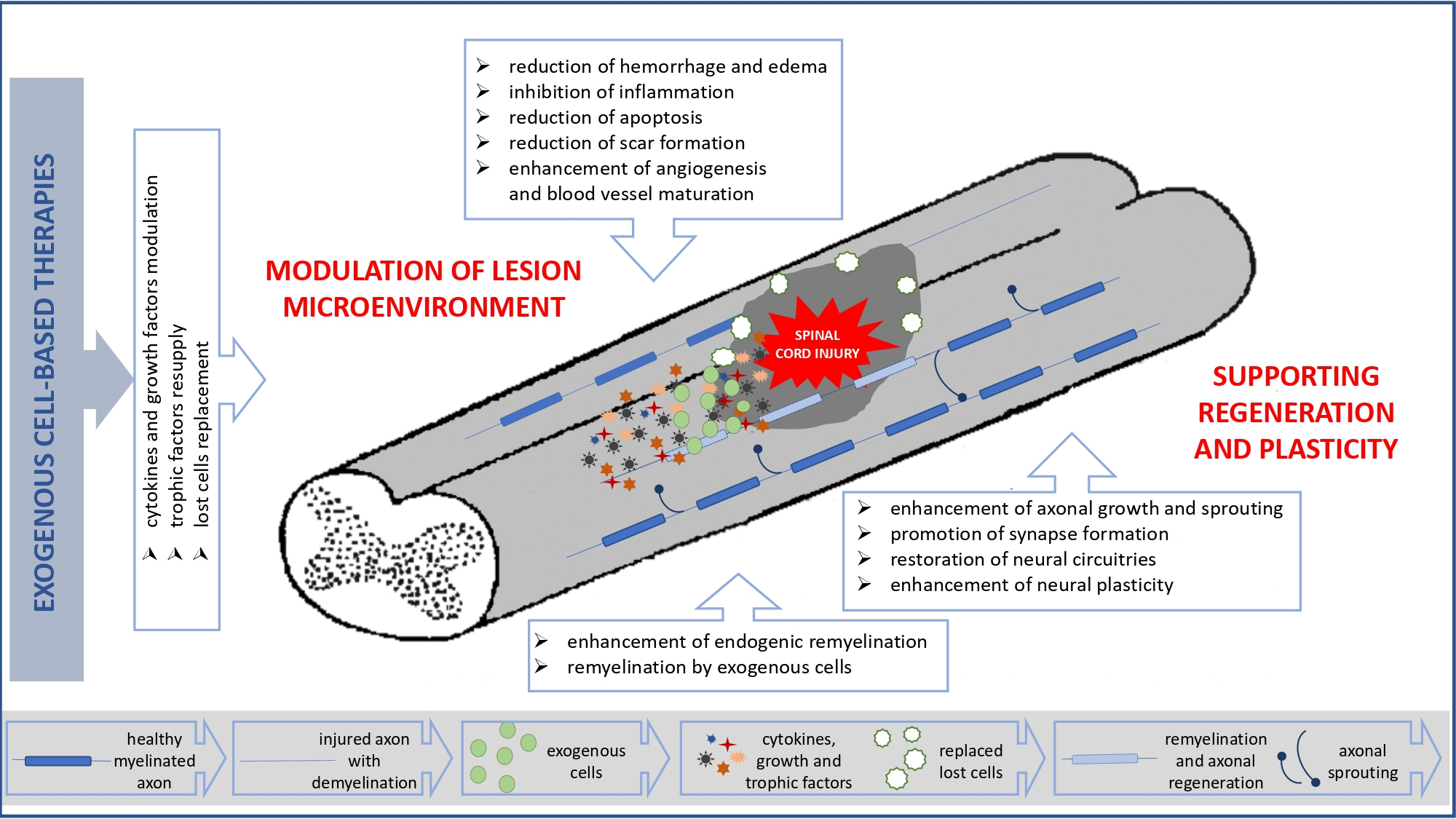
:
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Inhibition of Inflammatory Reaction
3. Keeping a Balance between Neuronal Apoptosis and Autophagy
4. Enhancing Angiogenesis
5. Increasing Axonal Sprouting and Regeneration
6. Supporting Remyelination
7. Rebuilding and/or Activation of Spinal Cord Network
8. Concluding Remarks
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| 5-HT | 5-hydroxytryptamine, serotonin |
| 5-HT2 | 5-HT2 receptors for serotonin |
| 5-HT7 | 5-HT7 receptors for serotonin |
| AMPA | α-amino-3-hydroxy-5-methyl-4-isoxazole propionic acid |
| Bax | pro-apoptotic Bcl-2 protein factor |
| BBB | Basso-Beattie-Bresnahan locomotor rating score |
| Bcl-2 | anti-apoptotic protein inhibiting Beclin 1-dependent autophagy |
| BDNF | brain-derived neurotrophic factor |
| BM-MSCs | bone marrow mesenchymal stem cells |
| BMS | Basso Mouse Scale for locomotor rating |
| CNS | central nervous system |
| CREB | cAMP response element-binding protein div days in vitro |
| dpl | days post lesion |
| DPSCs | dental pulp stem cells |
| EPO | erythropoietin ESC embryonic stem |
| FAK | focal adhesion kinase |
| FasL | ligand for Fas receptor |
| FGF2 | fibroblast growth factor 2 |
| GABA | gamma-aminobutyric acid |
| GDAsBMP | bone morphogenetic protein-induced astrocytes derived from embryonic glial-restricted precursors |
| GDNF | glial cell-derived neurotrophic factor |
| GMCSF | granulocyte-macrophage colony-stimulating factor |
| HGF | hepatocyte growth factor |
| HIF-1α | hypoxia-inducible factor 1-alpha |
| IFN-γ | interferon γ |
| IL-10 | interleukin 10 |
| IL-12 | interleukin 12 |
| IL-1β | interleukin 1β |
| IL-6 | interleukin 6 |
| iNOS | inducible nitric oxide synthase |
| iNSCs | inducible neural stem cells |
| iPSCs | inducible pluripotenet stem cells |
| iPS-NSCs | induced pluripotent stem cell-derived neural stem cells |
| iPSs | induced pluripotent stem cells |
| KA | kainic acid |
| LC3 | autophagy factor |
| M1 | pro-inflammatory type of macrophages |
| M2 | anti-inflammatory type of macrophages |
| MAG | myelin-associated glycoprotein |
| MCP1 | monocyte chemoattractant protein 1 |
| MIP-1α | macrophage inflammatory protein 1-alpha |
| MSCs | mesenchymal stem cells |
| NF-κB | nuclear factor kappa B |
| NGF | nerve growth factor |
| NMDA | N-methyl-D-aspartate glutamate |
| Nogo-A | neurite outgrowth inhibitor A |
| NSCs | neural stem cells |
| NPCs | neural progenitor cells |
| NT-4 | neurotrophin-4 |
| Olig2 | oligodendrocyte transcription factor |
| OECs | olfactory ensheathing cells |
| OPCs | oligodendrocyte precursor cells |
| PNS | peripheral nervous system |
| RhoA | ras homolog family member A |
| ROCK | Rho-associated protein kinase |
| SCI | spinal cord injury |
| SCs | Schwann cells |
| sEV | small extracellular vesicles |
| SKP-SCs | skin-derived precursors differentiated into Schwann cells |
| STAT3 | signal transducer and activator of transcription 3 |
| TH | tyrosine hydroxylase |
| TNF-α | tumor necrosis factor-α |
| TT | total spinal cord transection |
| VEGF | vascular endothelial growth factor |
| Wnt/β | catenin |
References
- Benton, R.L.; Hagg, T. Vascular Pathology as a Potential Therapeutic Target in SCI. Transl. Stroke Res. 2011, 2, 556–574. [Google Scholar] [CrossRef] [PubMed]
- Li, X.Q.; Fang, B.; Tan, W.F.; Wang, Z.L.; Sun, X.J.; Zhang, Z.L.; Ma, H. miR-320a affects spinal cord edema through negatively regulating aquaporin-1 of blood-spinal cord barrier during bimodal stage after ischemia reperfusion injury in rats. BMC Neurosci. 2016, 17, 10. [Google Scholar] [CrossRef] [Green Version]
- Park, E.; Velumian, A.A.; Fehlings, M.G. The role of excitotoxicity in secondary mechanisms of spinal cord injury: A review with an emphasis on the implications for white matter degeneration. J. Neurotrauma 2004, 21, 754–774. [Google Scholar] [CrossRef]
- McAdoo, D.J.; Hughes, M.G.; Nie, L.; Shah, B.; Clifton, C.; Fullwood, S.; Hulsebosch, C.E. The effect of glutamate receptor blockers on glutamate release following spinal cord injury. Lack of evidence for an ongoing feedback cascade of damage→ glutamate release→ damage→ glutamate release→ etc. Brain Res. 2005, 1038, 92–99. [Google Scholar] [CrossRef] [PubMed]
- McDonald, J.W.; Sadowsky, C. Spinal-cord injury. Lancet 2002, 359, 417–425. [Google Scholar] [CrossRef]
- Gensel, J.C.; Zhang, B. Macrophage activation and its role in repair and pathology after spinal cord injury. Brain Res. 2015, 1619, 1–11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Silver, J.; Miller, J.H. Regeneration beyond the glial scar. Nat. Rev. Neurosci. 2004, 5, 146–156. [Google Scholar] [CrossRef]
- Soderblom, C.; Luo, X.; Blumenthal, E.; Bray, E.; Lyapichev, K.; Ramos, J.; Krishnan, V.; Lai-Hsu, C.; Park, K.K.; Tsoulfas, P.; et al. Perivascular fibroblasts form the fibrotic scar after contusive spinal cord injury. J. Neurosci. 2013, 33, 13882–13887. [Google Scholar] [CrossRef] [Green Version]
- Faulkner, J.R.; Herrmann, J.E.; Woo, M.J.; Tansey, K.E.; Doan, N.B.; Sofroniew, M.V. Reactive astrocytes protect tissue and preserve function after spinal cord injury. J. Neurosci. 2004, 24, 2143–2155. [Google Scholar] [CrossRef] [Green Version]
- Bradbury, E.J.; Burnside, E.R. Moving beyond the glial scar for spinal cord repair. Nat. Commun. 2019, 10, 3879. [Google Scholar] [CrossRef]
- Tran, A.P.; Warren, P.M.; Silver, J. New insights into glial scar formation after spinal cord injury. Cell Tissue Res. 2021. [Google Scholar] [CrossRef]
- Dolma, S.; Kumar, H. Neutrophil, Extracellular Matrix Components, and Their Interlinked Action in Promoting Secondary Pathogenesis After Spinal Cord Injury. Mol. Neurobiol. 2021, 58, 4652–4665. [Google Scholar] [CrossRef]
- Pernet, V.; Schwab, M.E. The role of Nogo-A in axonal plasticity, regrowth and repair. Cell Tissue Res. 2012, 349, 97–104. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mu, X.; Azbill, R.D.; Springer, J.E. NBQX treatment improves mitochondrial function and reduces oxidative events after spinal cord injury. J. Neurotrauma 2002, 19, 917–927. [Google Scholar] [CrossRef]
- Wrathall, J.R.; Teng, Y.D.; Choiniere, D. Amelioration of functional deficits from spinal cord trauma with systemically administered NBQX, an antagonist of non-N-methyl-D-aspartate receptors. Exp. Neurol. 1996, 137, 119–126. [Google Scholar] [CrossRef] [PubMed]
- Nagoshi, N.; Nakashima, H.; Fehlings, M.G. Riluzole as a neuroprotective drug for spinal cord injury: From bench to bedside. Molecules 2015, 20, 7775–7789. [Google Scholar] [CrossRef]
- Hermanns, S.; Reiprich, P.; Muller, H.W. A reliable method to reduce collagen scar formation in the lesioned rat spinal cord. J. Neurosci. Methods 2001, 110, 141–146. [Google Scholar] [CrossRef]
- Antri, M.; Barthe, J.Y.; Mouffle, C.; Orsal, D. Long-lasting recovery of locomotor function in chronic spinal rat following chronic combined pharmacological stimulation of serotonergic receptors with 8-OHDPAT and quipazine. Neurosci. Lett. 2005, 384, 162–167. [Google Scholar] [CrossRef]
