
What Do We Know about Pruritus in Very Young Infants? A Literature Review

 , ,
, ,  and
and 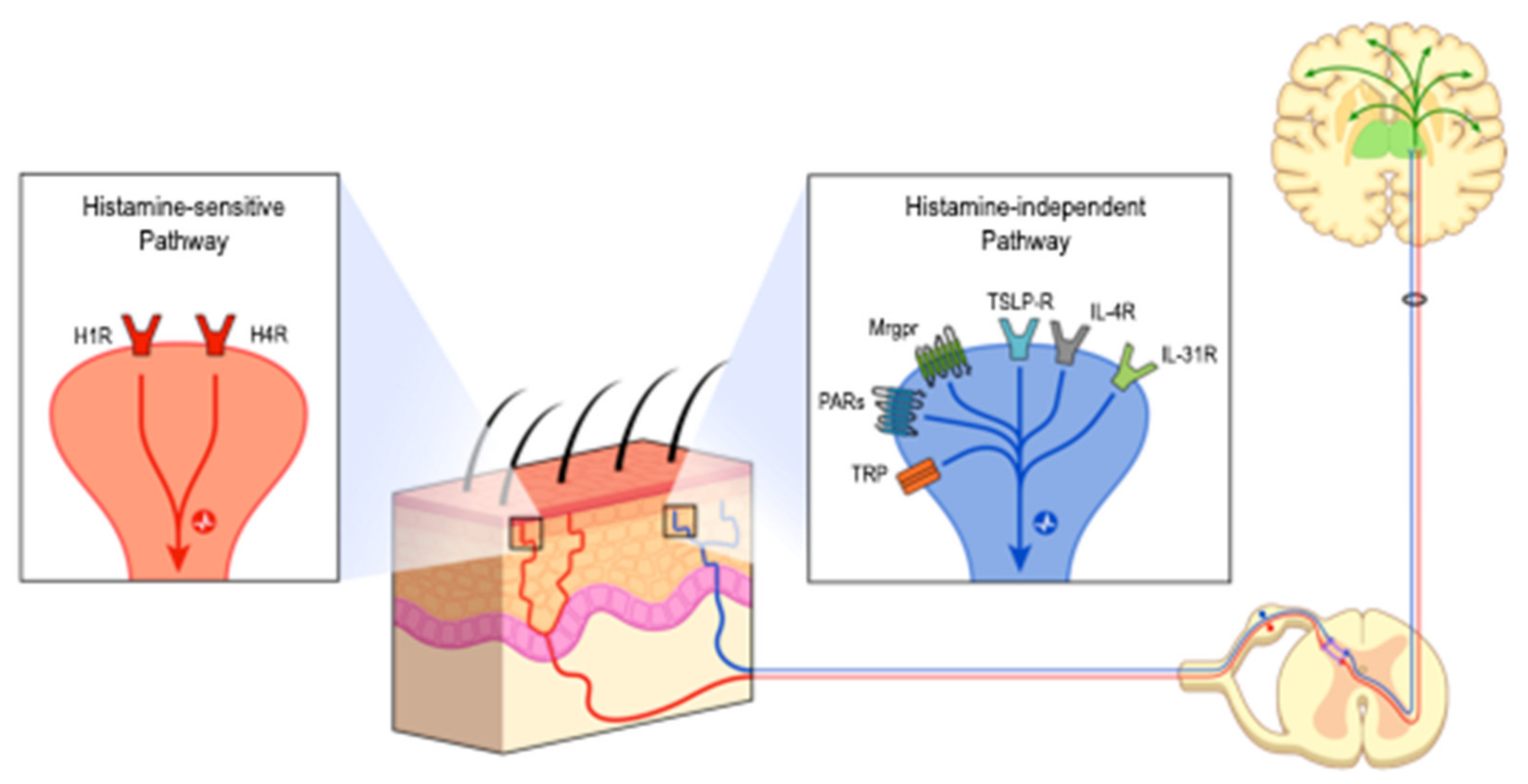{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
3. Results
3.1. Neurophysiology of Pruritus—Link between Pain and Pruritus
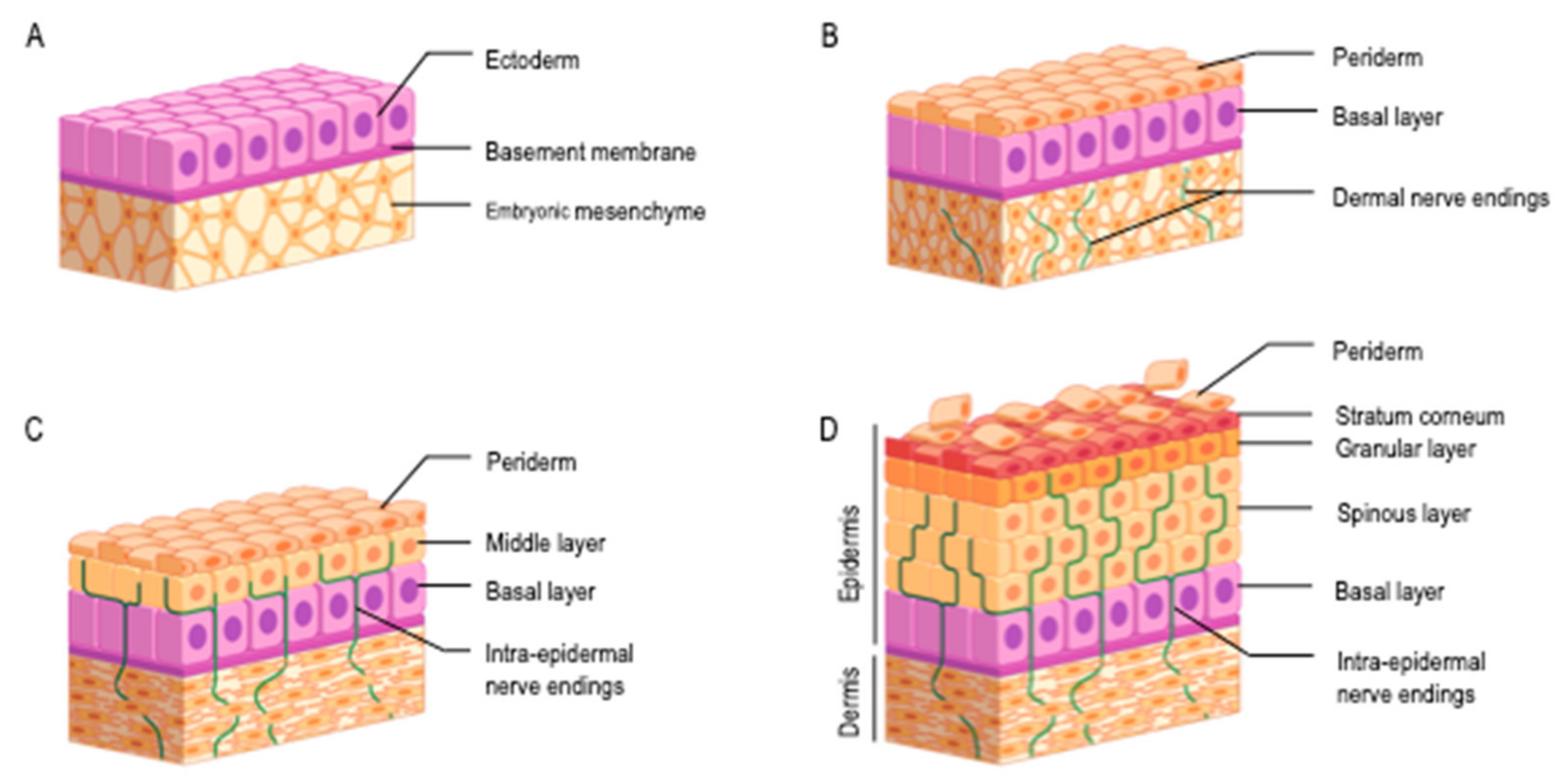
3.2. Newborn Skin: From Intra-Uterine Development to Neonatal Adaptation
3.3. Neurosensory System and Infant Skin Features Bring Clues in Favor of Infants’ Ability to Experience Pruritus
3.4. Pruritus during Infancy: Etiology, Epidemiology
3.5. Impact of Pruritus: The Example of Atopic Dermatitis (AD)
3.6. Assessment of Pruritus in Young Infants: A Need for Further Research
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Ikoma, A.; Steinhoff, M.; Ständer, S.; Yosipovitch, G.; Schmelz, M. The Neurobiology of Itch. Nat. Rev. Neurosci. 2006, 7, 535–547. [Google Scholar] [CrossRef]
- Misery, L.; Ständer, S. Pruritus, 2nd ed.; Springer: London, UK, 2016; ISBN 978-3-319-33140-9. [Google Scholar]
- Yosipovitch, G.; Rosen, J.D.; Hashimoto, T. Itch: From Mechanism to (Novel) Therapeutic Approaches. J. Allergy Clin. Immunol. 2018, 142, 1375–1390. [Google Scholar] [CrossRef] [Green Version]
- Cevikbas, F.; Lerner, E.A. Physiology and Pathophysiology of Itch. Physiol. Rev. 2020, 100, 945–982. [Google Scholar] [CrossRef]
- Ständer, S.; Schmelz, M. Chronic Itch and Pain-Similarities and Differences. Eur. J. Pain 2006, 10, 473–478. [Google Scholar] [CrossRef] [PubMed]
- Yosipovitch, G.; Carstens, E.; McGlone, F. Chronic Itch and Chronic Pain: Analogous Mechanisms. Pain 2007, 131, 4–7. [Google Scholar] [CrossRef]
- Ersch, J. Assessing Gestational Age from Histology of Fetal Skin: An Autopsy Study of 379 Fetuses. Obstet. Gynecol. 1999, 94, 753–757. [Google Scholar] [CrossRef]
- Serri, F.; Montagna, W.; Mescon, H. Studies of the Skin of the Fetus and the Child. J. Investig. Dermatol. 1962, 39, 199–217. [Google Scholar] [CrossRef] [Green Version]
- Terenghi, G.; Sundaresan, M.; Moscoso, G.; Polak, J.M. Neuropeptides and a Neuronal Marker in Cutaneous Innervation during Human Foetal Development. J. Comp. Neurol. 1993, 328, 595–603. [Google Scholar] [CrossRef] [PubMed]
