Metabolite-Targeted Analysis and Physiological Traits of Zea mays L. in Response to Application of a Leonardite-Humate and Lignosulfonate-Based Products for Their Evaluation as Potential Biostimulants

 ,
,  , , ,
, , ,
Abstract
:1. Introduction
2. Results
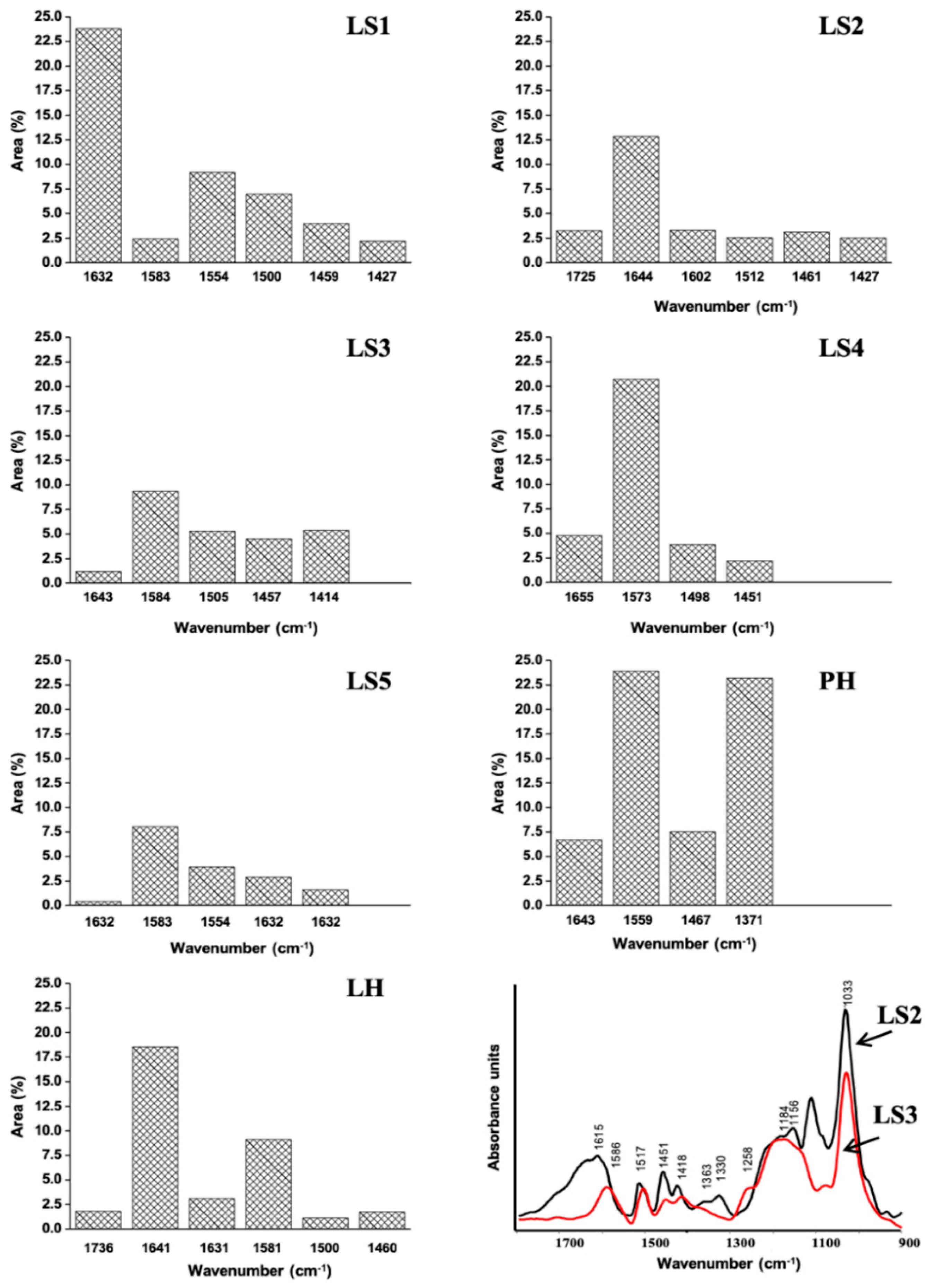
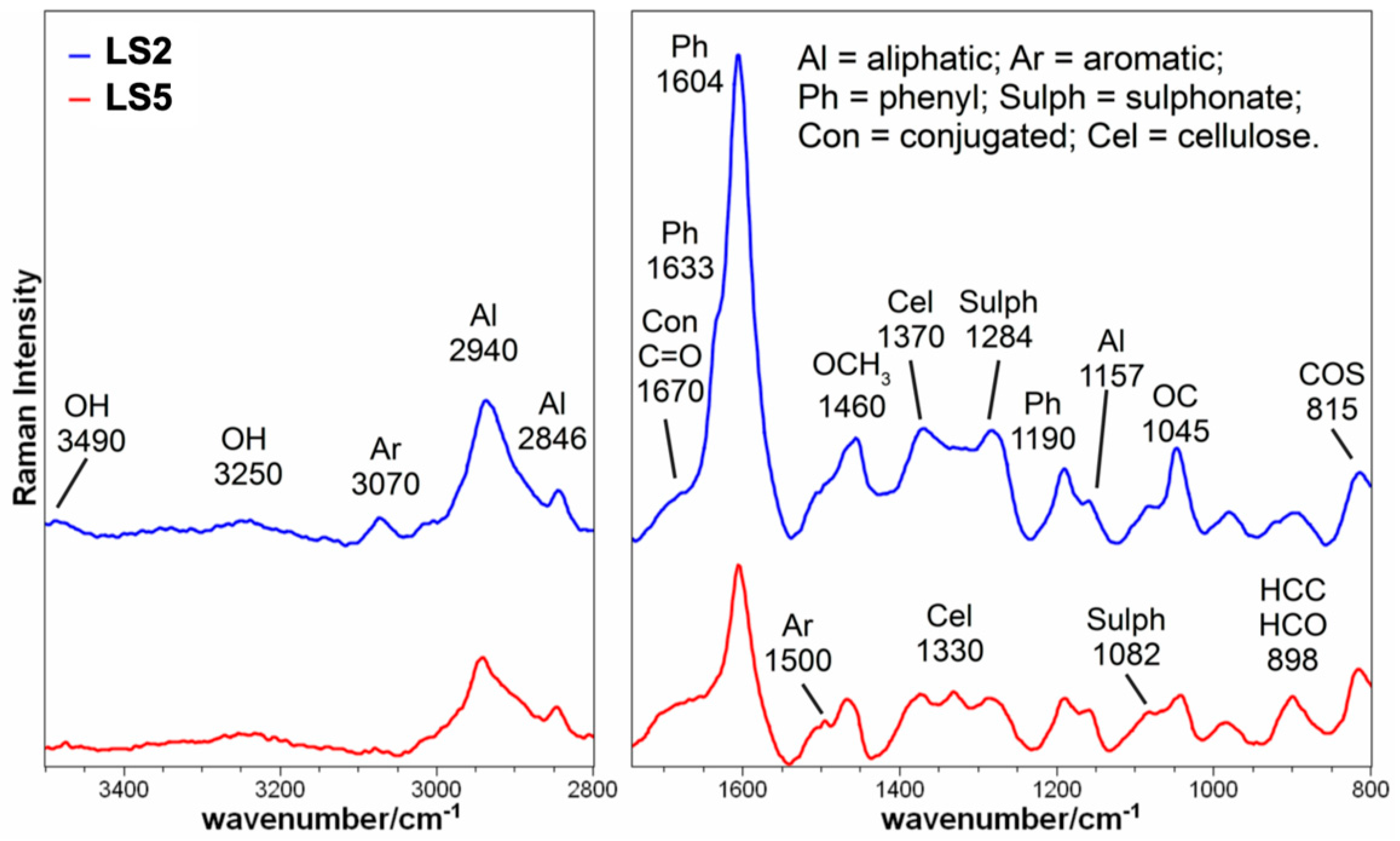
2.1. Chemical and Spectroscopic Features of Hs
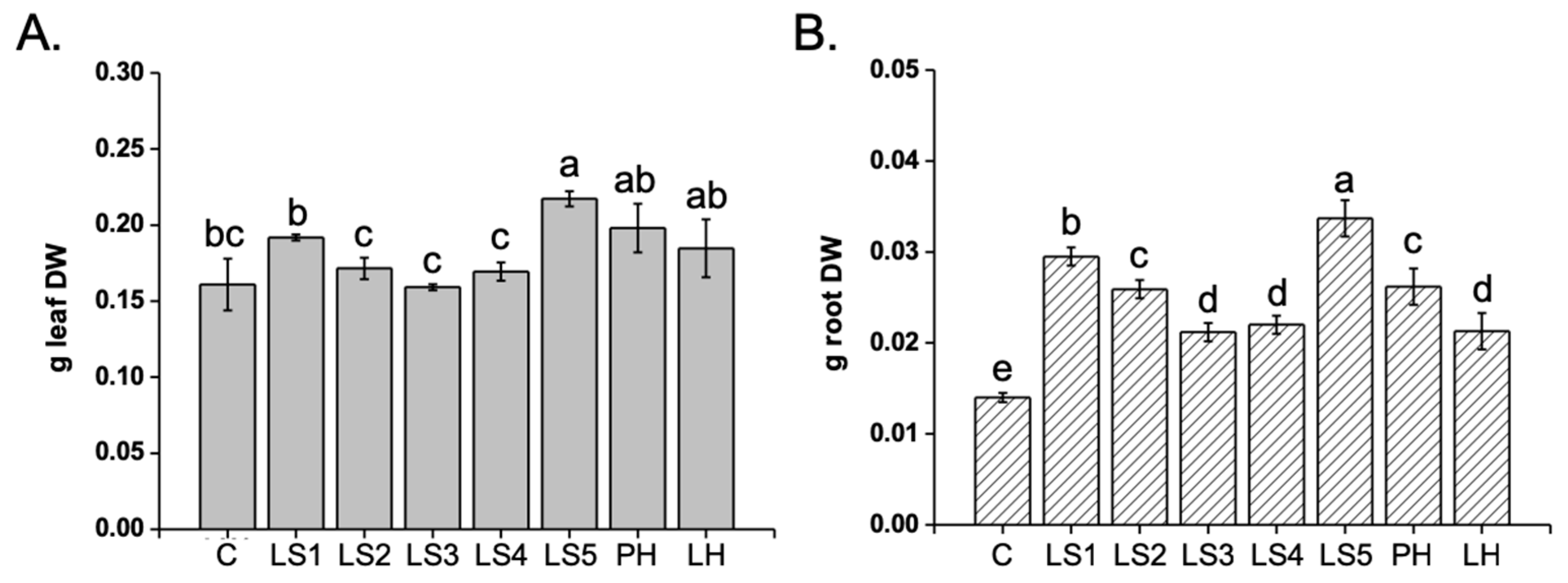
2.2. Effect of Hs on Maize Plant Growth
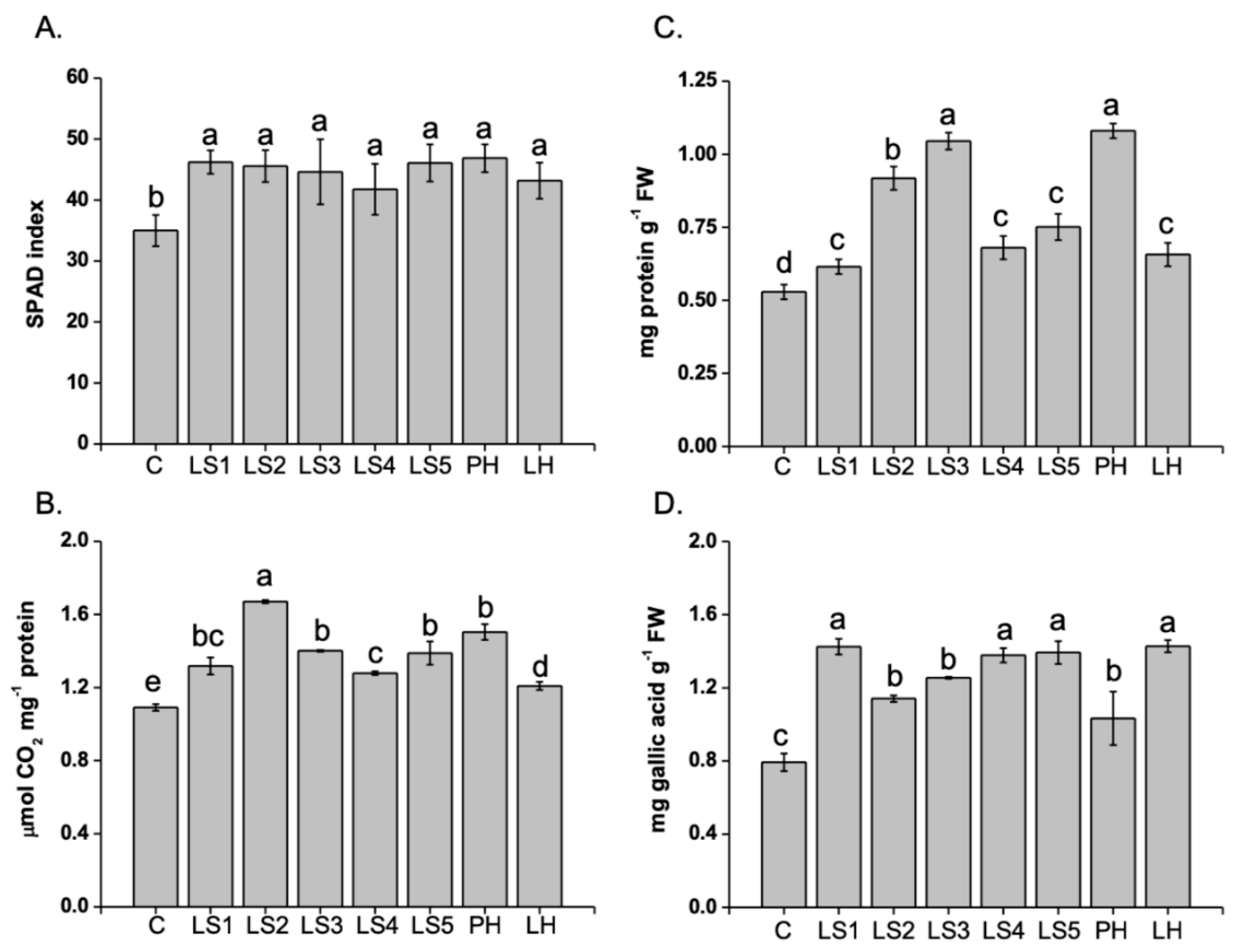
2.3. Effects of Hs on SPAD, RuBisCO activity, and N-compounds (Proteins and Phenolics)
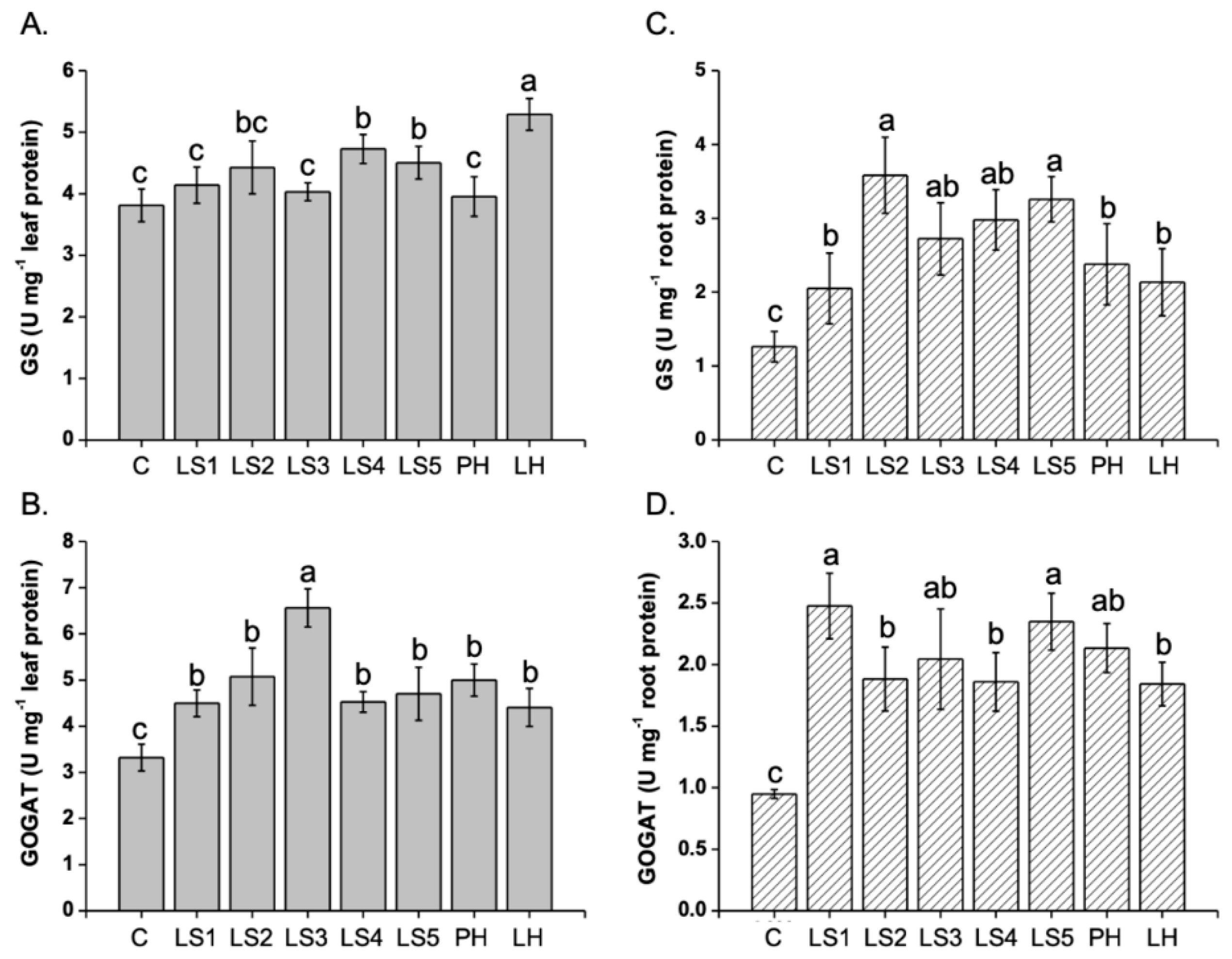
2.4. Effects of Hs on GS and GOGAT Activities
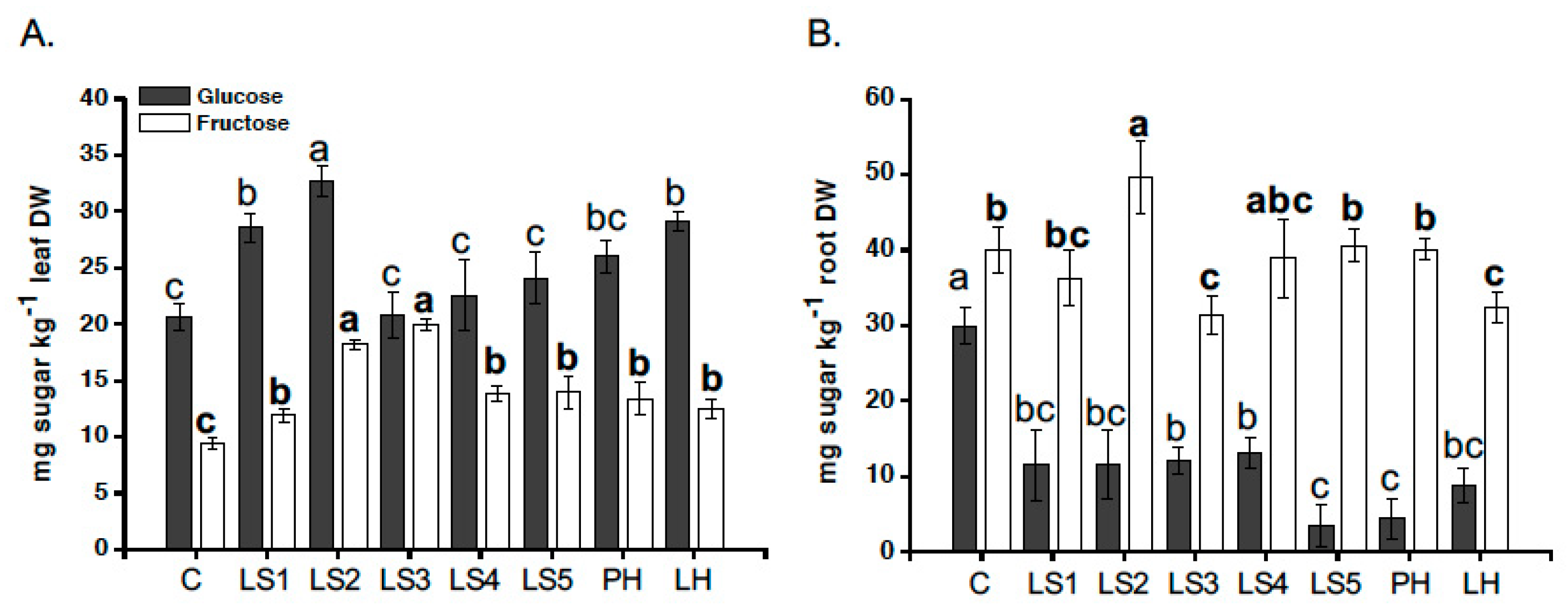
2.5. Effects of Hs on Reducing Sugar Accumulation
2.6. Statistical Analysis of Data
3. Discussion
4. Materials and Methods
4.1. Elemental Composition and Spectroscopic Analysis of Hs
4.2. Plant Material and Experimental Design
4.3. Determination of the SPAD Index
4.4. Analysis of Soluble Proteins and Reducing Sugars
4.5. Analysis of Total and Individual Phenolic Compounds
4.6. Determination of GS, GOGAT and RuBisCO Activity
4.7. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Karamesouti, M.; Detsis, V.; Kounalaki, A.; Vasiliou, P.; Salvati, L.; Kosmas, C. Land-use and land degradation processes affecting soil resources: Evidence from a traditional Mediterranean cropland. Catena 2015, 132, 45–55. [Google Scholar] [CrossRef]
- Shaviv, A.; Mikkelsen, R.L. Controlled-release fertilizers to increase efficiency of nutrient use and minimize environmental degradation-A review. Fertil. Res. 1993, 35, 1–12. [Google Scholar] [CrossRef]
- Trivedi, P.; Delgado-Baquerizo, M.; Anderson, I.C.; Singh, B.K. Response of soil properties and microbial communities to agriculture: Implications for primary productivity and soil health indicators. Front. Plant Sci. 2016, 7, 990. [Google Scholar] [CrossRef] [PubMed]
- Canellas, L.P.; Olivares, F.L.; Aguiar, N.O.; Jones, D.; Nebbioso, A.; Mazzei, P.; Piccolo, A. Humic and fulvic acids as biostimulants in horticulture. Sci. Hortic. 2015, 196, 15–27. [Google Scholar] [CrossRef]