- Rossignol, S.; Chau, C.; Brustein, E.; Giroux, N.; Bouyer, L.; Barbeau, H.; Reader, T.A. Pharmacological activation and modulation of the central pattern generator for locomotion in the cat. Ann. N. Y. Acad. Sci. 1998, 860, 346–359. [Google Scholar] [CrossRef]
- Slawinska, U.; Miazga, K.; Jordan, L.M. 5-HT(2) and 5-HT(7) receptor agonists facilitate plantar stepping in chronic spinal rats through actions on different populations of spinal neurons. Front. Neural Circuits 2014, 8, 95. [Google Scholar]
- Wang, X.; Budel, S.; Baughman, K.; Gould, G.; Song, K.H.; Strittmatter, S.M. Ibuprofen enhances recovery from spinal cord injury by limiting tissue loss and stimulating axonal growth. J. Neurotrauma 2009, 26, 81–95. [Google Scholar] [CrossRef]
- Shultz, R.B.; Zhong, Y. Minocycline targets multiple secondary injury mechanisms in traumatic spinal cord injury. Neural Regen. Res. 2017, 12, 702–713. [Google Scholar] [PubMed]
- Wang, H.; Liu, N.K.; Zhang, Y.P.; Deng, L.; Lu, Q.B.; Shields, C.B.; Walker, M.J.; Li, J.; Xu, X.M. Treadmill training induced lumbar motoneuron dendritic plasticity and behavior recovery in adult rats after a thoracic contusive spinal cord injury. Exp. Neurol. 2015, 271, 368–378. [Google Scholar] [CrossRef] [PubMed]
- Skup, M.; Ziemlinska, E.; Gajewska-Wozniak, O.; Platek, R.; Maciejewska, A.; Czarkowska-Bauch, J. The impact of training and neurotrophins on functional recovery after complete spinal cord transection: Cellular and molecular mechanisms contributing to motor improvement. Acta Neurobiol. Exp. 2014, 74, 121–141. [Google Scholar]
- Stummer, W.; Weber, K.; Tranmer, B.; Baethmann, A.; Kempski, O. Reduced mortality and brain damage after locomotor activity in gerbil forebrain ischemia. Stroke 1994, 25, 1862–1869. [Google Scholar] [CrossRef] [Green Version]
- Loy, K.; Schmalz, A.; Hoche, T.; Jacobi, A.; Kreutzfeldt, M.; Merkler, D.; Bareyre, F.M. Enhanced Voluntary Exercise Improves Functional Recovery following Spinal Cord Injury by Impacting the Local Neuroglial Injury Response and Supporting the Rewiring of Supraspinal Circuits. J. Neurotrauma 2018, 35, 2904–2915. [Google Scholar] [CrossRef]
- Loy, K.; Bareyre, F.M. Rehabilitation following spinal cord injury: How animal models can help our understanding of exercise-induced neuroplasticity. Neural Regen. Res. 2019, 14, 405–412. [Google Scholar]
- Aravind, N.; Harvey, L.A.; Glinsky, J.V. Physiotherapy interventions for increasing muscle strength in people with spinal cord injuries: A systematic review. Spinal Cord 2019, 57, 449–460. [Google Scholar] [CrossRef]
- Calvert, J.S.; Grahn, P.J.; Zhao, K.D.; Lee, K.H. Emergence of Epidural Electrical Stimulation to Facilitate Sensorimotor Network Functionality After Spinal Cord Injury. Neuromodulation 2019, 22, 244–252. [Google Scholar] [CrossRef] [PubMed]
- Rejc, E.; Angeli, C.A. Spinal Cord Epidural Stimulation for Lower Limb Motor Function Recovery in Individuals with Motor Complete Spinal Cord Injury. Phys. Med. Rehabil. Clin. N. Am. 2019, 30, 337–354. [Google Scholar] [CrossRef] [PubMed]
- Mousavi, M.; Hedayatpour, A.; Mortezaee, K.; Mohamadi, Y.; Abolhassani, F.; Hassanzadeh, G. Schwann cell transplantation exerts neuroprotective roles in rat model of spinal cord injury by combating inflammasome activation and improving motor recovery and remyelination. Metab. Brain Dis. 2019, 34, 1117–1130. [Google Scholar] [CrossRef]
- Raisman, G.; Barnett, S.C.; Ramon-Cueto, A. Repair of central nervous system lesions by transplantation of olfactory ensheathing cells. Handb. Clin. Neurol. 2012, 109, 541–549. [Google Scholar]
- Tabakow, P.; Jarmundowicz, W.; Czapiga, B.; Fortuna, W.; Miedzybrodzki, R.; Czyz, M.; Huber, J.; Szarek, D.; Okurowski, S.; Szewczyk, P.; et al. Transplantation of autologous olfactory ensheathing cells in complete human spinal cord injury. Cell Transplant. 2013, 22, 1591–1612. [Google Scholar] [CrossRef] [PubMed]
- Yamamoto, M.; Raisman, G.; Li, D.; Li, Y. Transplanted olfactory mucosal cells restore paw reaching function without regeneration of severed corticospinal tract fibres across the lesion. Brain Res. 2009, 1303, 26–31. [Google Scholar] [CrossRef] [Green Version]
- Lien, B.V.; Tuszynski, M.H.; Lu, P. Astrocytes migrate from human neural stem cell grafts and functionally integrate into the injured rat spinal cord. Exp. Neurol. 2019, 314, 46–57. [Google Scholar] [CrossRef] [PubMed]
- Zhao, X.M.; He, X.Y.; Liu, J.; Xu, Y.; Xu, F.F.; Tan, Y.X.; Zhang, Z.B.; Wang, T.H. Neural Stem Cell Transplantation Improves Locomotor Function in Spinal Cord Transection Rats Associated with Nerve Regeneration and IGF-1 R Expression. Cell Transplant. 2019, 28, 1197–1211. [Google Scholar] [CrossRef] [Green Version]
- Kim, C.; Kim, H.J.; Lee, H.; Lee, H.; Lee, S.J.; Lee, S.T.; Yang, S.R.; Chung, C.K. Mesenchymal Stem Cell Transplantation Promotes Functional Recovery through MMP2/STAT3 Related Astrogliosis after Spinal Cord Injury. Int. J. Stem Cells 2019, 12, 331–339. [Google Scholar] [CrossRef] [Green Version]
- Ma, Z.; Lu, Y.; Yang, Y.; Wang, J.; Kang, X. Research progress and prospects of tissue engineering scaffolds for spinal cord injury repair and protection. Regen. Med. 2019, 14, 887–898. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.W.; Ha, K.Y.; Molon, J.N.; Kim, Y.H. Bone marrow-derived mesenchymal stem cell transplantation for chronic spinal cord injury in rats: Comparative study between intralesional and intravenous transplantation. Spine 2013, 38, E1065–E1074. [Google Scholar] [CrossRef] [PubMed]
- Cui, L.L.; Kerkela, E.; Bakreen, A.; Nitzsche, F.; Andrzejewska, A.; Nowakowski, A.; Janowski, M.; Walczak, P.; Boltze, J.; Lukomska, B.; et al. The cerebral embolism evoked by intra-arterial delivery of allogeneic bone marrow mesenchymal stem cells in rats is related to cell dose and infusion velocity. Stem Cell Res. Ther. 2015, 6, 11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gyak, K.W.; Jeon, S.; Ha, L.; Kim, S.; Kim, J.Y.; Lee, K.S.; Choi, H.; Kim, D.P. Magnetically Actuated SiCN-Based Ceramic Microrobot for Guided Cell Delivery. Adv. Healthc. Mater. 2019, 8, e1900739. [Google Scholar] [CrossRef] [PubMed]
- Rong, Y.; Liu, W.; Wang, J.; Fan, J.; Luo, Y.; Li, L.; Kong, F.; Chen, J.; Tang, P.; Cai, W. Neural stem cell-derived small extracellular vesicles attenuate apoptosis and neuroinflammation after traumatic spinal cord injury by activating autophagy. Cell Death Dis. 2019, 10, 340. [Google Scholar] [CrossRef] [PubMed]
- Anwar, M.A.; Al Shehabi, T.S.; Eid, A.H. Inflammogenesis of Secondary Spinal Cord Injury. Front. Cell. Neurosci. 2016, 10, 98. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Van Broeckhoven, J.; Sommer, D.; Dooley, D.; Hendrix, S.; Franssen, A. Macrophage phagocytosis after spinal cord injury: When friends become foes. Brain 2021. [Google Scholar] [CrossRef]