- Holbrook, K.A. Human Epidermal Embryogenesis. Int. J. Dermatol. 1979, 18, 329–356. [Google Scholar] [CrossRef]
- Holbrook, K.A.; Odland, G.F. Regional Development of the Human Epidermis in the First Trimester Embryo and the Second Trimester Fetus (Ages Related to the Timing of Amniocentesis and Fetal Biopsy). J. Investig. Dermatol. 1980, 74, 161–168. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Visscher, M.; Narendran, V. The Ontogeny of Skin. Adv. Wound Care 2014, 3, 291–303. [Google Scholar] [CrossRef] [Green Version]
- Coughlin, C.C.; Taïeb, A. Evolving Concepts of Neonatal Skin. Pediatr. Dermatol. 2014, 31, 5–8. [Google Scholar] [CrossRef] [Green Version]
- Lane, A.T. Human Fetal Skin Development. Pediatr. Dermatol. 1986, 3, 487–491. [Google Scholar] [CrossRef]
- Chiou, Y.B.; Blume-Peytavi, U. Stratum Corneum Maturation. Skin. Pharmacol. Physiol. 2004, 17, 57–66. [Google Scholar] [CrossRef] [PubMed]
- Williams, M.L.; Hanley, K.; Elias, P.M.; Feingold, K.R. Ontogeny of the Epidermal Permeability Barrier. J. Investig. Dermatol. Symp. Proc. 1998, 3, 75–79. [Google Scholar] [CrossRef] [Green Version]
- Scott, I.R.; Harding, C.R. Filaggrin Breakdown to Water Binding Compounds during Development of the Rat Stratum Corneum Is Controlled by the Water Activity of the Environment. Dev. Biol. 1986, 115, 84–92. [Google Scholar] [CrossRef]
- Coolen, N.A.; Schouten, K.C.W.M.; Middelkoop, E.; Ulrich, M.M.W. Comparison between Human Fetal and Adult Skin. Arch. Dermatol. Res. 2010, 302, 47–55. [Google Scholar] [CrossRef] [Green Version]
- Valman, H.B.; Pearson, J.F. What the fetus feels. Br. Med. J. 1980, 280, 233–234. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lecanuet, J.-P.; Schaal, B. Fetal Sensory Competencies. Eur. J. Obstet. Gynecol. Reprod. Biol. 1996, 68, 1–23. [Google Scholar] [CrossRef]
- Moore, S.J.; Munger, B.L. The Early Ontogeny of the Afferent Nerves and Papillary Ridges in Human Digital Glabrous Skin. Dev. Brain Res. 1989, 48, 119–141. [Google Scholar] [CrossRef]