- Yakhin, O.I.; Lubyanov, A.A.; Yakhin, I.A.; Brown, P.H. Biostimulants in Plant Science: A Global Perspective. Front. Plant Sci. 2017, 7, 2049. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Colla, G.; Nardi, S.; Cardarelli, M.; Ertani, A.; Lucini, L.; Canaguier, R.; Rouphael, Y. Protein hydrolysates as biostimulants in horticulture. Sci. Hortic. 2015, 196, 28–38. [Google Scholar] [CrossRef]
- Nardi, S.; Pizzeghello, D.; Schiavon, M.; Ertani, A. Plant biostimulants: Physiological responses induced by protein hydrolyzed-based products and humic substances in plant metabolism. Sci. Agric. 2016, 73, 18–23. [Google Scholar] [CrossRef]
- Du Jardin, P. Plant biostimulants: Definition, concept, main categories and regulation. Sci. Hortic. 2015, 196, 3–14. [Google Scholar] [CrossRef] [Green Version]
- Calvo, P.; Nelson, L.; Kloepper, J.W. Agricultural uses of plant biostimulants. Plant Soil 2014, 383, 3–41. [Google Scholar] [CrossRef] [Green Version]
- Nardi, S.; Carletti, P.; Pizzeghello, D.; Muscolo, A. Biological activities of humic substances. In Biophysico-Chemical Processes Involving Natural Nonliving Organic Matter in Environmental Systems; Part, I. Fundamentals and Impact of Mineral-Organic-Biota Interactions on the Formation, Transformation, Turnover, and Storage of Natural Nonliving Organic Matter (NOM); Senesi, N., Xing, B., Huang, P.M., Eds.; John Wiley & Sons: Hoboken, NJ, USA, 2009; Volume 2, pp. 305–339. [Google Scholar]
- Nardi, S.; Ertani, A.; Francioso, O. Soil–root cross-talking: The role of humic substances. J. Plant Nutr. Soil Sci. 2017, 180, 5–13. [Google Scholar] [CrossRef]
- Zandonadi, D.B.; Santos, M.P.; Dobbss, L.B.; Olivares, F.L.; Canellas, L.P.; Binzel, M.L.; Okorokova, A.L.; Façanha, A.R. Nitric oxide mediates humic acids-induced root development and plasma membrane H+-ATPase activation. Planta 2010, 231, 1025. [Google Scholar] [CrossRef]
- Montoneri, E.; Adani, F.; Genevini, P.L.; Ricca, G.; Cherubini, S.; Spitaleri, C. Polyalkylphenyl-sulphonic acids with acid groups of variable strength from animal-vegetable wastes. Waste Manag. 2004, 24, 513–522. [Google Scholar] [CrossRef]
- Gargulak, J.D.; Lebo, S.E. Commercial use of lignin-based materials. In Lignin: Historical, Biological and Haterials Perspectives; Glass, W.G., Northey, R.A., Schultz, T.P., Eds.; ACS Symposium Series 742; American Chemical Society: Washington, DC, USA, 2000; Volume 15, pp. 2–559. [Google Scholar]