- Wolf, S.A.; Boddeke, H.W.; Kettenmann, H. Microglia in Physiology and Disease. Annu. Rev. Physiol. 2017, 79, 619–643. [Google Scholar] [CrossRef] [PubMed]
- Dong, Y.; Benveniste, E.N. Immune function of astrocytes. Glia 2001, 36, 180–190. [Google Scholar] [CrossRef]
- Rothwell, N.J. Annual review prize lecture cytokines—Killers in the brain? J. Physiol. 1999, 514 Pt 1, 3–17. [Google Scholar] [CrossRef]
- David, S.; Lopez-Vales, R.; Wee Yong, V. Harmful and beneficial effects of inflammation after spinal cord injury: Potential therapeutic implications. Handb. Clin. Neurol. 2012, 109, 485–502. [Google Scholar]
- Okada, S. The pathophysiological role of acute inflammation after spinal cord injury. Inflamm. Regen. 2016, 36, 20. [Google Scholar] [CrossRef] [Green Version]
- Ren, Y.; Young, W. Managing inflammation after spinal cord injury through manipulation of macrophage function. Neural Plast. 2013, 2013, 945034. [Google Scholar] [CrossRef] [Green Version]
- Fawcett, J.W.; Asher, R.A. The glial scar and central nervous system repair. Brain Res. Bull. 1999, 49, 377–391. [Google Scholar] [CrossRef]
- Bush, T.G.; Puvanachandra, N.; Horner, C.H.; Polito, A.; Ostenfeld, T.; Svendsen, C.N.; Mucke, L.; Johnson, M.H.; Sofroniew, M.V. Leukocyte infiltration, neuronal degeneration, and neurite outgrowth after ablation of scar-forming, reactive astrocytes in adult transgenic mice. Neuron 1999, 23, 297–308. [Google Scholar] [CrossRef] [Green Version]
- David, S.; Kroner, A. Repertoire of microglial and macrophage responses after spinal cord injury. Nat. Rev. Neurosci. 2011, 12, 388–399. [Google Scholar] [CrossRef] [PubMed]
- Milich, L.M.; Ryan, C.B.; Lee, J.K. The origin, fate, and contribution of macrophages to spinal cord injury pathology. Acta Neuropathol. 2019, 137, 785–797. [Google Scholar] [CrossRef]
- Kong, X.; Gao, J. Macrophage polarization: A key event in the secondary phase of acute spinal cord injury. J. Cell. Mol. Med. 2017, 21, 941–954. [Google Scholar] [CrossRef] [PubMed]
- Nakajima, H.; Uchida, K.; Guerrero, A.R.; Watanabe, S.; Sugita, D.; Takeura, N.; Yoshida, A.; Long, G.; Wright, K.T.; Johnson, W.E.; et al. Transplantation of mesenchymal stem cells promotes an alternative pathway of macrophage activation and functional recovery after spinal cord injury. J. Neurotrauma 2012, 29, 1614–1625. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sun, G.; Li, G.; Li, D.; Huang, W.; Zhang, R.; Zhang, H.; Duan, Y.; Wang, B. hucMSC derived exosomes promote functional recovery in spinal cord injury mice via attenuating inflammation. Mater. Sci. Eng. C Mater. Biol. Appl. 2018, 89, 194–204. [Google Scholar] [CrossRef]
- Cheng, Z.; Zhu, W.; Cao, K.; Wu, F.; Li, J.; Wang, G.; Li, H.; Lu, M.; Ren, Y.; He, X. Anti-Inflammatory Mechanism of Neural Stem Cell Transplantation in Spinal Cord Injury. Int. J. Mol. Sci. 2016, 17, 1380. [Google Scholar] [CrossRef] [Green Version]
- Riemann, L.; Younsi, A.; Scherer, M.; Zheng, G.; Skutella, T.; Unterberg, A.W.; Zweckberger, K. Transplantation of Neural Precursor Cells Attenuates Chronic Immune Environment in Cervical Spinal Cord Injury. Front. Neurol. 2018, 9, 428. [Google Scholar] [CrossRef] [Green Version]
- Cusimano, M.; Biziato, D.; Brambilla, E.; Donega, M.; Alfaro-Cervello, C.; Snider, S.; Salani, G.; Pucci, F.; Comi, G.; Garcia-Verdugo, J.M.; et al. Transplanted neural stem/precursor cells instruct phagocytes and reduce secondary tissue damage in the injured spinal cord. Brain 2012, 135, 447–460. [Google Scholar] [CrossRef] [Green Version]
- Wang, T.; Fang, X.; Yin, Z.S. Endothelial progenitor cell-conditioned medium promotes angiogenesis and is neuroprotective after spinal cord injury. Neural Regen. Res. 2018, 13, 887–895. [Google Scholar]
- Cheng, Z.; Bosco, D.B.; Sun, L.; Chen, X.; Xu, Y.; Tai, W.; Didier, R.; Li, J.; Fan, J.; He, X.; et al. Neural Stem Cell-Conditioned Medium Suppresses Inflammation and Promotes Spinal Cord Injury Recovery. Cell Transplant. 2017, 26, 469–482. [Google Scholar] [CrossRef]
- Karova, K.; Wainwright, J.V.; Machova-Urdzikova, L.; Pisal, R.V.; Schmidt, M.; Jendelova, P.; Jhanwar-Uniyal, M. Transplantation of neural precursors generated from spinal progenitor cells reduces inflammation in spinal cord injury via NF-kappaB pathway inhibition. J. Neuroinflamm. 2019, 16, 12. [Google Scholar] [CrossRef] [Green Version]
- Kolar, M.K.; Kingham, P.J.; Novikova, L.N.; Wiberg, M.; Novikov, L.N. The therapeutic effects of human adipose-derived stem cells in a rat cervical spinal cord injury model. Stem Cells Dev. 2014, 23, 1659–1674. [Google Scholar] [CrossRef]
- Pajer, K.; Bellak, T.; Nogradi, A. The mutual interaction between the host spinal cord and grafted undifferentiated stem cells fosters the production of a lesion-induced secretome. Neural Regen. Res. 2020, 15, 1844–1845. [Google Scholar] [PubMed]
- Hakim, R.; Covacu, R.; Zachariadis, V.; Frostell, A.; Sankavaram, S.R.; Brundin, L.; Svensson, M. Mesenchymal stem cells transplanted into spinal cord injury adopt immune cell-like characteristics. Stem Cell Res. Ther. 2019, 10, 115. [Google Scholar] [CrossRef]
- Pajer, K.; Nemes, C.; Berzsenyi, S.; Kovacs, K.A.; Pirity, M.K.; Pajenda, G.; Nogradi, A.; Dinnyes, A. Grafted murine induced pluripotent stem cells prevent death of injured rat motoneurons otherwise destined to die. Exp. Neurol. 2015, 269, 188–201. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pajer, K.; Feichtinger, G.A.; Marton, G.; Sabitzer, S.; Klein, D.; Redl, H.; Nogradi, A. Cytokine signaling by grafted neuroectodermal stem cells rescues motoneurons destined to die. Exp. Neurol. 2014, 261, 180–189. [Google Scholar] [CrossRef] [Green Version]
- Pajer, K.; Bellak, T.; Redl, H.; Nogradi, A. Neuroectodermal Stem Cells Grafted into the Injured Spinal Cord Induce Both Axonal Regeneration and Morphological Restoration via Multiple Mechanisms. J. Neurotrauma 2019, 36, 2977–2990. [Google Scholar] [CrossRef]
- Liu, X.Z.; Xu, X.M.; Hu, R.; Du, C.; Zhang, S.X.; McDonald, J.W.; Dong, H.X.; Wu, Y.J.; Fan, G.S.; Jacquin, M.F.; et al. Neuronal and glial apoptosis after traumatic spinal cord injury. J. Neurosci. 1997, 17, 5395–5406. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jia, Z.; Zhu, H.; Li, J.; Wang, X.; Misra, H.; Li, Y. Oxidative stress in spinal cord injury and antioxidant-based intervention. Spinal Cord 2012, 50, 264–274. [Google Scholar] [CrossRef] [Green Version]
- Los, M.; Wesselborg, S.; Schulze-Osthoff, K. The role of caspases in development, immunity, and apoptotic signal transduction: Lessons from knockout mice. Immunity 1999, 10, 629–639. [Google Scholar] [CrossRef] [Green Version]
- Springer, J.E.; Azbill, R.D.; Knapp, P.E. Activation of the caspase-3 apoptotic cascade in traumatic spinal cord injury. Nat. Med. 1999, 5, 943–946. [Google Scholar] [CrossRef]
