
Ethylene Induced by Sound Stimulation Enhances Anthocyanin Accumulation in Grape Berry Skin through Direct Upregulation of UDP-Glucose: Flavonoid 3-O-Glucosyltransferase


 and
and 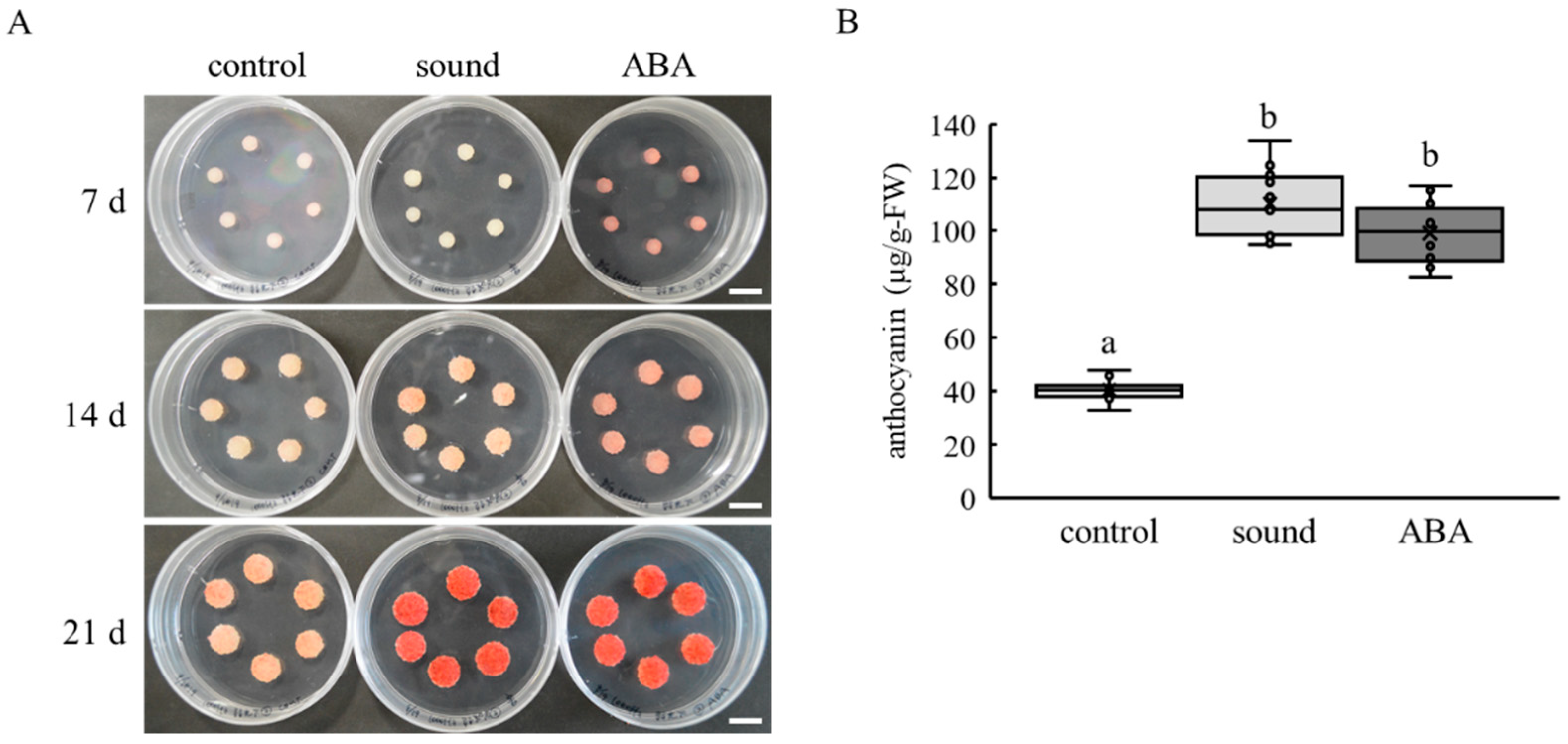{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Materials
2.2. Sound Stimulation of VR Cells
2.3. Sound Stimulation of Bunches on Field-Grown Grapevine
2.4. Measurement of Total Anthocyanin
2.5. Berry Characteristics
2.6. RNA Isolation
2.7. Real-Time RT-PCR Analysis
2.8. Ethephon Treatment of VR Cells
2.9. Ethylene Biosynthesis Inhibitor Treatment of VR Cells Exposed to Sound Stimulation
2.10. Assay of UFGT Promoter in Response to Ethylene
2.11. Statistical Analysis
3. Results
3.1. Sound Stimulation Enhances Anthocyanin Accumulation in VR Cells
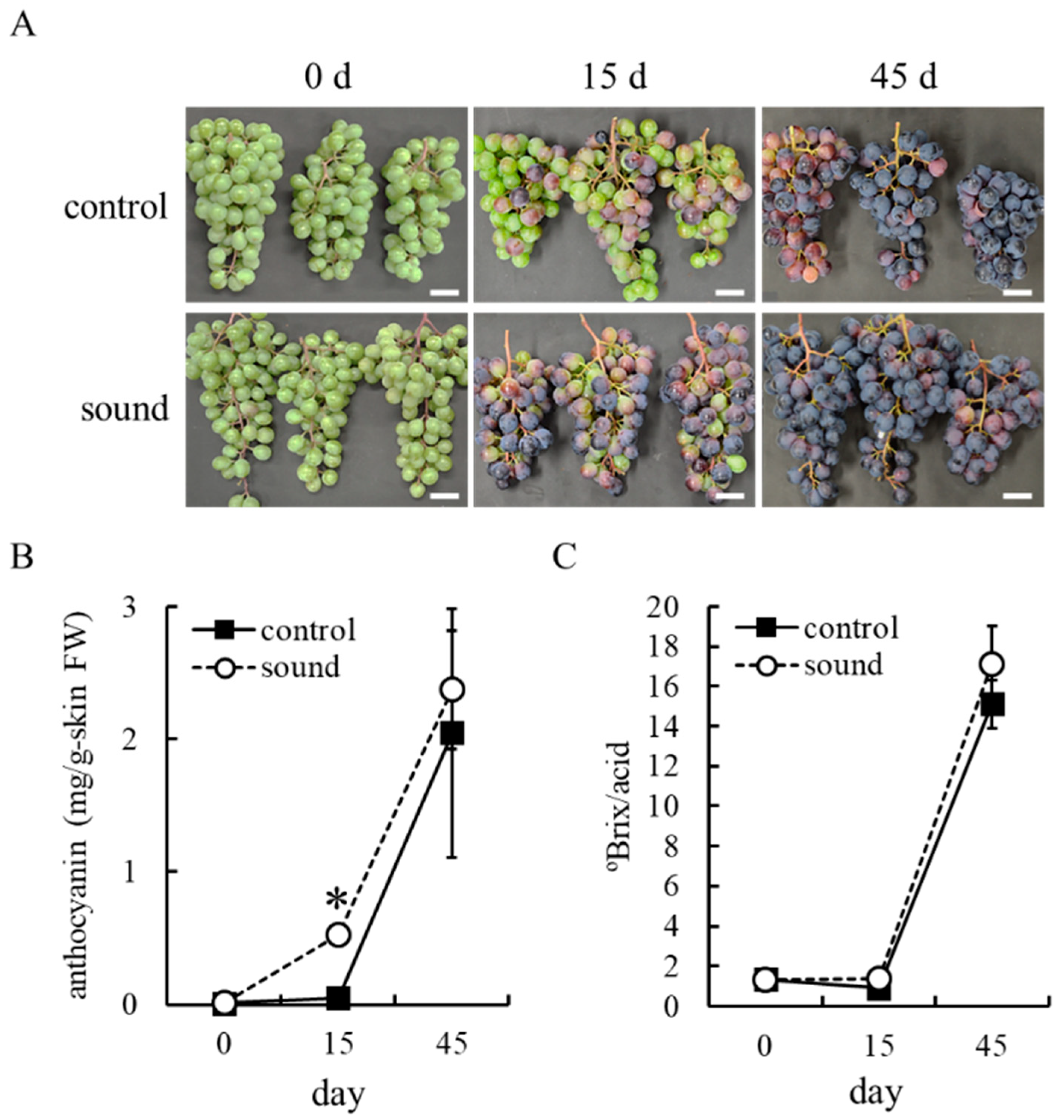
3.2. Sound Stimulation Accelerates Anthocyanin Accumulation in Berry Skins of Field-Grown Grapevines
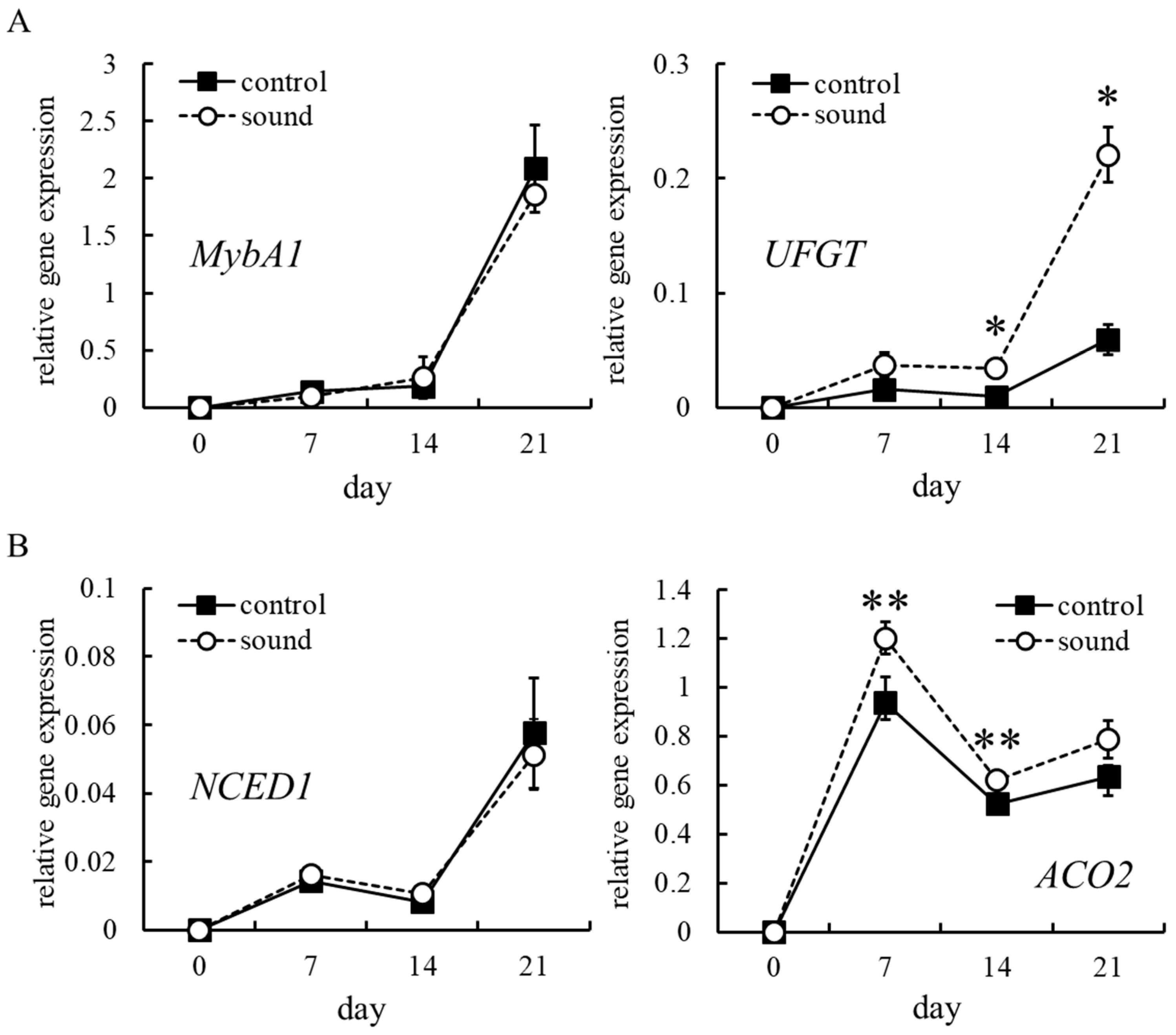
3.3. Effect of Sound Stimulation on MybA1 and UFGT Transcription
3.4. Effect of Sound Stimulation on ABA and Ethylene Biosynthesis
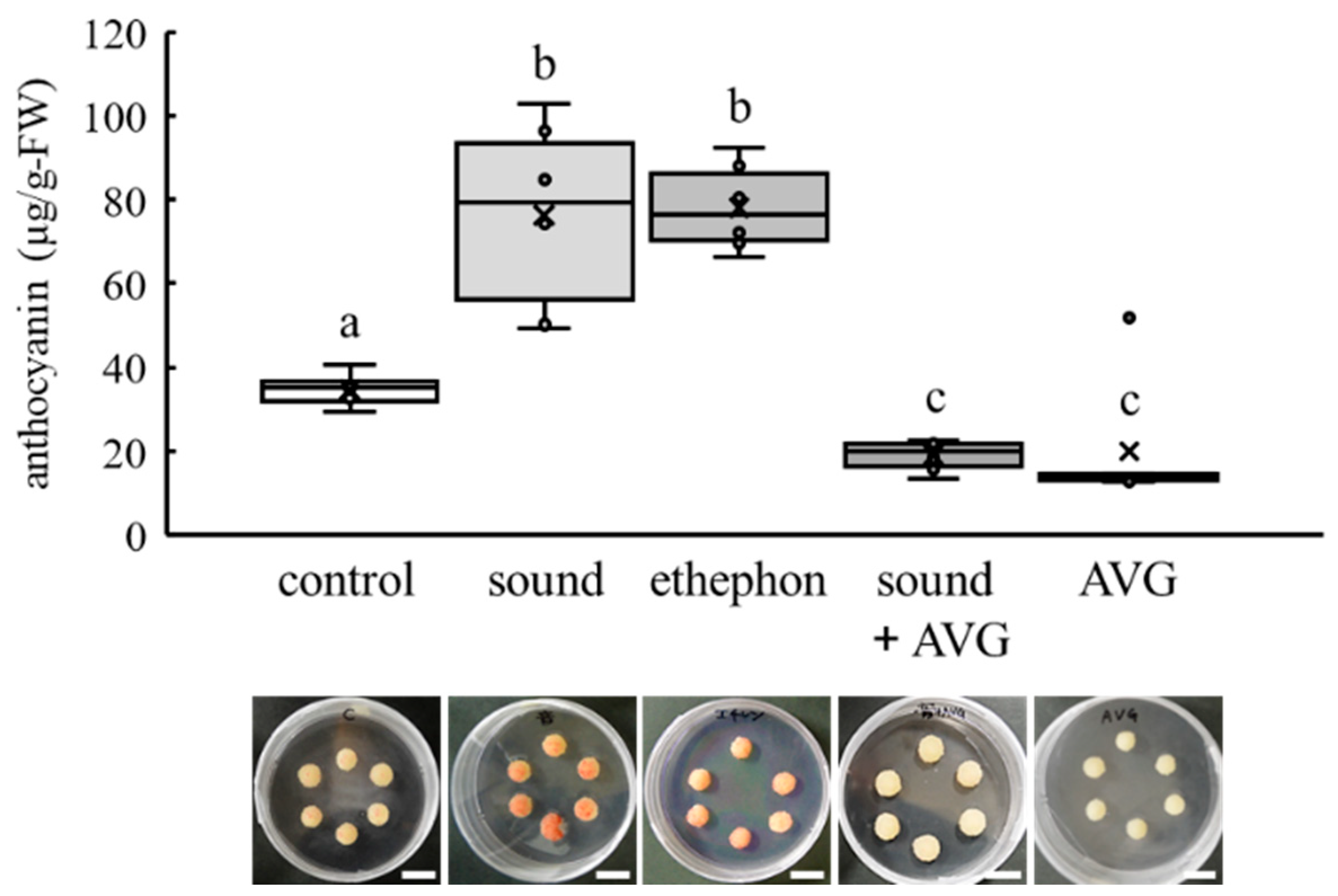
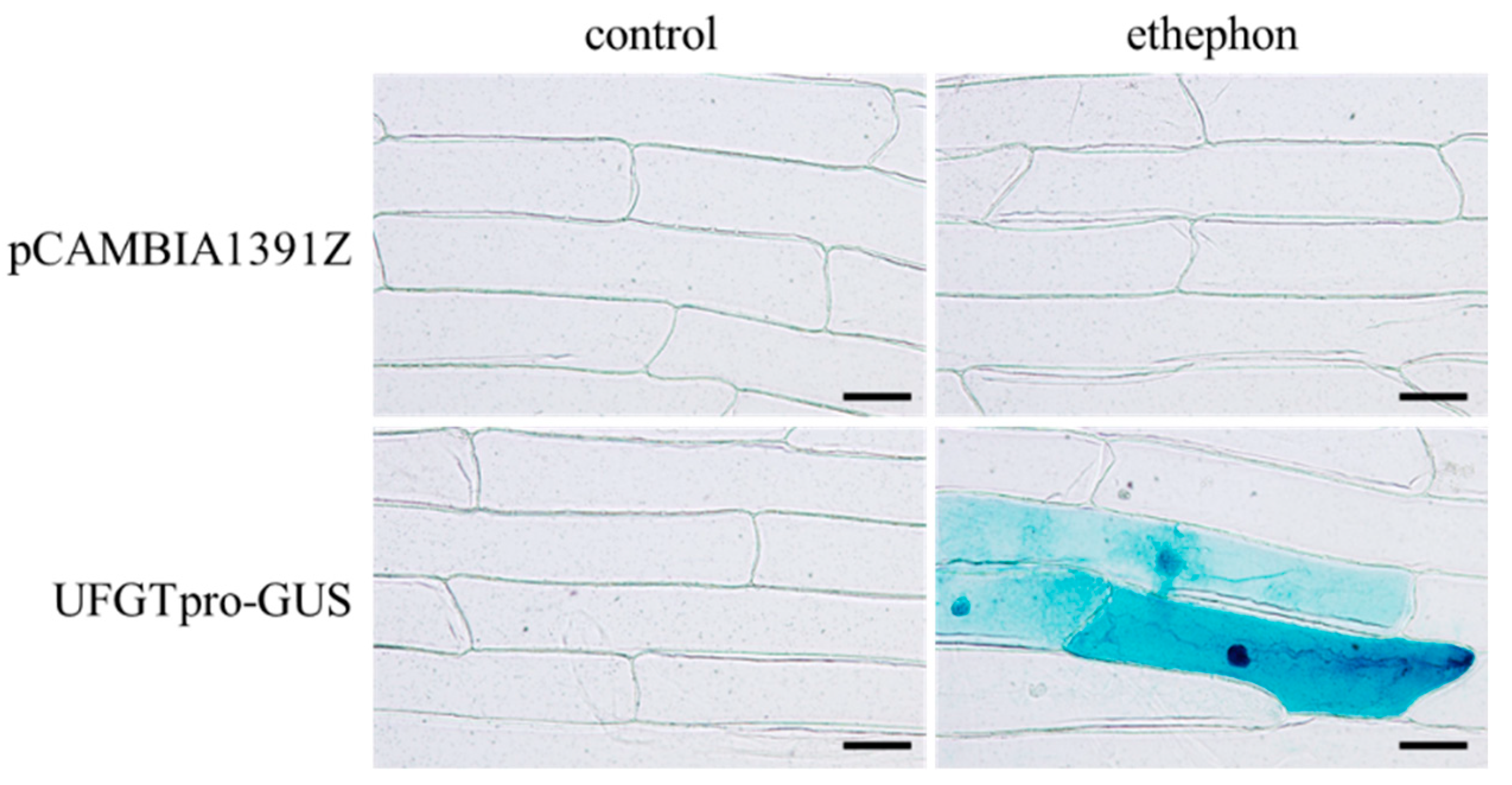
3.5. Activation of UFGT Promoter by Ethephon
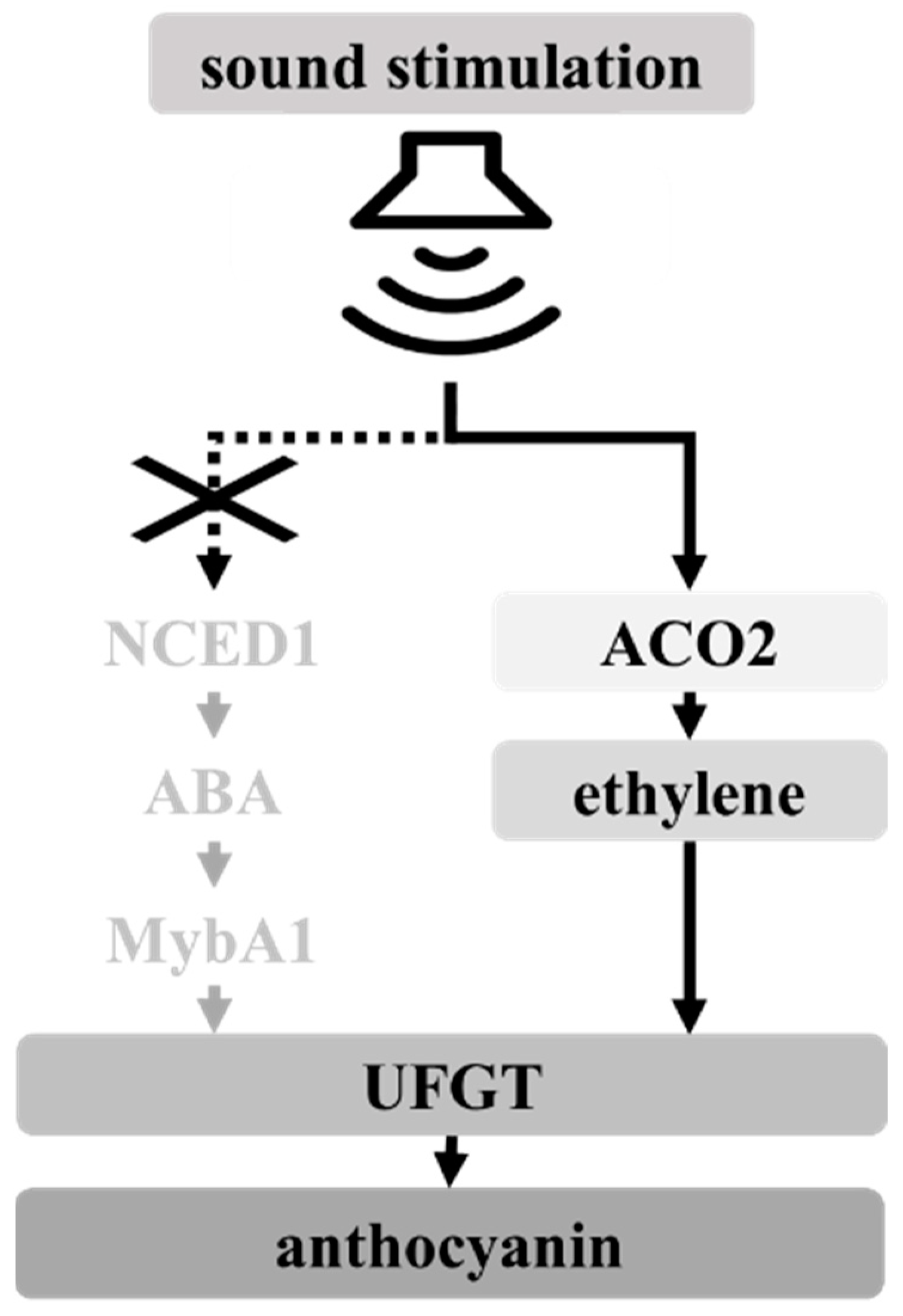
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Pilati, S.; Bagagli, G.; Sonego, P.; Moretto, M.; Brazzale, D.; Castorina, G.; Simoni, L.; Tonelli, C.; Guella, G.; Engelen, K.; et al. Abscisic acid is a major regulator of grape berry ripening onset: New insights into ABA signaling network. Front. Plant Sci. 2017, 8, 1093. [Google Scholar] [CrossRef] [Green Version]
- Koshita, Y.; Asakura, T.; Fukuda, H.; Tsuchida, Y. Nighttime temperature treatment of fruit clusters of ‘Aki Queen’ grapes during maturation and its effect on the skin color and abscisic acid content. Vitis 2007, 46, 208–209. [Google Scholar] [CrossRef]