- Lutnaes, B.F.; Myrvold, B.O.; Lauten, R.A.; Endeshaw, M.M. 1H and 13C NMR data of benzylsulfonic acids model compounds for lignosulfonate. Magn. Reson. Chem. 2008, 46, 299–305. [Google Scholar] [CrossRef]
- McGovern, J.N. Pulp and Paper: Chemistry and Chemical Technology, 3rd ed.; Casey, J.P., Ed.; Wiley: New York, NY, USA, 1980; Volume 4. [Google Scholar]
- Ertani, A.; Francioso, O.; Tugnoli, V.; Righi, V.; Nardi, S. Effect of commercial lignosulfonate-humate on Zea mays L. metabolism. J. Agric. Food Chem. 2011, 59, 11940–11948. [Google Scholar] [CrossRef]
- Matsushita, Y. Conversion of technical lignins to functional materials with retained polymeric properties. J. Wood Sci. 2015, 61, 230–250. [Google Scholar] [CrossRef] [Green Version]
- Cieschi, M.T.; Lucena, J.J. Iron and Humic Acid Accumulation on Soybean Roots Fertilized with Leonardite Iron Humates under Calcareous Conditions. J. Agric. Food Chem. 2018, 66, 13386–13396. [Google Scholar] [CrossRef]
- Kovács, K.; Czech, V.; Fodor, F.; Solti, A.; Lucena, J.J.; Santos-Rosell, S.; Hernández-Apaolaza, L. Characterization of Fe-Leonardite complexes as novel natural iron fertilizers. J. Agric. Food Chem. 2013, 61, 12200–12210. [Google Scholar] [CrossRef]
- Aguirre, E.; Leménager, D.; Bacaicoa, E.; Fuentes, M.; Baigiorri, R.; Zamarreño, A.M.; García-Mina, J.M. The root application of a purified leonardite humic acid modifies the transcriptional regulation of the main physiological root responses to Fe deficiency in Fe-sufficient cucumber plants. Plant Physiol. Biochem. 2009, 47, 215–223. [Google Scholar] [CrossRef]
- Kulikova, N.A.; Stepanova, E.V.; Koroleva, O.V. Mitigating activity of humic substances: Direct influence on biota. In Use of Humic Substances to Remediate Polluted Environments: From Theory to Practice; Springer: Dordrecht, The Netherlands, 2005; pp. 285–309. [Google Scholar]
- Billingham, K. Humic products potential or presumption for agriculture? Can humic products improve my soil? In Proceedings of the 27th Annual Conference of the Grassland Society of NSW Inc., Orange, NSW, Australia, 24–26 July 2012; Harris, C., Lodge, G., Waters, C., Eds.; Grassland Society of NSW Inc.: Wagga, Australia, 2012; pp. 43–50. [Google Scholar]