- Saxod, R. Ontogeny of the Cutaneous Sensory Organs. Micros. Res. Tech. 1996, 34, 313–333. [Google Scholar] [CrossRef]
- Belle, M.; Godefroy, D.; Couly, G.; Malone, S.A.; Collier, F.; Giacobini, P.; Chédotal, A. Tridimensional Visualization and Analysis of Early Human Development. Cell 2017, 169, 161–173. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fluhr, J.W.; Darlenski, R.; Taieb, A.; Hachem, J.-P.; Baudouin, C.; Msika, P.; De Belilovsky, C.; Berardesca, E. Functional Skin Adaptation in Infancy—Almost Complete but Not Fully Competent: Functional Skin Adaptation. Exp. Dermatol. 2010, 19, 483–492. [Google Scholar] [CrossRef] [PubMed]
- Fluhr, J.W.; Darlenski, R.; Lachmann, N.; Baudouin, C.; Msika, P.; Belilovsky, C.D.; Hachem, J.-P. Infant Epidermal Skin Physiology: Adaptation after Birth. Br. J. Dermatol. 2012, 166, 483–490. [Google Scholar] [CrossRef] [PubMed]
- Nikolovski, J.; Stamatas, G.N.; Kollias, N.; Wiegand, B.C. Barrier Function and Water-Holding and Transport Properties of Infant Stratum Corneum Are Different from Adult and Continue to Develop through the First Year of Life. J. Investig. Dermatol. 2008, 128, 1728–1736. [Google Scholar] [CrossRef]
- Stamatas, G.N.; Nikolovski, J.; Mack, M.C.; Kollias, N. Infant Skin Physiology and Development during the First Years of Life: A Review of Recent Findings Based on in Vivo Studies: Infant Skin Physiology and Development. Int. J. Cosmet. Sci. 2011, 33, 17–24. [Google Scholar] [CrossRef]
- Hoeger, P.H.; Enzmann, C.C. Skin Physiology of the Neonate and Young Infant: A Prospective Study of Functional Skin Parameters During Early Infancy. Pediatr. Dermatol. 2002, 19, 256–262. [Google Scholar] [CrossRef] [PubMed]
- Harpin, V.A.; Rutter, N. Development of Emotional Sweating in the Newborn Infant. Arch. Dis. Child. 1982, 57, 691–695. [Google Scholar] [CrossRef] [PubMed]
- Anand, K.J.; Hickey, P.R. Pain and Its Effects in the Human Neonate and Fetus. N. Engl. J. Med. 1987, 317, 1321–1329. [Google Scholar] [CrossRef] [Green Version]
- Flower, M.J. Neuromaturation of the Human Fetus. J. Med. Philos. 1985, 10, 237–252. [Google Scholar] [CrossRef]