- Kliewer, W.M.; Torres, R.E. Effect of controlled day and night temperatures on grape coloration. Am. J. Enol. Vitic. 1972, 23, 71–77. [Google Scholar]
- Jones, G.V.; White, M.A.; Cooper, O.R.; Storchmann, K. Climate change and global wine quality. Clim. Chang. 2005, 73, 319–343. [Google Scholar] [CrossRef]
- Saito, N.; Inomata, M.; Tomita, A.; Tezuka, T.; Murakami, Y.; Hagihara, E.; Udo, Y. Global warming tendency in recent years and influence forecasts to fruit production in Yamanashi. In Proceedings of the Annual Meeting of the Society of Agricultural Meteorology of Japan 2008, Yamaguchi, Japan, 20–22 March 2008; p. 81. (In Japanese). [Google Scholar]
- Hannah, L.; Roehrdanz, P.R.; Ikegami, M.; Shepard, A.V.; Shaw, M.R.; Tabor, G.; Zhi, L.; Marquet, P.A.; Hijmans, R.J. Climate change, wine, and conservation. Proc. Natl. Acad. Sci. USA 2013, 110, 6907–6912. [Google Scholar] [CrossRef] [Green Version]
- Morales-Castilla, I.; de Cortázar-Atauri, I.G.; Cook, B.I.; Lacombe, T.; Parker, A.; van Leeuwen, C.; Nicholas, K.A.; Wolkovich, E.M. Diversity buffers winegrowing regions from climate change losses. Proc. Natl. Acad. Sci. USA 2020, 117, 2864–2869. [Google Scholar] [CrossRef] [PubMed]
- Gutiérrez-Gamboa, G.; Zheng, W.; de Toda, F.M. Current viticultural techniques to mitigate the effects of global warming on grape and wine quality: A comprehensive review. Food Res. Int. 2021, 139, 109946. [Google Scholar] [CrossRef] [PubMed]
- Guidoni, S.; Allara, P.; Schubert, A. Effect of cluster thinning on berry skin anthocyanin composition of Vitis vinifera cv. Nebbiolo. Am. J. Enol. Vitic. 2002, 53, 224–226. [Google Scholar]