- Mikkelsen, R.L. Humic materials for agriculture. Better Crop 2005, 89, 6–10. [Google Scholar]
- Lobartini, J.C.; Tan, K.H.; Rema, J.A.; Gingle, A.R.; Pape, C.; Himmelsbach, D.S. The geochemical nature and agricultural importance of commercial humic matter. Sci. Total Environ. 1992, 113, 1–15. [Google Scholar] [CrossRef]
- Gaur, A.C.; Bhardwaj, K.K.R. Influence of sodium humate on the crop plants inoculated with bacteria of agricultural importance. Plant Soil 1971, 35, 613–621. [Google Scholar] [CrossRef]
- Mora, V.; Bacaicoa, E.; Baigorri, R.; Zamarreno, A.M.; Garcia-Mina, J.M. NO and IAA key regulators in the shoot growth promoting action of humic acid in Cucumis sativus L. J. Plant Growth Regul. 2014, 33, 430–439. [Google Scholar] [CrossRef]
- Iakimenko, O.S. Commercial humates from coal and their influence on soil properties and initial plant development. In Use of Humic Substances to Remediate Polluted Environments: From Theory to Practice; Springer: Dordrecht, The Netherlands, 2005; pp. 365–378. [Google Scholar]
- Schulz, H.; Baranska, M. Identification and quantification of valuable plant substances by IR and Raman spectroscopy. Vib. Spectrosc. 2007, 43, 13–25. [Google Scholar] [CrossRef]
- Suganuma, S.; Nakajima, K.; Kitano, M.; Yamaguchi, D.; Kato, H.; Hayashiet, S. Hydrolysis of cellulose by amorphous carbon bearing SO3H, COOH, and OH groups. J. Am. Chem. Soc. 2008, 130, 12787–12793. [Google Scholar] [CrossRef]
- Semwal, S.; Gaur, R.; Mukherjee, S.; Chopra, A.; Gupta, R.P.; Kumar, R.; Tuli, D.K. Structural features of dilute acid pretreated Acacia mangium and impact of sodium sulfite supplementation on enzymatic hydrolysis. Sustain. Chem. Eng. 2016, 4, 4635–4644. [Google Scholar] [CrossRef]
- Nadji, H.; Diouf, P.N.; Benaboura, A.; Bedard, Y.; Riedl, B.; Stevanovic, T. Comparative study of lignins isolated from Alfa grass (Stipa tenacissima L.). Bioresour. Technol. 2009, 100, 3585–3592. [Google Scholar] [CrossRef]
- Hsu, T.C.; Guo, G.L.; Chen, W.H.; Hwang, W.S. Effect of dilute acid pretreatment of rice straw on structural properties and enzymatic hydrolysis. Bioresour. Technol. 2010, 101, 4907–4913. [Google Scholar] [CrossRef]