- Zhou, H.L.; Zhang, X.J.; Zhang, M.Y.; Yan, Z.J.; Xu, Z.M.; Xu, R.X. Transplantation of Human Amniotic Mesenchymal Stem Cells Promotes Functional Recovery in a Rat Model of Traumatic Spinal Cord Injury. Neurochem. Res. 2016, 41, 2708–2718. [Google Scholar] [CrossRef] [PubMed]
- Yin, F.; Guo, L.; Meng, C.Y.; Liu, Y.J.; Lu, R.F.; Li, P.; Zhou, Y.B. Transplantation of mesenchymal stem cells exerts anti-apoptotic effects in adult rats after spinal cord ischemia-reperfusion injury. Brain Res. 2014, 1561, 1–10. [Google Scholar] [CrossRef]
- Liu, W.; Ding, Y.; Zhang, X.; Wang, L. Bone marrow stromal cells inhibit caspase-12 expression in rats with spinal cord injury. Exp. Ther. Med. 2013, 6, 671–674. [Google Scholar] [CrossRef] [PubMed]
- Wu, S.; Cui, G.; Shao, H.; Du, Z.; Ng, J.C.; Peng, C. The Cotransplantation of Olfactory Ensheathing Cells with Bone Marrow Mesenchymal Stem Cells Exerts Antiapoptotic Effects in Adult Rats after Spinal Cord Injury. Stem Cells Int. 2015, 2015, 516215. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nicola, F.D.C.; Marques, M.R.; Odorcyk, F.; Arcego, D.M.; Petenuzzo, L.; Aristimunha, D.; Vizuete, A.; Sanches, E.F.; Pereira, D.P.; Maurmann, N.; et al. Neuroprotector effect of stem cells from human exfoliated deciduous teeth transplanted after traumatic spinal cord injury involves inhibition of early neuronal apoptosis. Brain Res. 2017, 1663, 95–105. [Google Scholar] [CrossRef]
- Gao, S.; Ding, J.; Xiao, H.J.; Li, Z.Q.; Chen, Y.; Zhou, X.S.; Wang, J.E.; Wu, J.; Shi, W.Z. Anti-inflammatory and anti-apoptotic effect of combined treatment with methylprednisolone and amniotic membrane mesenchymal stem cells after spinal cord injury in rats. Neurochem. Res. 2014, 39, 1544–1552. [Google Scholar] [CrossRef]
- Lu, Y.; Zhou, Y.; Zhang, R.; Wen, L.; Wu, K.; Li, Y.; Yao, Y.; Duan, R.; Jia, Y. Bone Mesenchymal Stem Cell-Derived Extracellular Vesicles Promote Recovery Following Spinal Cord Injury via Improvement of the Integrity of the Blood-Spinal Cord Barrier. Front. Neurosci. 2019, 13, 209. [Google Scholar] [CrossRef] [Green Version]
- Huang, J.H.; Yin, X.M.; Xu, Y.; Xu, C.C.; Lin, X.; Ye, F.B.; Cao, Y.; Lin, F.Y. Systemic Administration of Exosomes Released from Mesenchymal Stromal Cells Attenuates Apoptosis, Inflammation, and Promotes Angiogenesis after Spinal Cord Injury in Rats. J. Neurotrauma 2017, 34, 3388–3396. [Google Scholar] [CrossRef] [PubMed]
- Zhao, S.; Liu, Y.; Pu, Z. Bone marrow mesenchymal stem cell-derived exosomes attenuate D-GaIN/LPS-induced hepatocyte apoptosis by activating autophagy in vitro. Drug Des. Devel. Ther. 2019, 13, 2887–2897. [Google Scholar] [CrossRef] [PubMed]
- Maiuri, M.C.; Zalckvar, E.; Kimchi, A.; Kroemer, G. Self-eating and self-killing: Crosstalk between autophagy and apoptosis. Nat. Rev. Mol. Cell Biol. 2007, 8, 741–752. [Google Scholar] [CrossRef]
- Hou, H.; Zhang, L.; Zhang, L.; Tang, P. Acute spinal cord injury in rats should target activated autophagy. J. Neurosurg. Spine 2014, 20, 568–577. [Google Scholar] [CrossRef]
- Liu, S.; Sarkar, C.; Dinizo, M.; Faden, A.I.; Koh, E.Y.; Lipinski, M.M.; Wu, J. Disrupted autophagy after spinal cord injury is associated with ER stress and neuronal cell death. Cell Death Dis. 2015, 6, e1582. [Google Scholar] [CrossRef] [PubMed]
- He, M.; Ding, Y.; Chu, C.; Tang, J.; Xiao, Q.; Luo, Z.G. Autophagy induction stabilizes microtubules and promotes axon regeneration after spinal cord injury. Proc. Natl. Acad. Sci. USA 2016, 113, 11324–11329. [Google Scholar] [CrossRef] [Green Version]
- Maiti, P.; Peruzzaro, S.; Kolli, N.; Andrews, M.; Al-Gharaibeh, A.; Rossignol, J.; Dunbar, G.L. Transplantation of mesenchymal stem cells overexpressing interleukin-10 induces autophagy response and promotes neuroprotection in a rat model of TBI. J. Cell. Mol. Med. 2019, 23, 5211–5224. [Google Scholar] [CrossRef] [Green Version]
- Li, H.; Wang, C.; He, T.; Zhao, T.; Chen, Y.Y.; Shen, Y.L.; Zhang, X.; Wang, L.L. Mitochondrial Transfer from Bone Marrow Mesenchymal Stem Cells to Motor Neurons in Spinal Cord Injury Rats via Gap Junction. Theranostics 2019, 9, 2017–2035. [Google Scholar] [CrossRef] [PubMed]
- Nieto-Diaz, M.; Esteban, F.J.; Reigada, D.; Munoz-Galdeano, T.; Yunta, M.; Caballero-Lopez, M.; Navarro-Ruiz, R.; Del Aguila, A.; Maza, R.M. MicroRNA dysregulation in spinal cord injury: Causes, consequences and therapeutics. Front. Cell Neurosci. 2014, 8, 53. [Google Scholar] [CrossRef]
- Ning, B.; Gao, L.; Liu, R.H.; Liu, Y.; Zhang, N.S.; Chen, Z.Y. microRNAs in spinal cord injury: Potential roles and therapeutic implications. Int. J. Biol. Sci. 2014, 10, 997–1006. [Google Scholar] [CrossRef] [Green Version]
- Zhou, X.; Chu, X.; Yuan, H.; Qiu, J.; Zhao, C.; Xin, D.; Li, T.; Ma, W.; Wang, H.; Wang, Z.; et al. Mesenchymal stem cell derived EVs mediate neuroprotection after spinal cord injury in rats via the microRNA-21-5p/FasL gene axis. Biomed. Pharmacother. 2019, 115, 108818. [Google Scholar] [CrossRef] [PubMed]
- Song, J.L.; Zheng, W.; Chen, W.; Qian, Y.; Ouyang, Y.M.; Fan, C.Y. Lentivirus-mediated microRNA-124 gene-modified bone marrow mesenchymal stem cell transplantation promotes the repair of spinal cord injury in rats. Exp. Mol. Med. 2017, 49, e332. [Google Scholar] [CrossRef] [PubMed]
- Xu, H.; Cao, J.; Xu, J.; Li, H.; Shen, H.; Li, X.; Wang, Z.; Wu, J.; Chen, G. GATA-4 regulates neuronal apoptosis after intracerebral hemorrhage via the NF-kappaB/Bax/Caspase-3 pathway both in vivo and in vitro. Exp. Neurol. 2019, 315, 21–31. [Google Scholar] [CrossRef] [PubMed]
- Yuan, H.X.; Feng, X.E.; Liu, E.L.; Ge, R.; Zhang, Y.L.; Xiao, B.G.; Li, Q.S. 5,2′-dibromo-2,4′,5′-trihydroxydiphenylmethanone attenuates LPS-induced inflammation and ROS production in EA.hy926 cells via HMBOX1 induction. J. Cell. Mol. Med. 2019, 23, 453–463. [Google Scholar] [CrossRef]
- Figley, S.A.; Khosravi, R.; Legasto, J.M.; Tseng, Y.F.; Fehlings, M.G. Characterization of vascular disruption and blood-spinal cord barrier permeability following traumatic spinal cord injury. J. Neurotrauma 2014, 31, 541–552. [Google Scholar] [CrossRef] [Green Version]
- Graumann, U.; Ritz, M.F.; Hausmann, O. Necessity for re-vascularization after spinal cord injury and the search for potential therapeutic options. Curr. Neurovasc. Res. 2011, 8, 334–341. [Google Scholar] [CrossRef]
- Casella, G.T.; Marcillo, A.; Bunge, M.B.; Wood, P.M. New vascular tissue rapidly replaces neural parenchyma and vessels destroyed by a contusion injury to the rat spinal cord. Exp. Neurol. 2002, 173, 63–76. [Google Scholar] [CrossRef]
- Imperato-Kalmar, E.L.; McKinney, R.A.; Schnell, L.; Rubin, B.P.; Schwab, M.E. Local changes in vascular architecture following partial spinal cord lesion in the rat. Exp. Neurol. 1997, 145, 322–328. [Google Scholar] [CrossRef]
- Benton, R.L.; Whittemore, S.R. VEGF165 therapy exacerbates secondary damage following spinal cord injury. Neurochem. Res. 2003, 28, 1693–1703. [Google Scholar] [CrossRef]