- Brar, H.S.; Singh, Z.; Swinny, E.; Cameron, I. Girdling and grapevine leafroll associated viruses affect berry weight, colour development and accumulation of anthocyanins in ‘Crimson Seedless’ grapes during maturation and ripening. Plant Sci. 2008, 175, 885–897. [Google Scholar] [CrossRef]
- Matus, J.T.; Loyola, R.; Vega, A.; Neira, A.P.; Bordeu, E.; Johnson, P.A.; Alcalde, J.A. Post-veraison sunlight exposure induces MYB-mediated transcriptional regulation of anthocyanin and flavonol synthesis in berry skins of Vitis vinifera. J. Exp. Bot. 2009, 60, 853–867. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Coventry, J.M.; Fisher, K.H.; Strommer, J.N.; Reynolds, A. Reflective mulch to enhance berry quality in Ontario wine grapes. Acta Hortic. 2005, 689, 95–102. [Google Scholar] [CrossRef]
- Peppi, M.C.; Walker, M.A.; Fidelibus, M.W. Application of abscisic acid rapidly upregulated UFGT gene expression and improved color of grape berries. Vitis 2008, 47, 11–24. [Google Scholar] [CrossRef]
- Ferrara, G.; Mazzeo, A.; Matarrese, A.M.S.; Pacucci, C.; Pacifico, A.; Gambacorta, G.; Faccia, M.; Trani, A.; Gallo, V.; Cafagna, I.; et al. Application of abscisic acid (a-ABA) to ‘Crimson Seedless’ grape berries in a Mediterranean climate: Effects on color, chemical characteristics, metabolic profile, and s-ABA concentration. J. Plant Growth Regul. 2013, 32, 491–505. [Google Scholar] [CrossRef]
- Kataoka, I.; Sugiura, A.; Utsunomiya, N.; Tomana, T. Effect of abscisic acid and defoliation on anthocyanin accumulation in Kyoho grapes (Vitis vinifera L. × V. labruscana Bailey). Vitis 1982, 21, 325–332. [Google Scholar] [CrossRef]