- Hergert, H.L. Infrared spectra, In Lignins: Occurrence, Formation, Structure and Reactions; Sarkanen, K.V., Ludwig, C.H., Eds.; John Wiley & Sons, Inc.: New York, NY, USA, 1971; pp. 267–296. [Google Scholar]
- Ozgenc, O.; Durmaz, S.; Hakki Boyaci, I.; Eksi-Kocak, H. Determination of chemical changes in heat-treated wood using ATR-FTIR and FT Raman spectrometry. Spectrochim. Acta Part A Mol. Biomol. Spectrosc. 2017, 171, 395–400. [Google Scholar] [CrossRef]
- Séné, C.; McCann, M.C.; Wilson, R.H.; Crinter, R. Fourier-Transform Raman and Fourier-Transform lnfrared Spectroscopy. Plant Physiol. 1994, 106, 1623–1631. [Google Scholar]
- Synytsya, A.; Bleha, R.; Synytsya, A.; Pohl, R.; Hayashi, K.; Yoshinaga, K.; Nakano, T.; Hayashi, T. Mekabu fucoidan: Structural complexity and defensive effects against avian influenza A viruses. Carbohydr. Polym. 2014, 111, 633–644. [Google Scholar] [CrossRef]
- Marinval, N.; Saboural, P.; Haddad, O.; Maire, M.; Bassand, K.; Geinguenaud, F.; Djaker, N.; Ben Akrout, K.; Lamy de la Chapelle, M.; Robert, R.; et al. Identification of a pro-angiogenic potential and cellular uptake mechanism of a LMW highly sulfated fraction of fucoidan from Ascophyllum nodosum. Mar. Drugs 2016, 14, 185. [Google Scholar] [CrossRef]
- Mora, V.; Baigorri, R.; Bacaicoa, E.; Zamarrenõ, A.M.; García-Mina, J.M. The humic acid induced changes in the root concentration of nitric oxide, IAA and ethylene do not explain the changes in root architecture caused by humic acid in cucumber. Environ. Exp. Bot. 2012, 76, 24–32. [Google Scholar] [CrossRef]
- Mora, V.; Bacaicoa, E.; Zamarrenõ, A.M.; Aguirre, E.; Garnica, M.; Fuentes, M.; García-Mina, J.M. Action of humic acid on promotion of cucumber shoot growth involves nitrate-related changes associated with the root-to-shoot distribution of cytokinins, polyamines and mineral nutrients. J. Plant Physiol. 2010, 167, 633–642. [Google Scholar] [CrossRef]
- Pizzeghello, D.; Nicolini, G.; Nardi, S. Hormone-like activity of humic substances in Fagus sylvaticae L. forests. New Phytol. 2001, 151, 647–657. [Google Scholar] [CrossRef]
- Muscolo, A.; Sidari, M.; Francioso, O.; Tugnoli, V.; Nardi, S. The auxin-like activity of humic substances is related to membrane interactions in carrot cell cultures. J. Chem. Ecol. 2007, 33, 115–129. [Google Scholar] [CrossRef]