- Lutton, C.; Young, Y.W.; Williams, R.; Meedeniya, A.C.; Mackay-Sim, A.; Goss, B. Combined VEGF and PDGF treatment reduces secondary degeneration after spinal cord injury. J. Neurotrauma 2012, 29, 957–970. [Google Scholar] [CrossRef] [Green Version]
- Gomes, E.D.; Mendes, S.S.; Assuncao-Silva, R.C.; Teixeira, F.G.; Pires, A.O.; Anjo, S.I.; Manadas, B.; Leite-Almeida, H.; Gimble, J.M.; Sousa, N.; et al. Co-Transplantation of Adipose Tissue-Derived Stromal Cells and Olfactory Ensheathing Cells for Spinal Cord Injury Repair. Stem Cells 2018, 36, 696–708. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Reshamwala, R.; Shah, M.; St John, J.; Ekberg, J. Survival and Integration of Transplanted Olfactory Ensheathing Cells are Crucial for Spinal Cord Injury Repair: Insights from the Last 10 Years of Animal Model Studies. Cell Transplant. 2019, 28, 132S–159S. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.; Guo, G.H.; Wang, G.S.; Guan, C.X.; Yue, L. Influence of neural stem cell transplantation on angiogenesis in rats with spinal cord injury. Genet. Mol. Res. 2014, 13, 6083–6092. [Google Scholar] [CrossRef]
- Zeng, Y.; Han, H.; Tang, B.; Chen, J.; Mao, D.; Xiong, M. Transplantation of Recombinant Vascular Endothelial Growth Factor (VEGF)189-Neural Stem Cells Downregulates Transient Receptor Potential Vanilloid 1 (TRPV1) and Improves Motor Outcome in Spinal Cord Injury. Med. Sci. Monit. 2018, 24, 1089–1096. [Google Scholar] [CrossRef] [PubMed]
- Nori, S.; Okada, Y.; Yasuda, A.; Tsuji, O.; Takahashi, Y.; Kobayashi, Y.; Fujiyoshi, K.; Koike, M.; Uchiyama, Y.; Ikeda, E.; et al. Grafted human-induced pluripotent stem-cell-derived neurospheres promote motor functional recovery after spinal cord injury in mice. Proc. Natl. Acad. Sci. USA 2011, 108, 16825–16830. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, Y.T.; Tsai, M.J.; Hsieh, N.; Lo, M.J.; Lee, M.J.; Cheng, H.; Huang, W.C. The superiority of conditioned medium derived from rapidly expanded mesenchymal stem cells for neural repair. Stem Cell Res. Ther. 2019, 10, 390. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, X.; Wang, J.; Li, G.; Lv, H. Effect of combined chondroitinase ABC and hyperbaric oxygen therapy in a rat model of spinal cord injury. Mol. Med. Rep. 2018, 18, 25–30. [Google Scholar] [CrossRef] [PubMed]
- Rocha, L.A.; Sousa, R.A.; Learmonth, D.A.; Salgado, A.J. The Role of Biomaterials as Angiogenic Modulators of Spinal Cord Injury: Mimetics of the Spinal Cord, Cell and Angiogenic Factor Delivery Agents. Front. Pharmacol. 2018, 9, 164. [Google Scholar] [CrossRef]
- Yu, S.; Yao, S.; Wen, Y.; Wang, Y.; Wang, H.; Xu, Q. Angiogenic microspheres promote neural regeneration and motor function recovery after spinal cord injury in rats. Sci. Rep. 2016, 6, 33428. [Google Scholar] [CrossRef] [Green Version]
- Wang, L.; Shi, Q.; Dai, J.; Gu, Y.; Feng, Y.; Chen, L. Increased vascularization promotes functional recovery in the transected spinal cord rats by implanted vascular endothelial growth factor-targeting collagen scaffold. J. Orthop. Res. 2018, 36, 1024–1034. [Google Scholar] [CrossRef] [Green Version]
- Ropper, A.E.; Thakor, D.K.; Han, I.; Yu, D.; Zeng, X.; Anderson, J.E.; Aljuboori, Z.; Kim, S.W.; Wang, H.; Sidman, R.L.; et al. Defining recovery neurobiology of injured spinal cord by synthetic matrix-assisted hMSC implantation. Proc. Natl. Acad. Sci. USA 2017, 114, E820–E829. [Google Scholar] [CrossRef] [Green Version]
- Kitamura, K.; Iwanami, A.; Nakamura, M.; Yamane, J.; Watanabe, K.; Suzuki, Y.; Miyazawa, D.; Shibata, S.; Funakoshi, H.; Miyatake, S.; et al. Hepatocyte growth factor promotes endogenous repair and functional recovery after spinal cord injury. J. Neurosci. Res. 2007, 85, 2332–2342. [Google Scholar] [CrossRef] [PubMed]
- Kitamura, M.; Hirano, S.; Kanemaru, S.I.; Kitani, Y.; Ohno, S.; Kojima, T.; Nakamura, T.; Ito, J.; Rosen, C.A.; Gilbert, T.W. Glottic regeneration with a tissue-engineering technique, using acellular extracellular matrix scaffold in a canine model. J. Tissue Eng. Regen. Med. 2016, 10, 825–832. [Google Scholar] [CrossRef] [Green Version]
- Yamane, K.; Mazaki, T.; Shiozaki, Y.; Yoshida, A.; Shinohara, K.; Nakamura, M.; Yoshida, Y.; Zhou, D.; Kitajima, T.; Tanaka, M.; et al. Collagen-Binding Hepatocyte Growth Factor (HGF) alone or with a Gelatin- furfurylamine Hydrogel Enhances Functional Recovery in Mice after Spinal Cord Injury. Sci. Rep. 2018, 8, 917. [Google Scholar] [CrossRef] [Green Version]
- Takano, M.; Kawabata, S.; Shibata, S.; Yasuda, A.; Nori, S.; Tsuji, O.; Nagoshi, N.; Iwanami, A.; Ebise, H.; Horiuchi, K.; et al. Enhanced Functional Recovery from Spinal Cord Injury in Aged Mice after Stem Cell Transplantation through HGF Induction. Stem Cell Rep. 2017, 8, 509–518. [Google Scholar] [CrossRef] [Green Version]
- Cheng, C.H.; Lin, C.T.; Lee, M.J.; Tsai, M.J.; Huang, W.H.; Huang, M.C.; Lin, Y.L.; Chen, C.J.; Huang, W.C.; Cheng, H. Local Delivery of High-Dose Chondroitinase ABC in the Sub-Acute Stage Promotes Axonal Outgrowth and Functional Recovery after Complete Spinal Cord Transection. PLoS ONE 2015, 10, e0138705. [Google Scholar] [CrossRef] [Green Version]
- Barritt, A.W.; Davies, M.; Marchand, F.; Hartley, R.; Grist, J.; Yip, P.; McMahon, S.B.; Bradbury, E.J. Chondroitinase ABC promotes sprouting of intact and injured spinal systems after spinal cord injury. J. Neurosci. 2006, 26, 10856–10867. [Google Scholar] [CrossRef]
- Rosenzweig, E.S.; Salegio, E.A.; Liang, J.J.; Weber, J.L.; Weinholtz, C.A.; Brock, J.H.; Moseanko, R.; Hawbecker, S.; Pender, R.; Cruzen, C.L.; et al. Chondroitinase improves anatomical and functional outcomes after primate spinal cord injury. Nat. Neurosci. 2019, 22, 1269–1275. [Google Scholar] [CrossRef] [PubMed]
- Alilain, W.J.; Horn, K.P.; Hu, H.; Dick, T.E.; Silver, J. Functional regeneration of respiratory pathways after spinal cord injury. Nature 2011, 475, 196–200. [Google Scholar] [CrossRef] [Green Version]
- Janzadeh, A.; Sarveazad, A.; Yousefifard, M.; Dameni, S.; Samani, F.S.; Mokhtarian, K.; Nasirinezhad, F. Combine effect of Chondroitinase ABC and low level laser (660 nm) on spinal cord injury model in adult male rats. Neuropeptides 2017, 65, 90–99. [Google Scholar] [CrossRef] [PubMed]
- Fuhrmann, T.; Anandakumaran, P.N.; Payne, S.L.; Pakulska, M.M.; Varga, B.V.; Nagy, A.; Tator, C.; Shoichet, M.S. Combined delivery of chondroitinase ABC and human induced pluripotent stem cell-derived neuroepithelial cells promote tissue repair in an animal model of spinal cord injury. Biomed. Mater. 2018, 13, 024103. [Google Scholar] [CrossRef] [PubMed]
- Suzuki, H.; Ahuja, C.S.; Salewski, R.P.; Li, L.; Satkunendrarajah, K.; Nagoshi, N.; Shibata, S.; Fehlings, M.G. Neural stem cell mediated recovery is enhanced by Chondroitinase ABC pretreatment in chronic cervical spinal cord injury. PLoS ONE 2017, 12, e0182339. [Google Scholar]
- Shinozaki, M.; Iwanami, A.; Fujiyoshi, K.; Tashiro, S.; Kitamura, K.; Shibata, S.; Fujita, H.; Nakamura, M.; Okano, H. Combined treatment with chondroitinase ABC and treadmill rehabilitation for chronic severe spinal cord injury in adult rats. Neurosci. Res. 2016, 113, 37–47. [Google Scholar] [CrossRef] [PubMed]