- Zhu, Z.; Li, G.; Yan, C.; Liu, L.; Zhang, Q.; Han, Z.; Li, B. DRL1, encoding a NAC transcription factor, is involved in leaf senescence in grapevine. Int. J. Mol. Sci. 2019, 20, 2678. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hassanien, R.H.; Hou, T.Z.; Li, Y.F.; Li, B.M. Advances in effects of sound waves on plants. J. Integr. Agric. 2014, 13, 335–348. [Google Scholar] [CrossRef]
- Chuanren, D.; Bochu, W.; Wanqian, L.; Jing, C.; Jie, L.; Huan, Z. Effect of chemical and physical factors to improve the germination rate of Echinacea angustifolia seeds. Colloids Surf. B Biointerfaces 2004, 37, 101–105. [Google Scholar] [CrossRef]
- Yi, J.; Bochu, W.; Xiujuan, W.; Chuanren, D.; Xiaocheng, Y. Effect of sound stimulation on roots growth and plasmalemma H+-ATPase activity of chrysanthemum (Gerbera jamesonii). Colloids Surf. B Biointerfaces 2003, 27, 65–69. [Google Scholar] [CrossRef]
- López-Ribera, I.; Vicient, C.M. Drought tolerance induced by sound in Arabidopsis plants. Plant Signal. Behav. 2017, 12, 10. [Google Scholar] [CrossRef] [PubMed]
- Ghosh, R.; Mishra, R.; Choi, B.; Kwon, Y.S.; Bae, D.W.; Park, S.-C.; Jeong, M.-J.; Bae, H. Exposure to sound vibrations lead to transcriptomic, proteomic and hormonal changes in Arabidopsis. Sci. Rep. 2016, 6, 33370. [Google Scholar] [CrossRef]
- Yamakawa, T.; Kato, S.; Ishida, K.; Kodama, T.; Minoda, Y. Production of anthocyanins by Vitis cells in suspension culture. Agric. Biol. Chem. 1983, 47, 2185–2191. [Google Scholar] [CrossRef]
- Moriyama, A.; Nojiri, M.; Watanabe, G.; Enoki, S.; Suzuki, S. Exogenous allantoin improves anthocyanin accumulation in grape berry skin at early stage of ripening. J. Plant Physiol. 2020, 253, 153253. [Google Scholar] [CrossRef] [PubMed]
- Bakker, J.; Preston, N.W.; Timberlake, C.F. The determination of anthocyanins in aging red wines: Comparison of HPLC and spectral methods. Am. J. Enol. Vitic. 1986, 37, 121–126. [Google Scholar]