- Pizzeghello, D.; Zanella, A.; Carletti, P.; Nardi, S. Chemical and biochemical characterization of dissolved organic matter from Silver fir and beech forest soils. Chemosphere 2006, 65, 190–200. [Google Scholar] [CrossRef]
- Muscolo, A.; Sidari, M.; Attinà, E.; Francioso, O.; Tugnoli, V.; Nardi, S. Biological activity of humic substances is related to their chemical structure. Soil Sci. Soc. Am. J. 2007, 71, 75–85. [Google Scholar] [CrossRef]
- Jannin, L.; Arkoun, M.; Ourry, A.; Laîné, P.; Goux, D.; Garnica, M.; Fuentes, M.; San Francisco, S.; Baigorri, R.; Cruz, F.; et al. Microarray analysis of humic acid effects on Brassica napus growth: Involvement of N, C and S metabolisms. Plant Soil 2012, 359, 297–319. [Google Scholar] [CrossRef]
- Vance, C.P.; Gantt, J.S. Control of nitrogen and carbon metabolism in root nodules. Physiol. Plant. 1992, 85, 266–274. [Google Scholar] [CrossRef]
- Forde, B.G. Nitrate transporters in plants: Structure, function and regulation. BBA-Biomembr. 2000, 1465, 219–235. [Google Scholar] [CrossRef]
- Wang, M.; Shen, Q.; Xu, G.; Guo, S. New insight into the strategy for nitrogen metabolism in plant cells. Int. Rev. Cell Mol. Biol. 2014, 310, 1–37. [Google Scholar]
- Ertani, A.; Schiavon, M.; Nardi, S. Transcriptome-wide identification of differentially expressed genes in Solanum lycopersicon L. in response to an alfalfa-protein hydrolysate using microarrays. Front. Plant Sci. 2017, 8, 1159. [Google Scholar] [CrossRef]
- Lattanzio, V.; Lattanzio, V.M.T.; Cardinali, A. Role of phenolics in the resistance mechanisms of plants against fungal pathogens and insects. In Phytochemistry Advances in Research; Imperato, F., Ed.; Research Signpost: Kerala, India, 2006; pp. 23–67. [Google Scholar]
- Vermerris, W.; Nicholson, R. Phenolic compound biochemistry. In The Role of Phenols in Plant Defense; Springer: Dordrecht, The Netherlands, 2006; pp. 211–234. [Google Scholar]
- Chen, F.; Tholl, D.; Bohlmann, J.; Pichersky, E. The family of terpene synthases in plants: A mid-size family of genes for specialized metabolism that is highly diversified throughout the kingdom. Plant J. 2011, 66, 212–229. [Google Scholar] [CrossRef]
- Zhang, F.P.; Yang, Q.Y.; Zhang, S.B. Dual effect of phenolic nectar on three floral visitors of Elsholtzia rugulosa (Lamiaceae) in SW China. PLoS ONE 2016, 11, e0154381. [Google Scholar] [CrossRef]