- Lee, C.-H.; Chuang, H.-Y.; Shih, C.-C.; Jong, S.-B.; Chang, C.-H.; Yu, H.-S. Transepidermal Water Loss, Serum IgE and Beta-Endorphin as Important and Independent Biological Markers for Development of Itch Intensity in Atopic Dermatitis. Br. J. Dermatol. 2006, 154, 1100–1107. [Google Scholar] [CrossRef]
- Fagard, J. Le Développement des Habiletés de l’Enfant; CNRS Editions: Paris, France, 2001; ISBN 978-2-271-09137-6. [Google Scholar]
- Weisshaar, E.; Szepietowski, J.; Darsow, U.; Misery, L.; Wallengren, J.; Mettang, T.; Gieler, U.; Lotti, T.; Lambert, J.; Maisel, P.; et al. European Guideline on Chronic Pruritus. Acta Derm. Venerol. 2012, 92, 563–581. [Google Scholar] [CrossRef] [Green Version]
- Ständer, S.; Schäfer, I.; Phan, N.Q.; Blome, C.; Herberger, K.; Heigel, H.; Augustin, M. Prevalence of Chronic Pruritus in Germany: Results of a Cross-Sectional Study in a Sample Working Population of 11,730. Dermatology 2010, 221, 229–235. [Google Scholar] [CrossRef]
- Dalgard, F.; Svensson, A.; Holm, J.O.; Sundby, J. Self-Reported Skin Morbidity in Oslo. Associations with Sociodemographic Factors among Adults in a Cross-Sectional Study. Br. J. Dermatol. 2004, 151, 452–457. [Google Scholar] [CrossRef]
- Misery, L.; Rahhali, N.; Duhamel, A.; Taieb, C. Epidemiology of Pruritus in France. Acta Derm. Venerol. 2012, 92, 541–542. [Google Scholar] [CrossRef]
- Halvorsen, J.A.; Olesen, A.B.; Thoresen, M.; Holm, J.-Ø.; Bjertness, E.; Dalgard, F. Comparison of Self-Reported Skin Complaints with Objective Skin Signs Among Adolescents. Acta Derm. Venereol. 2008, 88, 573–577. [Google Scholar] [CrossRef] [PubMed]
- Fölster-Holst, R. Itch Management in Childhood. In Current Problems in Dermatology; Szepietowski, J., Weisshaar, E., Eds.; S. Karger AG: Basel, Switzerland, 2016; Volume 50, pp. 173–191. ISBN 978-3-318-05888-8. [Google Scholar]
- Metz, M.; Wahn, U.; Gieler, U.; Stock, P.; Schmitt, J.; Blume-Peytavi, U. Chronic Pruritus Associated with Dermatologic Disease in Infancy and Childhood: Update from an Interdisciplinary Group of Dermatologists and Pediatricians. Pediatr. Allergy Immunol. 2013, 24, 527–539. [Google Scholar] [CrossRef] [PubMed]