- Kobayashi, S.; Ishimaru, M.; Ding, C.K.; Yakushiji, H.; Goto, N. Comparison of UDP-glucose:flavonoid 3-O-glucosyltransferase (UFGT) gene sequences between white grapes (Vitis vinifera) and their sports with red skin. Plant Sci. 2001, 160, 543–550. [Google Scholar] [CrossRef]
- Boss, P.K.; Davies, C.; Robinson, S.P. Analysis of the expression of anthocyanin pathway genes in developing Vitis vinifera L. cv Shiraz grape berries and the implications for pathway regulation. Plant Physiol. 1996, 111, 1059–1066. [Google Scholar] [CrossRef] [Green Version]
- Kobayashi, S.; Goto-Yamamoto, N.; Hirokawa, H. Retrotransposon-induced mutations in grape skin color. Science 2004, 304, 982. [Google Scholar] [CrossRef]
- Enoki, S.; Hattori, T.; Ishiai, S.; Tanaka, S.; Mikami, M.; Arita, K.; Nagasaka, S.; Suzuki, S. Vanillylacetone up-regulates expression of genes leading to anthocyanin accumulation by inducing endogenous abscisic acid in grape cell cultures. J. Plant Physiol. 2017, 219, 22–27. [Google Scholar] [CrossRef]
- Iuchi, S.; Kobayashi, M.; Taji, T.; Naramoto, M.; Seki, M.; Kato, T.; Tabata, S.; Kakubari, Y.; Yamaguchi-Shinozaki, K.; Shinozaki, K. Regulation of drought tolerance by gene manipulation of 9-cis-epoxycarotenoid dioxygenase, a key enzyme in abscisic acid biosynthesis in Arabidopsis. Plant J. 2001, 27, 325–333. [Google Scholar] [CrossRef] [Green Version]
- Tira-Umphon, A.; Roustan, J.P.; Chervin, C. The stimulation by ethylene of the UDP glucose-flavonoid 3-O-glucosyltransferase (UFGT) in grape tissues is independent from the MybA transcription factors. Vitis 2007, 46, 210–211. [Google Scholar] [CrossRef]
- Sun, X.; Li, Y.; He, W.; Ji, C.; Xia, P.; Wang, Y.; Du, S.; Li, H.; Raikhel, N.; Xiao, J.; et al. Pyrazinamide and derivatives block ethylene biosynthesis by inhibiting ACC oxidase. Nat. Commun. 2017, 8, 15758. [Google Scholar] [CrossRef]