- Di Marco, G.; D’Ambrosio, N.; Giardi, M.T.; Massacci, A.; Tricoli, D. Photosynthetic properties of leaves of a yellow green mutant of wheat compared to its wild type. Photosynth. Res. 1989, 21, 117–122. [Google Scholar]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Product | C | N | S |
|---|---|---|---|
| % (w/w) | |||
| LS1 | 54.56 ± 1.02 | 2.18 ± 0.13 | 5.56 ± 0.34 |
| LS2 | 41.08 ± 1.10 | 1.70 ± 0.15 | 6.12 ± 0.40 |
| LS3 | 41.28 ± 1.32 | 1.95 ± 0.21 | 5.49 ± 0.12 |
| LS4 | 37.11 ± 1.15 | 1.58 ± 0.17 | 7.83 ± 0.21 |
| LS5 | 48.15 ± 1.50 | 2.12 ± 0.13 | 5.13 ± 0.34 |
| PH | 38.05 ± 1.01 | 1.67 ± 0.20 | 1.30 ± 0.23 |
| LH | 33.04 ± 1.14 | 1.64 ± 0.22 | 5.33 ± 0.31 |
| Attributions | LS2 | LS5 |
|---|---|---|
| OH stretching | 3490 w | 3490 vw |
| OH stretching | 3250 w | 3250 w |
| Aromatic CH stretching | 3070 w | 3070 vw |
| Aliphatic CH stretching | 2940 m | 2940 w |
| Aliphatic CH stretching | 2846 m–w | 2846 m–w |
| 1670 conjugated C=O | 1670 w. sh | 1670 w. sh |
| Phenolic peak | 1633 s. sh | |
| Aryl ring stretch, symmetric (lignin); Phenolic peak | 1604 vs | 1604 s |
| Car-H in plane bend, CO(H) str. | About 1500 vw. sh | About 1500 w. sh |
| CH3 bending in OCH3 (lignin and carbohydrates) | 1460m–w | 1460 vw |
| C-H bend in R3C-H (cellulose) | 1370 m | 1370 m |
| Aliphatic O-H bend (cellulose) | 1330 m | |
| Sulfate group, asymmetric stretching | 1284 m | 1284 m |
| Phenol (lignin) | 1190 m–w | 1190 m–w |
| C-C skeletal mode OCH3 loop rocking | 1157 w. sh | 1157 w. sh |
| Sulfate group, symmetric stretching | 1082 w. sh | 1082 m–w. sh |
| OC(H3) stretching and rocking | 1045 m | 1045 m |
| H-C-C and H-C-O bending at C6 (cellulose) | 898 w | 898 m |
| bending of primary C6-O-S | 815 m | 815 m |
| Chlorogenic | Caffeic | p-Cumaric | Ferulic | p-Hydroxybenzoic | ||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Leaves (mg k−1 FW) | ||||||||||
| C | 30.29 | ± 4.11e | 0.52 | ± 0.02d | 1.28 | ± 0.05d | 0.58 | ± 0.02c | 16.32 | ± 0.13e |
| LS1 | 67.65 | ± 8.54b | 1.41 | ± 0.02c | 3.02 | ± 0.05a | 3.32 | ± 0.03a | 47.88 | ± 3.45a |
| LS2 | 81.13 | ± 12.37a | 3.90 | ± 0.02a | 1.33 | ± 0.03c | 2.48 | ± 0.06a | 46.88 | ± 5.33a |