- Carroll, C.L.; Balkrishnan, R.; Feldman, S.R.; Fleischer, A.B., Jr.; Manuel, J.C. The Burden of Atopic Dermatitis: Impact on the Patient, Family, and Society. Pediatr. Dermatol. 2005, 22, 192–199. [Google Scholar] [CrossRef] [PubMed]
- Drucker, A.M.; Wang, A.R.; Li, W.-Q.; Sevetson, E.; Block, J.K.; Qureshi, A.A. The Burden of Atopic Dermatitis: Summary of a Report for the National Eczema Association. J. Investig. Dermatol. 2017, 137, 26–30. [Google Scholar] [CrossRef] [Green Version]
- Schmitt, J.; Chen, C.-M.; Apfelbacher, C.; Romanos, M.; Lehmann, I.; Herbarth, O.; Schaaf, B.; Kraemer, U.; von Berg, A.; Wichmann, H.-E.; et al. Infant Eczema, Infant Sleeping Problems, and Mental Health at 10 Years of Age: The Prospective Birth Cohort Study LISAplus: Infant Eczema, Sleep Problems & Mental Health at Age 10. Allergy 2011, 66, 404–411. [Google Scholar] [CrossRef]
- Nettis, E.; Ortoncelli, M.; Pellacani, G.; Foti, C.; Di Leo, E.; Patruno, C.; Rongioletti, F.; Argenziano, G.; Ferrucci, S.; Macchia, L.; et al. A Multicenter Study on the Prevalence of Clinical Patterns and Clinical Phenotypes in Adult Atopic Dermatitis. J. Investig. Allergol. Clin. Immunol. 2020, 30, 448–450. [Google Scholar] [CrossRef] [PubMed]
- Yaghmaie, P.; Koudelka, C.W.; Simpson, E.L. Mental Health Comorbidity in Patients with Atopic Dermatitis. J. Allergy Clin. Immunol. 2013, 131, 428–433. [Google Scholar] [CrossRef] [Green Version]
- Lewis-Jones, S. Quality of Life and Childhood Atopic Dermatitis: The Misery of Living with Childhood Eczema: Atopic eczema/quality of life. Int. J. Clin. Pract. 2006, 60, 984–992. [Google Scholar] [CrossRef]
- Chamlin, S.L.; Frieden, I.J.; Williams, M.L.; Chren, M.-M. Effects of Atopic Dermatitis on Young American Children and Their Families. Pediatrics 2004, 114, 607–611. [Google Scholar] [CrossRef]
- Fang, M.M.; Nowinski, C.J.; Lai, J.; Shaunfield, S.; Silverberg, J.I.; Rangel, S.M.; Cella, D.; Paller, A.S. Characteristics and Impacts of Itch in Children with Inflammatory Skin Disorders. Br. J. Dermatol. 2021, 184, 896–904. [Google Scholar] [CrossRef] [PubMed]