- Itzhaki, H.; Maxson, J.M.; Woodson, W.R. An ethylene-responsive enhancer element is involved in the senescence-related expression of the carnation glutathione-S-transferase (GST1) gene. Proc. Nat. Acad. Sci. USA 1994, 91, 8925–8929. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, S.F.; Hoffman, N.E. Ethylene biosynthesis and its regulation in higher plants. Annu. Rev. Plant Physiol. 1984, 35, 155–189. [Google Scholar] [CrossRef]
- El-Kereamy, A.; Chervin, C.; Roustan, J.-P.; Cheynier, V.; Souquet, J.-M.; Moutounet, M.; Raynal, J.; Ford, C.; Latché, A.; Pech, J.-C.; et al. Exogenous ethylene stimulates the long-term expression of genes related to anthocyanin biosynthesis in grape berries. Physiol. Plant 2003, 119, 175–182. [Google Scholar] [CrossRef]
- Chervin, C.; Tira-Umphom, A.; Chatelet, P.; Jauneau, A.; Boss, P.K.; Tesniereet, C. Ethylene and other stimuli affect expression of the UDP glucose-flavonoid 3-O-glucosyltransferase in a non-climacteric fruit. Vitis 2009, 48, 11–16. [Google Scholar] [CrossRef]
- Coombe, B.G.; Hale, C.R. The hormone content of ripening grape berries and the effect of growth substance treatments. Plant Physiol. 1973, 51, 629–634. [Google Scholar] [CrossRef] [Green Version]
- Giovannoni, J. Molecular biology of fruit maturation and ripening. Ann. Rev. Plant Physiol. Plant Mol. Biol. 2001, 52, 725–749. [Google Scholar] [CrossRef]
- Chervin, C.; El-Kereamy, A.; Roustan, J.P.; Latche, A.; Lamon, J.; Bouzayen, M. Ethylene seems required for the ripening of grape, a non-climacteric fruit. Plant Sci. 2004, 167, 1301–1305. [Google Scholar] [CrossRef] [Green Version]