- Tester, N.J.; Plaas, A.H.; Howland, D.R. Effect of body temperature on chondroitinase ABC’s ability to cleave chondroitin sulfate glycosaminoglycans. J. Neurosci. Res. 2007, 85, 1110–1118. [Google Scholar] [CrossRef]
- Carwardine, D.; Prager, J.; Neeves, J.; Muir, E.M.; Uney, J.; Granger, N.; Wong, L.F. Transplantation of canine olfactory ensheathing cells producing chondroitinase ABC promotes chondroitin sulphate proteoglycan digestion and axonal sprouting following spinal cord injury. PLoS ONE 2017, 12, e0188967. [Google Scholar] [CrossRef] [Green Version]
- Kanno, H.; Pressman, Y.; Moody, A.; Berg, R.; Muir, E.M.; Rogers, J.H.; Ozawa, H.; Itoi, E.; Pearse, D.D.; Bunge, M.B. Combination of engineered Schwann cell grafts to secrete neurotrophin and chondroitinase promotes axonal regeneration and locomotion after spinal cord injury. J. Neurosci. 2014, 34, 1838–1855. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zukor, K.; Belin, S.; Wang, C.; Keelan, N.; Wang, X.; He, Z. Short hairpin RNA against PTEN enhances regenerative growth of corticospinal tract axons after spinal cord injury. J. Neurosci. 2013, 33, 15350–15361. [Google Scholar] [CrossRef]
- Jin, Y.; Shumsky, J.S.; Fischer, I. Axonal regeneration of different tracts following transplants of human glial restricted progenitors into the injured spinal cord in rats. Brain Res. 2018, 1686, 101–112. [Google Scholar] [CrossRef]
- Goulao, M.; Ghosh, B.; Urban, M.W.; Sahu, M.; Mercogliano, C.; Charsar, B.A.; Komaravolu, S.; Block, C.G.; Smith, G.M.; Wright, M.C.; et al. Astrocyte progenitor transplantation promotes regeneration of bulbospinal respiratory axons, recovery of diaphragm function, and a reduced macrophage response following cervical spinal cord injury. Glia 2019, 67, 452–466. [Google Scholar] [CrossRef]
- Shih, C.H.; Lacagnina, M.; Leuer-Bisciotti, K.; Proschel, C. Astroglial-derived periostin promotes axonal regeneration after spinal cord injury. J. Neurosci. 2014, 34, 2438–2443. [Google Scholar] [CrossRef] [Green Version]
- Nagoshi, N.; Khazaei, M.; Ahlfors, J.E.; Ahuja, C.S.; Nori, S.; Wang, J.; Shibata, S.; Fehlings, M.G. Human Spinal Oligodendrogenic Neural Progenitor Cells Promote Functional Recovery After Spinal Cord Injury by Axonal Remyelination and Tissue Sparing. Stem Cells Transl. Med. 2018, 7, 806–818. [Google Scholar] [CrossRef] [Green Version]
- Sanchis-Gomar, F.; Perez-Quilis, C.; Lippi, G. Erythropoietin receptor (EpoR) agonism is used to treat a wide range of disease. Mol. Med. 2013, 19, 62–64. [Google Scholar] [CrossRef]
- Subiros, N.; Del Barco, D.G.; Coro-Antich, R.M. Erythropoietin: Still on the neuroprotection road. Ther. Adv. Neurol. Disord. 2012, 5, 161–173. [Google Scholar] [CrossRef] [Green Version]
- Ozkunt, O.; Sariyilmaz, K.; Gemalmaz, H.C.; Gurgen, S.G.; Yener, U.; Dikici, F. Investigation of efficacy of treatment in spinal cord injury: Erythropoietin versus methylprednisolone. J. Orthop. Surg. 2017, 25, 2309499017739481. [Google Scholar] [CrossRef] [Green Version]
- Tanaka, T.; Murata, H.; Miyazaki, R.; Yoshizumi, T.; Sato, M.; Ohtake, M.; Tateishi, K.; Kim, P.; Yamamoto, T. Human recombinant erythropoietin improves motor function in rats with spinal cord compression-induced cervical myelopathy. PLoS ONE 2019, 14, e0214351. [Google Scholar] [CrossRef] [PubMed]
- Carelli, S.; Giallongo, T.; Gombalova, Z.; Merli, D.; Di Giulio, A.M.; Gorio, A. EPO-releasing neural precursor cells promote axonal regeneration and recovery of function in spinal cord traumatic injury. Restor. Neurol. Neurosci. 2017, 35, 583–599. [Google Scholar] [CrossRef] [Green Version]
- Potdar, P.D.; Jethmalani, Y.D. Human dental pulp stem cells: Applications in future regenerative medicine. World J. Stem Cells 2015, 7, 839–851. [Google Scholar] [CrossRef] [PubMed]
- Kabatas, S.; Demir, C.S.; Civelek, E.; Yilmaz, I.; Kircelli, A.; Yilmaz, C.; Akyuva, Y.; Karaoz, E. Neuronal regeneration in injured rat spinal cord after human dental pulp derived neural crest stem cell transplantation. Bratisl. Lek. Listy 2018, 119, 143–151. [Google Scholar] [CrossRef] [Green Version]
- Nagashima, K.; Miwa, T.; Soumiya, H.; Ushiro, D.; Takeda-Kawaguchi, T.; Tamaoki, N.; Ishiguro, S.; Sato, Y.; Miyamoto, K.; Ohno, T.; et al. Priming with FGF2 stimulates human dental pulp cells to promote axonal regeneration and locomotor function recovery after spinal cord injury. Sci. Rep. 2017, 7, 13500. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, D.; Zhang, P.; Yao, X.; Li, H.; Shen, H.; Li, X.; Wu, J.; Lu, X. Exosomes Derived From miR-133b-Modified Mesenchymal Stem Cells Promote Recovery After Spinal Cord Injury. Front. Neurosci. 2018, 12, 845. [Google Scholar] [CrossRef] [Green Version]
- Nave, K.A.; Werner, H.B. Myelination of the nervous system: Mechanisms and functions. Annu. Rev. Cell Dev. Biol. 2014, 30, 503–533. [Google Scholar] [CrossRef]
- Almad, A.; Sahinkaya, F.R.; McTigue, D.M. Oligodendrocyte fate after spinal cord injury. Neurotherapeutics 2011, 8, 262–273. [Google Scholar] [CrossRef] [Green Version]
- Plemel, J.R.; Keough, M.B.; Duncan, G.J.; Sparling, J.S.; Yong, V.W.; Stys, P.K.; Tetzlaff, W. Remyelination after spinal cord injury: Is it a target for repair? Prog. Neurobiol. 2014, 117, 54–72. [Google Scholar] [CrossRef]
- Franklin, R.J.; Ffrench-Constant, C. Remyelination in the CNS: From biology to therapy. Nat. Rev. Neurosci. 2008, 9, 839–855. [Google Scholar] [CrossRef]
- Zawadzka, M.; Rivers, L.E.; Fancy, S.P.; Zhao, C.; Tripathi, R.; Jamen, F.; Young, K.; Goncharevich, A.; Pohl, H.; Rizzi, M.; et al. CNS-resident glial progenitor/stem cells produce Schwann cells as well as oligodendrocytes during repair of CNS demyelination. Cell Stem Cell 2010, 6, 578–590. [Google Scholar] [CrossRef] [Green Version]
- Duncan, G.J.; Manesh, S.B.; Hilton, B.J.; Assinck, P.; Plemel, J.R.; Tetzlaff, W. The fate and function of oligodendrocyte progenitor cells after traumatic spinal cord injury. Glia 2020, 68, 227–245. [Google Scholar] [CrossRef]
- Assinck, P.; Duncan, G.J.; Hilton, B.J.; Plemel, J.R.; Tetzlaff, W. Cell transplantation therapy for spinal cord injury. Nat. Neurosci. 2017, 20, 637–647. [Google Scholar] [CrossRef] [PubMed]
- Ulanska-Poutanen, J.; Mieczkowski, J.; Zhao, C.; Konarzewska, K.; Kaza, B.; Pohl, H.B.; Bugajski, L.; Kaminska, B.; Franklin, R.J.; Zawadzka, M. Injury-induced perivascular niche supports alternative differentiation of adult rodent CNS progenitor cells. Elife 2018, 7, e30325. [Google Scholar] [CrossRef]
- Woodruff, R.H.; Franklin, R.J. Demyelination and remyelination of the caudal cerebellar peduncle of adult rats following stereotaxic injections of lysolecithin, ethidium bromide, and complement/anti-galactocerebroside: A comparative study. Glia 1999, 25, 216–228. [Google Scholar] [CrossRef]