| LS3 | 51.24 | ± 6.32c | 1.34 | ± 0.05c | 2.61 | ± 0.06b | 1.87 | ± 0.07b | 30.04 | ± 7.34c |
| LS4 | 85.98 | ± 7.10a | 1.88 | ± 0.05b | 2.41 | ± 0.07b | 4.13 | ± 0.05a | 49.24 | ± 6.13a |
| LS5 | 57.10 | ± 10.22c | 1.47 | ± 0.01c | 2.35 | ± 0.05b | 1.24 | ± 0.05b | 39.94 | ± 5.28b |
| PH | 34.09 | ± 5.13d | 0.72 | ± 0.02d | 3.09 | ± 0.07a | 1.22 | ± 0.05b | 40.69 | ± 6.13b |
| LH | 37.48 | ± 5.08d | 0.92 | ± 0.03d | 2.03 | ± 0.03c | 0.72 | ± 0.03c | 24.78 | ± 5.13d |
| Roots (mg kg−1 FW) | ||||||||||
| C | 3.54 | ± 0.12c | n.d. | 5.71 | ± 0.30b | 0.58 | ± 0.01c | 29.97 | ± 4.14a | |
| LS1 | 3.56 | ± 0.11c | n.d. | 1.04 | ± 0.62c | 3.09 | ± 0.03a | 3.65 | ± 0.84d | |
| LS2 | 6.05 | ± 0.13a | n.d. | 3.16 | ± 0.61b | 2.86 | ± 0.05a | 11.57 | ± 3.12c | |
| LS3 | 3.44 | ± 0.12c | n.d. | 6.29 | ± 0.82a | 0.85 | ± 0.03c | 21.07 | ± 2.34b | |
| LS4 | 5.67 | ± 0.23b | n.d. | 5.10 | ± 0.72b | 1.12 | ± 0.03b | 19.46 | ± 4.13b | |
| LS5 | 7.61 | ± 0.14a | n.d. | 4.89 | ± 0.53b | 1.69 | ± 0.05b | 24.39 | ± 6.81b | |
| PH | 7.54 | ± 0.17a | n.d. | 6.46 | ± 0.52a | 2.65 | ± 0.05a | 31.12 | ± 5.68a | |
| LH | 3.48 | ± 0.18c | n.d. | 6.15 | ± 0.51a | 0.75 | ± 0.01c | 20.29 | ± 3.12b | |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ertani, A.; Nardi, S.; Francioso, O.; Pizzeghello, D.; Tinti, A.; Schiavon, M. Metabolite-Targeted Analysis and Physiological Traits of Zea mays L. in Response to Application of a Leonardite-Humate and Lignosulfonate-Based Products for Their Evaluation as Potential Biostimulants. Agronomy 2019, 9, 445. https://doi.org/10.3390/agronomy9080445
Ertani A, Nardi S, Francioso O, Pizzeghello D, Tinti A, Schiavon M. Metabolite-Targeted Analysis and Physiological Traits of Zea mays L. in Response to Application of a Leonardite-Humate and Lignosulfonate-Based Products for Their Evaluation as Potential Biostimulants. Agronomy. 2019; 9(8):445. https://doi.org/10.3390/agronomy9080445
Chicago/Turabian StyleErtani, Andrea, Serenella Nardi, Ornella Francioso, Diego Pizzeghello, Anna Tinti, and Michela Schiavon. 2019. "Metabolite-Targeted Analysis and Physiological Traits of Zea mays L. in Response to Application of a Leonardite-Humate and Lignosulfonate-Based Products for Their Evaluation as Potential Biostimulants" Agronomy 9, no. 8: 445. https://doi.org/10.3390/agronomy9080445