- Mishra, R.C.; Ghosh, R.; Bae, H. Plant acoustics: In the search of a sound mechanism for sound signaling in plants. J. Exp. Bot. 2016, 67, 4483–4494. [Google Scholar] [CrossRef]
- Telewski, F.W. A unified hypothesis of mechanoperception in plants. Am. J. Bot. 2006, 93, 1466–1476. [Google Scholar] [CrossRef] [PubMed]
- Jung, J.; Kim, S.-K.; Jung, S.-H.; Jeong, M.-J.; Ryu, C.-M. Sound vibration-triggered epigenetic modulation induces plant root immunity against Ralstonia solanacearum. Front. Microbiol. 2020, 11, 1978. [Google Scholar] [CrossRef] [PubMed]
- Jeong, M.-J.; Shim, C.-K.; Lee, J.-O.; Kwon, H.-B.; Kim, Y.-H.; Lee, S.-K.; Byun, M.-B.; Park, S.-C. Plant gene responses to frequency-specific sound signals. Mol. Breed. 2008, 21, 217–226. [Google Scholar] [CrossRef]
- Ghosh, R.; Gururani, M.A.; Ponpandian, L.N.; Mishra, R.C.; Park, S.-C.; Jeong, M.-J.; Bae, H. Expression analysis of sound vibration-regulated genes by touch treatment in Arabidopsis. Front. Plant Sci. 2017, 8, 100. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fraga, H.; Malheiro, A.C.; Moutinho-Pereira, J.; Santos, J.A. An overview of climate change impacts on European viticulture. Food Energy Secur. 2012, 1, 94–110. [Google Scholar] [CrossRef]
- Portu, J.; López, R.; Baroja, E.; Santamaría, P.; Garde-Cerdán, T. Improvement of grape and wine phenolic content by foliar application to grapevine of three different elicitors: Methyl jasmonate, chitosan, and yeast extract. Food Chem. 2016, 201, 213–221. [Google Scholar] [CrossRef] [PubMed]
- Mikami, M.; Mori, D.; Masumura, Y.; Aoki, Y.; Suzuki, S. Electrical stimulation: An abiotic stress generator for enhancing anthocyanin and resveratrol accumulation in grape berry. Sci. Hortic. 2017, 226, 285–292. [Google Scholar] [CrossRef]