- Assinck, P.; Duncan, G.J.; Plemel, J.R.; Lee, M.J.; Stratton, J.A.; Manesh, S.B.; Liu, J.; Ramer, L.M.; Kang, S.H.; Bergles, D.E.; et al. Myelinogenic Plasticity of Oligodendrocyte Precursor Cells following Spinal Cord Contusion Injury. J. Neurosci. 2017, 37, 8635–8654. [Google Scholar] [CrossRef] [PubMed]
- Hesp, Z.C.; Goldstein, E.Z.; Miranda, C.J.; Kaspar, B.K.; McTigue, D.M. Chronic oligodendrogenesis and remyelination after spinal cord injury in mice and rats. J. Neurosci. 2015, 35, 1274–1290. [Google Scholar] [CrossRef] [PubMed]
- Powers, B.E.; Lasiene, J.; Plemel, J.R.; Shupe, L.; Perlmutter, S.I.; Tetzlaff, W.; Horner, P.J. Axonal thinning and extensive remyelination without chronic demyelination in spinal injured rats. J. Neurosci. 2012, 32, 5120–5125. [Google Scholar] [CrossRef]
- Manley, N.C.; Priest, C.A.; Denham, J.; Wirth, E.D., 3rd; Lebkowski, J.S. Human Embryonic Stem Cell-Derived Oligodendrocyte Progenitor Cells: Preclinical Efficacy and Safety in Cervical Spinal Cord Injury. Stem Cells Transl. Med. 2017, 6, 1917–1929. [Google Scholar] [CrossRef]
- Sharp, J.; Frame, J.; Siegenthaler, M.; Nistor, G.; Keirstead, H.S. Human embryonic stem cell-derived oligodendrocyte progenitor cell transplants improve recovery after cervical spinal cord injury. Stem Cells 2010, 28, 152–163. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- All, A.H.; Gharibani, P.; Gupta, S.; Bazley, F.A.; Pashai, N.; Chou, B.K.; Shah, S.; Resar, L.M.; Cheng, L.; Gearhart, J.D.; et al. Early intervention for spinal cord injury with human induced pluripotent stem cells oligodendrocyte progenitors. PLoS ONE 2015, 10, e0116933. [Google Scholar] [CrossRef] [Green Version]
- Hawryluk, G.W.; Spano, S.; Chew, D.; Wang, S.; Erwin, M.; Chamankhah, M.; Forgione, N.; Fehlings, M.G. An examination of the mechanisms by which neural precursors augment recovery following spinal cord injury: A key role for remyelination. Cell Transplant. 2014, 23, 365–380. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shin, J.E.; Jung, K.; Kim, M.; Hwang, K.; Lee, H.; Kim, I.S.; Lee, B.H.; Lee, I.S.; Park, K.I. Brain and spinal cord injury repair by implantation of human neural progenitor cells seeded onto polymer scaffolds. Exp. Mol. Med. 2018, 50, 39. [Google Scholar] [CrossRef] [Green Version]
- Salewski, R.P.; Mitchell, R.A.; Li, L.; Shen, C.; Milekovskaia, M.; Nagy, A.; Fehlings, M.G. Transplantation of Induced Pluripotent Stem Cell-Derived Neural Stem Cells Mediate Functional Recovery Following Thoracic Spinal Cord Injury Through Remyelination of Axons. Stem Cells Transl. Med. 2015, 4, 743–754. [Google Scholar] [CrossRef]
- Sankavaram, S.R.; Hakim, R.; Covacu, R.; Frostell, A.; Neumann, S.; Svensson, M.; Brundin, L. Adult Neural Progenitor Cells Transplanted into Spinal Cord Injury Differentiate into Oligodendrocytes, Enhance Myelination, and Contribute to Recovery. Stem Cell Rep. 2019, 12, 950–966. [Google Scholar] [CrossRef] [Green Version]
- Kawabata, S.; Takano, M.; Numasawa-Kuroiwa, Y.; Itakura, G.; Kobayashi, Y.; Nishiyama, Y.; Sugai, K.; Nishimura, S.; Iwai, H.; Isoda, M.; et al. Grafted Human iPS Cell-Derived Oligodendrocyte Precursor Cells Contribute to Robust Remyelination of Demyelinated Axons after Spinal Cord Injury. Stem Cell Rep. 2016, 6, 1–8. [Google Scholar] [CrossRef] [Green Version]
- Hwang, D.H.; Kim, B.G.; Kim, E.J.; Lee, S.I.; Joo, I.S.; Suh-Kim, H.; Sohn, S.; Kim, S.U. Transplantation of human neural stem cells transduced with Olig2 transcription factor improves locomotor recovery and enhances myelination in the white matter of rat spinal cord following contusive injury. BMC Neurosci. 2009, 10, 117. [Google Scholar] [CrossRef] [Green Version]
- Assinck, P.; Sparling, J.S.; Dworski, S.; Duncan, G.J.; Wu, D.L.; Liu, J.; Kwon, B.K.; Biernaskie, J.; Miller, F.D.; Tetzlaff, W. Transplantation of Skin Precursor-Derived Schwann Cells Yields Better Locomotor Outcomes and Reduces Bladder Pathology in Rats with Chronic Spinal Cord Injury. Stem Cell Rep. 2020, 15, 140–155. [Google Scholar] [CrossRef] [PubMed]
- Hou, B.; Ye, Z.; Ji, W.; Cai, M.; Ling, C.; Chen, C.; Guo, Y. Comparison of the Effects of BMSC-derived Schwann Cells and Autologous Schwann Cells on Remyelination Using a Rat Sciatic Nerve Defect Model. Int. J. Biol. Sci. 2018, 14, 1910–1922. [Google Scholar] [CrossRef]
- Deng, L.X.; Deng, P.; Ruan, Y.; Xu, Z.C.; Liu, N.K.; Wen, X.; Smith, G.M.; Xu, X.M. A novel growth-promoting pathway formed by GDNF-overexpressing Schwann cells promotes propriospinal axonal regeneration, synapse formation, and partial recovery of function after spinal cord injury. J. Neurosci. 2013, 33, 5655–5667. [Google Scholar] [CrossRef] [Green Version]
- Feng, S.Q.; Kong, X.H.; Liu, Y.; Ban, D.X.; Ning, G.Z.; Chen, J.T.; Guo, S.F.; Wang, P. Regeneration of spinal cord with cell and gene therapy. Orthop. Surg. 2009, 1, 153–163. [Google Scholar] [CrossRef]
- Zhang, X.; Zeng, Y.; Zhang, W.; Wang, J.; Wu, J.; Li, J. Co-transplantation of neural stem cells and NT-3-overexpressing Schwann cells in transected spinal cord. J. Neurotrauma 2007, 24, 1863–1877. [Google Scholar] [CrossRef] [PubMed]
- Sparling, J.S.; Bretzner, F.; Biernaskie, J.; Assinck, P.; Jiang, Y.; Arisato, H.; Plunet, W.T.; Borisoff, J.; Liu, J.; Miller, F.D.; et al. Schwann cells generated from neonatal skin-derived precursors or neonatal peripheral nerve improve functional recovery after acute transplantation into the partially injured cervical spinal cord of the rat. J. Neurosci. 2015, 35, 6714–6730. [Google Scholar] [CrossRef] [Green Version]
- Gao, S.; Guo, X.; Zhao, S.; Jin, Y.; Zhou, F.; Yuan, P.; Cao, L.; Wang, J.; Qiu, Y.; Sun, C.; et al. Differentiation of human adipose-derived stem cells into neuron/motoneuron-like cells for cell replacement therapy of spinal cord injury. Cell Death Dis. 2019, 10, 597. [Google Scholar] [CrossRef] [PubMed]
- Hong, J.Y.; Lee, S.H.; Lee, S.C.; Kim, J.W.; Kim, K.P.; Kim, S.M.; Tapia, N.; Lim, K.T.; Kim, J.; Ahn, H.S.; et al. Therapeutic potential of induced neural stem cells for spinal cord injury. J. Biol. Chem. 2014, 289, 32512–32525. [Google Scholar] [CrossRef] [Green Version]
- Rosenzweig, E.S.; Brock, J.H.; Lu, P.; Kumamaru, H.; Salegio, E.A.; Kadoya, K.; Weber, J.L.; Liang, J.J.; Moseanko, R.; Hawbecker, S.; et al. Restorative effects of human neural stem cell grafts on the primate spinal cord. Nat. Med. 2018, 24, 484–490. [Google Scholar] [CrossRef]
- Kumamaru, H.; Lu, P.; Rosenzweig, E.S.; Kadoya, K.; Tuszynski, M.H. Regenerating Corticospinal Axons Innervate Phenotypically Appropriate Neurons within Neural Stem Cell Grafts. Cell Rep. 2019, 26, 2329–2339 e2324. [Google Scholar] [CrossRef] [Green Version]
- Kadoya, K.; Lu, P.; Nguyen, K.; Lee-Kubli, C.; Kumamaru, H.; Yao, L.; Knackert, J.; Poplawski, G.; Dulin, J.N.; Strobl, H.; et al. Spinal cord reconstitution with homologous neural grafts enables robust corticospinal regeneration. Nat. Med. 2016, 22, 479–487. [Google Scholar] [CrossRef] [Green Version]