- Pereira, M.P.; Ständer, S. Measurement Tools for Chronic Pruritus: Assessment of the Symptom and the Associated Burden. Itch 2019, 4, e29. [Google Scholar] [CrossRef]
- Kong, H.E.; Francois, S.; Smith, S.; Spraker, M.; Lawley, L.P.; Lee, G.; Chen, K.-H.; Roberts, J.S.; Chen, S.C. Pruritus Assessment Tools for 6 to 7-year-old Children: KidsItchyQoL and ItchyQuant. Pediatr. Dermatol. 2021, 38, 591–601. [Google Scholar] [CrossRef] [PubMed]
- Bringhurst, C.; Waterston, K.; Schofield, O.; Benjamin, K.; Rees, J.L. Measurement of Itch Using Actigraphy in Pediatric and Adult Populations. J. Am. Acad. Dermatol. 2004, 51, 893–898. [Google Scholar] [CrossRef]
- Rioualen, S.; Roué, J.-M.; Lefranc, J.; Gouillou, M.; Noak, E.; Alavi, Z.; Dubourg, M.; Sizun, J. Actigraphy Is Not a Reliable Method for Measuring Sleep Patterns in Neonates. Acta Paediatr. 2015, 104, 478–482. [Google Scholar] [CrossRef] [PubMed]
- Grunau, R.V.E.; Craig, K.D. Pain Expression in Neonates Facial Action and Cry. Pain 1987, 28, 395–410. [Google Scholar] [CrossRef]
- Grunau, R.V.E.; Johnston, C.C.; Craig, K.D. Neonatal Facial and Cry Responsesto Invasive and Non-Invasive Procedures. Pain 1990, 42, 295–305. [Google Scholar] [CrossRef]
- Anand, K.J.; Stevens, B.J.; McGrath, P. Pain in Neonates and Infants: Pain Research and Clinical Management; Elsevier: Amsterdam, The Netherlands, 2007; Volume 10, ISBN 978-0-444-52061-6. [Google Scholar]


Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Le Pors, C.; Talagas, M.; Abasq-Thomas, C.; Henry, S.; Misery, L.; Roué, J.-M. What Do We Know about Pruritus in Very Young Infants? A Literature Review. Cells 2021, 10, 2788. https://doi.org/10.3390/cells10102788
Le Pors C, Talagas M, Abasq-Thomas C, Henry S, Misery L, Roué J-M. What Do We Know about Pruritus in Very Young Infants? A Literature Review. Cells. 2021; 10(10):2788. https://doi.org/10.3390/cells10102788
Chicago/Turabian StyleLe Pors, Camille, Matthieu Talagas, Claire Abasq-Thomas, Séverine Henry, Laurent Misery, and Jean-Michel Roué. 2021. "What Do We Know about Pruritus in Very Young Infants? A Literature Review" Cells 10, no. 10: 2788. https://doi.org/10.3390/cells10102788