- Tsavkelova, E.A.; Klimova, S.Y.; Cherdyntseva, T.A.; Netrusov, A.I. Hormones and hormone-like substances of microorganisms: A review. Appl. Biochem. Microbiol. 2006, 42, 229–235. [Google Scholar] [CrossRef]
- Yu, M.; Chen, J.C.; Qu, J.Z.; Liu, F.; Zhou, M.; Ma, Y.M.; Xiang, S.Y.; Pan, X.X.; Zhang, H.B.; Yang, M.Z. Exposure to endophytic fungi quantitatively and compositionally alters anthocyanins in grape cells. Plant Physiol. Biochem. 2020, 149, 144–152. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.-Y.; Lee, J.-S.; Kwon, T.-R.; Lee, S.-I.; Kim, J.-A.; Lee, G.-M.; Park, S.-C.; Jeong, M.-J. Sound waves delay tomato fruit ripening by negatively regulating ethylene biosynthesis and signaling genes. Postharvest Biol. Technol. 2015, 110, 43–50. [Google Scholar] [CrossRef] [Green Version]






Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yamazaki, M.; Ishida, A.; Suzuki, Y.; Aoki, Y.; Suzuki, S.; Enoki, S. Ethylene Induced by Sound Stimulation Enhances Anthocyanin Accumulation in Grape Berry Skin through Direct Upregulation of UDP-Glucose: Flavonoid 3-O-Glucosyltransferase. Cells 2021, 10, 2799. https://doi.org/10.3390/cells10102799
Yamazaki M, Ishida A, Suzuki Y, Aoki Y, Suzuki S, Enoki S. Ethylene Induced by Sound Stimulation Enhances Anthocyanin Accumulation in Grape Berry Skin through Direct Upregulation of UDP-Glucose: Flavonoid 3-O-Glucosyltransferase. Cells. 2021; 10(10):2799. https://doi.org/10.3390/cells10102799
Chicago/Turabian StyleYamazaki, Mone, Akari Ishida, Yutaka Suzuki, Yoshinao Aoki, Shunji Suzuki, and Shinichi Enoki. 2021. "Ethylene Induced by Sound Stimulation Enhances Anthocyanin Accumulation in Grape Berry Skin through Direct Upregulation of UDP-Glucose: Flavonoid 3-O-Glucosyltransferase" Cells 10, no. 10: 2799. https://doi.org/10.3390/cells10102799