- Meisner, J.G.; Marsh, A.D.; Marsh, D.R. Loss of GABAergic interneurons in laminae I-III of the spinal cord dorsal horn contributes to reduced GABAergic tone and neuropathic pain after spinal cord injury. J. Neurotrauma 2010, 27, 729–737. [Google Scholar] [CrossRef]
- Drew, G.M.; Siddall, P.J.; Duggan, A.W. Mechanical allodynia following contusion injury of the rat spinal cord is associated with loss of GABAergic inhibition in the dorsal horn. Pain 2004, 109, 379–388. [Google Scholar] [CrossRef] [PubMed]
- Kim, D.S.; Jung, S.J.; Nam, T.S.; Jeon, Y.H.; Lee, D.R.; Lee, J.S.; Leem, J.W.; Kim, D.W. Transplantation of GABAergic neurons from ESCs attenuates tactile hypersensitivity following spinal cord injury. Stem Cells 2010, 28, 2099–2108. [Google Scholar] [CrossRef] [PubMed]
- Hwang, I.; Hahm, S.C.; Choi, K.A.; Park, S.H.; Jeong, H.; Yea, J.H.; Kim, J.; Hong, S. Intrathecal Transplantation of Embryonic Stem Cell-Derived Spinal GABAergic Neural Precursor Cells Attenuates Neuropathic Pain in a Spinal Cord Injury Rat Model. Cell Transplant. 2016, 25, 593–607. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chau, C.; Barbeau, H.; Rossignol, S. Effects of intrathecal alpha1- and alpha2-noradrenergic agonists and norepinephrine on locomotion in chronic spinal cats. J. Neurophysiol. 1998, 79, 2941–2963. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kiehn, O.; Sillar, K.T.; Kjaerulff, O.; McDearmid, J.R. Effects of noradrenaline on locomotor rhythm-generating networks in the isolated neonatal rat spinal cord. J. Neurophysiol. 1999, 82, 741–746. [Google Scholar] [CrossRef] [Green Version]
- Cowley, K.C.; Schmidt, B.J. A comparison of motor patterns induced by N-methyl-D-aspartate, acetylcholine and serotonin in the in vitro neonatal rat spinal cord. Neurosci. Lett. 1994, 171, 147–150. [Google Scholar] [CrossRef]
- Feraboli-Lohnherr, D.; Barthe, J.Y.; Orsal, D. Serotonin-induced activation of the network for locomotion in adult spinal rats. J. Neurosci. Res. 1999, 55, 87–98. [Google Scholar] [CrossRef]
- Kwaśniewska, A.; Miazga, K.; Majczyński, H.; Jordan, L.M.; Zawadzka, M.; Sławińska, U. Noradrenergic Components of Locomotor Recovery Induced by Intraspinal Grafting of the Embryonic Brainstem in Adult Paraplegic Rats. Int. J. Mol. Sci. 2020, 21, 5520. [Google Scholar] [CrossRef]
- Liu, J.; Jordan, L.M. Stimulation of the parapyramidal region of the neonatal rat brain stem produces locomotor-like activity involving spinal 5-HT7 and 5-HT2A receptors. J. Neurophysiol. 2005, 94, 1392–1404. [Google Scholar] [CrossRef] [Green Version]
- Miazga, K.; Fabczak, H.; Joachimiak, E.; Zawadzka, M.; Krzemien-Ojak, L.; Bekisz, M.; Bejrowska, A.; Jordan, L.M.; Slawinska, U. Intraspinal Grafting of Serotonergic Neurons Modifies Expression of Genes Important for Functional Recovery in Paraplegic Rats. Neural Plast. 2018, 2018, 4232706. [Google Scholar] [CrossRef] [PubMed]
- Nishimaru, H.; Takizawa, H.; Kudo, N. 5-Hydroxytryptamine-induced locomotor rhythm in the neonatal mouse spinal cord in vitro. Neurosci. Lett. 2000, 280, 187–190. [Google Scholar] [CrossRef]
- Schmidt, B.J.; Jordan, L.M. The role of serotonin in reflex modulation and locomotor rhythm production in the mammalian spinal cord. Brain Res. Bull. 2000, 53, 689–710. [Google Scholar] [CrossRef]
- Sławińska, U.; Miazga, K.; Cabaj, A.M.; Leszczyńska, A.N.; Majczyński, H.; Nagy, J.I.; Jordan, L.M. Grafting of fetal brainstem 5-HT neurons into the sublesional spinal cord of paraplegic rats restores coordinated hindlimb locomotion. Exp. Neurol. 2013, 247, 572–581. [Google Scholar] [CrossRef]
- Slawinska, U.; Miazga, K.; Jordan, L.M. The role of serotonin in the control of locomotor movements and strategies for restoring locomotion after spinal cord injury. Acta Neurobiol. Exp. 2014, 74, 172–187. [Google Scholar]
- Nygren, L.G.; Olson, L. A new major projection from locus coeruleus: The main source of noradrenergic nerve terminals in the ventral and dorsal columns of the spinal cord. Brain Res. 1977, 132, 85–93. [Google Scholar] [CrossRef]
- Rajaofetra, N.; Poulat, P.; Marlier, L.; Geffard, M.; Privat, A. Pre- and postnatal development of noradrenergic projections to the rat spinal cord: An immunocytochemical study. Brain Res. Dev. Brain Res. 1992, 67, 237–246. [Google Scholar] [CrossRef]
- Rajaofetra, N.; Sandillon, F.; Geffard, M.; Privat, A. Pre- and post-natal ontogeny of serotonergic projections to the rat spinal cord. J. Neurosci. Res. 1989, 22, 305–321. [Google Scholar] [CrossRef]
- Nygren, L.G.; Olson, L.; Seiger, A. Mnoaminergic reinnervation of the transected spinal cord by homologous fetal brain grafts. Brain Res. 1977, 129, 227–235. [Google Scholar] [CrossRef]
- Privat, A.; Mansour, H.; Pavy, A.; Geffard, M.; Sandillon, F. Transplantation of dissociated foetal serotonin neurons into the transected spinal cord of adult rats. Neurosci. Lett. 1986, 66, 61–66. [Google Scholar] [CrossRef]
- Ribotta, M.G.; Provencher, J.; Feraboli-Lohnherr, D.; Rossignol, S.; Privat, A.; Orsal, D. Activation of locomotion in adult chronic spinal rats is achieved by transplantation of embryonic raphe cells reinnervating a precise lumbar level. J. Neurosci. 2000, 20, 5144–5152. [Google Scholar] [CrossRef]
- Sławińska, U.; Majczyński, H.; Djavadian, R. Recovery of hindlimb motor functions after spinal cord transection is enhanced by grafts of the embryonic raphe nuclei. Exp. Brain Res. 2000, 132, 27–38. [Google Scholar] [CrossRef] [PubMed]
- Majczynski, H.; Maleszak, K.; Cabaj, A.; Slawinska, U. Serotonin-related enhancement of recovery of hind limb motor functions in spinal rats after grafting of embryonic raphe nuclei. J. Neurotrauma 2005, 22, 590–604. [Google Scholar] [CrossRef]
- Gaspar, P.; Nedelec, S. Serotonin neurons in a dish. Nat. Biotechnol. 2016, 34, 41–42. [Google Scholar] [CrossRef]
- Bartlett, R.D.; Burley, S.; Ip, M.; Phillips, J.B.; Choi, D. Cell Therapies for Spinal Cord Injury: Trends and Challenges of Current Clinical Trials. Neurosurgery 2020, 87, E456–E472. [Google Scholar] [CrossRef]


Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zawadzka, M.; Kwaśniewska, A.; Miazga, K.; Sławińska, U. Perspectives in the Cell-Based Therapies of Various Aspects of the Spinal Cord Injury-Associated Pathologies: Lessons from the Animal Models. Cells 2021, 10, 2995. https://doi.org/10.3390/cells10112995
Zawadzka M, Kwaśniewska A, Miazga K, Sławińska U. Perspectives in the Cell-Based Therapies of Various Aspects of the Spinal Cord Injury-Associated Pathologies: Lessons from the Animal Models. Cells. 2021; 10(11):2995. https://doi.org/10.3390/cells10112995
Chicago/Turabian StyleZawadzka, Małgorzata, Anna Kwaśniewska, Krzysztof Miazga, and Urszula Sławińska. 2021. "Perspectives in the Cell-Based Therapies of Various Aspects of the Spinal Cord Injury-Associated Pathologies: Lessons from the Animal Models" Cells 10, no. 11: 2995. https://doi.org/10.3390/cells10112995